Functionality of Fatty Acid Chemoreception: A Potential Factor in the Development of Obesity?

{kind=link}
Abstract
:1. Introduction
2. Overweight/Obesity
3. The Sense of Taste and Its Function
4. Possibility of Oral Fatty Acid Chemoreception: But Fat Taste?
4.1. Animal Evidence for Fatty Acid Chemoreception
4.2. Human Evidence for Fatty Acid Chemoreception
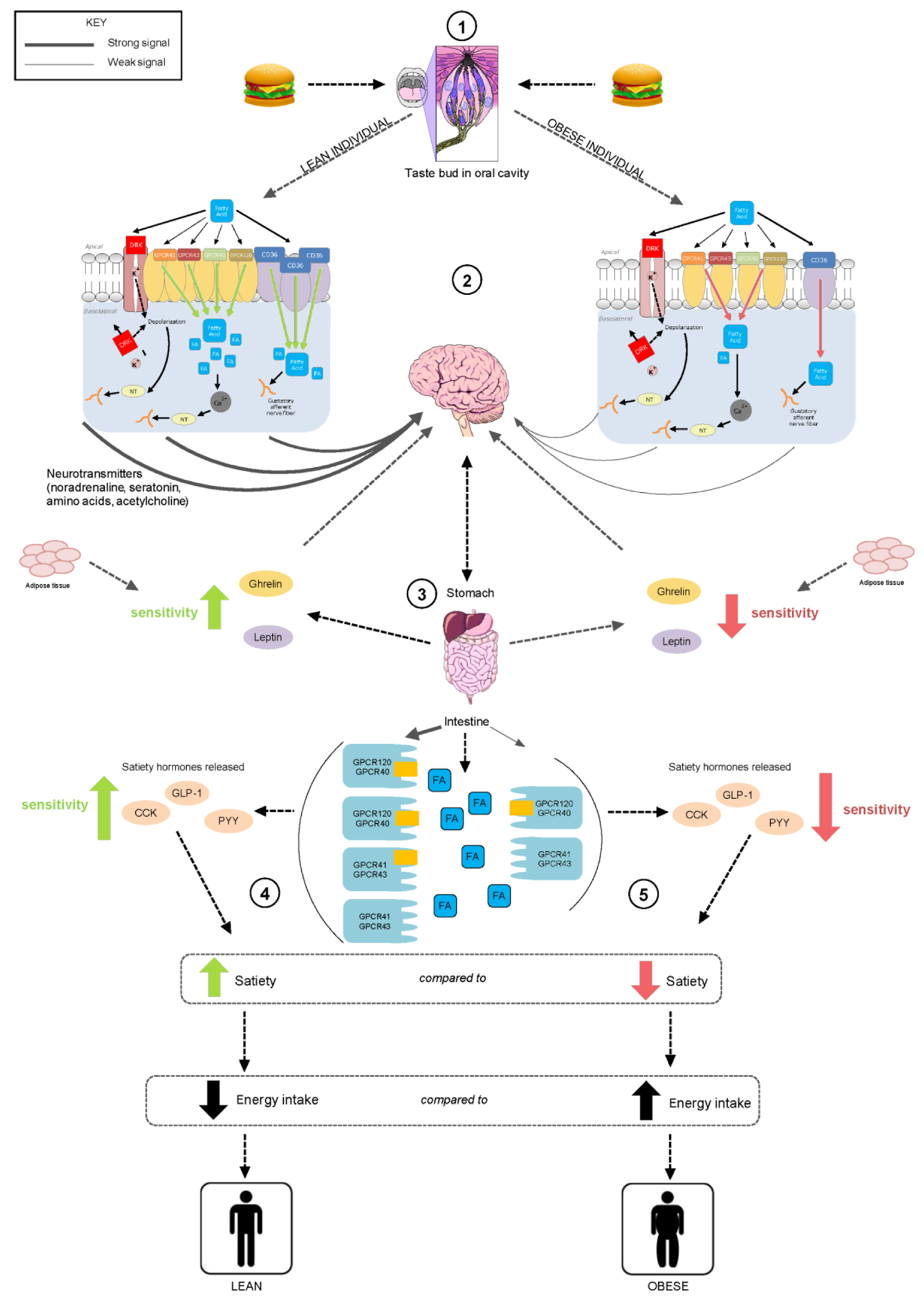
5. Putative Mechanisms for Fatty Acid Chemoreception
5.1. CD36 Transporter and Fatty Acid Transporter (FAT)
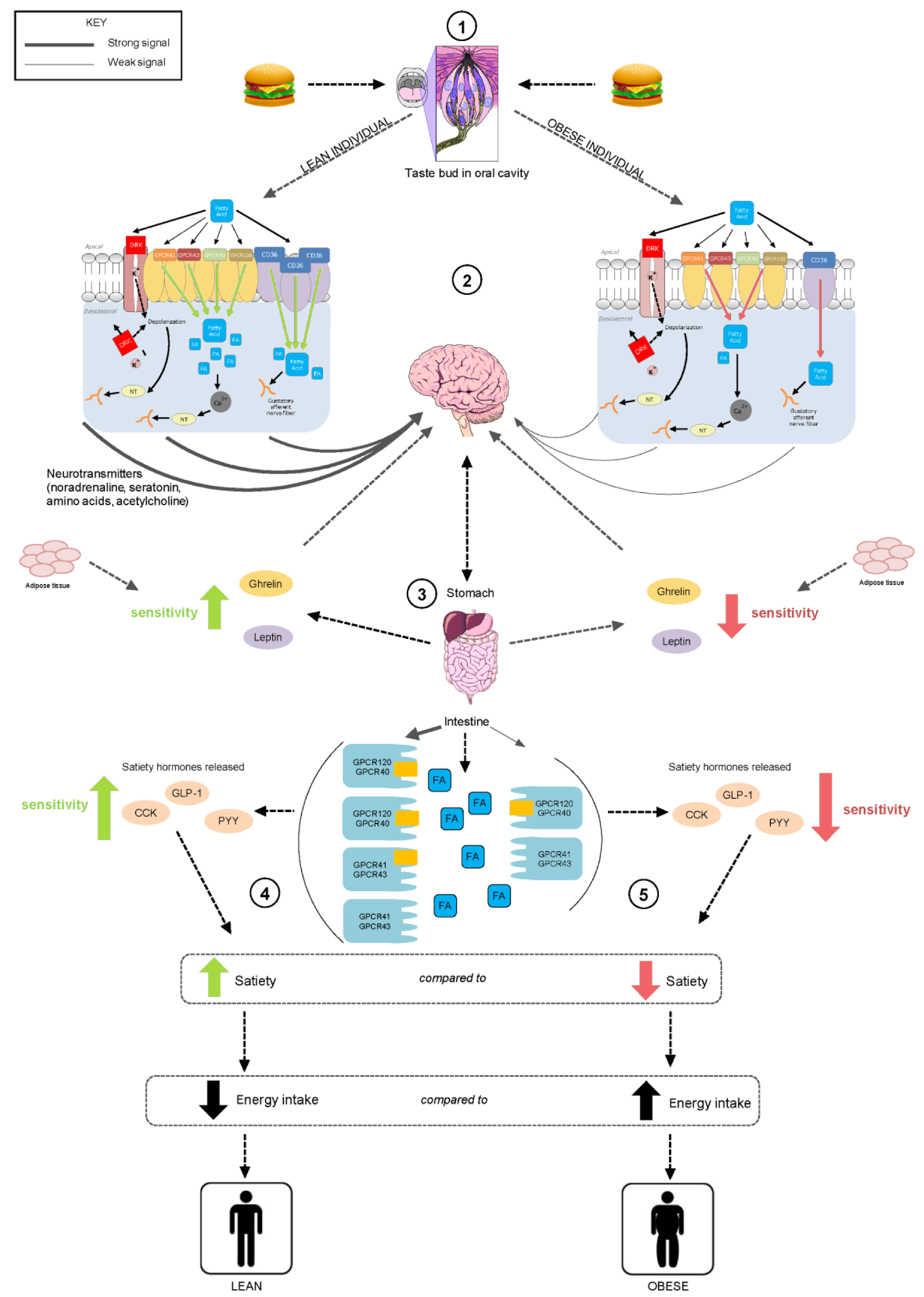
5.2. GPCRs
5.3. DRK Channels
5.4. Lingual Lipase
6. Possible Functions of Fatty Acid Chemoreception
7. Fatty Acid Sensitivity in the Oral Cavity and GI Tract
8. Dietary Influences on Fatty Acid Chemoreception
9. Environmental Influences on Gene Expression
10. Conclusions
Acknowledgments
Conflict of Interest
References
- Swinburn, B.A.; Sacks, G.; Hall, K.D.; McPherson, K.; Finegood, D.T.; Moodie, M.L.; Gortmaker, S.L. The global obesity pandemic: Shaped by global drivers and local environments. Lancet 2011, 378, 804–814. [Google Scholar] [CrossRef]
- Wang, Y.C.; McPherson, K.; Marsh, T.; Gortmaker, S.L.; Brown, M. Health and economic burden of the projected obesity trends in the USA and the UK. Lancet 2011, 378, 815–825. [Google Scholar] [CrossRef]
- Snoek, H.T.M.; Huntjens, L.; van Gemert, L.J.; de Graaf, C.; Weenen, H. Sensory-specific satiety in obese and normal-weight women. Am. J. Clin. Nutr. 2004, 80, 823–831. [Google Scholar]
- Feltrin, K.L.; Little, T.J.; Meyer, J.H.; Horowitz, M.; Smout, A.J.P.M.; Wishart, J.; Pilichiewicz, A.N.; Rades, T.; Chapman, I.M.; Feinle-Bisset, C. Effects of intraduodenal fatty acids on appetite, antropyloroduodenal motility, and plasma CCK and GLP-1 in humans vary with their chain length. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 56, R524–R533. [Google Scholar]
- Cummings, D.E.; Overduin, J. Gastrointestinal regulation of food intake. J. Clin. Invest. 2007, 117, 13–23. [Google Scholar] [CrossRef]
- Blundell, J.E.; Macdiarmid, J.I. Fat as a risk factor for overconsumption: Satiation, satiety, and patterns of eating. J. Am. Diet. Assoc. 1997, 97, S63–S69. [Google Scholar] [CrossRef]
- Rolls, B.J.; Kim-Harris, S.; Fischman, M.W.; Foltin, R.W.; Moran, T.H.; Stoner, S.A. Satiety after preloads with different amounts of fat and carbohydrate: Implications for obesity. Am. J. Clin. Nutr. 1994, 60, 476–487. [Google Scholar]
- Speechly, D.P.; Buffenstein, R. Appetite dysfunction in obese males: Evidence for role of hyperinsulinaemia in passive overconsumption with a high fat diet. Eur. J. Clin. Nutr. 2000, 54, 225–233. [Google Scholar] [CrossRef]
- Westerterp, K.R. Perception, passive overfeeding and energy metabolism. Physiol. Behav. 2006, 89, 62–65. [Google Scholar] [CrossRef]
- Hoad, V.; Somerford, P.; Katzenellenbogen, J. High body mass index overtakes tobacco as the leading independent risk factor contributing to disease burden in Western Australia. Aust. N. Z. J. Public Health 2010, 34, 214–215. [Google Scholar] [CrossRef]
- Rokholm, B.; Baker, J.L.; Sørensen, T.I.A. The levelling off of the obesity epidemic since the year 1999—A review of evidence and perspectives. Obes. Rev. 2010, 11, 835–846. [Google Scholar] [CrossRef]
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and evolution of the Western diet: Health implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar]
- Bray, G.A.; Paeratakul, S.; Popkin, B.M. Dietary fat and obesity: A review of animal, clinical and epidemiological studies. Physiol. Behav. 2004, 83, 549–555. [Google Scholar] [CrossRef]
- Bachmanov, A.A.; Beauchamp, G.K. Taste receptor genes. Annu. Rev. Nutr. 2007, 27, 389–414. [Google Scholar] [CrossRef]
- Gilbertson, T.A.; Damak, S.; Margolskee, R.F. The molecular physiology of taste transduction. Curr. Opin. Neurobiol. 2000, 10, 519–527. [Google Scholar] [CrossRef]
- Keast, R.S.J.; Roper, J. A complex relationship among chemical concentration, detection threshold, and suprathreshold intensity of bitter compounds. Chem. Senses 2007, 32, 245–253. [Google Scholar] [CrossRef]
- Laugerette, F.; Gaillard, D.; Passilly-Degrace, P.; Niot, I.; Besnard, P. Do we taste fat? Biochimie 2007, 89, 265–269. [Google Scholar] [CrossRef]
- Mattes, R.D. Accumulating evidence supports a taste component for free fatty acids in humans. Physiol. Behav. 2011, 104, 624–631. [Google Scholar] [CrossRef]
- Gibson, J.J. The Senses Considered as Perceptual System; Houghton Mifflin: Boston, MA, USA, 1966. [Google Scholar]
- Green, B.G. Studying taste as a cutaneous sense. Food Qual. Prefer. 2003, 14, 99–109. [Google Scholar] [CrossRef]
- Gilbertson, T.A.; Liu, L.; Kim, I.; Burks, C.A.; Hansen, D.R. Fatty acid responses in taste cells from obesity-prone and -resistant rats. Physiol. Behav. 2005, 86, 681–690. [Google Scholar] [CrossRef]
- Takeda, M.; Sawano, S.; Imaizumi, M.; Fushiki, T. Preference for corn oil in olfactory-blocked mice in the conditioned place preference test and the two-bottle choice test. Life Sci. 2001, 69, 847–854. [Google Scholar] [CrossRef]
- Gilbertson, T.A.; Fontenot, D.T. Fatty acid modulation of K+ channels in taste receptor cells: Gustatory cues for dietary fat. Am. J. Physiol. 1997, 272, C1203. [Google Scholar]
- Gilbertson, T.A. Gustatory mechanisms for the detection of fat. Curr. Opin. Neurobiol. 1998, 8, 447–452. [Google Scholar] [CrossRef]
- Abumrad, N.A. CD36 may determine our desire for dietary fats. J. Clin. Invest. 2005, 115, 2965–2967. [Google Scholar] [CrossRef]
- Simons, P.J.; Kummer, J.A.; Luiken, J.J.F.P.; Boon, L. Apical CD36 immunolocalization in human and porcine taste buds from circumvallate and foliate papillae. Acta Histochem. 2010, 113, 839–843. [Google Scholar]
- Kawai, T.; Fushiki, T. Importance of lipolysis in oral cavity for orosensory detection of fat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 54, R447–R454. [Google Scholar]
- Zhang, X.-J.; Zhou, L.-H.; Ban, X.; Liu, D.-X.; Jiang, W.; Liu, X.-M. Decreased expression of CD36 in circumvallate taste buds of high-fat diet induced obese rats. Acta Histochem. 2011, 113, 663–667. [Google Scholar] [CrossRef]
- Naville, D.; Duchampt, A.; Vigier, M.; Oursel, D.; Lessire, R.; Poirier, H.; Niot, I.; Bégeot, M.; Besnard, P.; Mithieux, G. Link between intestinal CD36 ligand binding and satiety induced by a high protein diet in mice. PLoS One 2012, 7, e30686–e30686. [Google Scholar] [CrossRef]
- Pepino, M.Y.; Love-Gregory, L.; Klein, S.; Abumrad, N.A. The fatty acid translocase gene, CD36, and lingual lipase influence oral sensitivity to fat in obese subjects. J. Lipid Res. 2011, 53, 561–566. [Google Scholar]
- Galindo, M.M.; Voigt, N.; Stein, J.; van Lengerich, J.; Raguse, J.-D.; Hofmann, T.; Meyerhof, W.; Behrens, M. G protein-coupled receptors in human fat taste perception. Chem. Senses 2012, 37, 123–139. [Google Scholar] [CrossRef]
- Cartoni, C.; Yasumatsu, K.; Ohkuri, T.; Shigemura, N.; Yoshida, R.; Godinot, N.; le Coutre, J.; Ninomiya, Y.; Damak, S. Taste preference for fatty acids is mediated by GPR40 and GPR120. J. Neurosci. 2010, 30, 8376–8382. [Google Scholar] [CrossRef]
- Gotoh, C.; Hong, Y.-H.; Iga, T.; Hishikawa, D.; Suzuki, Y.; Song, S.-H.; Choi, K.-C.; Adachi, T.; Hirasawa, A.; Tsujimoto, G.; et al. The regulation of adipogenesis through GPR120. Biochem. Biophys. Res. Commun. 2007, 354, 591–597. [Google Scholar] [CrossRef]
- Matsumura, S.; Eguchi, A.; Mizushige, T.; Kitabayashi, N.; Tsuzuki, S.; Inoue, K.; Fushiki, T. Colocalization of GPR120 with phospholipase-Cβ2 and α-gustducin in the taste bud cells in mice. Neurosci. Lett. 2009, 450, 186–190. [Google Scholar]
- Ichimura, A.; Hirasawa, A.; Poulain-Godefroy, O.; Bonnefond, A.; Hara, T.; Yengo, L.; Kimura, I.; Leloire, A.; Liu, N.; Iida, K.; et al. Dysfunction of lipid sensor GPR120 leads to obesity in both mouse and human. Nature 2012, 483, 350–354. [Google Scholar] [CrossRef] [Green Version]
- Laugerette, F.; Passilly-Degrace, P.; Patris, B.; Niot, I.; Febbraio, M.; Montmayeur, J.P.; Besnard, P. CD36 involvement in orosensory detection of dietary lipids, spontaneous fat preference, and digestive secretions. J. Clin. Invest. 2005, 115, 3177–3184. [Google Scholar] [CrossRef]
- Tsuruta, M.; Kawada, T.; Fukuwatari, T.; Fushiki, T. The orosensory recognition of long-chain fatty acids in rats. Physiol. Behav. 1999, 66, 285–288. [Google Scholar] [CrossRef]
- Fukuwatari, T.; Shibata, K.; Iguchi, K.; Saeki, T.; Iwata, A.; Tani, K.; Sugimoto, E.; Fushiki, T. Role of gustation in the recognition of oleate and triolein in anosmic rats. Physiol. Behav. 2003, 78, 579–583. [Google Scholar] [CrossRef]
- Mindell, S.; Smith, G.P.; Greenberg, D. Corn oil and mineral oil stimulate sham feeding in rats. Physiol. Behav. 1990, 48, 283–287. [Google Scholar] [CrossRef]
- Mattes, R.D. Oral detection of short-, medium-, and long-chain free fatty acids in humans. Chem. Senses 2009, 34, 145–150. [Google Scholar] [CrossRef]
- Mattes, R.D. Oral thresholds and suprathreshold intensity ratings for free fatty acids on 3 tongue sites in humans: Implications for transduction mechanisms. Chem. Senses 2009, 34, 415–423. [Google Scholar] [CrossRef]
- Mattes, R.D. Brief oral stimulation, but especially oral fat exposure, elevates serum triglycerides in humans. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 59, G365–G371. [Google Scholar]
- Stewart, J.E.; Feinle-Bisset, C.; Golding, M.; Delahunty, C.; Clifton, P.M.; Keast, R.S.J. Oral sensitivity to fatty acids, food consumption and BMI in human subjects. Br. J. Nutr. 2010, 104, 145–152. [Google Scholar] [CrossRef]
- Stewart, J.E.; Newman, L.P.; Keast, R.S.J. Oral sensitivity to oleic acid is associated with fat intake and body mass index. Clin. Nutr. 2011, 6, 838–844. [Google Scholar] [CrossRef]
- Stewart, J.E.; Seimon, R.V.; Otto, B.R.; Keast, R.S.J.; Clifton, P.M.; Feinle-Bisset, C. Marked differences in gustatory and gastrointestinal sensitivity to oleic acid between lean and obese men. Am. J. Clin. Nutr. 2011, 93, 703–711. [Google Scholar] [CrossRef]
- Mattes, R.D. Oral exposure to butter, but not fat replacers elevates postprandial triacylglycerol concentration in humans. J. Nutr. 2001, 131, 1491–1496. [Google Scholar]
- Mattes, R.D. Fat taste and lipid metabolism in humans. Physiol. Behav. 2005, 86, 691–697. [Google Scholar] [CrossRef]
- Crystal, S.R.; Teff, K.L. Tasting fat: Cephalic phase hormonal responses and food intake in restrained and unrestrained eaters. Physiol. Behav. 2006, 89, 213–220. [Google Scholar] [CrossRef]
- Ebba, S.; Abarintos, R.A.; Kim, D.G.; Tiyouh, M.; Stull, J.C.; Movalia, A.; Smutzer, G. The examination of fatty acid taste with edible strips. Physiol. Behav. 2012, 106, 579–586. [Google Scholar] [CrossRef]
- Ahne, G.; Erras, A.; Hummel, T.; Kobal, G. Assessment of gustatory function by means of tasting tablets. Laryngoscope 2000, 110, 1396–1401. [Google Scholar] [CrossRef]
- Newman, L.P.; Keast, R.S.J. The test retest reliability of fatty acid taste thresholds. Chemosens. Percept. 2013, in press. [Google Scholar]
- Keller, K.L.; Liang, L.C.H.; Sakimura, J.; May, D.; van Belle, C.; Breen, C.; Driggin, E.; Tepper, B.J.; Lanzano, P.C.; Deng, L.; et al. Common variants in the CD36 gene are associated with oral fat perception, fat preferences, and obesity in African Americans. Obesity (Silver Spring) 2012, 20, 1066–1073. [Google Scholar] [CrossRef]
- Liu, P.; Shah, B.P.; Croasdell, S.; Gilbertson, T.A. Transient receptor potential channel Type M5 is essential for fat taste. J. Neurosci. 2011, 31, 8634–8642. [Google Scholar] [CrossRef]
- Matsumura, S.; Mizushige, T.; Yoneda, T.; Iwanaga, T.; Tsuzuki, S.; Inoue, K.; Fushiki, T. GPR expression in the rat taste bud relating to fatty acid sensing. Biomed. Res. 2007, 28, 49–55. [Google Scholar] [CrossRef]
- Covington, D.; Briscoe, C.; Brown, A.; Jayawickreme, C. The G-protein-coupled receptor 40 family (GPR40-GPR43) and its role in nutrient sensing. Biochem. Soc. Trans. 2006, 34, 770–773. [Google Scholar] [CrossRef]
- Laffargue, A.; de Kochko, A.; Dussert, S. Development of solid-phase extraction and methylation procedures to analyse free fatty acids in lipid-rich seeds. Plant. Physiol. Biochem. 2007, 45, 250–257. [Google Scholar] [CrossRef]
- Kulkarni, B.; Mattes, R. Evidence for presence of nonesterified Fatty acids as potential gustatory signaling molecules in humans. Chem. Senses 2013, 38, 119–127. [Google Scholar] [CrossRef]
- Chale-Rush, A.; Burgess, J.R.; Mattes, R.D. Multiple routes of chemosensitivity to free fatty acids in humans. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1206–G1212. [Google Scholar] [CrossRef]
- Drayna, D. Human taste genetics. Annu. Rev. Genomics Hum. Genet. 2005, 6, 217–235. [Google Scholar] [CrossRef]
- Drewnowski, A.; Henderson, S.A.; Hann, C.S.; Berg, W.A.; Ruffin, M.T. Genetic taste markers and preferences for vegetables and fruit of female breast care patients. J. Am. Diet. Assoc. 2000, 100, 191–197. [Google Scholar] [CrossRef]
- Keller, K.L.; Steinmann, L.; Nurse, R.J.; Tepper, B.J. Genetic taste sensitivity to 6-n-propylthiouracil influences food preference and reported intake in preschool children. Appetite 2002, 38, 3–12. [Google Scholar] [CrossRef]
- Turnbull, B.; Matisoo-Smith, E. Taste sensitivity to 6-n-propylthiouracil predicts acceptance of bitter-tasting spinach in 3–6-y-old children. Am. J. Clin. Nutr. 2002, 76, 1101–1105. [Google Scholar]
- Kamphuis, M.M.; Westerterp-Plantenga, M.S.; Saris, W.H. Fat-specific satiety in humans for fat high in linoleic acid vs fat high in oleic acid. Eur. J. Clin. Nutr. 2001, 55, 499–508. [Google Scholar] [CrossRef]
- Heini, A.F.; Lara-Castro, C.; Kirk, K.A.; Considine, R.V.; Caro, J.F.; Weinsier, R.L. Association of leptin and hunger-satiety ratings in obese women. Int. J. Obes. Relat. Metab. Disord. 1998, 22, 1084–1087. [Google Scholar]
- Little, T.J.; Feinle-Bisset, C. Oral and gastrointestinal sensing of dietary fat and appetite regulation in humans: Modification by diet and obesity. Front. Neurosci. 2010, 4, 178–178. [Google Scholar]
- Stewart, J.E.; Keast, R.S.J. Recent fat intake modulates fat taste sensitivity in lean and overweight subjects. Int. J. Obes. 2011, 6, 834–842. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Newman, L.; Haryono, R.; Keast, R. Functionality of Fatty Acid Chemoreception: A Potential Factor in the Development of Obesity? Nutrients 2013, 5, 1287-1300. https://doi.org/10.3390/nu5041287
Newman L, Haryono R, Keast R. Functionality of Fatty Acid Chemoreception: A Potential Factor in the Development of Obesity? Nutrients. 2013; 5(4):1287-1300. https://doi.org/10.3390/nu5041287
Chicago/Turabian StyleNewman, Lisa, Rivkeh Haryono, and Russell Keast. 2013. "Functionality of Fatty Acid Chemoreception: A Potential Factor in the Development of Obesity?" Nutrients 5, no. 4: 1287-1300. https://doi.org/10.3390/nu5041287