One possible explanation for the absence of an ethanol-paired flavor preference is the unconditionally aversive qualities of 10% ethanol. A problem with this explanation is that HiS and LoS rats conditioned with 10% ethanol in solution express an ethanol-paired flavor preference despite the noxious taste of the conditioning solution [
17,
21]. Potentially, though, a stronger aversive taste-taste association develops in a gelatin medium, preventing expression of a flavor preference. A somewhat more likely explanation centers on differential intake of the 10% ethanol and no-ethanol gelatins during conditioning. Examining caloric compensation in Experiment 1 required providing unlimited access to both gelatins during conditioning. Rats consumed fewer calories as ethanol gelatin than no-ethanol gelatin, and lower caloric intake associated with the former could contribute to preference for the alternative flavor [
34]. In addition, the relative novelty of the ethanol-paired flavor could have reduced selection of it in the choice test (neophobia). Indeed, using solutions during conditioning, preference for an ethanol-paired flavor was less reliable when it was relatively novel [
21]. In that study, only LoS rats displayed sensitivity to relative novelty. However, in Experiment 1, the ethanol/no-ethanol gelatin difference was greater among HiS, so it is at least possible that the larger difference in relative novelty contributed to their tendency to choose the no-ethanol flavor in the test. In Experiments 2–8, a yoking procedure was used to roughly equate each rat’s exposure to the two flavors prior to the preference test, which also ensured that both greater caloric density and total calorie intake would be associated with the ethanol-paired flavor.
Table 1 summarizes Experiment 1 and subsequent manipulation of variables that might influence choice of an ethanol-paired flavor when the conditioning medium is gelatin. Baseline measurement of water intake and bodyweight, the 10-day alternating flavor conditioning procedure, days off, and two-jar flavor preference tests were as described for Experiment 1. In Experiments 2–8, more than one flavor preference test was administered, with one day of free access to chow and water separating the tests; when two gelatin tests were given with different conditions (e.g.,
ad lib. feeding
versus chow deprivation), test order was counterbalanced. Given the uniformity of basic conditioning and test procedures, only key differences from preceding experiments are explained here. In the 10 conditioning experiments conducted, three HiS rats (in different experiments) and two LoS rats (in different experiments) ate virtually none of the ethanol gelatin and were excluded from data analyses.
3.1. Experiment 2
Experiment 2 directly replicated Experiment 1 with two exceptions. First, after the two-gelatin flavor test and a day off, rats received a two-fluid flavor test (0.25% cherry and grape KoolAid in tap water). Second, during conditioning, no-ethanol gelatin intake was yoked to ethanol gelatin intake. Female HiS and LoS rats (
ns = 15) weighing approximately 305 g (no line difference) were given a full jar of ethanol gelatin on Day 1; on Day 2, each rat was given a ration of no-ethanol gelatin equal to the amount of ethanol gelatin (in grams) she had consumed the day before. Yoking was repeated for each of the remaining four ethanol/no-ethanol pairs of conditioning days. Because the caloric density of the ethanol gelatin was higher than that of the no-ethanol gelatin, yoking meant that unlike Experiment 1, rats consumed more calories as ethanol gelatin than no-ethanol gelatin. Greater caloric density and total calorie intake should favor a preference for an associated flavor [
8,
13,
34,
35]. If, in Experiment 1, either having consumed more calories as no-ethanol gelatin or the greater familiarity of the no-ethanol gelatin flavor contributed to indifference or aversion to the ethanol-paired flavor, yoking should increase ethanol-paired flavor preference—at minimum, it should eliminate rejection of it in favor of the no-ethanol flavor.
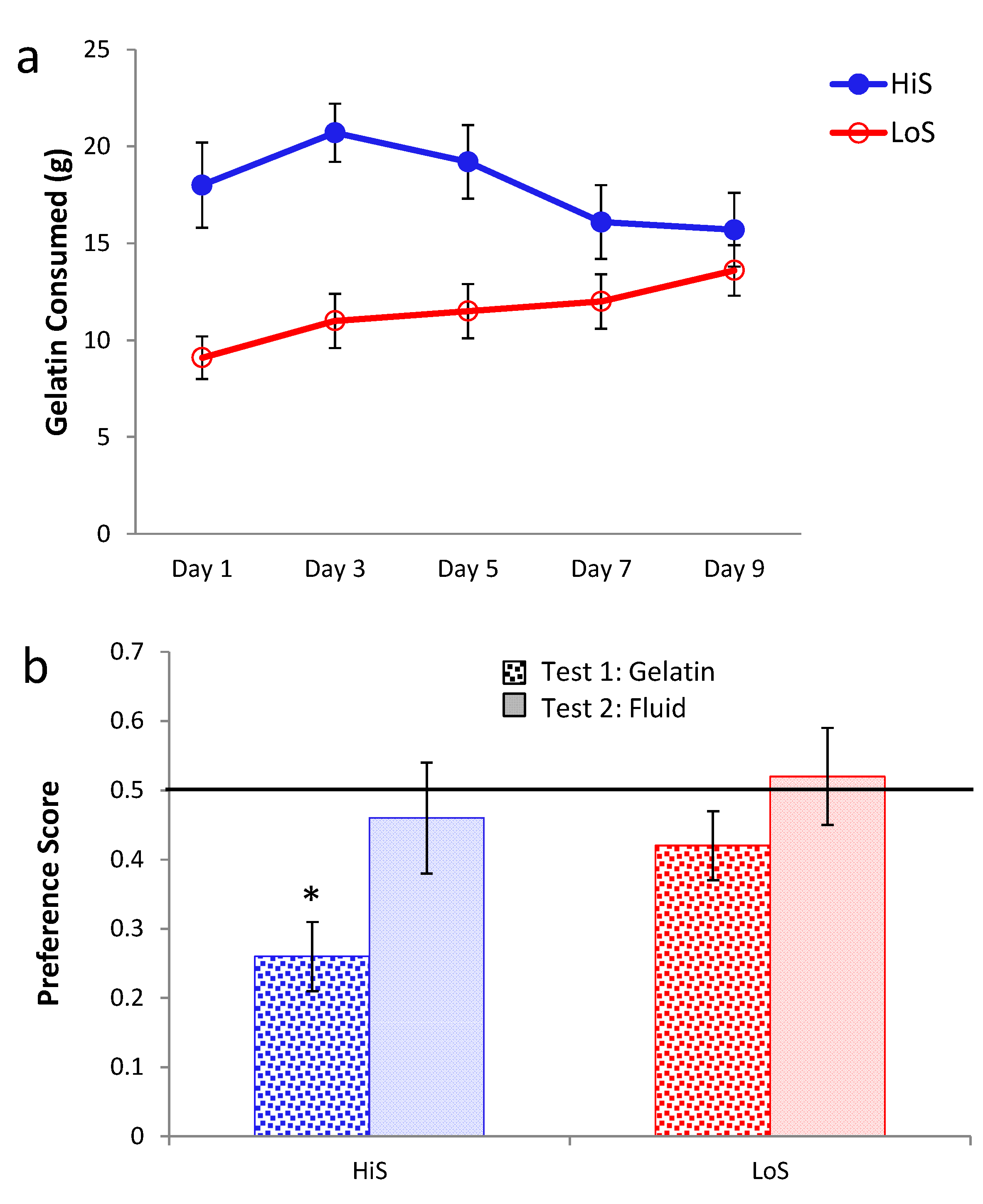
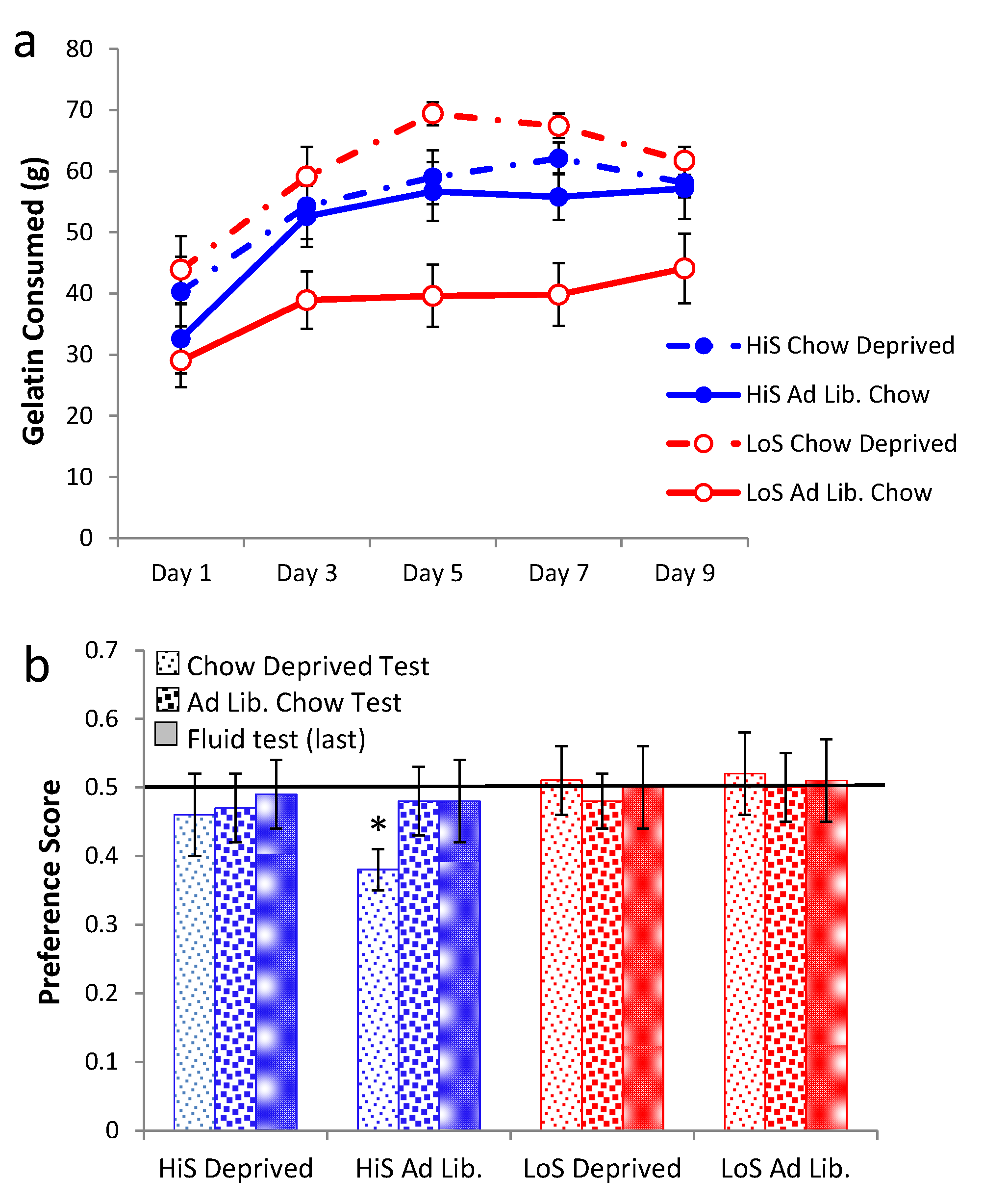
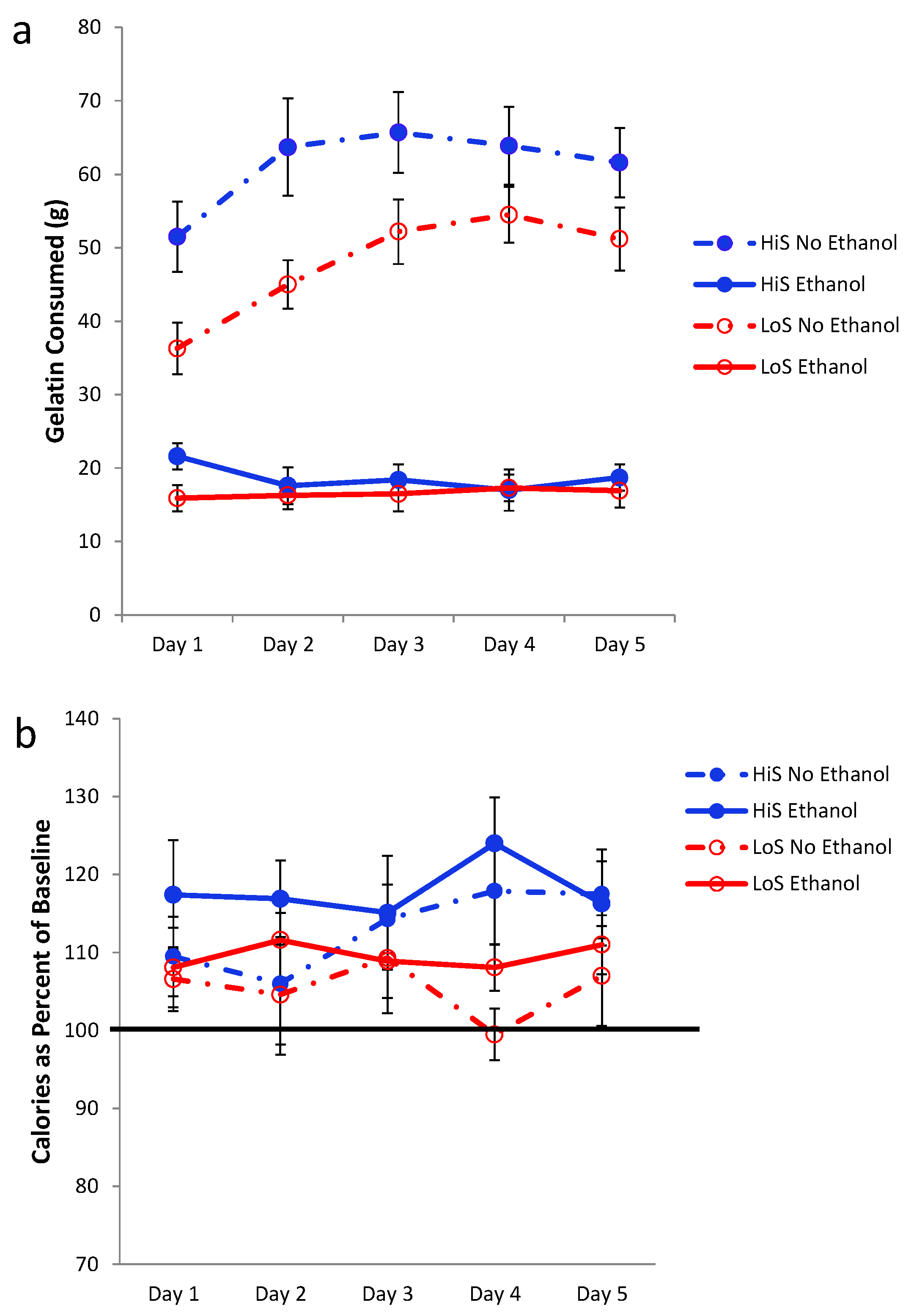
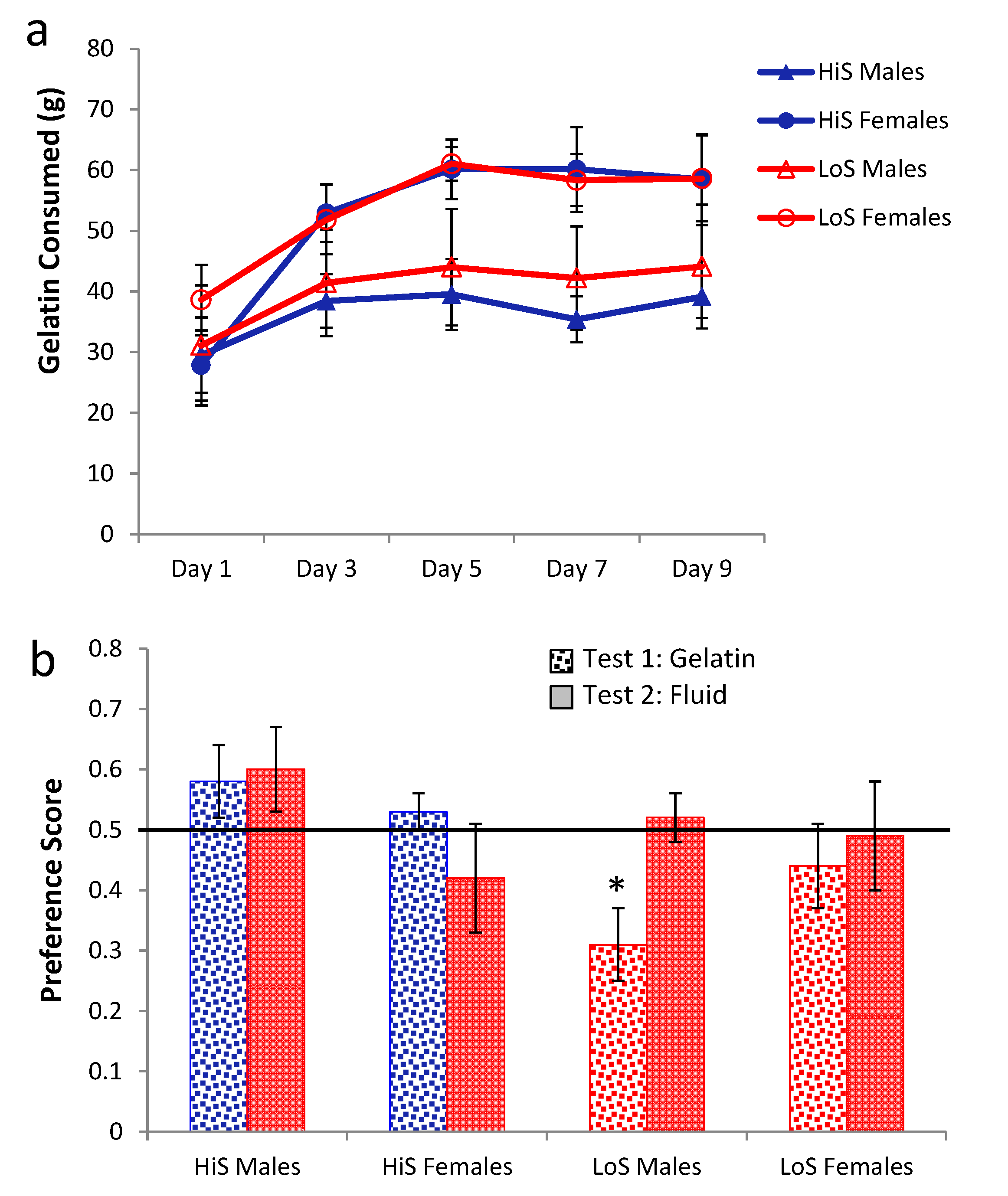
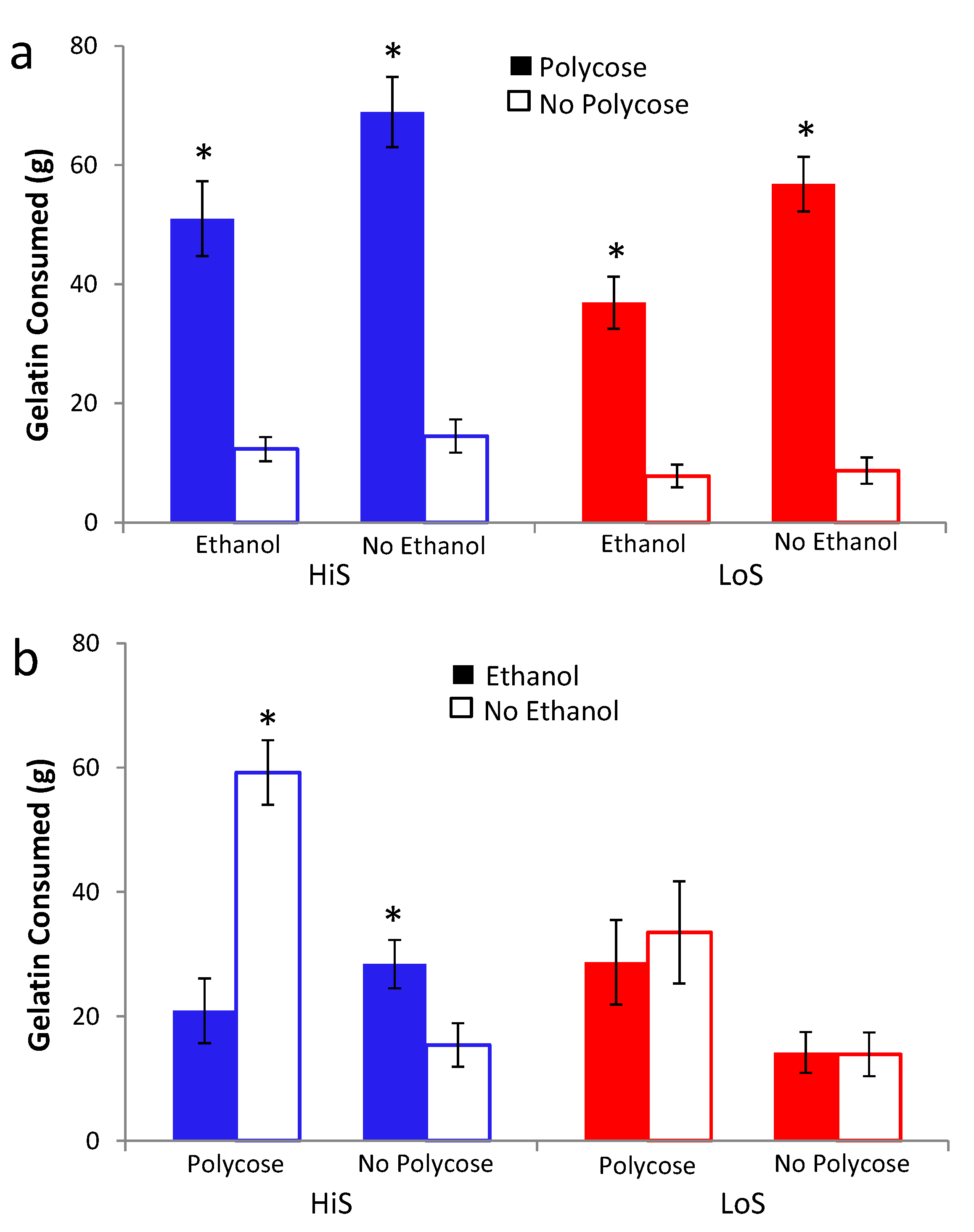
Ethanol gelatin intake during conditioning is shown in
Figure 2a. HiS rats consumed more gelatin than did LoS rats [line main effect,
F(1,22) = 11.32]. However, the line × day interaction also was significant,
F(4,88) = 5.55: Bonferroni-adjusted contrasts showed that LoS rats’ intake did not change significantly across Days 1–5, whereas for HiS rats, intake on Day 2 was significantly higher than on Days 4 and 5. Ethanol-paired flavor preference is shown in
Figure 2b. HiS rats rejected the ethanol-paired flavor in favor of the no-ethanol paired flavor; in the subsequent two-solution test, they were indifferent, a result to which extinction during the first test might have contributed. LoS rats were indifferent in both tests. A line × test medium ANOVA yielded a main effect of test medium,
F(1,22) = 10.00. One-sample
t tests comparing each of the four means to 0.5 showed indifference except for significant aversion among HiS rats tested with gelatin,
t(14) = 4.99.
Table 1.
Summary of Flavor Conditioning and Unconditioned Flavor Test series. Procedural changes within the first series are
bolded. The gelatin type given to rats as a ration (yoked to
ad lib. intake of the alternative on the preceding day) is shown in
![Nutrients 05 04685 i001]()
. Preferences/increases are shown in
![Nutrients 05 04685 i002]()
, aversions/decreases in
![Nutrients 05 04685 i003]()
.
Figure 2.
Intake of 10% ethanol/polycose gelatin during conditioning (Panel a) and ethanol-paired flavor preference in polycose gelatin and no-polycose fluid (Panel b) in Experiment 2. Mean preference scores differing significantly from indifference (0.5) are marked with an asterisk (*).
Figure 2.
Intake of 10% ethanol/polycose gelatin during conditioning (Panel a) and ethanol-paired flavor preference in polycose gelatin and no-polycose fluid (Panel b) in Experiment 2. Mean preference scores differing significantly from indifference (0.5) are marked with an asterisk (*).
Thus, with flavor exposure during conditioning matched and total flavor-associated calorie intake reversed from Experiment 1, HiS rats’ tendency toward aversion to the ethanol-paired flavor—which also signaled higher caloric density—was strengthened, not weakened. This aversion is surprising, even if eating the 10% ethanol gelatin shifts brain ethanol levels toward those resulting from drinking 4% solution [
31], because 4% and 10% ethanol
solutions both support a preference in both lines [
21].
3.2. Experiment 3
Experiment 3 directly replicated Experiment 2 with two exceptions. First, male as well as female rats were used (
ns = 13–14). Second, test gelatins were made without polycose and test fluids were made with polycose. The polycose gelatin used in Experiments 1 and 2 is a palatable, caloric flavor medium. In contrast, the flavored test solutions used in our previous studies [
17,
21] were neither palatable nor caloric. Perhaps, then, rats only express a preference for an ethanol-paired flavor when the test medium otherwise has little positive incentive value. If so, rats might express an ethanol-paired flavor preference when tested with minimally caloric no-polycose gelatins and an aversion when tested with polycose fluids.
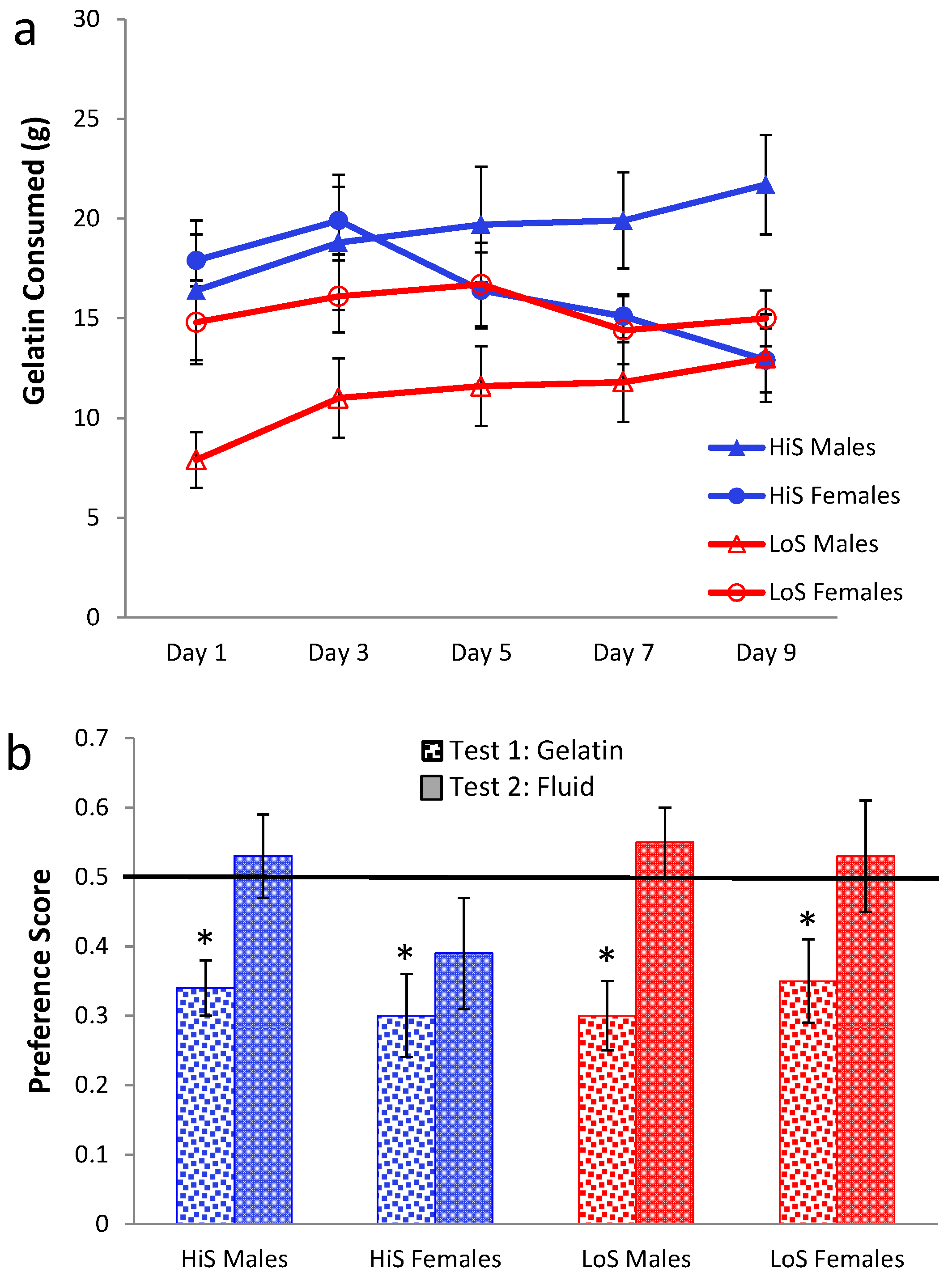
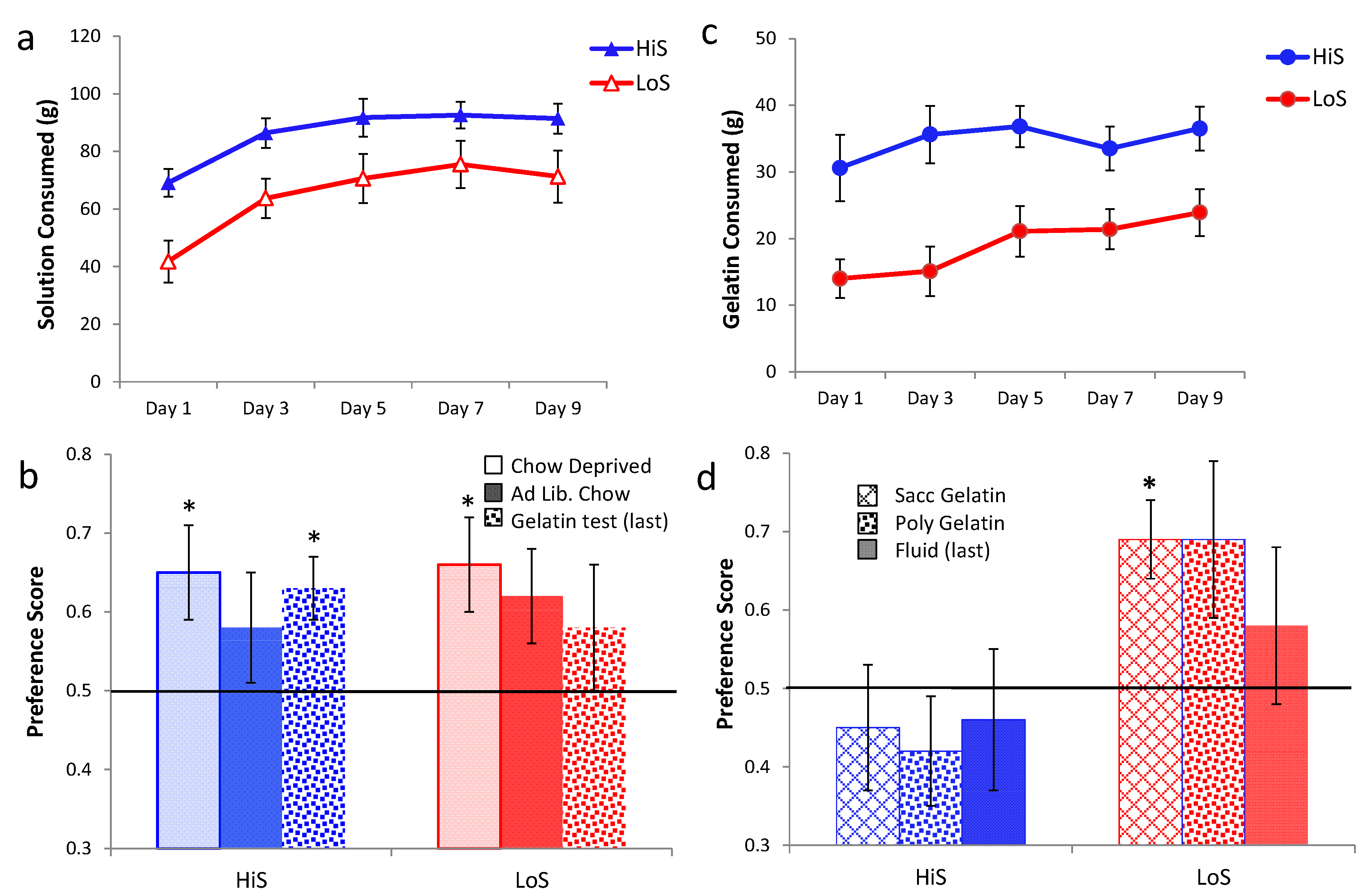
Ethanol gelatin intake during the yoked conditioning procedure is shown in
Figure 3a. As in Experiment 2, female rats’ gelatin intake decreased, somewhat more so among HiS. In contrast, males’ intake increased, with HiS males eating more than LoS males throughout. A line × sex × day ANOVA yielded a line main effect,
F(1,50) = 6.79, and interactions of sex with line,
F(1,50) = 4.01, and day,
F(4,200) = 5.77. The interactions were interpreted with Bonferroni-adjusted contrasts. HiS males’ overall gelatin intake exceeded LoS males’; marginal means for females did not differ. Females’ intake decreased from Day 2 to Day 5; although the interaction with line was not significant, this decrease clearly derives more from HiS than from LoS females, whose intake was relatively stable. Males’ intake increased significantly from Day 1 to Day 5. In contrast, increasing intake of ethanol gelatin among males is consistent with the development of a preference for the ethanol-paired flavor. Yet, as shown in
Figure 3b, males and females in both lines rejected the ethanol-paired flavor in favor of the no-ethanol flavor in the no-polycose gelatin choice test. A line × sex × test medium ANOVA yielded a main effect of test medium,
F(1,50) = 22.13. Comparison of each mean to 0.5 confirmed aversion when all four groups were tested with gelatin,
ts(14) ≥ 2.69, and indifference when tested with solution. Testing with no-polycose gelatin produced more uniform aversion to the ethanol-paired flavor rather than “unmasking” a preference for it, and testing with polycose fluids still yielded indifference.
Figure 3.
Intake of 10% ethanol/polycose gelatin during conditioning (Panel a) and ethanol-paired flavor preference in no-polycose gelatin and polycose fluid (Panel b) in Experiment 3.
Figure 3.
Intake of 10% ethanol/polycose gelatin during conditioning (Panel a) and ethanol-paired flavor preference in no-polycose gelatin and polycose fluid (Panel b) in Experiment 3.
3.3. Experiment 4
Experiment 4 directly replicated Experiment 3 with three exceptions. First, gelatin was made with 4% ethanol (0.9 kcal/g). Unlike 10% ethanol solution, 4% ethanol solution is unconditionally preferred to water by both HiS and LoS rats [
16]; like 10% ethanol, it supports expression of ethanol-paired flavor preference among freely chow fed rats [
21]. Second, during conditioning, yoking was reversed such that no-ethanol gelatin was provided on odd days and a matched ration of 4% ethanol gelatin was provided on even days. This change was made because rats’ preference for 4% ethanol over water leads reasonably to the expectation that they would consume more 4% gelatin than no-ethanol gelatin. Finally, we returned to testing with polycose gelatins and no-polycose fluids. If a relatively unpalatable conditioning and/or test gelatin limits expression of an ethanol-paired flavor preference, then using more palatable media might reveal a preference. We know that rats will express an ethanol-paired flavor preference in a no-polycose fluid test [
17,
21] and so returned to that type of fluid test as well.
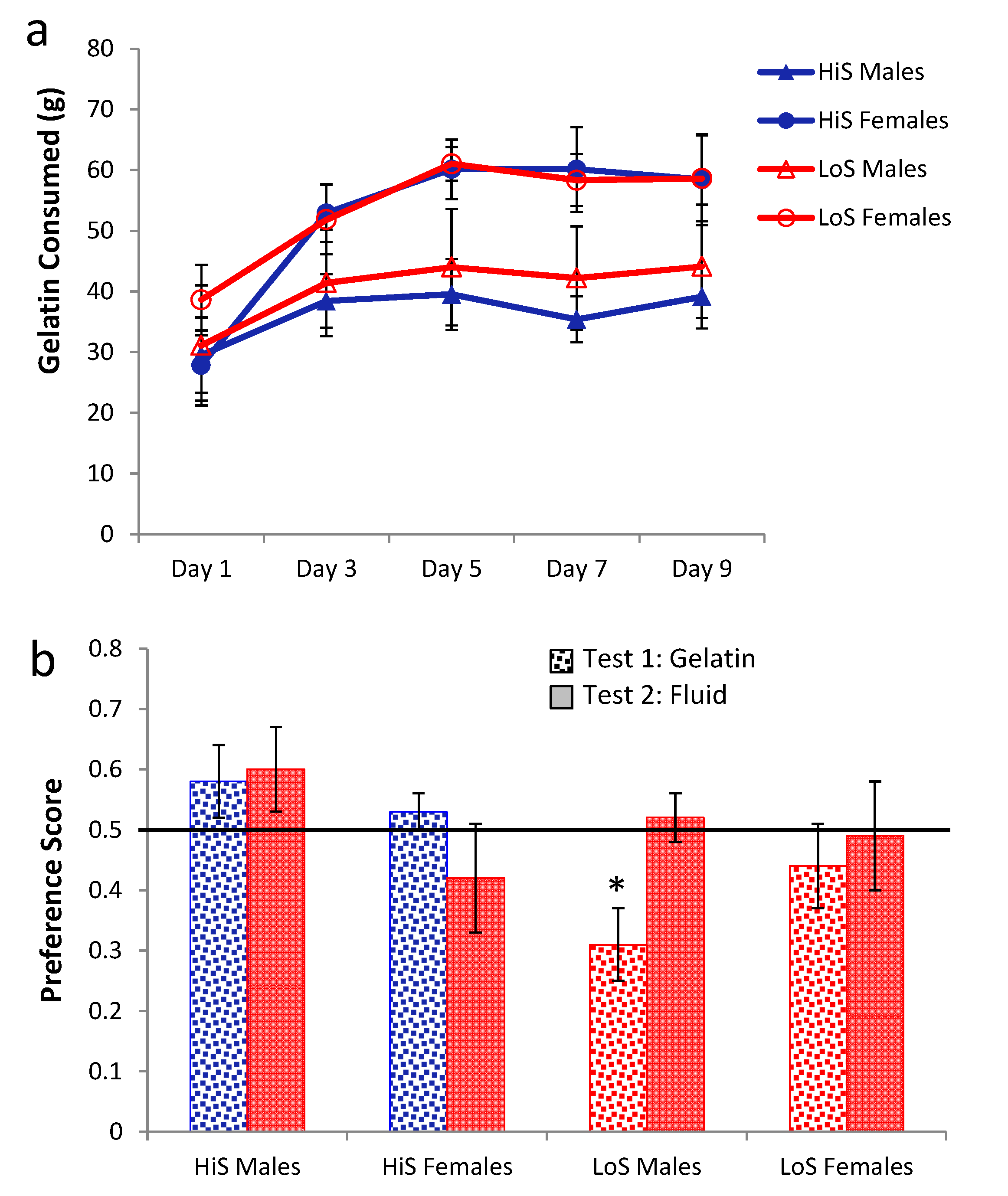
No-ethanol gelatin intake during conditioning by HiS and LoS females and males (
ns = 7 or 8) is shown in
Figure 4a. Overall intake was 2–3 times greater than for 10% ethanol gelatin in Experiments 1–3 and was greater among females than males. Intake increased across days in both males and females, somewhat more so in the latter. Intake was comparable in HiS and LoS rats.
Figure 4.
Intake of 4% ethanol/polycose gelatin during conditioning (Panel a) and ethanol-paired flavor preference in polycose gelatin and no-polycose fluid (Panel b) in Experiment 4.
Figure 4.
Intake of 4% ethanol/polycose gelatin during conditioning (Panel a) and ethanol-paired flavor preference in polycose gelatin and no-polycose fluid (Panel b) in Experiment 4.
A line × sex × day ANOVA yielded main effects of sex,
F(1,27) = 5.82, and day,
F(4,108) = 19.90, and a sex × day interaction,
F(4,108) = 3.90. The interaction was ordinal; Bonferroni-adjusted contrasts showed that in both males and females, intake was significantly lower on Day 1 than on Day 2, which was not significantly different from subsequent days. Expressed as ethanol dose, by the end of conditioning females were consuming a slightly higher dose than they had of 10% ethanol in Experiment 1 (7.6 ± 0.6 g/kg) and more than males (3.6 ± 0.5). This sex difference and the male’s ethanol dose are comparable to results in our original ethanol-paired flavor conditioning study [
21].
Ethanol-paired flavor preference is shown in
Figure 4b. Results for LoS males were the same as when 10% ethanol gelatin was used during conditioning: They were averse to the ethanol-paired flavor in gelatin and were indifferent to it in the subsequent fluid test. All other groups were indifferent in both tests. A line × sex × test medium ANOVA yielded a line × test medium interaction,
F(1,27) = 5.66, and Bonferroni-adjusted contrasts showed that the lines differed in the gelatin test (HiS > LoS) but not in the fluid test. Comparison of each mean to 0.5 confirmed significant aversion when LoS males were tested with gelatin,
t(7) = 3.07, and indifference in all other cases. An unforeseen occurrence during conditioning, however, leaves these test results open to interpretation: Whereas in Experiments 2 and 3 nearly all rats consumed all of their no-ethanol ration on even-numbered days, in Experiment 4 most rats left most of the 4% ethanol rations uneaten. Contrary to the high acceptability of 4% ethanol solution, 4% ethanol did not seem to increase the acceptability of polycose gelatin. The Unconditioned Flavor Test Series (Experiments 8–10) below addresses unconditioned flavor acceptability and preference issues further. Here, the point is that despite the attempt at yoking, the ethanol-paired flavor was somewhat less familiar than the no-ethanol flavor, which would tend to reduce preference for it, especially among LoS rats [
21]. Thus, neophobia could account for LoS males’ rejection of the ethanol-paired flavor, and failure of other groups to prefer it.
3.4. Experiment 5
Experiments 5a and 5b directly replicated Experiment 4 with the following exceptions. First, only females were used (
ns = 15–19). Second, yoking was reversed (ethanol gelatin on odd days, no-ethanol rations on even days). Third, rats were chow deprived during gelatin access (free access to chow otherwise). In our studies with flavored ethanol solutions, those solutions were the only source of fluid available overnight and, because intake was lower than baseline water intake, chow intake likely was lower than baseline. In contrast, in Experiments 1–4, gelatin was not the only source of calories; chow and water were available, and total caloric intake was at or above baseline levels (Experiment 1). If consuming an ethanol-paired flavor during mild caloric restriction contributed to a preference for it in our prior work [
8,
10,
12,
35], then removing chow during gelatin intake might yield a preference. Fourth, the design was run with both 4% ethanol (Experiment 5a) and 10% ethanol (Experiment 5b). Finally, flavored gelatin preference tests were conducted twice, during free access to chow and chow deprivation, with order balanced, prior to the fluid test. Negative energy balance can enhance expression of preferences for an ethanol-paired flavor [
8,
10,
11], so testing during chow deprivation might reveal a preference based on gelatin conditioning that is not expressed when rats are freely feeding.
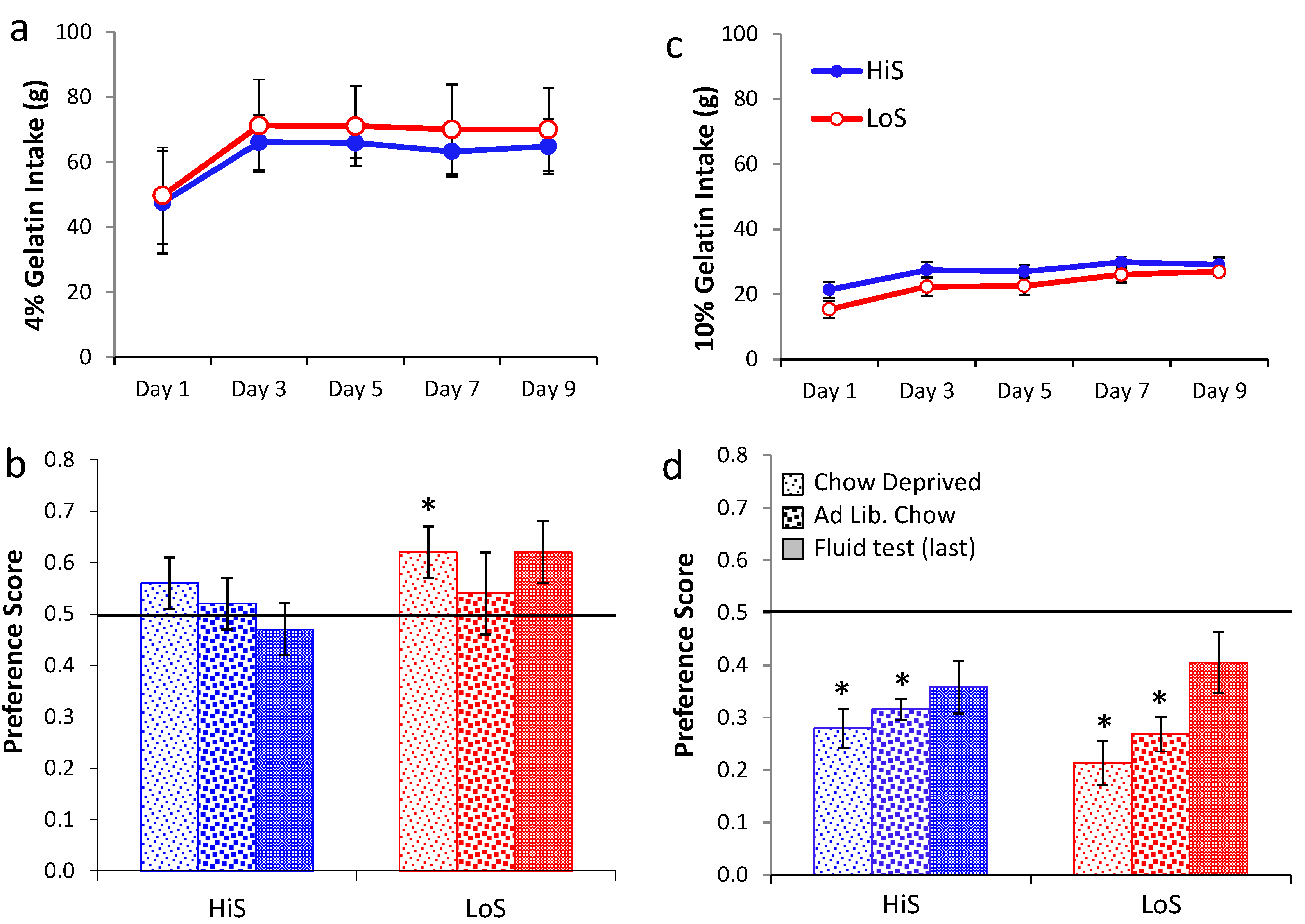
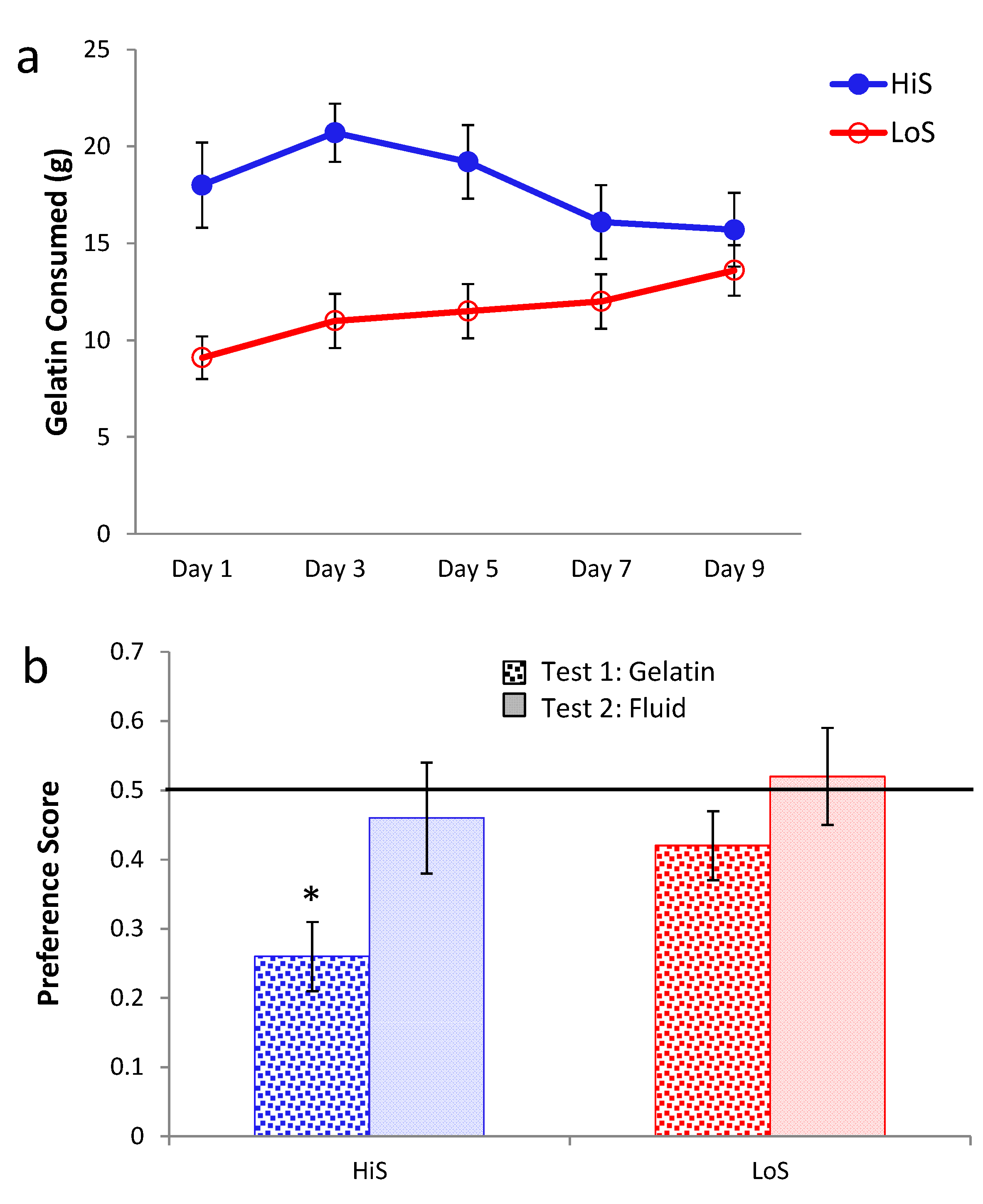
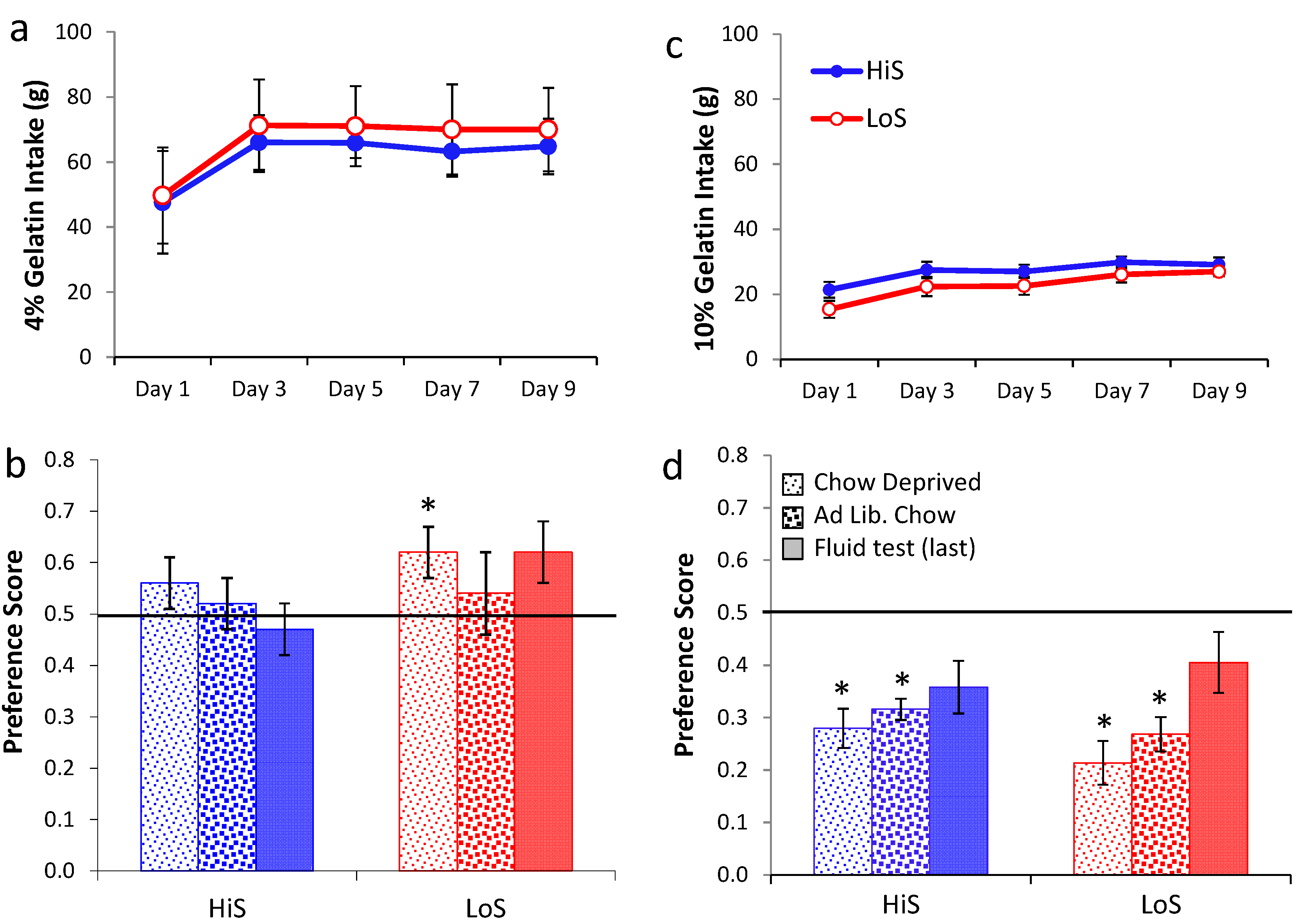
Ethanol gelatin intake during conditioning is shown
Figure 5a (4%) and 5c (10%). At both concentrations, ethanol gelatin increased over days. Line × day ANOVAs yielded a significant main effect of day at both 4%,
F(4,128) = 29.35, and 10%,
F(4,132) = 10.70. No other effects were significant. Preference for a flavor paired with 4% ethanol is shown in
Figure 5b. The first significant ethanol-paired flavor preference in this study was observed here, when LoS rats were conditioned and tested chow deprived. Otherwise, indifference prevailed. A line × test condition (gelatin chow deprived, gelatin
ad lib., fluid) ANOVA yielded no significant effects. Comparison of each mean to 0.5 revealed a significant preference for LoS/Chow Deprived,
t(14) = 2.22.
Rats conditioned with 10% ethanol were averse to the ethanol-paired flavor in gelatin regardless of feeding status, with stronger aversion expressed when rats were tested chow deprived. Trends toward aversion also occurred during the fluid test but neither achieved statistical significance (Ps > 0.058). Because some rats in Experiment 5b were inadvertently tested with fluid containing polycose, a line × feeding status ANOVA was run on data from the gelatin tests, and an independent t test was used to compare the subset of LoS and HiS for which valid fluid test data were available. The ANOVA yielded a significant feeding status effect, F(1,33) = 4.54; no other differences were significant. Comparison of means to 0.5 confirmed aversion in both lines during both gelatin tests, ts > 4.
Figure 5.
Ethanol/polycose gelatin intake by chow-deprived rats in Experiments 5a (4% EtOH) and 5b (10% EtOH) (respectively, Panel a and c; legend in latter). Ethanol-paired flavor preference in polycose gelatin as a function of feeding status, and in a final no-polycose fluid test in Experiments 5a and 5b (respectively, Panel b and d; legend in latter).
Figure 5.
Ethanol/polycose gelatin intake by chow-deprived rats in Experiments 5a (4% EtOH) and 5b (10% EtOH) (respectively, Panel a and c; legend in latter). Ethanol-paired flavor preference in polycose gelatin as a function of feeding status, and in a final no-polycose fluid test in Experiments 5a and 5b (respectively, Panel b and d; legend in latter).
Improved yoking (compared to Exp. 4) did not unmask preference for a flavor paired with 4% ethanol among HiS rats. Nor did chow deprivation during gelatin testing unmask preference for a flavor paired with 10% ethanol; in fact, it strengthened choice of the no-ethanol flavor. These results do suggest constraints within which preference for an ethanol-paired gelatin flavor might occur—specifically, when conditioning gelatin contains a low ethanol concentration (4%) and when rats sensitive to feeding status (LoS) are conditioned and tested during caloric deficit.
3.5. Experiment 6
Experiment 6 was designed to determine whether the ethanol-paired flavor preference observed within those constraints in Experiment 5a is properly attributed to conditioning with chow deprivation, by directly comparing rats conditioned with chow deprivation (as in Experiment 5a) to rats conditioned with continuous access to chow (as in Experiment 4). Procedures were the same as in Experiment 5a except for experimental manipulation of feeding status: HiS and LoS female rats (
ns = 14) were assigned to either
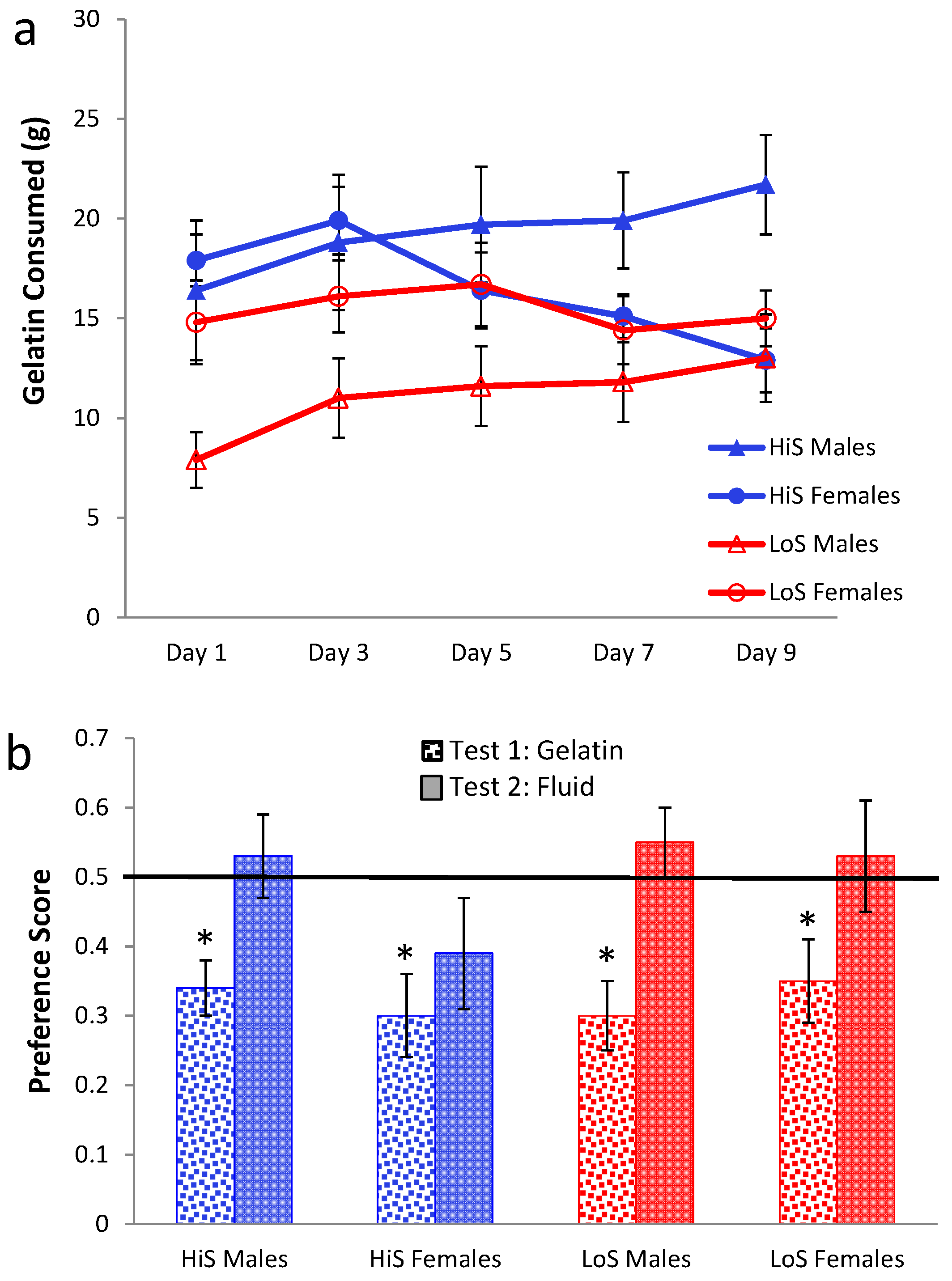
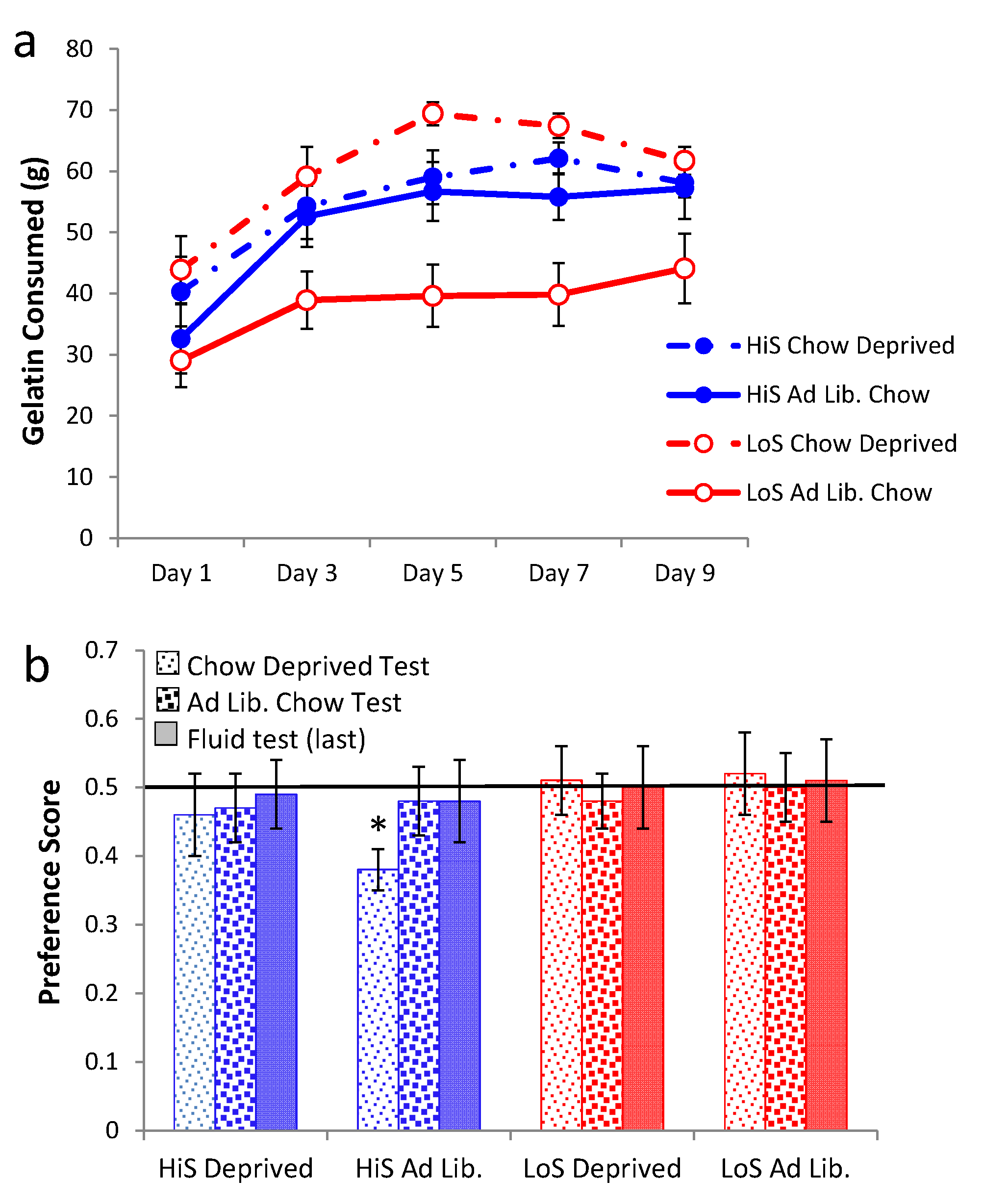
ad lib. chow or chow deprivation during conditioning, with litters balanced across the two conditions. Intake of 4% ethanol gelatin during conditioning is shown in
Figure 6a. Gelatin intake increased over days, and chow deprivation increased gelatin intake among LoS rats. Interestingly, chow deprivation did not affect gelatin intake among HiS rats, another indication of their reduced behavioral regulation of energy balance (Experiment 1) [
21,
22,
23]. A line × feeding status × day ANOVA yielded main effects of feeding status,
F(1,61) = 11.44, and day,
F(4,244) = 39.43, and a line × feeding status interaction,
F(1,61) = 5.72.
Figure 6.
Intake of 4% gelatin during conditioning (Panel a) and ethanol-paired flavor preference in gelatin and fluid (Panel b) in Experiment 6.
Figure 6.
Intake of 4% gelatin during conditioning (Panel a) and ethanol-paired flavor preference in gelatin and fluid (Panel b) in Experiment 6.
Ethanol-paired flavor preference is shown in
Figure 6b. Among rats fed
ad lib. during conditioning, LoS rats were indifferent, and HiS rats expressed frank aversion when chow deprived during gelatin testing. Rats that were chow deprived during conditioning were indifferent in all test conditions; the preference observed among chow-deprived LoS rats in Experiment 5 was not replicated. A line × test condition ANOVA yielded no significant effects. Comparison of each mean to 0.5 yielded a significant difference for HiS/
Ad Lib. Conditioned/Chow Deprived Test,
t(15) = 4.15. No other means differed from 0.5.
3.6. Experiment 7
A viable explanation for failure to observe a reliable ethanol-paired flavor preference after gelatin conditioning is that having 10% polycose in both ethanol and no-ethanol gelatin nullifies whatever differences favor later choice of a flavor associated with ethanol. For instance, flavors can be weaker in gelatin than in solution, at least in humans [
36]. The preceding experiments yielded many deviations from indifference, predominantly in the direction of aversion—especially, surprisingly, among HiS rats. Apparently, having polycose in the gelatins does not simply make their flavors undetectable or indiscriminable. Rather, by increasing caloric density and/or the palatability of the medium, a polycose base might increase the salience of ethanol-associated factors—noxious sensory qualities, for instance—that support aversion and/or attenuate preference. This account is consistent with our having observed ethanol-paired flavor preference among freely feeding HiS and LoS rats when noncaloric, relatively unpalatable solutions were used during conditioning [
17,
21]. The absence of polycose rather than the fluid medium in those studies might account for the discrepancy between those and the present findings.
Experiment 7 examined flavor preference after conditioning with polycose solutions (Experiment 7a) and with no-polycose gelatins (Experiment 7b). Rats in Experiment 3 consumed substantial amounts of no-polycose test gelatins (overall average ~26 g); however, those rats had a history of eating polycose gelatin, and pilot testing showed that many naive rats consume trivial amounts of KoolAid-flavored no-polycose gelatin. Thus, to achieve intake sufficient to support flavor learning, saccharin was added to the no-polycose gelatins in Experiment 7b. We expected HiS rats to consume more gelatin than did LoS rats given selection on a saccharin phenotype, but because HiS rats consumed more polycose gelatin than did LoS rats in the preceding experiments, such a difference would facilitate comparison of these results with earlier ones. Moreover, to date, we have not compared HiS and LoS rats on consumption of a saccharin flavored food that was eaten rather than drunk, so Experiment 7b bore on the generality of their differential propensity to consume saccharin-flavored substances.
In Experiment 7a, the same yoked-intake design was used as in Experiments 2–6, substituting 10% polycose (wt/vol) solutions for polycose gelatins: Male rats (ns = 12) received 4% ethanol/polycose solution flavored with cherry or grape KoolAid (0.25% wt/vol) on each odd day and an equal-weight ration of the alternate flavor in no-ethanol/polycose solution the next day. Two preference tests with flavored no-polycose fluids were conducted, once during chow deprivation and once during ad lib. feeding, with order balanced; a final preference test with polycose gelatin then occurred. Conditioning in Experiment 7b was identical to Experiment 4 except that only females (ns = 11 or 13) were used and gelatins were made with 0.2% (wt/wt) sodium saccharin (Sigma Aldrich Inc., St Louis, MO, USA) instead of polycose. Flavor preference tests were conducted with polycose gelatins and no-polycose saccharin gelatins and then with no-polycose fluids. The polycose and saccharin gelatin tests were balanced for order, and the fluid test was conducted last.
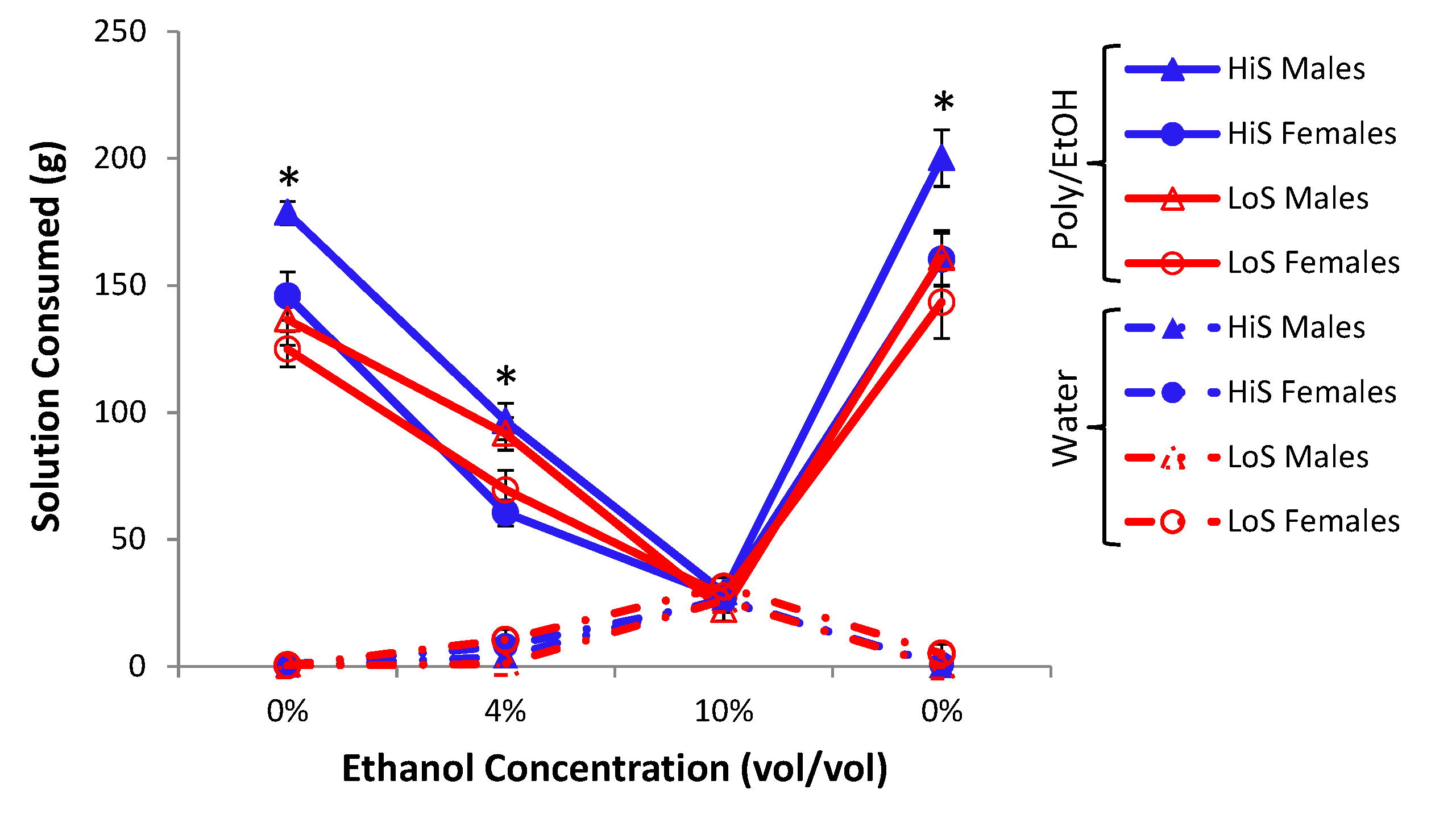
Intake of ethanol/polycose solution during conditioning in Experiment 7a is shown in
Figure 7a. HiS rats drank more than did LoS rats, and intake increased over days. A line × day ANOVA yielded main effects of day and line,
F(1,22) = 5.75, and day,
F(4,88) = 38.52.
Ethanol-paired flavor preference is shown in
Figure 7b. When chow deprived, both HiS and LoS rats expressed a significant preference for the ethanol-paired flavor in solution,
ts(11) 2.47 and 2.40, respectively. The preference also was significant among HiS rats in the gelatin test,
t(11) = 3.45. During
ad lib. testing, preference scores did not differ significantly from 0.5; that preference therefore was less robust than observed previously for both HiS and LoS rats tested during
ad lib. feeding [
19,
21], perhaps indicating some reduction due to use of the polycose vehicle during conditioning or to variability arising from the order-balanced re-testing. At any rate, both lines leaned toward preference, and not aversion, in all three tests when conditioned with polycose solutions. Also of note, HiS rats expressed an ethanol-paired flavor preference in the gelatin test, demonstrating that expression of an ethanol-flavor association is not always limited to the conditioning medium.
Figure 7.
Intake during conditioning (polycose/ethanol solution in Experiment 7a and saccharin/ethanol gelatin in Experiment 7b, respectively, Panels a and c). Ethanol-paired flavor preference is shown in Panels b and d.
Figure 7.
Intake during conditioning (polycose/ethanol solution in Experiment 7a and saccharin/ethanol gelatin in Experiment 7b, respectively, Panels a and c). Ethanol-paired flavor preference is shown in Panels b and d.
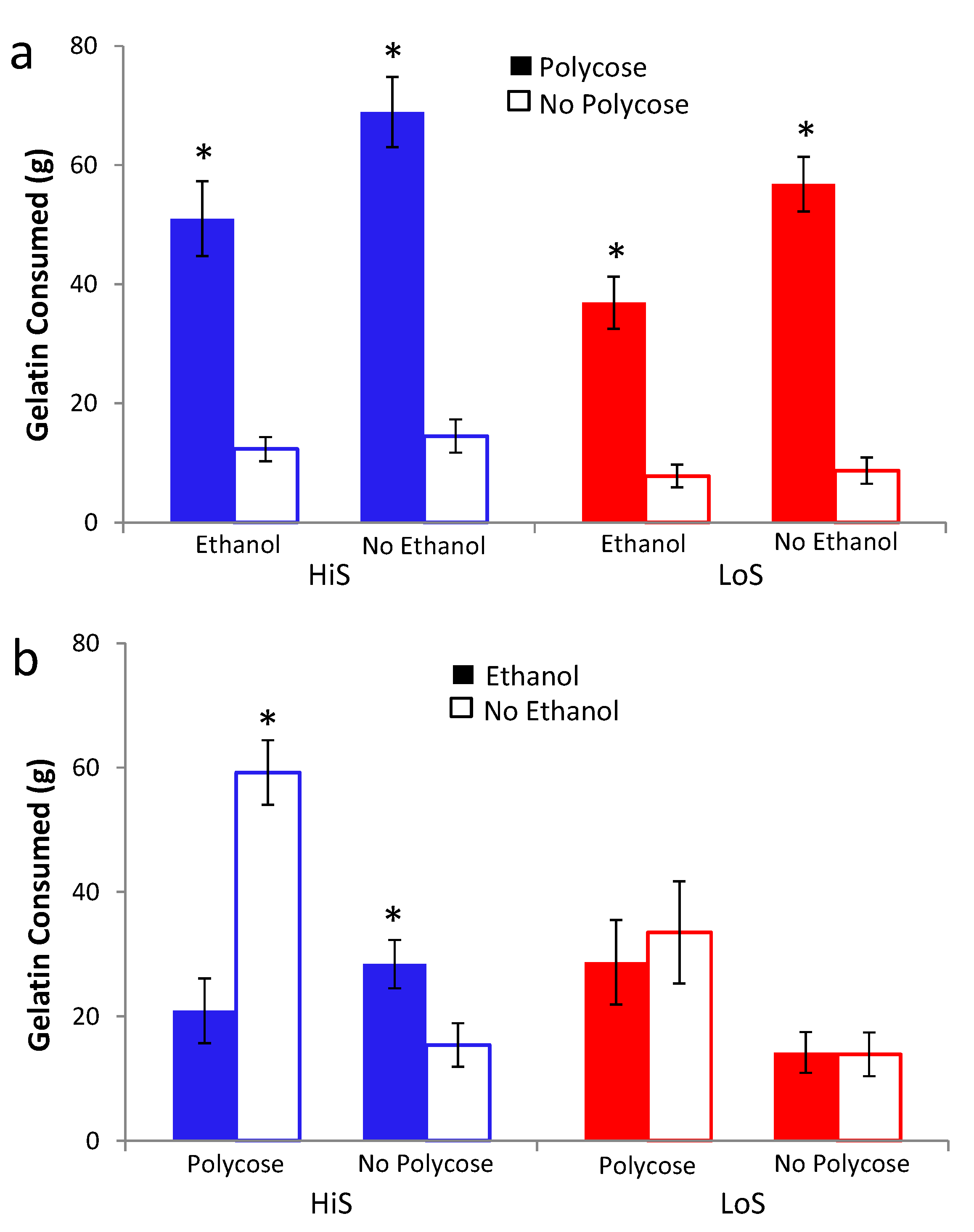
Intake of no-polycose saccharin/ethanol gelatin during conditioning in Experiment 7b is shown in
Figure 7c. One LoS rat, who ate an average amount of ethanol gelatin but rejected most of the no-ethanol gelatin ration, was excluded from the following analyses. HiS rats consumed more saccharin gelatin than did LoS rats. Although intake by the latter trends upwards over days, no effects in a line × day ANOVA were significant other than the line main effect,
F(1,21) = 17.84. Ethanol-paired flavor preference is shown in
Figure 7d. LoS rats preferred the ethanol-paired flavor in gelatin but not fluid; within-group variability was lower for saccharin than for polycose gelatin. HiS rats were indifferent regardless of test medium. A line × test medium ANOVA yielded a main effect of line,
F(1,21) = 4.33. Comparison of each mean to 0.5 confirmed LoS rats’ preference for the ethanol-paired flavor in saccharin gelatin,
t(9) = 3.78. (Note: The excluded LoS rat’s preference for the ethanol-paired flavor was 0.95, so including her would only have increased deviation of the mean from 0.5). Their preference was not significant in polycose gelatin or fluid or in any of the three tests among HiS rats.
The yoking direction in Experiment 7b (unlimited ethanol saccharin gelatin, matched no-ethanol saccharin gelatin rations) was an educated guess based on the preceding experiments and a small pilot study that showed both were acceptable in single-jar tests. However, more rats left a higher proportion of their no-ethanol rations uneaten than we had seen in prior experiments. Followup analyses were conducted to determine whether ration-leaving—i.e., relatively less exposure to the no-ethanol flavor—might have differentially biased LoS rats’ preference toward the ethanol-paired flavor. First, a line × day ANOVA was conducted on leftover ration amounts. It yielded no significant effects. Then, intercorrelations between amount of leftover gelatin and flavor preference in the three tests were assessed separately in each line with Pearson’s r. Among LoS rats, ethanol-paired flavor preference in the saccharin gelatin and fluid tests were strongly positively correlated, r(8) = 0.88, but neither was correlated with preference in the polycose gelatin test; none of the three preference scores was correlated with amount of leftover gelatin on any ration trial (Ps > 0.13–0.93). Results were different for HiS rats: Preference in the polycose gelatin test was strongly correlated with preference in both the saccharin gelatin and the fluid test, rs(11) = 0.84 and 0.74, respectively, and the correlation between saccharin gelatin and fluid preferences was nearly significant, r(11) = 0.54, P = 0.055. Moreover, amount of leftover gelatin on the last ration day (and not before) predicted ethanol-paired flavor preference in both the saccharin and polycose gelatin tests, rs(11) = 0.56 and 0.63, respectively, but not in the fluid test.
The greater stability of individual HiS rats’ relative flavor intake across contexts (
i.e., successive and simultaneous flavor presentations, three test media) suggests a difference in the basis on which HiS and LoS rats are responding to the two flavors, and not just a difference in the overall strength of associative conditioning or its expression. Lower reactivity to the relative novelty of test media among HiS rats is one possibility [
21]. Alternatively, the greater stability in the HiS rats’ preference scores might reflect conditioning based on ethanol or stronger unconditioned flavor preferences; it is impossible to tell from these data because ethanol pairing and flavor (cherry, grape) are perfectly confounded at the individual level. That ration leftovers achieved significance as a predictor of HiS rats’ preference only at the end of conditioning hints at differential conditioning. Experiment 10 (below) explored the last idea.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 . Preferences/increases are shown in
. Preferences/increases are shown in  , aversions/decreases in
, aversions/decreases in  .
.