Exercise-Induced Immunodepression in Endurance Athletes and Nutritional Intervention with Carbohydrate, Protein and Fat — What Is Possible, What Is Not?

Abstract
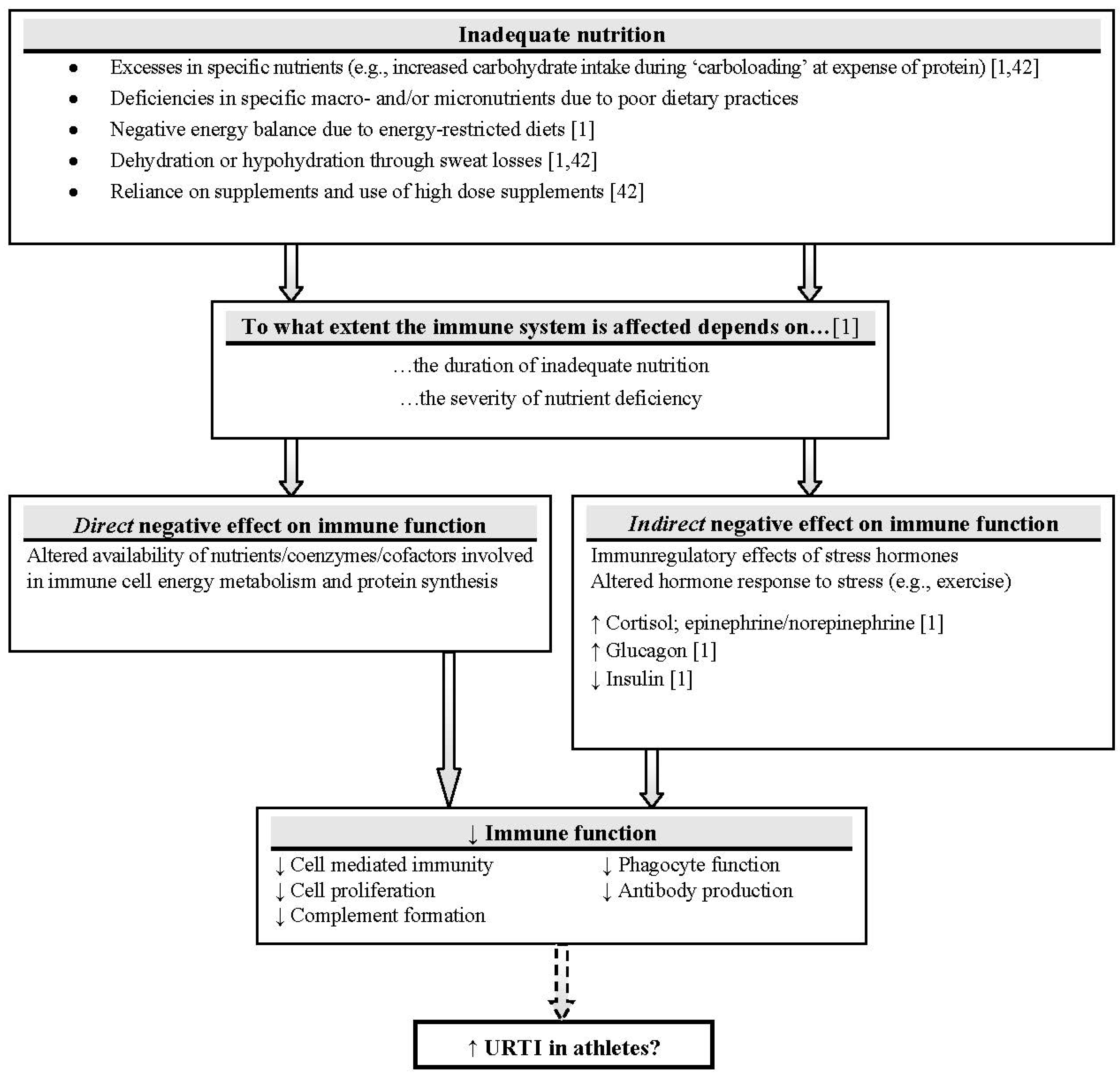
:1. Introduction
1.1. Endurance Exercise and Upper Respiratory Tract Infections
1.2. Effects of Heavy Exercise on Cellular Immune Function
Effects of Acute and Chronic Exercise on Immune Function
- (a) Consequent elevated levels of stress hormones, particularly cortisol;
- (b) Insufficient time between the bouts for immune system to recover fully;
- (c) Plasma glutamine levels may become chronically depressed.
1.3. Influence of Nutrition State on Pre-Exercise Immune Function
2. Nutritional Modulation of Exercise-Induced Immunodepression

2.1. Carbohydrate, Exercise and Immune Function
2.1.1. Availability of Dietary Carbohydrate

{kind=link}
| Hormonal/Immune Response | High-CHO Diet (70%–77% Dietary Intake from CHO/8.0–12.0 g CHO/kg BW/day) | Low-CHO Diet/Self Selected (7%–11% Dietary Intake from CHO/0.5 g CHO/kg BW/day) | |
|---|---|---|---|
| Glucose response | ↑ Glucose response [51,53] ↔ [56] | ↓ Low blood glucose level [56] | |
| Glutamine level | ↑ Glutamine level [51,57] ↔ [52] | ↓ Glutamine level [52] | |
| Cortisol response | ↓ Plasma cortisol [51,53,58] | ↑ Plasma or salivary cortisol [52,56] | |
| Leukocyte & lymphocyte cell counts | ↔ Circulating leukocytes [52] | ↑ Numbers of neutrophils [52], leukocytes, lymphocytes [51] | |
| ↓ Numbers of neutrophils [53,55]Trend to attenuate post-exercise leukocytosis [55] | |||
| ↑ Neutrophil:lymphocyte ratio [52,54] | |||
| ↔ Post-exercise lymphocytopenia [51] | |||
| Mucosal immunity | ↑ Post-exercise s-IgA concentration than pre-exercise [56] | ||
| Cytokine response | ↑ IL-6, IL-10, IL-1ra [54] |
2.1.2. Carbohydrate Ingestion during Exercise and Immune Function
| Ref. | Mode | Intensity | CHO dose | Leukocytes | Lymphocytes | Neutrophils | NK Cells | Cytokines | Mucosal Immunity | Cortisol | Blood Glucose | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [63] | 2.5 h running | 77% VO2max | 6% | - | - | - | - | ↓ IL-6 post, post 1.5 h | - | ↓ post | ↑ post | |
| ↓ IL-1ra post 1.5 h | ||||||||||||
| [64] | 2.5 h running | 77% VO2max | 6% every 15 min | ↓ (monocytes) post | ↓ post | ↓ post | - | - | - | ↓ post | ↑ post | |
| ↑ post 3 h | ↓ post 1.5 h | |||||||||||
| [65] | 2.5 h running | 77% VO2max | 6% every 15 min | ↓ post | - | - | ↓ post | ↑ post | ||||
| ↔ NKCA | ||||||||||||
| [82] | 2.5 h running or cycling | 75% VO2max | 6% every 15 min | ↓ (monocytes) post | - | ↓ post | - | - | - | ↓ post | ↑ post | |
| [69] | 2.5 h running or cycling | 75% VO2max | 6% | - | ↓ post-exercise lymphocytosis | - | ↓ NKCA | - | - | ↓ post (cycling & running) | ↑ post | |
| [68] | 2 h cycling | 60% VO2max | 6% | - | - | - | - | - | ↓ s-IgA concent-ration during exercise | - | ↑ post | |
| [66] | 2 h rowing | - | - | ↓ (monocytes) post | - | ↓ post | - | ↓ IL-1ra post | - | ↔ | ↑ post | |
| ↔ IL-6 post | ||||||||||||
| ↔ IL-8 post | ||||||||||||
| ↔ TNF-α | ||||||||||||
| [83] | 1 h cycling and running | At individual’s lactate threshold | 6.4% 12 mL/kg BW | - | - | - | - | ↓ IL-6 post in cycling & running | - | - | ↑ post | |
| [50] | Marathon run | - | 6% | - | - | - | - | ↓ IL-10 post | - | ↓ post | ↑ post | |
| ↓ IL-1ra post | ||||||||||||
| ↔ IL-6 post | ||||||||||||
| ↑ IL-8 post | ||||||||||||
| [84] | 6 × 20 min cycling | 90% of individual’s lactate threshold | 1 g/kg BW/h (10%) | - | - | - | - | ↓ cytokine response post | - | ↓ post | ↑ post | |
| [85] | 6 × 15 min intermittent running | - | Every 15 min | - | - | ↓ post 30 min | - | ↓ IL-6 post 30 min | - | ↓ post 30 min | ↑ post | |
| ↔ TNF-α post | ||||||||||||
| [70] | 3 h run | 70% VO2max | 6% every 15 min | ↓ post | ↓ post | - | - | ↓ IL-6 post | ↔ | ↓ post | ↑ post | |
| ↓ IL-10 post | ||||||||||||
| ↓ IL-1ra post | ||||||||||||
| ↔ IL-8 post | ||||||||||||
| [86] | 2 h cycling | - | 6.4% | - | - | - | - | ↓ IL-6 post | - | - | ↑ post | |
| ↓ muscle derived IL-6 post | ||||||||||||
| [87] | 2 h cycling | 75% VO2max | 6.4% | - | - | ↓ post | - | - | - | ↓ post | ↑ post | |
| ↓ post 1 h | ||||||||||||
| [75] | 2.5 h cycling | 85% VO2max | 6% | ↓ post | ↔ | ↓ post | - | - | - | ↓ post | ↑ post | |
| ↓ post 1 h | ↓ post 1 h | ↓ post 1 h | ||||||||||
| [76] | Marathon run | - | 6% | ↓ (monocytes) post | ↔ on post lymphocytopenia | ↓ post | ↔ | ↔ cytokine response post | - | ↓ post | ↑ post | |
| [77] | 2 × 1 h cycling | 75%–80% VO2max | 60 g/h | - | ↔ | - | ↔ | - | - | - | ↑ post | |
| [71] | 1 h cycling | 70% VO2max | 6% | - | ↓ post | ↓ post | ↓ post | ↔ IL-6 post | - | -. | ↑ post | |
| ↓ post 1 h | ↔ TNF-α post | |||||||||||
| [88] | 2.5 h cycling | 60% Wmax | 6% | - | - | - | - | ↓ IL-6 post | - | ↓ post | ↑ post | |
| ↓ IL-10 post | ||||||||||||
| ↓ IL-1ra post | ||||||||||||
| ↔ IL-8 post | ||||||||||||
| ↔ muscle IL-6, IL-8, TNF-α post | ||||||||||||
| [80] | 2 × 1.5 h cycling | 60% VO2max | 10% | - | - | - | - | - | ↔ | ↓ post | ↑ post | |
| [72] | 2.5 h cycling | 65% VO2max | 6.4%12.8% | ↓ post in 6.4% + 12.8% | ↓ post in 6.4% + 12.8% (T-cell subpopulations) | ↓ post in 6.4% + 12.8% | - | - | - | ↓ post in 6.4% + 12.8% | ↑ post in 6.4% + 12.8% | |
| ↓ post 2 h in 6.4% + 12.8% | ↓ post 2 h in 6.4% + 12.8% | |||||||||||
| ↓ post 2 h in 6.4% + 12.8% | ||||||||||||
| ↔ between 6.4% + 12.8% | ↔ between 6.4% + 12.8% | |||||||||||
| ↔ between 6.4% + 12.8% | ||||||||||||
| [67] | 1.5 h running on two consecutive days | 70%–80% VO2max | 6.4% | ↓ total count (D1 + D2) | ND (D1 + D2) but ↓ T-cell count post (D1 + D2) | ↓ post (D1 + D2) | ↔ | - | - | ↔ (D1 + D2) | ↑ post (D1 + D2) | |
| ↓ (monocytes) post (D1) | ||||||||||||
| ↓ post 1 h (D1 + D2) | ||||||||||||
| DAY1 (D1) | ↓ (monocytes) post 1 h (D1 + D2) | |||||||||||
| DAY2 (D2) | ||||||||||||
| [74] | 4 h cycling | 70% of individual anaerobic threshold | 6%12% | ↓ post in 6% + 12% | ↔ but trend to attenuate lymphocytopenia in 6% + 12% post 1 h | ↓ post in 6% + 12% | ↔ | ↓ IL-6 post in 6% + 12% | - | ↓ post in 6% + 12% | ↑ post in 6% + 12% | |
| ↓ IL-6 post 1 h in 6% + 12% | ↓ post 1 h in 6% + 12% | |||||||||||
| ↓ post 1 h in 6% + 12% | ||||||||||||
| ↔ between 6% + 12% | ↓ post 1 h in 6% + 12% | |||||||||||
| ↔ between 6% + 12% | ||||||||||||
| ↔ between 6% + 12% | ↔ between 6% + 12% | |||||||||||
| [78] | 2 h cycling | 64% Wmax | 6% every 15 min | ↓ (monocytes) post | ↔ | ↓ post | ↔ | - | - | ↓ post | ↑ post | |
| [89] | Duathlon (5 km run—20 km cycling—2.5 km run) | - | 6% malto-dextrin | - | - | - | - | - | - | ↓ post | ↑ post | |
| [73] | 2 h cycling | 65% VO2max | 6% CHO | ↓ post in experimental conditions with CHO | ↓ post in experimental conditions with CHO | ↓ post in experimental conditions with CHO | - | - | - | ↓ post in CHO/PLA condition | ↑ post in experimental conditions with CHO | |
| 6 mg/kg BW caff eine (CAF) | ↓ post 1 h in experimental conditions with CHO | ↓ post 1 h in experimental conditions with CHO | ||||||||||
| ↔ in CHO/CAF | ||||||||||||
| [68] | 1.5 h cycling followed by 16 km TT | - | 0,24 g/kg BW CHO gel every 15 min | ↓ (monocytes) post | ↔ | ↓ post | - | ↔ IL-6 post | - | ↔ | ↑ post | |
| ↔ IL-10 post | ||||||||||||
| ↔ IL-1ra post | ||||||||||||
| ↔ IL-8 post | ||||||||||||
| [90] | 1.5 h TT running | - | 8% | - | - | - | - | ↓ IL-6 post | - | - | ↑ post | |
| [91] | 2 h run, followed by 5 km TT | 60% VO2max | 8% | - | - | - | - | ↓ IL-6 post | - | - | ↑ post |
2.1.3. Post-Exercise Carbohydrate Ingestion and Immune Function
2.2. Dietary Protein, Amino Acids and Exercise Immune Function
2.2.1. Glutamine & the “Glutamine Hypothesis”
2.2.2. Branched Chain Amino Acids
2.3. Dietary Fat, Fatty Acids and Exercise Immune Function
2.3.1. Dietary Fat Intake
2.3.2. Omega-3 Polyunsaturated Fatty Acids
3. Results
| Nutrient/Strategy | Rationale | Evidence | Likely Impact |
|---|---|---|---|
| Adequate nutrient availability (e.g., micronutrients, fluid) | Adequate nutrient availability maintains immunocompetence | ++++ | ++++ |
| High-CHO diet | Maintained blood glucose level → lower stress hormone levels → attenuated post-ex immune response | ++ | ++ |
| CHO ingestion during exercise | Maintained blood glucose level → lower stress hormone levels → attenuated post-ex immune response | +++ | +++ |
| CHO ingestion post-exercise | Attenuating effect on some immune variables (prevents lymphocytopenia, faster IL-6 return to pre-exercise level) during recovery | − | − |
| Dietary protein availability | Protein is needed for production of immune variables | ++ | ++ |
| Glutamine | Glutamine hypothesis; protein synthesis | − | + |
| BCAA | Precursors of glutamine | ++ | + |
| Creatine | Muscle trauma from heavy exercise → higher inflammatory markers (TNF-α, prostaglandin). | + | + |
| Creatine prevents muscle trauma → attenuated inflammation markers | |||
| Cystine/theanine | Reinforced glutathione synthesis → reinforced anti-oxidative response & better immune function | + | + |
| Dietary fat intake | Low-fat: energy & micronutrient deficiency | ++ | ++ |
| High-fat: excessive intake at cost of protein/CHO | |||
| n-3 PUFA | Anti-inflammatory effects of n-3 PUFA | − | − |
4. Discussion & Future Perspectives
Conflict of Interest
References
- Jeukendrup, A.E.; Gleeson, M. Sport Nutrition: An Introduction to Energy Production and Performance, 2nd ed; Human Kinetics: Champaign, IL, USA, 2010. [Google Scholar]
- Gleeson, M.; Nieman, D.C.; Pedersen, B.K. Exercise, nutrition and immune function. J. Sports Sci. 2004, 22, 115–125. [Google Scholar] [CrossRef]
- Maughan, R.J.; Gleeson, M. The Biochemical Basis of Sports Performance, 2nd ed; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Walsh, N.P.; Gleeson, M.; Pyne, D.B.; Nieman, D.C.; Dhabhar, F.S.; Shephard, R.J.; Oliver, S.J.; Bermon, S.; Kajeniene, A. Position statement. Part two: Maintaining immune health. Exerc. Immunol. Rev. 2011, 17, 64–103. [Google Scholar]
- Peters, E.M.; Bateman, E.D. Ultramarathon running and upper respiratory tract infections. An epidemiological survey. S. Afr. Med. J. 1983, 64, 582–584. [Google Scholar]
- Nieman, D.C.; Johanssen, L.M.; Lee, J.W. Infectious episodes in runners before and after a roadrace. J. Sports Med. Phys. Fitness 1989, 29, 289–296. [Google Scholar]
- Nieman, D.C.; Johanssen, L.M.; Lee, J.W.; Arabatzis, K. Infectious episodes in runners before and after the Los Angeles Marathon. J. Sports Med. Phys. Fitness 1990, 30, 316–328. [Google Scholar]
- Heath, G.W.; Ford, E.S.; Craven, T.E.; Macera, C.A.; Jackson, K.L.; Pate, R.R. Exercise and the incidence of upper respiratory tract infections. Med. Sci. Sports Exerc. 1991, 23, 152–157. [Google Scholar]
- Peters, E.M.; Goetzsche, J.M.; Grobbelaar, B.; Noakes, T.D. Vitamin C supplementation reduces the incidence of postrace symptoms of upper-respiratory-tract infection in ultramarathon runners. Am. J. Clin. Nutr. 1993, 57, 170–174. [Google Scholar]
- Nieman, D.C. Immune function responses to ultramarathon race competition. Med. Sport. 2009, 13, 189–196. [Google Scholar] [CrossRef]
- Gleeson, M.; Bishop, N.C.; Oliveira, M.; Tauler, P. Influence of training load on upper respiratory tract infection incidence and antigen-stimulated cytokine production. Scand. J. Med. Sci. Sports 2011. [Google Scholar] [CrossRef] [Green Version]
- Ekblom, B.; Ekblom, O.; Malm, C. Infectious episodes before and after a marathon race. Scand. J. Med. Sci. Sports 2006, 16, 287–293. [Google Scholar] [CrossRef]
- Pacqué, P.F.; Booth, C.; Ball, M.; Dwyer, D. The effect of an ultra-endurance running race on mucosal and humoral immune function. J. Sports Med. Phys. Fitness 2007, 47, 496–501. [Google Scholar]
- Nieman, D.C.; Pedersen, B.K. Exercise and immune function. Recent developments. Sports Med. 1999, 27, 73–80. [Google Scholar] [CrossRef]
- Walsh, N.P.; Gleeson, M.; Shephard, R.J.; Gleeson, M.; Woods, J.A.; Bishop, N.C.; Fleshner, M.; Green, C.; Pedersen, B.K.; Hoffman-Goetz, L.; et al. Position statement. Part one: Immune function and exercise. Exerc. Immunol. Rev. 2011, 17, 6–63. [Google Scholar]
- Gleeson, M. Immune function in sport and exercise. J. Appl. Physiol. 2007, 103, 693–699. [Google Scholar] [CrossRef]
- Brolinson, P.G.; Elliott, D. Exercise and the immune system. Clin. Sports Med. 2007, 26, 311–319. [Google Scholar] [CrossRef]
- Nieman, D.C. Risk of upper respiratory tract infection in athletes: An epidemiologic and immunologic perspective. J. Athl. Train. 1997, 32, 344–349. [Google Scholar]
- Bishop, N.C. Exercise and Infection Risk. In Immune Function in Sport and Exercise. Advances in Sport and Exercise Science Series; Gleeson, M., Ed.; Churchill Livingstone Elsevier: Edinburgh, UK, 2006; pp. 1–14. [Google Scholar]
- Moreira, A.; Delgado, L.; Moreira, P.; Haahtela, T. Does exercise increase the risk of upper respiratory tract infections? Br. Med. Bull. 2009, 90, 111–131. [Google Scholar] [CrossRef]
- Schwellnus, M.; Lichaba, M.; Derman, E. Respiratory tract symptoms in endurance athletes—A review of causes and consequences. Curr. Allergy Clin. Immunol. 2010, 23, 52–57. [Google Scholar]
- Peters, E.M. Postrace upper respiratory tract ‘infections’ in ultramarathoners—Infection, allergy or inflammation? S. Afr. J. Sports Med. 2004, 16, 3–9. [Google Scholar]
- Nieman, D.C.; Henson, D.A.; Austin, M.D.; Sha, W. Upper respiratory tract infection is reduced in physically fit and active adults. Br. J. Sports Med. 2011, 45, 987–992. [Google Scholar] [CrossRef]
- Nieman, D.C. Special feature for the Olympics: Effects of exercise on the immune system: Exercise effects on systemic immunity. Immunol. Cell Biol. 2000, 78, 496–501. [Google Scholar] [CrossRef]
- Albers, R.; Antoine, J.M.; Bourdet-Sicard, R.; Calder, P.C.; Gleeson, M.; Lesourd, B.; Samartin, S.; Sanderson, I.R.; van Loo, J.; Vas Dias, F.W.; et al. Markers to measure immunomodulation in human nutrition intervention studies. Br. J. Nutr. 2005, 94, 452–481. [Google Scholar]
- Nieman, D.C. Immunonutrition support for athletes. Nutr. Rev. 2008, 66, 310–320. [Google Scholar] [CrossRef]
- Blannin, A.K. Acute Exercise and Innate Immune Function. In Immune Function in Sport and Exercise. Advances in Sport and Exercise Science Series; Gleeson, M., Ed.; Churchill Livingstone Elsevier: Edinburgh, UK, 2006; pp. 67–89. [Google Scholar]
- Timmons, B.W.; Tarnopolsky, M.A.; Snider, D.P.; Bar-Or, O. Immunological changes in response to exercise: Influence of age, puberty, and gender. Med. Sci. Sports Exerc. 2006, 38, 293–304. [Google Scholar]
- Pedersen, B.K.; Hoffman-Goetz, L. Exercise and the immune system: Regulation, integration, and adaptation. Physiol. Rev. 2000, 80, 1055–1081. [Google Scholar]
- Bishop, N.C. Acute Exercise and Aquired Immune Function. In Immune Function in Sport and Exercise. Advances in Sport and Exercise Science Series; Gleeson, M., Ed.; Churchill Livingstone Elsevier: Edinburgh, UK, 2006; pp. 91–113. [Google Scholar]
- Cooper, D.M.; Radom-Aizik, S.; Schwindt, C.; Zaldivar, F., Jr. Dangerous exercise: Lessons learned from dysregulated inflammatory responses to physical activity. J. Appl. Physiol. 2007, 103, 700–709. [Google Scholar] [CrossRef]
- Walsh, N.P. Exercise, Nutrition and Immune Function. I. Macronutrients and Amino Acids. In Immune Function in Sport and Exercise. Advances in Sport and Exercise Science Series; Gleeson, M., Ed.; Churchill Livingstone Elsevier: Edinburgh, UK, 2006; pp. 161–181. [Google Scholar]
- Gleeson, M.; Robson-Ansley, P. Immune Response to Intensified Training and Overtraining. In Immune Function in Sport and Exercise. Advances in Sport and Exercise Science Series; Gleeson, M., Ed.; Churchill Livingstone Elsevier: Edinburgh, UK, 2006; pp. 115–137. [Google Scholar]
- MacKinnon, L.T. Special feature for the Olympics: Effects of exercise on the immune system: Overtraining effects on immunity and performance in athletes. Immunol. Cell Biol. 2000, 78, 502–509. [Google Scholar] [CrossRef]
- Fahlman, M.M.; Engels, H.J. Mucosal IgA and URTI in American college football players: A year longitudinal study. Med. Sci. Sports Exerc. 2005, 37, 374–380. [Google Scholar] [CrossRef]
- Lancaster, G.I.; Halson, S.L.; Khan, Q.; Drysdale, P.; Jeukendrup, A.E.; Drayson, M.T.; Gleeson, M. Effects of acute exhaustive exercise and chronic exercise training on type 1 and type 2 T lymphocytes. Exerc. Immunol. Rev. 2004, 10, 91–106. [Google Scholar]
- Gleeson, M.; McDonald, W.A.; Cripps, A.W.; Pyne, D.B.; Clancy, R.L.; Fricker, P.A. The effect on immunity of long-term intensive training in elite swimmers. Clin. Exp. Immunol. 1995, 102, 210–216. [Google Scholar]
- Ronsen, O.; Kjeldsen-Kragh, J.; Haug, E.; Bahr, R.; Pedersen, B.K. Recovery time affects immunoendocrine responses to a second bout of endurance exercise. Am. J. Physiol. Cell Physiol. 2002, 283, C1612–C1620. [Google Scholar]
- Degerstrom, J.; Osterud, B. Increased inflammatory response of blood cells to repeated bout of endurance exercise. Med. Sci. Sports Exerc. 2006, 38, 1297–1303. [Google Scholar] [CrossRef]
- Calder, P.C.; Jackson, A.A. Undernutrition, infection and immune function. Nutr. Res. Rev. 2000, 13, 3–29. [Google Scholar] [CrossRef]
- Burke, L. Practical Sports Nutrition; Human Kinetics: Champaign, IL, USA, 2007. [Google Scholar]
- Calder, P.C. Immunodepression and Exercise: The Evidence & an Evaluation of Preventive Nutritional Strategies. In Exercise and Immunity in Athletic Performance and a Healthy Life, Proceedings of the 10th ISEI Symposium, Oxford, UK, 11–13 July 2011. Invited Speaker Presentation #7..
- Pyne, D.B. Nutrition for the Athlete’s Immune System: Eating to Stay Well during Training and Competition. In Clinical Sports Nutrition, 3rd; Burke, L., Deakin, V., Eds.; McGraw-Hill: Sydney, Australia, 2006; pp. 581–588. [Google Scholar]
- Scott, J.P.; Sale, C.; Greeves, J.P.; Casey, A.; Dutton, J.; Fraser, W.D. Effect of exercise intensity on the cytokine response to an acute bout of running. Med. Sci. Sports Exerc. 2011, 43, 2297–2306. [Google Scholar] [CrossRef]
- Nieman, D.C.; Konrad, M.; Henson, D.A.; Kennerly, K.; Shanely, R.A.; Wallner-Liebmann, S.J. Variance in the acute inflammatory response to prolonged cycling is linked to exercise intensity. J. Interferon Cytokine Res. 2012, 32, 12–17. [Google Scholar] [CrossRef]
- Peake, J.M.; Suzuki, K.; Hordern, M.; Wilson, G.; Nosaka, K.; Coombes, J.S. Plasma cytokine changes in relation to exercise intensity and muscle damage. Eur. J. Appl. Physiol. 2005, 95, 514–521. [Google Scholar] [CrossRef]
- Gleeson, M.; Bishop, N.C. Special feature for the Olympics: Effects of exercise on the immune system: Modification of immune responses to exercise by carbohydrate, glutamine and anti-oxidant supplements. Immunol. Cell Biol. 2000, 78, 554–561. [Google Scholar] [CrossRef]
- Nieman, D.C. Exercise Testing and Prescription: A Health-Related Approach, 7th ed; McGraw-Hill: New York, NY, USA, 2011. [Google Scholar]
- Burke, L.M. Fueling strategies to optimize performance: Training high or training low? Scand. J. Med. Sci. Sports 2010, 20, 48–58. [Google Scholar] [CrossRef]
- Nieman, D.C.; Henson, D.A.; Smith, L.L.; Utter, A.C.; Vinci, D.M.; Davis, J.M.; Kaminsky, D.E.; Shute, M. Cytokine changes after a marathon race. J. Appl. Physiol. 2001, 91, 109–114. [Google Scholar]
- Mitchell, J.B.; Pizza, F.X.; Paquet, A.; Davis, B.J.; Forrest, M.B.; Braun, W.A. Influence of carbohydrate status on immune responses before and after endurance exercise. J. Appl. Physiol. 1998, 84, 1917–1925. [Google Scholar]
- Gleeson, M.; Blannin, A.K.; Walsh, N.P.; Bishop, N.C.; Clark, A.M. Effect of low- and high-carbohydrate diets on the plasma glutamine and circulating leukocyte responses to exercise. Int. J. Sport Nutr. 1998, 8, 49–59. [Google Scholar]
- Bishop, N.C.; Walsh, N.P.; Haines, D.L.; Richards, E.E.; Gleeson, M. Pre-Exercise carbohydrate status and immune responses to prolonged cycling: I. Effect on neutrophil degranulation. Int. J. Sport Nutr. Exerc. Metab. 2001, 11, 490–502. [Google Scholar]
- Bishop, N.C.; Walsh, N.P.; Haines, D.L.; Richards, E.E.; Gleeson, M. Pre-Exercise carbohydrate status and immune responses to prolonged cycling: II. Effect on plasma cytokine concentration. Int. J. Sport Nutr. Exerc. Metab. 2001, 11, 503–512. [Google Scholar]
- Close, G.L.; Ashton, T.; Cable, T.; Doran, D.; Noyes, C.; McArdle, F.; MacLaren, D.P. Effects of dietary carbohydrate on delayed onset muscle soreness and reactive oxygen species after contraction induced muscle damage. Br. J. Sports Med. 2005, 39, 948–953. [Google Scholar] [CrossRef]
- Costa, R.J.; Jones, G.E.; Lamb, K.L.; Coleman, R.; Williams, J.H. The effects of a high carbohydrate diet on cortisol and salivary immunoglobulin A (s-IgA) during a period of increase exercise workload amongst Olympic and Ironman triathletes. Int. J. Sports Med. 2005, 26, 880–885. [Google Scholar] [CrossRef]
- Blanchard, M.A.; Jordan, G.; Desbrow, B.; MacKinnon, L.T.; Jenkins, D.G. The influence of diet and exercise on muscle and plasma glutamine concentrations. Med. Sci. Sports Exerc. 2001, 33, 69–74. [Google Scholar]
- De Sousa, M.V.; Madsen, K.; Simoes, H.G.; Pereira, R.M.; Negrao, C.E.; Mendonca, R.Z.; Takayama, L.; Fukui, R.; da Silva, M.E. Effects of carbohydrate supplementation on competitive runners undergoing overload training followed by a session of intermittent exercise. Eur. J. Appl. Physiol. 2010, 109, 507–516. [Google Scholar] [CrossRef]
- Chen, Y.J.; Wong, S.H.; Wong, C.K.; Lam, C.W.; Huang, Y.J.; Siu, P.M. The effect of a pre-exercise carbohydrate meal on immune responses to an endurance performance run. Br. J. Nutr. 2008, 100, 1260–1268. [Google Scholar] [CrossRef]
- Chen, Y.J.; Wong, S.H.; Chan, C.O.; Wong, C.K.; Lam, C.W.; Siu, P.M. Effects of glycemic index meal and CHO-electrolyte drink on cytokine response and run performance in endurance athletes. J. Sci. Med. Sport 2009, 12, 697–703. [Google Scholar] [CrossRef]
- Li, T.L.; Wu, C.L.; Gleeson, M.; Williams, C. The effects of pre-exercise high carbohydrate meals with different glycemic indices on blood leukocyte redistribution, IL-6, and hormonal responses during a subsequent prolonged exercise. Int. J. Sport Nutr. Exerc. Metab. 2004, 14, 647–656. [Google Scholar]
- Gleeson, M. Can nutrition limit exercise-induced immunodepression? Nutr. Rev. 2006, 64, 119–131. [Google Scholar] [CrossRef]
- Nehlsen-Cannarella, S.L.; Fagoaga, O.R.; Nieman, D.C.; Henson, D.A.; Butterworth, D.E.; Schmitt, R.L.; Bailey, E.M.; Warren, B.J.; Utter, A.; Davis, J.M. Carbohydrate and the cytokine response to 2.5 h of running. J. Appl. Physiol. 1997, 82, 1662–1667. [Google Scholar]
- Nieman, D.C.; Fagoaga, O.R.; Butterworth, D.E.; Warren, B.J.; Utter, A.; Davis, J.M.; Henson, D.A.; Nehlsen-Cannarella, S.L. Carbohydrate supplementation affects blood granulocyte and monocyte trafficking but not function after 2.5 h or running. Am. J. Clin. Nutr. 1997, 66, 153–159. [Google Scholar]
- Nieman, D.C.; Henson, D.A.; Garner, E.B.; Butterworth, D.E.; Warren, B.J.; Utter, A.; Davis, J.M.; Fagoaga, O.R.; Nehlsen-Cannarella, S.L. Carbohydrate affects natural killer cell redistribution but not activity after running. Med. Sci. Sports Exerc. 1997, 29, 1318–1324. [Google Scholar] [CrossRef]
- Henson, D.A.; Nieman, D.C.; Nehlsen-Cannarella, S.L.; Fagoaga, O.R.; Shannon, M.; Bolton, M.R.; Davis, J.M.; Gaffney, C.T.; Kelln, W.J.; Austin, M.D.; et al. Influence of carbohydrate on cytokine and phagocytic responses to 2 h of rowing. Med. Sci. Sports Exerc. 2000, 32, 1384–1389. [Google Scholar] [CrossRef]
- Bishop, N.C.; Walker, G.J.; Bowley, L.A.; Evans, K.F.; Molyneux, K.; Wallace, F.A.; Smith, A.C. Lymphocyte responses to influenza and tetanus toxoid in vitro following intensive exercise and carbohydrate ingestion on consecutive days. J. Appl. Physiol. 2005, 99, 1327–1335. [Google Scholar]
- Peake, J.; Peiffer, J.J.; Abbiss, C.R.; Nosaka, K.; Laursen, P.B.; Suzuki, K. Carbohydrate gel ingestion and immunoendocrine responses to cycling in temperate and hot conditions. Int. J. Sport Nutr. Exerc. Metab. 2008, 18, 229–246. [Google Scholar]
- Henson, D.A.; Nieman, D.C.; Blodgett, A.D.; Butterworth, D.E.; Utter, A.; Davis, J.M.; Sonnenfeld, G.; Morton, D.S.; Fagoaga, O.R.; Nehlsen-Cannarella, S.L. Influence of exercise mode and carbohydrate on the immune response to prolonged exercise. Int. J. Sport Nutr. 1999, 9, 213–228. [Google Scholar]
- Nieman, D.C.; Davis, J.M.; Henson, D.A.; Walberg-Rankin, J.; Shute, M.; Dumke, C.L.; Utter, A.C.; Vinci, D.M.; Carson, J.A.; Brown, A.; et al. Carbohydrate ingestion influences skeletal muscle cytokine mRNA and plasma cytokine levels after a 3-h run. J. Appl. Physiol. 2003, 94, 1917–1925. [Google Scholar]
- Timmons, B.W.; Tarnopolsky, M.A.; Bar-Or, O. Immune responses to strenuous exercise and carbohydrate intake in boys and men. Pediatr. Res. 2004, 56, 227–234. [Google Scholar]
- Lancaster, G.I.; Khan, Q.; Drysdale, P.T.; Wallace, F.; Jeukendrup, A.E.; Drayson, M.T.; Gleeson, M. Effect of prolonged exercise and carbohydrate ingestion on type 1 and type 2 T lymphocyte distribution and intracellular cytokine production in humans. J. Appl. Physiol. 2005, 98, 565–571. [Google Scholar]
- Walker, G.J.; Finlay, O.; Griffiths, H.; Sylvester, J.; Williams, M.; Bishop, N.C. Immunoendocrine response to cycling following ingestion of caffeine and carbohydrate. Med. Sci. Sports Exerc. 2007, 39, 1554–1560. [Google Scholar] [CrossRef]
- Scharhag, J.; Meyer, T.; Auracher, M.; Gabriel, H.H.; Kindermann, W. Effects of graded carbohydrate supplementation on the immune response in cycling. Med. Sci. Sports Exerc. 2006, 38, 286–292. [Google Scholar] [CrossRef]
- Green, K.J.; Croaker, S.J.; Rowbottom, D.G. Carbohydrate supplementation and exercise-induced changes in T-lymphocyte function. J. Appl. Physiol. 2003, 95, 1216–1223. [Google Scholar]
- Henson, D.A.; Nieman, D.C.; Pistilli, E.E.; Schilling, B.; Colacino, A.; Utter, A.C.; Fagoaga, O.R.; Vinci, D.M.; Nehlsen-Cannarella, S.L. Influence of carbohydrate and age on lymphocyte function following a marathon. Int. J. Sport Nutr. Exerc. Metab. 2004, 14, 308–322. [Google Scholar]
- McFarlin, B.K.; Flynn, M.G.; Stewart, L.K.; Timmerman, K.L. Carbohydrate intake during endurance exercise increases natural killer cell responsiveness to IL-2. J. Appl. Physiol. 2004, 96, 271–275. [Google Scholar]
- Nieman, D.C.; Henson, D.A.; Gojanovich, G.; Davis, J.M.; Murphy, E.A.; Mayer, E.P.; Pearce, S.; Dumke, C.L.; Utter, A.C.; McAnulty, S.R.; et al. Influence of carbohydrate on immune function following 2 h cycling. Res. Sports Med. 2006, 14, 225–237. [Google Scholar] [CrossRef]
- Gleeson, M. Introduction to the Immune System. In Immune Function in Sport and Exercise. Advances in Sport and Exercise Science Series; Gleeson, M., Ed.; Churchill Livingstone Elsevier: Edinburgh, UK, 2006; pp. 15–43. [Google Scholar]
- Li, T.L.; Gleeson, M. The effects of carbohydrate supplementation during repeated bouts of prolonged exercise on saliva flow rate and immunoglobulin A. J. Sports Sci. 2005, 23, 713–722. [Google Scholar] [CrossRef]
- Bishop, N.C.; Blannin, A.K.; Armstrong, E.; Rickman, M.; Gleeson, M. Carbohydrate and fluid intake affect the saliva flow rate and IgA response to cycling. Med. Sci. Sports Exerc. 2000, 32, 2046–2051. [Google Scholar]
- Nieman, D.C.; Nehlsen-Cannarella, S.L.; Fagoaga, O.R.; Henson, D.A.; Utter, A.; Davis, J.M.; Williams, F.; Butterworth, D.E. Effects of mode and carbohydrate on the granulocyte and monocyte response to intensive, prolonged exercise. J. Appl. Physiol. 1998, 84, 1252–1259. [Google Scholar]
- Starkie, R.L.; Arkinstall, M.J.; Koukoulas, I.; Hawley, J.A.; Febbraio, M.A. Carbohydrate ingestion attenuates the increase in plasma interleukin-6, but not skeletal muscle interleukin-6 mRNA, during exercise in humans. J. Physiol. 2001, 533, 585–591. [Google Scholar]
- Bacurau, R.F.; Bassit, R.A.; Sawada, L.; Navarro, F.; Martins, E., Jr.; Costa Rosa, L.F. Carbohydrate supplementation during intense exercise and the immune response of cyclists. Clin. Nutr. 2002, 21, 423–429. [Google Scholar] [CrossRef]
- Bishop, N.C.; Gleeson, M.; Nicholas, C.W.; Ali, A. Influence of carbohydrate supplementation on plasma cytokine and neutrophil degranulation responses to high intensity intermittent exercise. Int. J. Sport Nutr. Exerc. Metab. 2002, 12, 145–156. [Google Scholar]
- Febbraio, M.A.; Steensberg, A.; Keller, C.; Starkie, R.L.; Nielsen, H.B.; Krustrup, P.; Ott, P.; Secher, N.H.; Pedersen, B.K. Glucose ingestion attenuates interleukin-6 release from contracting skeletal muscle in humans. J. Physiol. 2003, 549, 607–612. [Google Scholar]
- Bishop, N.C.; Walsh, N.P.; Scanlon, G.A. Effect of prolonged exercise and carbohydrate on total neutrophil elastase content. Med. Sci. Sports Exerc. 2003, 35, 1326–1332. [Google Scholar] [CrossRef]
- Nieman, D.C.; Davis, J.M.; Henson, D.A.; Gross, S.J.; Dumke, C.L.; Utter, A.C.; Vinci, D.M.; Carson, J.A.; Brown, A.; McAnulty, S.R.; et al. Muscle cytokine mRNA changes after 2.5 h of cycling: Influence of carbohydrate. Med. Sci. Sports Exerc. 2005, 37, 1283–1290. [Google Scholar]
- Mamus, R.T.; Dos Santos, M.G.; Campbell, B.; Kreider, R. Biochemical effects of carbohydrate supplementation in a simulated competition of short terrestrial duathlon. J. Int. Soc. Sports Nutr. 2006, 3, 6–11. [Google Scholar] [CrossRef]
- Robson-Ansley, P.; Barwood, M.; Eglin, C.; Ansley, L. The effect of carbohydrate ingestion on the interleukin-6 response to a 90-minute run time trial. Int. J. Sports Physiol. Perform. 2009, 4, 186–194. [Google Scholar]
- Robson-Ansley, P.; Walshe, I.; Ward, D. The effect of carbohydrate ingestion on plasma interleukin-6, hepcidin and iron concentrations following prolonged exercise. Cytokine 2011, 53, 196–200. [Google Scholar] [CrossRef]
- Cox, A.J.; Pyne, D.B.; Cox, G.R.; Callister, R.; Gleeson, M. Effect of Chronic Carbohydrate Consumption on Cytokine Responses to Cycle Ergometry. In Inflammation in Exercise: Friend or Foe? Proceeding of the 8th International Society of Exercise and Immunology Symposium, Sendai, Japan, 25–27 October 2007.
- Cox, A.J.; Pyne, D.B.; Cox, G.R.; Callister, R.; Gleeson, M. Pre-Exercise carbohydrate status influences carbohydrate-mediated attenuation of post-exercise cytokine responses. Int. J. Sports Med. 2008, 29, 1003–1009. [Google Scholar] [CrossRef]
- Kerksick, C.; Harvey, T.; Stout, J.; Campbell, B.; Wilborn, C.; Kreider, R.; Kalman, D.; Ziegenfuss, T.; Lopez, H.; Landis, J.; et al. International society of sports nutrition position stand: Nutrient timing. J. Int. Soc. Sports Nutr. 2008, 5, 17. [Google Scholar] [CrossRef]
- Ferguson-Stegall, L.; McCleave, E.; Ding, Z.; Doerner, P.G., III; Liu, Y.; Wang, B.; Healy, M.; Kleinert, M.; Dessard, B.; Lassiter, D.G.; et al. Aerobic exercise training adaptations are increased by postexercise carbohydrate-protein supplementation. J. Nutr. Metab. 2011, 2011. [Google Scholar] [CrossRef]
- Costa, R.J.; Walters, R.; Bilzon, J.L.; Walsh, N.P. Effects of immediate postexercise carbohydrate ingestion with and without protein on neutrophil degranulation. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 205–213. [Google Scholar]
- Gusba, J.E.; Wilson, R.J.; Robinson, D.L.; Graham, T.E. Interleukin-6 and its mRNA responses in exercise and recovery: Relationship to muscle glycogen. Scand. J. Med. Sci. Sports 2008, 18, 77–85. [Google Scholar]
- Li, P.; Yin, Y.L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef]
- American Dietetic Association; Dietitians of Canada; American College of Sports Medicine; Rodriguez, N.R.; Di Marco, N.M.; Langley, S. American college of sports medicine position stand. Nutrition and athletic performance. Med. Sci. Sports Exerc. 2009, 41, 709–731. [Google Scholar] [CrossRef]
- Murakami, S.; Kurihara, S.; Koikawa, N.; Nakamura, A.; Aoki, K.; Yosigi, H.; Sawaki, K.; Ohtani, M. Effects of oral supplementation with cystine and theanine on the immune function of athletes in endurance exercise: Randomized, double-blind, placebo-controlled trial. Biosci. Biotechnol. Biochem. 2009, 73, 817–821. [Google Scholar] [CrossRef]
- Murakami, S.; Kurihara, S.; Titchenal, C.A.; Ohtani, M. Suppression of exercise-induced neutrophilia and lymphopenia in athletes by cystine/theanine intake: A randomized, double-blind, placebo-controlled trial. J. Int. Soc. Sports Nutr. 2010, 7, 23. [Google Scholar]
- Bassit, R.A.; Curi, R.; Costa Rosa, L.F. Creatine supplementation reduces plasma levels of pro-inflammatory cytokines and PGE2 after a half-ironman competition. Amino Acids 2008, 35, 425–431. [Google Scholar] [CrossRef]
- Santos, R.V.; Bassit, R.A.; Caperuto, E.C.; Costa Rosa, L.F. The effect of creatine supplementation upon inflammatory and muscle soreness markers after a 30km race. Life Sci. 2004, 75, 1917–1924. [Google Scholar] [CrossRef]
- Braun, W.A.; von Duvillard, S.P. Influence of carbohydrate delivery on the immune response during exercise and recovery from exercise. Nutrition 2004, 20, 645–650. [Google Scholar]
- Gleeson, M. Dosing and efficacy of glutamine supplementation in human exercise and sport training. J. Nutr. 2008, 138, 2045S–2049S. [Google Scholar]
- Castell, L.M. Can glutamine modify the apparent immunodepression observed after prolonged, exhaustive exercise? Nutrition 2002, 18, 371–375. [Google Scholar] [CrossRef]
- Nieman, D.C. Marathon training and immune function. Sports Med. 2007, 37, 412–415. [Google Scholar] [CrossRef]
- Castell, L.M.; Poortmans, J.R.; Newsholme, E.A. Does glutamine have a role in reducing infections in athletes? Eur. J. Appl. Physiol. Occup. Physiol. 1996, 73, 488–490. [Google Scholar] [CrossRef]
- Krzywkowski, K.; Petersen, E.W.; Ostrowski, K.; Link-Amster, H.; Boza, J.; Halkjaer-Kristensen, J.; Pedersen, B.K. Effect of glutamine and protein supplementation on exercise-induced decreases in salivary IgA. J. Appl. Physiol. 2001, 91, 832–838. [Google Scholar]
- Roshan, V.D.; Barzegarzadeh, H. The effect of the short-term glutamine supplementation on exhaustive exercise-induced changes in immune system of active boys. World J. Sport Sci. 2009, 2, 222–230. [Google Scholar]
- Krieger, J.W.; Crowe, M.; Blank, S.E. Chronic glutamine supplementation increases nasal but not salivary IgA during 9 days of interval training. J. Appl. Physiol. 2004, 97, 585–591. [Google Scholar]
- Hoffman, J.R.; Ratamess, N.A.; Kang, J.; Rashti, S.L.; Kelly, N.; Gonzalez, A.M.; Stec, M.; Anderson, S.; Bailey, B.L.; Yamamoto, L.M.; et al. Examination of the efficacy of acute L-alanyl-L-glutamine ingestion during hydration stress in endurance exercise. J. Int. Soc. Sports Nutr. 2010, 7. [Google Scholar] [CrossRef]
- Hiscock, N.; Petersen, E.W.; Krzywkowski, K.; Boza, J.; Halkjaer-Kristensen, J.; Pedersen, B.K. Glutamine supplementation further enhances exercise-induced plasma IL-6. J. Appl. Physiol. 2003, 95, 145–148. [Google Scholar]
- Krzywkowski, K.; Petersen, E.W.; Ostrowski, K.; Kristensen, J.H.; Boza, J.; Pedersen, B.K. Effect of glutamine supplementation on exercise-induced changes in lymphocyte function. Am. J. Physiol. Cell Physiol. 2001, 281, C1259–C1265. [Google Scholar]
- Walsh, N.P.; Blannin, A.K.; Bishop, N.C.; Robson, P.J.; Gleeson, M. Effect of oral glutamine supplementation on human neutrophil lipopolysaccharide-stimulated degranulation following prolonged exercise. Int. J. Sport Nutr. Exerc. Metab. 2000, 10, 39–50. [Google Scholar]
- Hiscock, N.; Pedersen, B.K. Exercise-Induced immunodepression—plasma glutamine is not the link. J. Appl. Physiol. 2002, 93, 813–822. [Google Scholar]
- Negro, M.; Giardina, S.; Marzani, B.; Marzatico, F. Branched-Chain amino acid supplementation does not enhance athletic performance but affects muscle recovery and the immune system. J. Sports Med. Phys. Fit. 2008, 48, 347–351. [Google Scholar]
- Calder, P.C. Branched-Chain amino acids and immunity. J. Nutr. 2006, 136, 288S–293S. [Google Scholar]
- Bassit, R.A.; Sawada, L.A.; Bacurau, R.F.; Navarro, F.; Costa Rosa, L.F. The effect of BCAA supplementation upon the immune response of triathletes. Med. Sci. Sports Exerc. 2000, 32, 1214–1219. [Google Scholar]
- Bassit, R.A.; Sawada, L.A.; Bacurau, R.F.; Navarro, F.; Martins, E., Jr.; Santos, R.V.; Caperuto, E.C.; Rogeri, P.; Costa Rosa, L.F. Branched-Chain amino acid supplementation and the immune response of long-distance athletes. Nutrition 2002, 18, 376–379. [Google Scholar]
- Gleeson, M. Interrelationship between physical activity and branched-chain amino acids. J. Nutr. 2005, 135, 1591S–1595S. [Google Scholar]
- Galli, C.; Calder, P.C. Effects of fat and fatty acid intake on inflammatory and immune responses: A critical review. Ann. Nutr. Metab. 2009, 55, 123–139. [Google Scholar] [CrossRef]
- Wall, R.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Fatty acids from fish: The anti-inflammatory potential of long-chain omega-3 fatty acids. Nutr. Rev. 2010, 68, 280–289. [Google Scholar]
- Meksawan, K.; Venkatraman, J.T.; Awad, A.B.; Pendergast, D.R. Effect of dietary fat intake and exercise on inflammatory mediators of the immune system in sedentary men and women. J. Am. Coll. Nutr. 2004, 23, 331–340. [Google Scholar]
- Venkatraman, J.T.; Feng, X.; Pendergast, D. Effects of dietary fat and endurance exercise on plasma cortisol, prostaglandin E2, interferon-gamma and lipid peroxides in runners. J. Am. Coll. Nutr. 2001, 20, 529–536. [Google Scholar]
- Pedersen, B.K.; Helge, J.W.; Richter, E.A.; Rohde, T.; Kiens, B. Training and natural immunity: Effects of diets rich in fat or carbohydrate. Eur. J. Appl. Physiol. 2000, 82, 98–102. [Google Scholar]
- Venkatraman, J.T.; Pendergast, D. Effects of the level of dietary fat intake and endurance exercise on plasma cytokines in runners. Med. Sci. Sports Exerc. 1998, 30, 1198–1204. [Google Scholar] [CrossRef]
- Calder, P.C. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, 1505S–1519S. [Google Scholar]
- Simopoulos, A.P. Omega-3 fatty acids and athletics. Curr. Sports Med. Rep. 2007, 6, 230–236. [Google Scholar] [CrossRef]
- Gray, P.; Gabriel, B.; Thies, F.; Gray, S.R. Fish oil supplementation augments post-exercise immune function in young males. Brain Behav. Immun. 2012, in press.. [Google Scholar]
- Nieman, D.C.; Henson, D.A.; McAnulty, S.R.; Jin, F.; Maxwell, K.R. n-3 polyunsaturated fatty acids do not alter immune and inflammation measures in endurance athletes. Int. J. Sport Nutr. Exerc. Metab. 2009, 19, 536–546. [Google Scholar]
- Callister, R.; Plunkett, B.; Garg, M. Effects of Fish Oil and Lycopene Supplements on Cytokine Response to Exercise. In Exercise Immunology—Emerging Relevance in Clinical Medicine, Proceeding of 9th Symposium of the International Society of Exercise and Immunology, Tübingen, Germany, 21–23 September 2009; p. 59.
- Bloomer, R.J.; Larson, D.E.; Fisher-Wellman, K.H.; Galpin, A.J.; Schilling, B.K. Effect of eicosapentaenoic and docosahexaenoic acid on resting and exercise-induced inflammatory and oxidative stress biomarkers: A randomized, placebo controlled, cross-over study. Lipids Health Dis. 2009, 8, 36. [Google Scholar] [CrossRef]
- Toft, A.D.; Thorn, M.; Ostrowski, K.; Asp, S.; Moller, K.; Iversen, S.; Hermann, C.; Sondergaard, S.R.; Pedersen, B.K. n-3 polyunsaturated fatty acids do not affect cytokine response to strenuous exercise. J. Appl. Physiol. 2000, 89, 2401–2406. [Google Scholar]
- Bakker, G.C.; van Erk, M.J.; Pellis, L.; Wopereis, S.; Rubingh, C.M.; Cnubben, N.H.; Kooistra, T.; van Ommen, B.; Hendriks, H.F. An antiinflammatory dietary mix modulates inflammation and oxidative and metabolic stress in overweight men: a nutrigenomics approach. Am. J. Clin. Nutr. 2010, 91, 1044–1059. [Google Scholar] [CrossRef]
- Konrad, M.; Nieman, D.C.; Henson, D.A.; Kennerly, K.M.; Jin, F.; Wallner-Liebmann, S.J. The acute effect of ingesting a quercetin-based supplement on exercise-induced inflammation and immune changes in runners. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 338–346. [Google Scholar]
- Moreira, A.; Kekkonen, R.A.; Delgado, L.; Fonseca, J.; Korpela, R.; Haahtela, T. Nutritional modulation of exercise-induced immunodepression in athletes: A systematic review and meta-analysis. Eur. J. Clin. Nutr. 2007, 61, 443–460. [Google Scholar]
- Petersen, A.M.; Pedersen, B.K. The anti-inflammatory effect of exercise. J. Appl. Physiol. 2005, 98, 1154–1162. [Google Scholar] [CrossRef]
- Davison, G.; Diment, B.C. Bovine colostrum supplementation attenuates the decrease of salivary lysozyme and enhances the recovery of neutrophil function after prolonged exercise. Br. J. Nutr. 2010, 103, 1425–1432. [Google Scholar] [CrossRef]
- Shing, C.M.; Peake, J.; Suzuki, K.; Okutsu, M.; Pereira, R.; Stevenson, L.; Jenkins, D.G.; Coombes, J.S. Effects of bovine colostrum supplementation on immune variables in highly trained cyclists. J. Appl. Physiol. 2007, 102, 1113–1122. [Google Scholar]
- Brinkworth, G.D.; Buckley, J.D. Concentrated bovine colostrum protein supplementation reduces the incidence of self-reported symptoms of upper respiratory tract infection in adult males. Eur. J. Nutr. 2003, 42, 228–232. [Google Scholar] [CrossRef]
- West, N.P.; Pyne, D.B.; Peake, J.M.; Cripps, A.W. Probiotics, immunity and exercise: A review. Exerc. Immunol. Rev. 2009, 15, 107–126. [Google Scholar]
- Gleeson, M.; Bishop, N.C.; Oliveira, M.; Tauler, P. Daily probiotic’s (Lactobacillus casei Shirota) reduction of infection incidence in athletes. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 55–64. [Google Scholar]
- West, N.P.; Pyne, D.B.; Cripps, A.W.; Hopkins, W.G.; Eskesen, D.C.; Jairath, A.; Christophersen, C.T.; Conlon, M.A.; Fricker, P.A. Lactobacillus fermentum (PCC®) supplementation and gastrointestinal and respiratory-tract illness symptoms: A randomised control trial in athletes. Nutr. J. 2011, 10, 30. [Google Scholar]
- Talbott, S.; Talbott, J. Effect of BETA 1,3/1,6 GLUCAN on upper respiratory tract infection symptoms and mood state in marathon athletes. J. Sports Sci. Med. 2009, 8, 509–515. [Google Scholar]
- Nieman, D.C.; Henson, D.A.; McMahon, M.; Wrieden, J.L.; Davis, J.M.; Murphy, E.A.; Gross, S.J.; McAnulty, L.S.; Dumke, C.L. Beta-Glucan, immune function, and upper respiratory tract infections in athletes. Med. Sci. Sports Exerc. 2008, 40, 1463–1471. [Google Scholar] [CrossRef]
- Murphy, E.A.; Davis, J.M.; Brown, A.S.; Carmichael, M.D.; Ghaffar, A.; Mayer, E.P. Oat beta-glucan effects on neutrophil respiratory burst activity following exercise. Med. Sci. Sports Exerc. 2007, 39, 639–644. [Google Scholar]
- Davis, J.M.; Murphy, E.A.; Brown, A.S.; Carmichael, M.D.; Ghaffar, A.; Mayer, E.P. Effects of moderate exercise and oat beta-glucan on innate immune function and susceptibility to respiratory infection. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R366–R372. [Google Scholar]
- McAnulty, S.R.; Nieman, D.C.; McAnulty, L.S.; Lynch, W.S.; Jin, F.; Henson, D.A. Effect of mixed flavonoids, n-3 fatty acids, and vitamin C on oxidative stress and antioxidant capacity before and after intense cycling. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 328–337. [Google Scholar]
- Davison, G.; Gleeson, M.; Phillips, S. Antioxidant supplementation and immunoendocrine responses to prolonged exercise. Med. Sci. Sports Exerc. 2007, 39, 645–652. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gunzer, W.; Konrad, M.; Pail, E. Exercise-Induced Immunodepression in Endurance Athletes and Nutritional Intervention with Carbohydrate, Protein and Fat — What Is Possible, What Is Not? Nutrients 2012, 4, 1187-1212. https://doi.org/10.3390/nu4091187
Gunzer W, Konrad M, Pail E. Exercise-Induced Immunodepression in Endurance Athletes and Nutritional Intervention with Carbohydrate, Protein and Fat — What Is Possible, What Is Not? Nutrients. 2012; 4(9):1187-1212. https://doi.org/10.3390/nu4091187
Chicago/Turabian StyleGunzer, Wolfgang, Manuela Konrad, and Elisabeth Pail. 2012. "Exercise-Induced Immunodepression in Endurance Athletes and Nutritional Intervention with Carbohydrate, Protein and Fat — What Is Possible, What Is Not?" Nutrients 4, no. 9: 1187-1212. https://doi.org/10.3390/nu4091187
APA StyleGunzer, W., Konrad, M., & Pail, E. (2012). Exercise-Induced Immunodepression in Endurance Athletes and Nutritional Intervention with Carbohydrate, Protein and Fat — What Is Possible, What Is Not? Nutrients, 4(9), 1187-1212. https://doi.org/10.3390/nu4091187