Targeting Cell Signaling and Apoptotic Pathways by Luteolin: Cardioprotective Role in Rat Cardiomyocytes Following Ischemia/Reperfusion

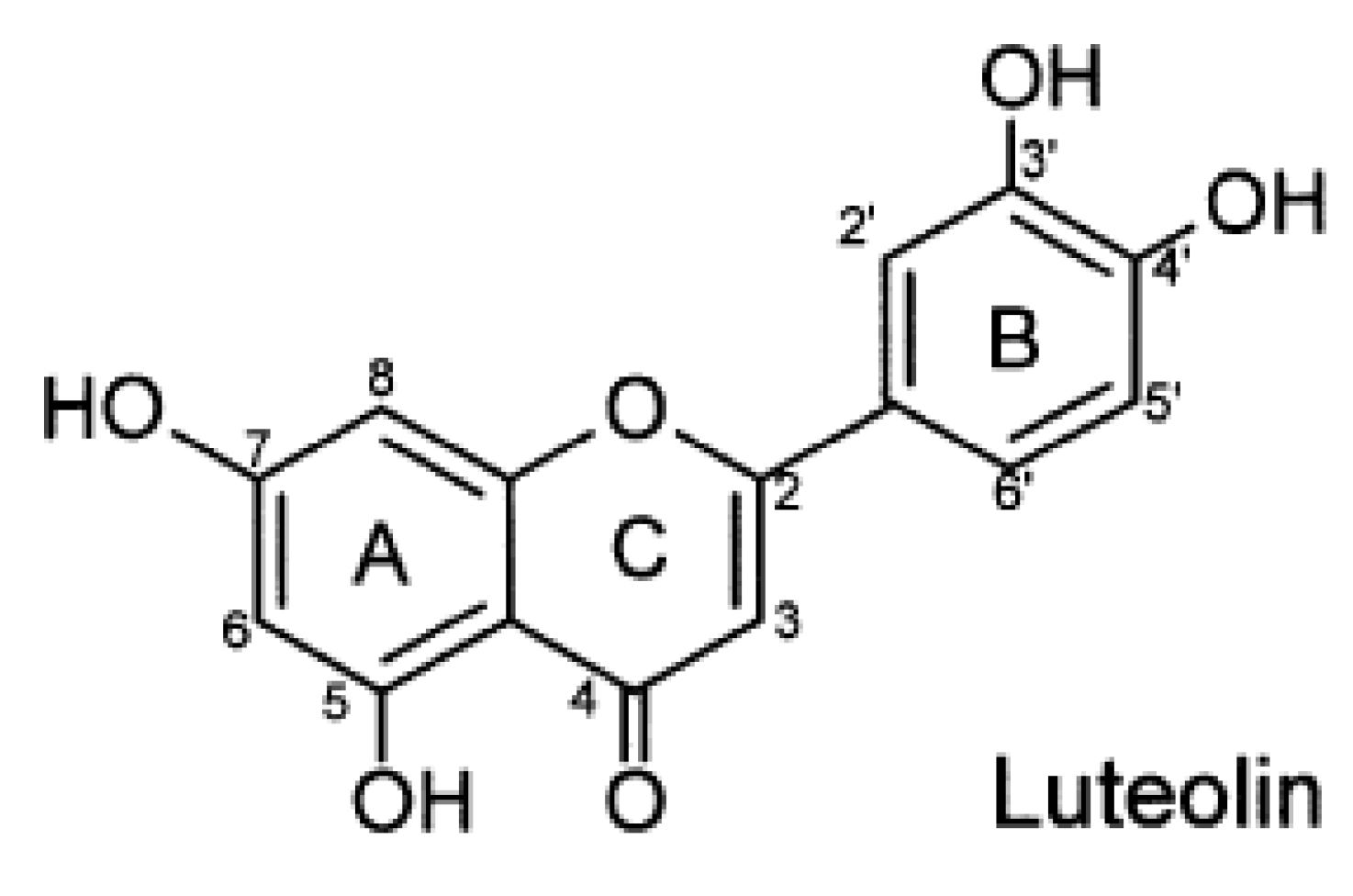{kind=link}
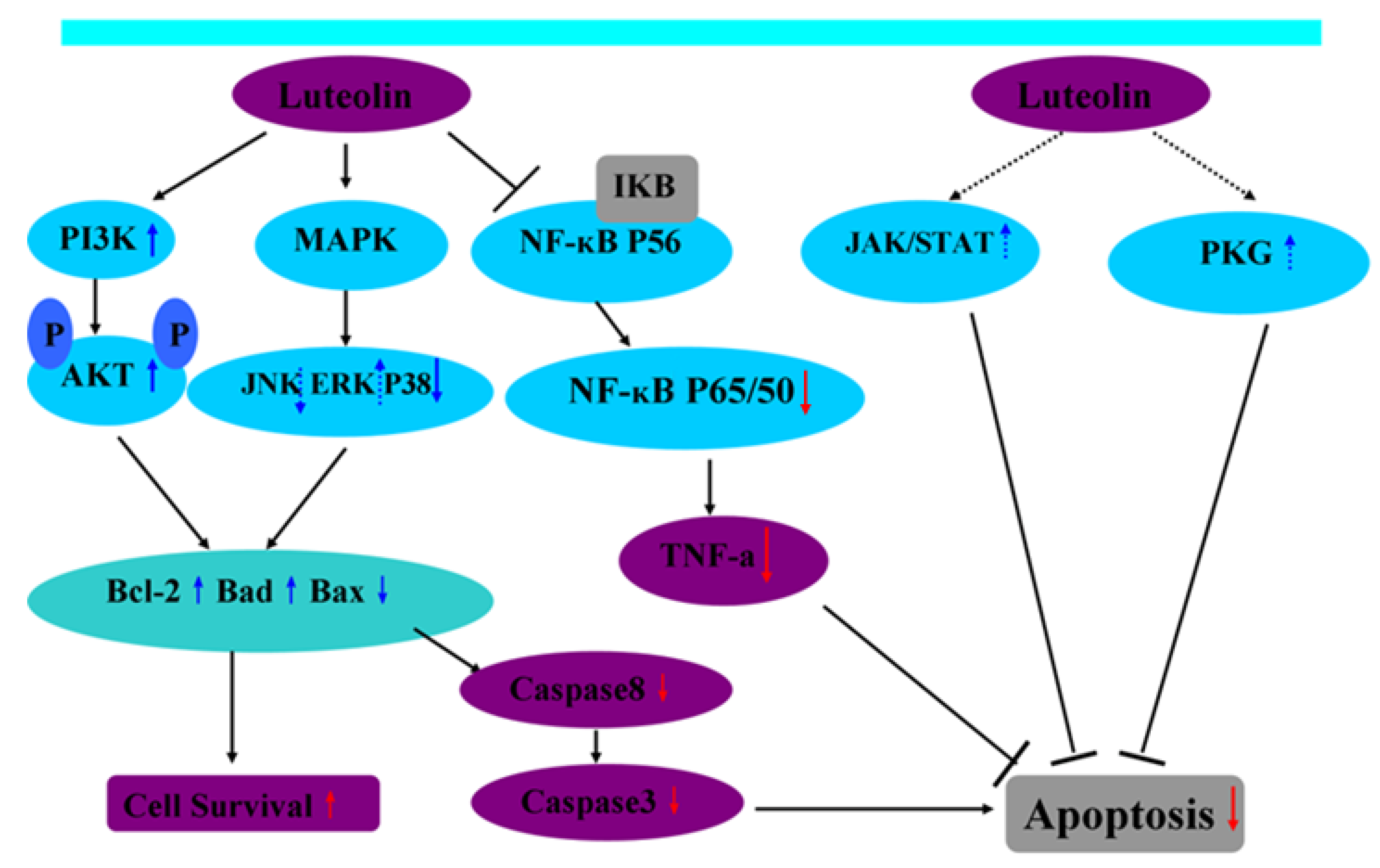{kind=link}
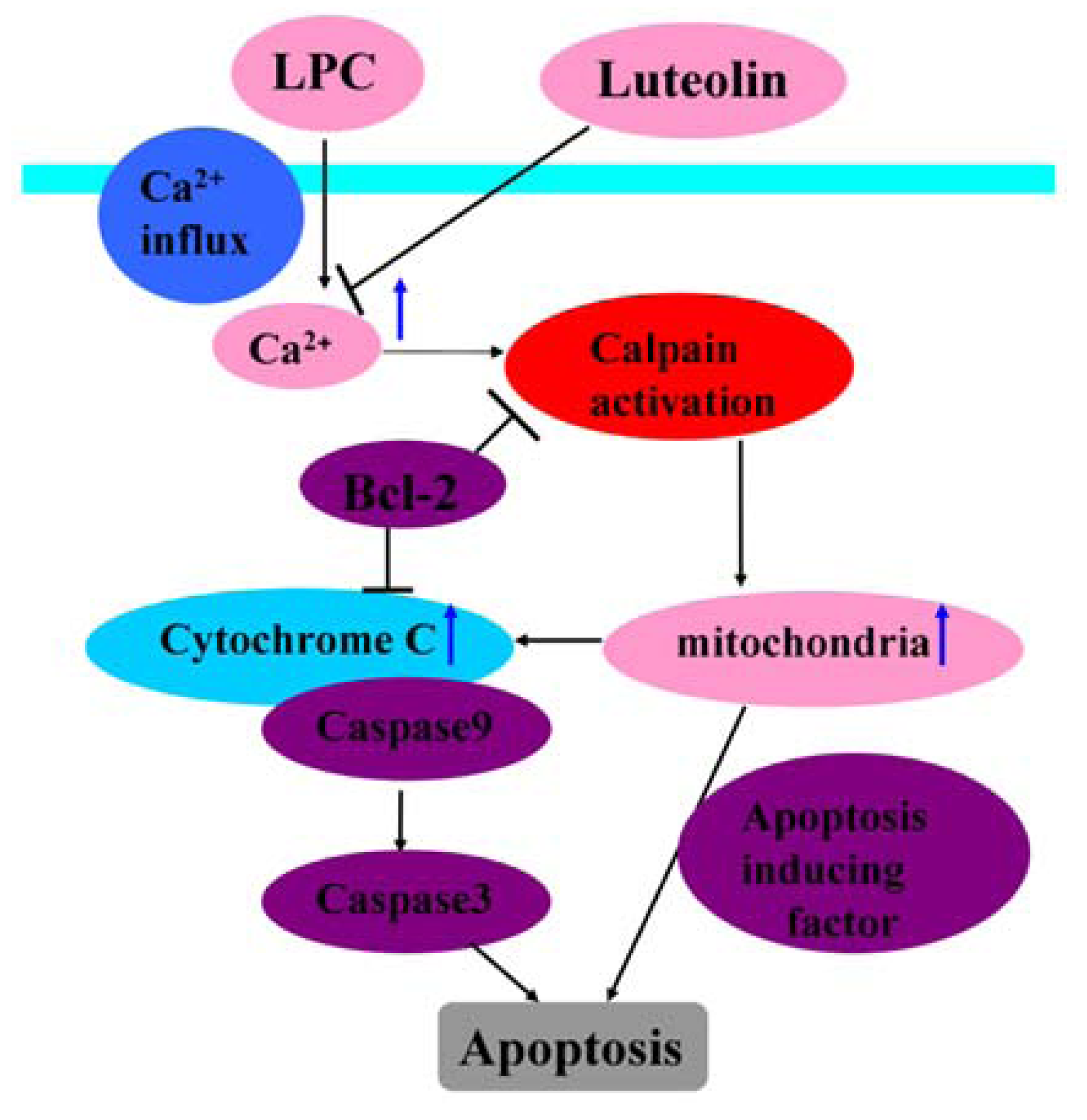{kind=link}
Abstract
:1. Introduction
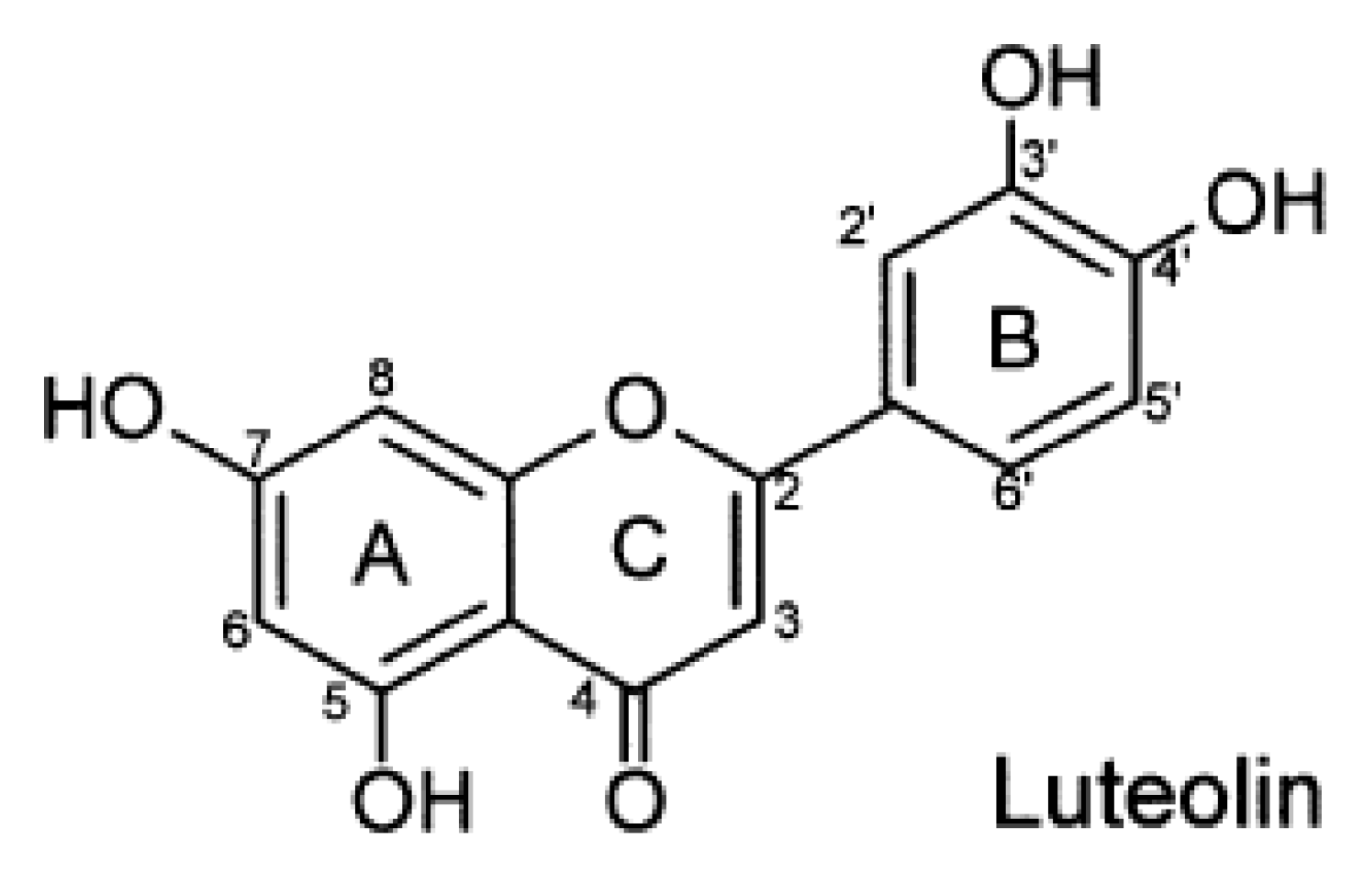
2. Mechanisms of Cell Apoptosis in Myocardial IRI
3. Luteolin and Signaling Pathways Involved Cell Apoptosis during I/R
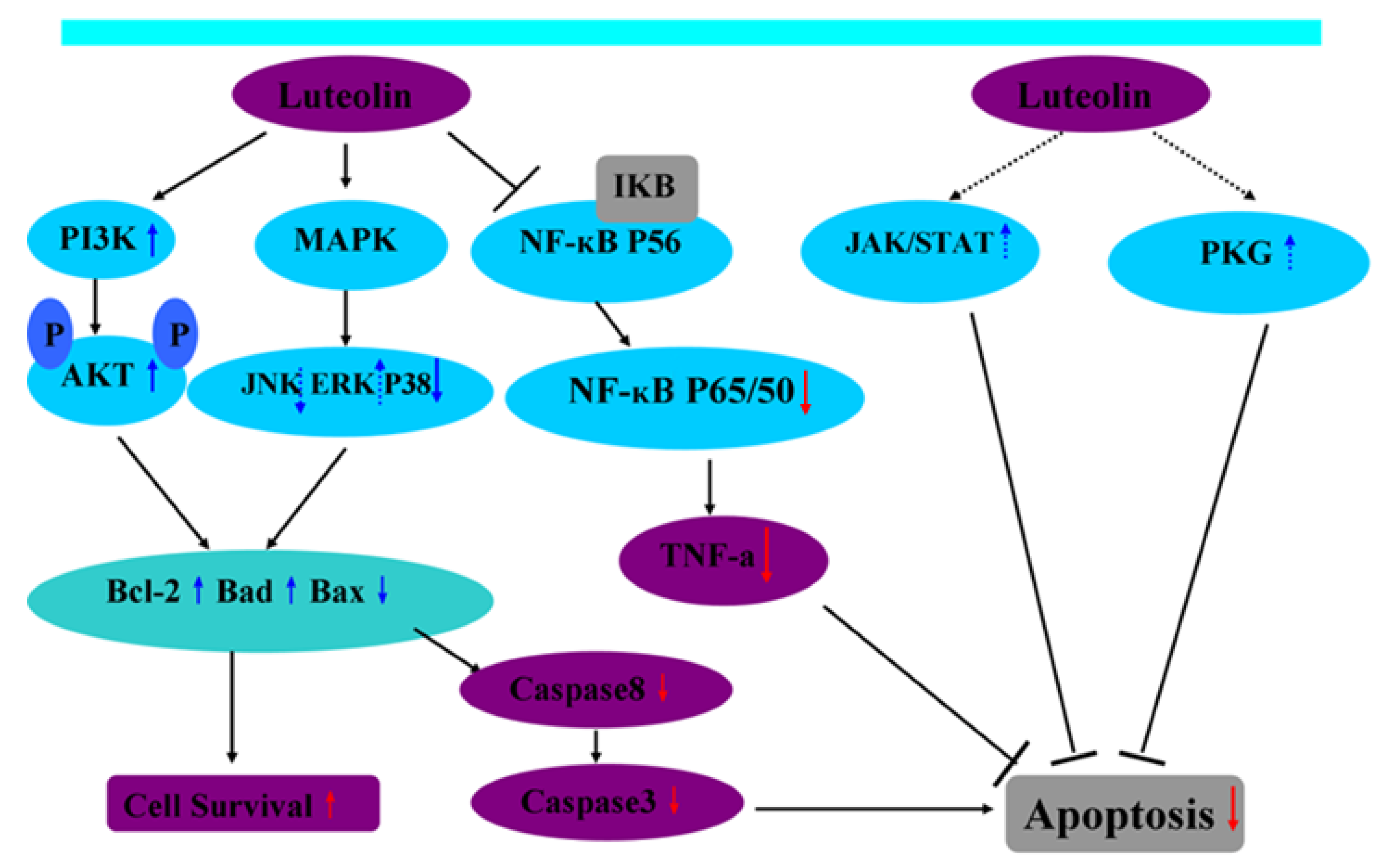
3.1. Luteolin and the PI3K/Akt Signaling Pathway Mediating Cell Apoptosis during I/R
3.2. Luteolin and the MAPKs Signaling Pathway Mediating Cell Apoptosis during I/R
3.3. Luteolin and the Caspase Signaling Pathway Mediating Cell Apoptosis during I/R

3.4. Luteolin and the JAK/STAT Signaling Pathway Mediating Cell Apoptosis during I/R
3.5. Luteolin and the cGMP/PKG Signaling Pathway Mediating Cell Apoptosis during I/R
3.6. Luteolin and the LOX-1 Signaling Pathway Mediating Cell Apoptosis during I/R
3.7. Luteolin and Other Signaling Pathway Mediating Cell Apoptosis during I/R
4. Conclusions and Perspectives
Conflict of Interest
References
- Minamino, T. Cardioprotection from ischemia/reperfusion injury: Basic and translational research. Circ. J. 2012, 76, 1074–1082. [Google Scholar] [CrossRef]
- Moens, A.L.; Claeys, M.J.; Timmermans, J.P.; Vrints, C.J. Myocardial ischemia/reperfusion-injury, a clinical view on a complex pathophysiological process. Int. J. Cardiol. 2005, 100, 179–190. [Google Scholar] [CrossRef]
- Crozier, S.J.; Zhang, X.; Wang, J.; Cheung, J.; Kimball, S.R.; Jefferson, L.S. Activation of signaling pathways and regulatory mechanisms of mRNA translation following myocardial ischemia-reperfusion. J. Appl. Physiol. 2006, 101, 576–582. [Google Scholar] [CrossRef]
- Tian, Z.; Zheng, H.; Li, J.; Li, Y.; Su, H.; Wang, X. Genetically induced moderate inhibition of the proteasome in cardiomyocytes exacerbates myocardial ischemia-reperfusion injury in mice. Circ. Res. 2012, 111, 532–542. [Google Scholar] [CrossRef]
- Fukuyama, N.; Tanaka, E.; Tabata, Y.; Fujikura, H.; Hagihara, M.; Sakamoto, H.; Ando, K.; Nakazawa, H.; Mori, H. Intravenous injection of phagocytes transfected ex vivo with FGF4 DNA/biodegradable gelatin complex promotes angiogenesis in a rat myocardial ischemia/reperfusion injury model. Basic Res. Cardiol. 2007, 102, 209–216. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Eckle, T. Ischemia and reperfusion—from mechanism to translation. Nat. Med. 2011, 17, 1391–1401. [Google Scholar] [CrossRef]
- Akhlaghi, M.; Bandy, B. Preconditioning and acute effects of flavonoids in protecting cardiomyocytes from oxidative cell death. Oxid. Med. Cell. Longev. 2012, 2012, 782321. [Google Scholar] [CrossRef]
- Middleton, E., Jr.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar]
- Marniemi, J.; Alanen, E.; Impivaara, O.; Seppanen, R.; Hakala, P.; Rajala, T.; Ronnemaa, T. Dietary and serum vitamins and minerals as predictors of myocardial infarction and stroke in elderly subjects. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 188–197. [Google Scholar] [CrossRef]
- Lopez-Lazaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.M. Luteolin, a flavonoid with potential for cancer prevention and therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef]
- Mulvihill, E.E.; Huff, M.W. Antiatherogenic properties of flavonoids: Implications for cardiovascular health. Can. J. Cardiol. 2010, 26, 17A–21A. [Google Scholar] [CrossRef]
- Cai, X.; Lu, W.; Ye, T.; Lu, M.; Wang, J.; Huo, J.; Qian, S.; Wang, X.; Cao, P. The molecular mechanism of luteolin-induced apoptosis is potentially related to inhibition of angiogenesis in human pancreatic carcinoma cells. Oncol. Rep. 2012, 28, 1353–1361. [Google Scholar]
- Liao, P.H.; Hung, L.M.; Chen, Y.H.; Kuan, Y.H.; Zhang, F.B.; Lin, R.H.; Shih, H.C.; Tsai, S.K.; Huang, S.S. Cardioprotective effects of luteolin during ischemia-reperfusion injury in rats. Circ. J. 2011, 75, 443–450. [Google Scholar] [CrossRef]
- Akhlaghi, M.; Bandy, B. Mechanisms of flavonoid protection against myocardial ischemia-reperfusion injury. J. Mol. Cell. Cardiol. 2009, 46, 309–317. [Google Scholar] [CrossRef]
- Kim, D.S.; Kwon, D.Y.; Kim, M.S.; Kim, H.K.; Lee, Y.C.; Park, S.J.; Yoo, W.H.; Chae, S.W.; Chung, M.J.; Kim, H.R.; et al. The involvement of endoplasmic reticulum stress in flavonoid-induced protection on cardiac cell death caused by ischaemia/reperfusion. J. Pharm. Pharmacol. 2010, 62, 197–204. [Google Scholar]
- Jiang, F.; Chang, C.W.; Dusting, G.J. Cytoprotection by natural and synthetic polyphenols in the heart: Novel mechanisms and perspectives. Curr. Pharm. Des. 2010, 16, 4103–4112. [Google Scholar] [CrossRef]
- Fliss, H.; Gattinger, D. Apoptosis in ischemic and reperfused rat myocardium. Circ. Res. 1996, 79, 949–956. [Google Scholar] [CrossRef]
- Zhao, Z.Q. Oxidative stress-elicited myocardial apoptosis during reperfusion. Curr. Opin. Pharmacol. 2004, 4, 159–165. [Google Scholar] [CrossRef]
- He, D.; Ma, X.; Chen, Y.; Cai, Y.; Ru, X.; Bruce, I.C.; Xia, Q.; Shi, G.; Jin, J. Luteolin inhibits pyrogallol-induced apoptosis through the extracellular signal-regulated kinase signaling pathway. FEBS J. 2012, 279, 1834–1843. [Google Scholar]
- Mullonkal, C.J.; Toledo-Pereyra, L.H. Akt in ischemia and reperfusion. J. Investig. Surg. 2007, 20, 195–203. [Google Scholar] [CrossRef]
- Matsui, T.; Tao, J.; del Monte, F.; Lee, K.H.; Li, L.; Picard, M.; Force, T.L.; Franke, T.F.; Hajjar, R.J.; Rosenzweig, A. Akt activation preserves cardiac function and prevents injury after transient cardiac ischemia in vivo. Circulation 2001, 104, 330–335. [Google Scholar]
- Li, C.; Tian, J.; Li, G.; Jiang, W.; Xing, Y.; Hou, J.; Zhu, H.; Xu, H.; Zhang, G.; Liu, Z.; et al. Asperosaponin VI protects cardiac myocytes from hypoxia-induced apoptosis via activation of the PI3K/Akt and CREB pathways. Eur. J. Pharmacol. 2010, 649, 100–107. [Google Scholar] [CrossRef]
- Fang, F.; Li, D.; Pan, H.; Chen, D.; Qi, L.; Zhang, R.; Sun, H. Luteolin inhibits apoptosis and improves cardiomyocyte contractile function through the PI3K/Akt pathway in simulated ischemia/reperfusion. Pharmacology 2011, 88, 149–158. [Google Scholar] [CrossRef]
- Sun, D.; Huang, J.; Zhang, Z.; Gao, H.; Li, J.; Shen, M.; Cao, F.; Wang, H. Luteolin limits infarct size and improves cardiac function after myocardium ischemia/reperfusion injury in diabetic rats. PLoS One 2012, 7, e33491. [Google Scholar]
- Qi, L.; Pan, H.; Li, D.; Fang, F.; Chen, D.; Sun, H. Luteolin improves contractile function and attenuates apoptosis following ischemia-reperfusion in adult rat cardiomyocytes. Eur. J. Pharmacol. 2011, 668, 201–207. [Google Scholar] [CrossRef]
- Ferrandi, C.; Ballerio, R.; Gaillard, P.; Giachetti, C.; Carboni, S.; Vitte, P.A.; Gotteland, J.P.; Cirillo, R. Inhibition of c-Jun N-terminal kinase decreases cardiomyocyte apoptosis and infarct size after myocardial ischemia and reperfusion in an aesthetized rats. Br. J. Pharmacol. 2004, 142, 953–960. [Google Scholar] [CrossRef]
- Strohm, C.; Barancik, T.; Bruhl, M.L.; Kilian, S.A.; Schaper, W. Inhibition of the ER-kinase cascade by PD98059 and UO126 counteracts ischemic preconditioning in pig myocardium. J. Cardiovasc. Pharmacol. 2000, 36, 218–229. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Hsieh, M.T.; Tsai, F.S.; Wu, C.R.; Chiu, C.S.; Lee, M.M.; Xu, H.X.; Zhao, Z.Z.; Peng, W.H. Neuroprotective effect of luteolin on amyloid beta protein (25-35)-induced toxicity in cultured rat cortical neurons. Phytother. Res. 2010, 24, S102–S108. [Google Scholar] [CrossRef]
- Lee, Y.; Gustafsson, A.B. Role of apoptosis in cardiovascular disease. Apoptosis 2009, 14, 536–548. [Google Scholar] [CrossRef]
- Maenpaa, C.J.; Shames, B.D.; van Why, S.K.; Johnson, C.P.; Nilakantan, V. Oxidant-mediated apoptosis in proximal tubular epithelial cells following ATP depletion and recovery. Free Radic. Biol. Med. 2008, 44, 518–526. [Google Scholar] [CrossRef]
- Nam, Y.J.; Mani, K.; Ashton, A.W.; Peng, C.F.; Krishnamurthy, B.; Hayakawa, Y.; Lee, P.; Korsmeyer, S.J.; Kitsis, R.N. Inhibition of both the extrinsic and intrinsic death pathways through nonhomotypic death-fold interactions. Mol. Cell 2004, 15, 901–912. [Google Scholar] [CrossRef]
- Song, J.; Liu, K.; Yi, J.; Zhu, D.; Liu, G.; Liu, B. Luteolin inhibits lysophosphatidylcholine-induced apoptosis in endothelial cells by a calcium/mitocondrion/caspases-dependent pathway. Planta Med. 2010, 76, 433–438. [Google Scholar] [CrossRef]
- Negoro, S.; Kunisada, K.; Tone, E.; Funamoto, M.; Oh, H.; Kishimoto, T.; Yamauchi-Takihara, K. Activation of JAK/STAT pathway transduces cytoprotective signal in rat acute myocardial infarction. Cardiovasc. Res. 2000, 47, 797–805. [Google Scholar] [CrossRef]
- Boengler, K.; Hilfiker-Kleiner, D.; Drexler, H.; Heusch, G.; Schulz, R. The myocardial JAK/STAT pathway: From protection to failure. Pharmacol. Ther. 2008, 120, 172–185. [Google Scholar] [CrossRef]
- Wagner, M.; Siddiqui, M.A. Signaling networks regulating cardiac myocyte survival and death. Curr. Opin. Investig. Drugs 2009, 10, 928–937. [Google Scholar]
- Burley, D.S.; Ferdinandy, P.; Baxter, G.F. Cyclic GMP and protein kinase-G in myocardial ischaemia-reperfusion: Opportunities and obstacles for survival signaling. Br. J. Pharmacol. 2007, 152, 855–869. [Google Scholar] [CrossRef]
- Han, J.; Kim, N.; Joo, H.; Kim, E.; Earm, Y.E. ATP-sensitive K(+) channel activation by nitric oxide and protein kinase G in rabbit ventricular myocytes. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H1545–H1554. [Google Scholar]
- Hu, C.; Chen, J.; Dandapat, A.; Fujita, Y.; Inoue, N.; Kawase, Y.; Jishage, K.; Suzuki, H.; Li, D.; Hermonat, P.L.; et al. LOX-1 abrogation reduces myocardial ischemia-reperfusion injury in mice. J. Mol. Cell. Cardiol. 2008, 44, 76–83. [Google Scholar] [CrossRef]
- Kataoka, K.; Hasegawa, K.; Sawamura, T.; Fujita, M.; Yanazume, T.; Iwai-Kanai, E.; Kawamura, T.; Hirai, T.; Kita, T.; Nohara, R. LOX-1 pathway affects the extent of myocardial ischemia-reperfusion injury. Biochem. Biophys. Res. Commun. 2003, 300, 656–660. [Google Scholar] [CrossRef]
- Lv, L.; Zhang, Y.; Kong, Q. Luteolin prevents LPS-induced TNF-α expression in cardiac myocytes through inhibiting NF-κB signaling pathway. Inflammation 2011, 34, 620–629. [Google Scholar] [CrossRef]
- Miura, T.; Liu, Y.; Goto, M.; Tsuchida, A.; Miki, T.; Nakano, A.; Nishino, Y.; Ohnuma, Y.; Shimamoto, K. Mitochondrial ATP-sensitive K+ channels play a role in cardioprotection by Na+-H+ exchange inhibition against ischemia/reperfusion injury. J. Am. Coll. Cardiol. 2001, 37, 957–963. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xu, T.; Li, D.; Jiang, D. Targeting Cell Signaling and Apoptotic Pathways by Luteolin: Cardioprotective Role in Rat Cardiomyocytes Following Ischemia/Reperfusion. Nutrients 2012, 4, 2008-2019. https://doi.org/10.3390/nu4122008
Xu T, Li D, Jiang D. Targeting Cell Signaling and Apoptotic Pathways by Luteolin: Cardioprotective Role in Rat Cardiomyocytes Following Ischemia/Reperfusion. Nutrients. 2012; 4(12):2008-2019. https://doi.org/10.3390/nu4122008
Chicago/Turabian StyleXu, Tongda, Dongye Li, and Dehua Jiang. 2012. "Targeting Cell Signaling and Apoptotic Pathways by Luteolin: Cardioprotective Role in Rat Cardiomyocytes Following Ischemia/Reperfusion" Nutrients 4, no. 12: 2008-2019. https://doi.org/10.3390/nu4122008