Lipophilic Micronutrients and Adipose Tissue Biology


{kind=link}
{kind=link}
{kind=link}
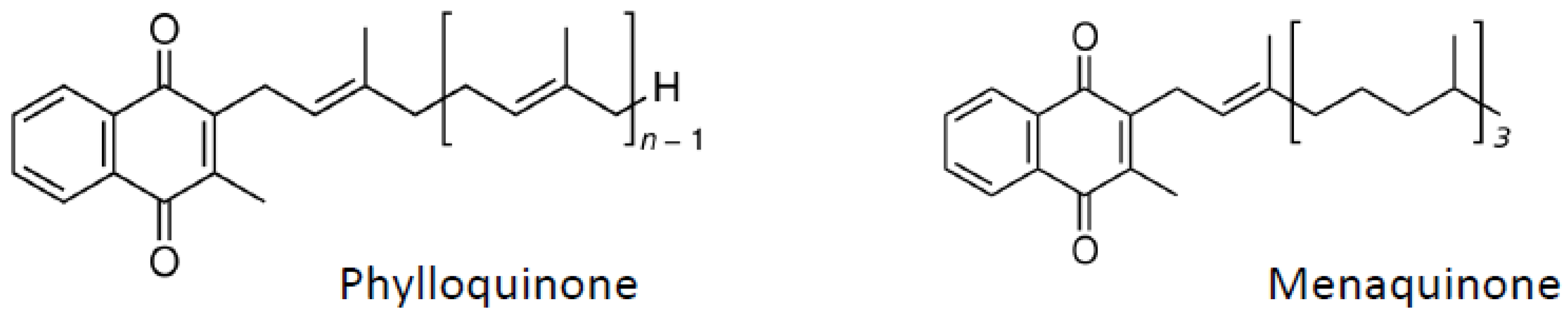{kind=link}
{kind=link}
Abstract
:1. Adipose Tissue, an Endocrine Tissue Involved in Obesity and Associated Pathologies
- Free fatty acids: continuously released from adipose tissue, with a peak in secretion during fasting and a decrease during postprandial periods. During obesity, when resistance of adipose tissue to insulin develops (partly because of hypoxia generated following adipocyte hypertrophia/hyperplasia), enhanced lipolysis leads to a massive increase in plasma free fatty acids. Free fatty acids will then perturb liver and muscle insulin action.
- Adiponectin: mainly synthesized by adipocytes [7,8]; elevated circulating concentrations are found in the plasma of lean individuals (5–30 mg/L). Conversely to other adipokines, its production and secretion are diminished in insulin resistant or Type 2 diabetic obese individuals. Adiponectin increases insulin sensitivity and modulates hepatic glucose synthesis by inhibiting the expression of enzymes essential to gluconeogenesis. Adiponectin also presents anti-inflammatory properties through its ability to modulate the expression of pro- and anti-inflammatory cytokines (especially TNF-α).
- Leptin: similarly to adiponectin, leptin is mainly produced by adipocytes [9]. Circulating levels of leptin as well as its expression in adipose tissue are positively correlated with the severity of obesity [10]. Leptin’s main action is on the central nervous system to regulate food intake. Furthermore, leptin possesses pro-inflammatory properties: It stimulates the production of TNF-α, IL-6 and IL-12 by macrophages. Conversely, it also improves insulin sensitivity by activating AMP-activated protein kinase (AMPK). However, obese individuals display very high circulating levels of leptin, suggesting the existence of leptin resistance in these subjects [10].
- Pro-inflammatory cytokines: TNF-α, IL-6 and IL-1β are synthesized by numerous tissues, including obese adipose tissue, which, in addition to their well described pro-inflammatory properties, are involved in the genesis of insulin resistance. TNF-α was the first pro-inflammatory mediator linked to inflammation, obesity and insulin resistance [11]. TNF-α and IL-1β interfere with insulin signaling at the level of insulin receptor substrate-1 (IRS-1) in different ways, whereas the mechanisms of action of IL-6 are still a matter of debate [10,12].
- Plasma retinol binding protein 4 (RBP4), produced mainly by the liver, has also been shown to be an adipokine, favoring insulin resistance in mice [13]. However, clinical observations did not show a consistent correlation between plasma RBP4 and obesity associated insulin resistance, but this lack of a correlation may be caused by confounding factors, including methodology [14]. Several authors have therefore proposed to use the retinol:RBP4 ratio rather than RBP4 levels alone as a better marker (a low retinol:RBP4 is associated with insulin resistance), suggesting that retinol-free RBP4 (apo-RBP4) might contribute more to insulin resistance than retinol-bound RBP4 (holo-RBP4) [15,16]. Recently, two mechanisms by which RBP4 induces insulin resistance have been proposed, suggesting that both apo- and holo-RBP4 could participate in tissue insulin resistance. In a first report, it was shown that binding of holo-RBP4 to its membrane receptor, sensitive to retinoic acid-6 (STRA6, mediating cellular uptake of retinol), triggers an inflammatory signaling cascade (janus activated kinase 2/signal transducer and activator of transcription 5, JAK2/STAT5), causing a blockade of insulin response via the induction of expression of suppressor of cytokine signaling-3 (SOCS3), but also an increase in PPARγ, which stimulates lipid accumulation [17]. On the other hand, Norseen et al. have shown that insulin resistance could be triggered indirectly in adipocytes by either apo- and holo-RBP4 [18]. Indeed, both forms of RBP4 could stimulate pro-inflammatory cytokine secretion by macrophages via JNK and TLR4 pathways, independently of STRA6 binding, hereby perturbating insulin pathways in adipocytes.
2. Impact of LM on Obesity and Associated Pathologies
3. Vitamin A
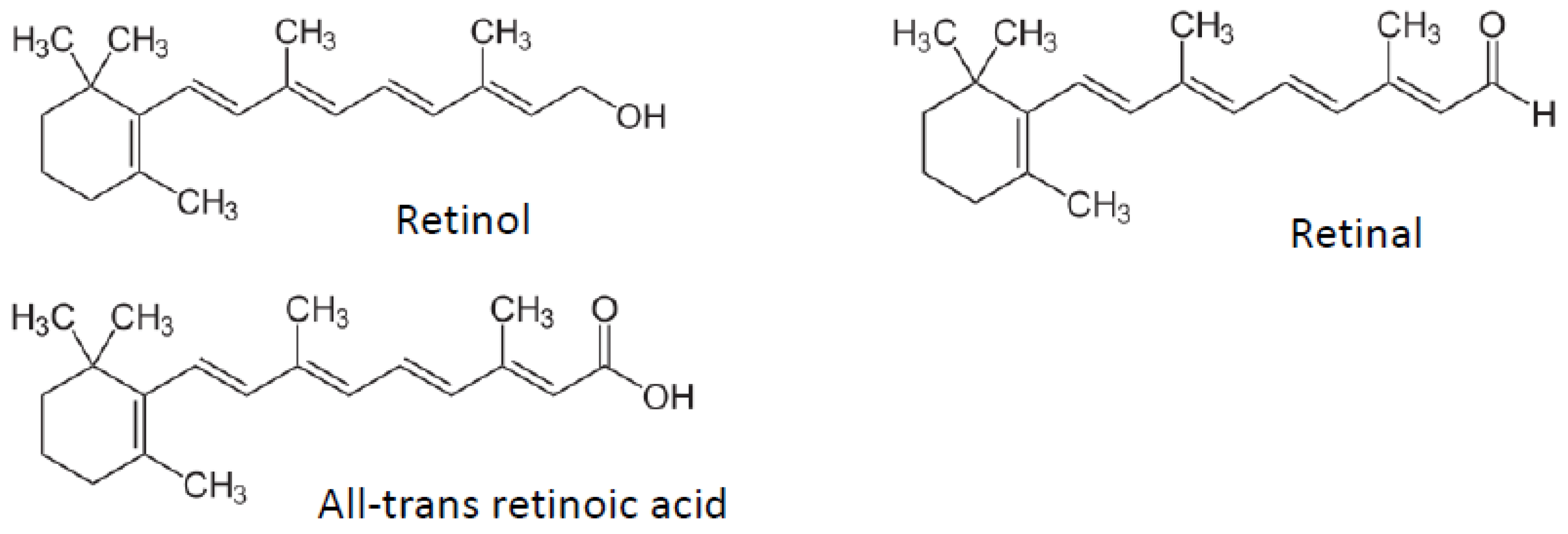
3.1. Adipogenesis
3.2. Inflammation
3.3. Metabolism
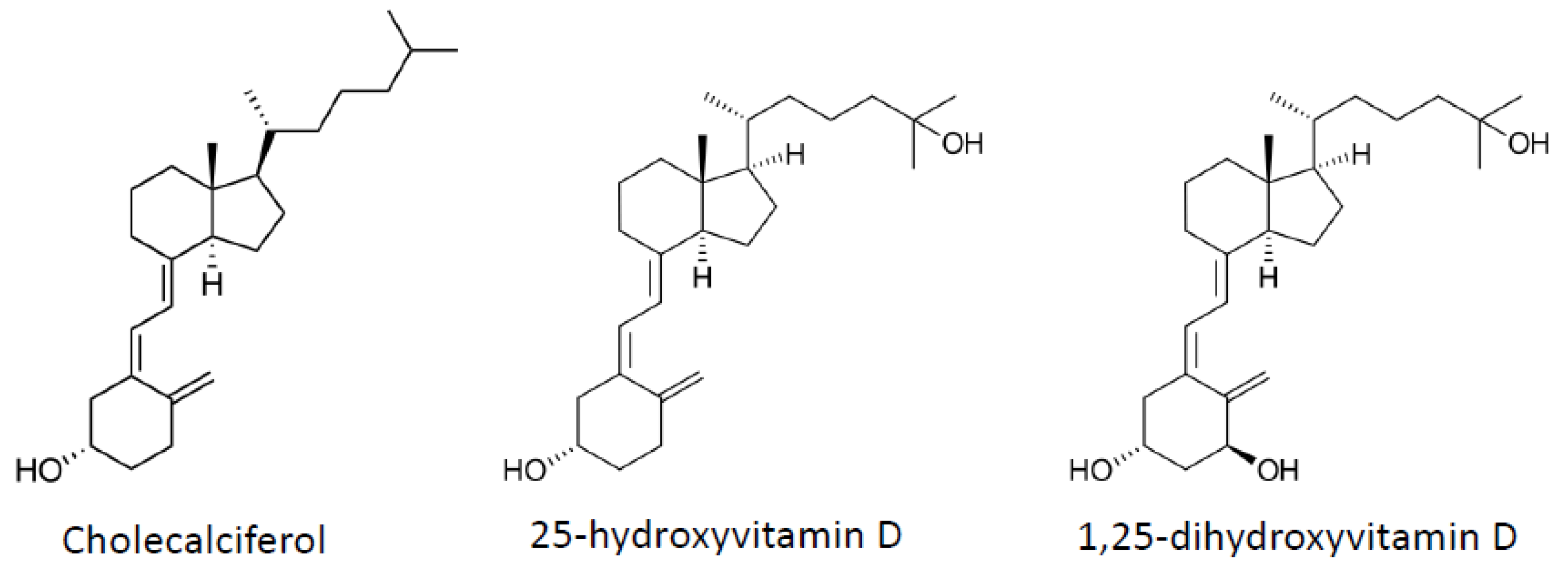
4. Vitamin D
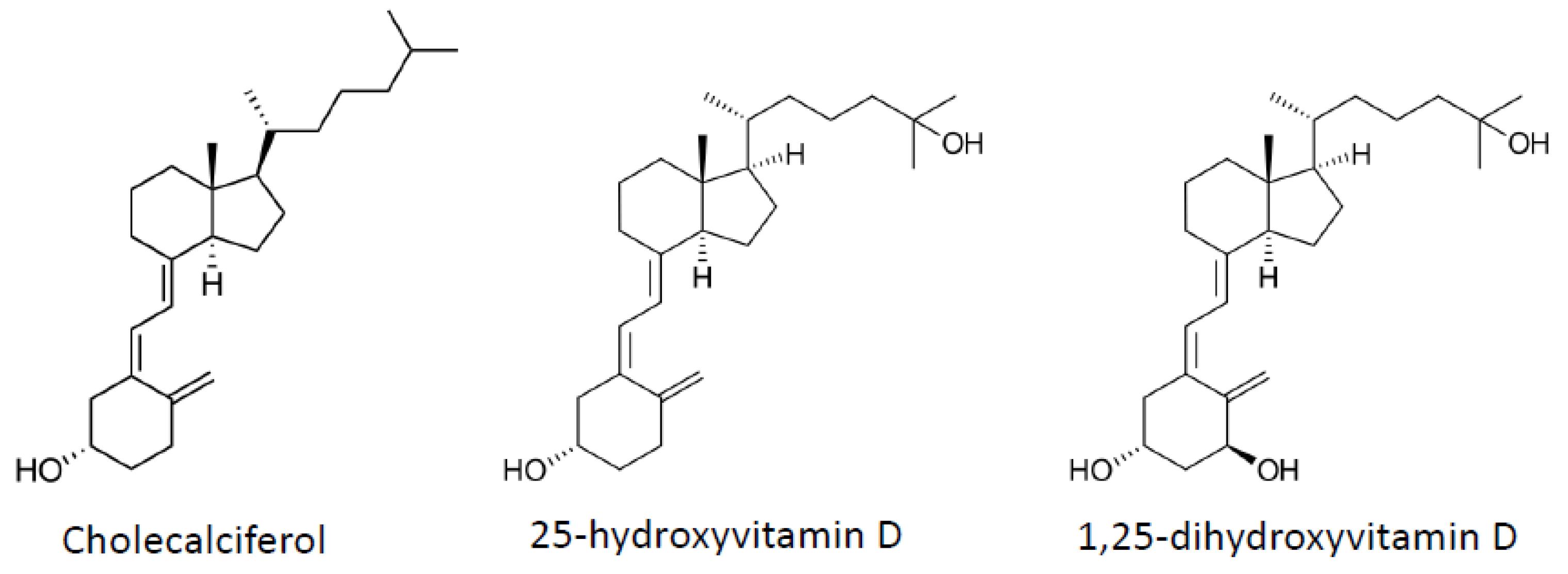
4.1. Adipogenesis
4.2. Inflammation
4.3. Metabolism
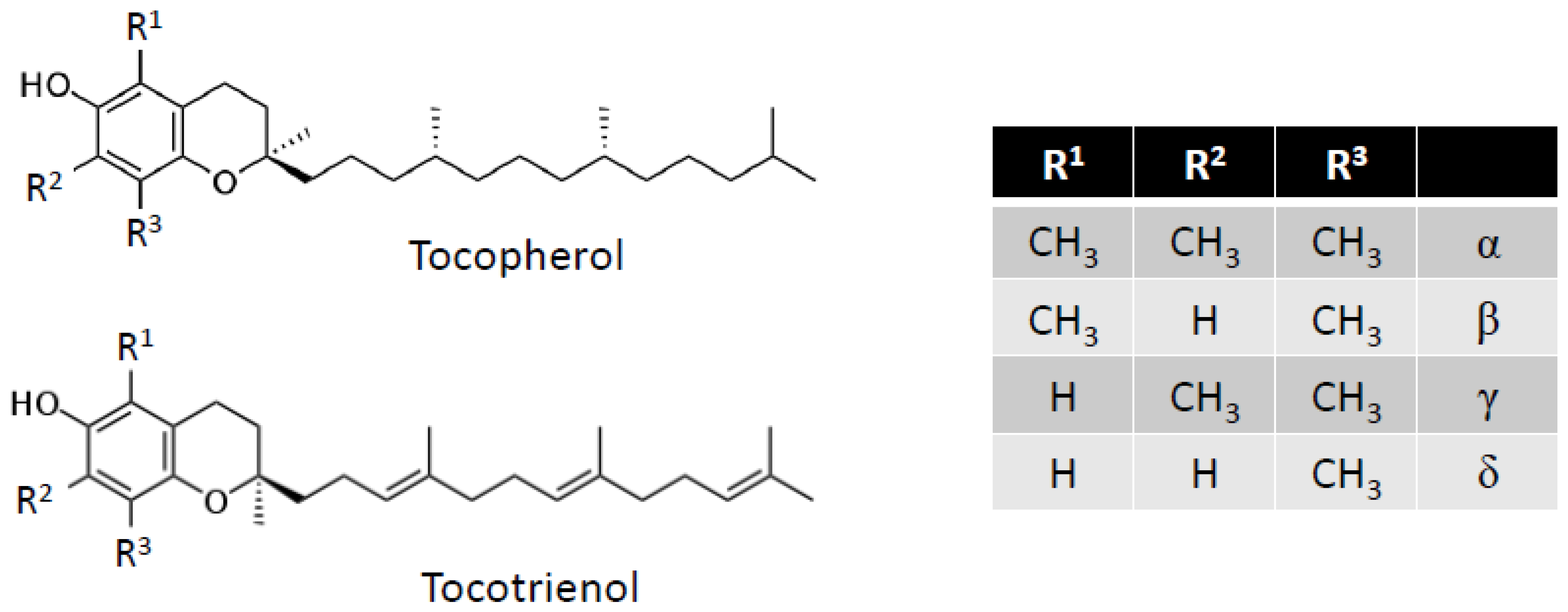
5. Vitamin E
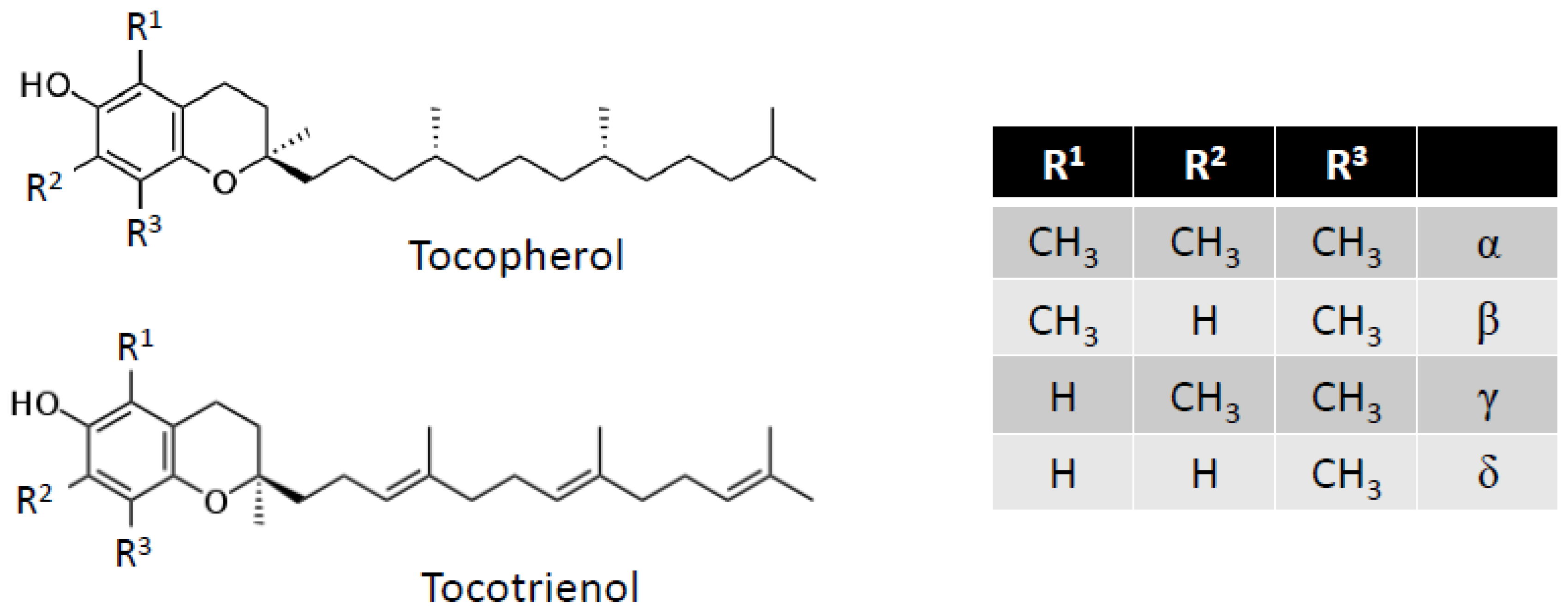
5.1. Adipogenesis
5.2. Inflammation
5.3. Metabolism
6. Vitamin K
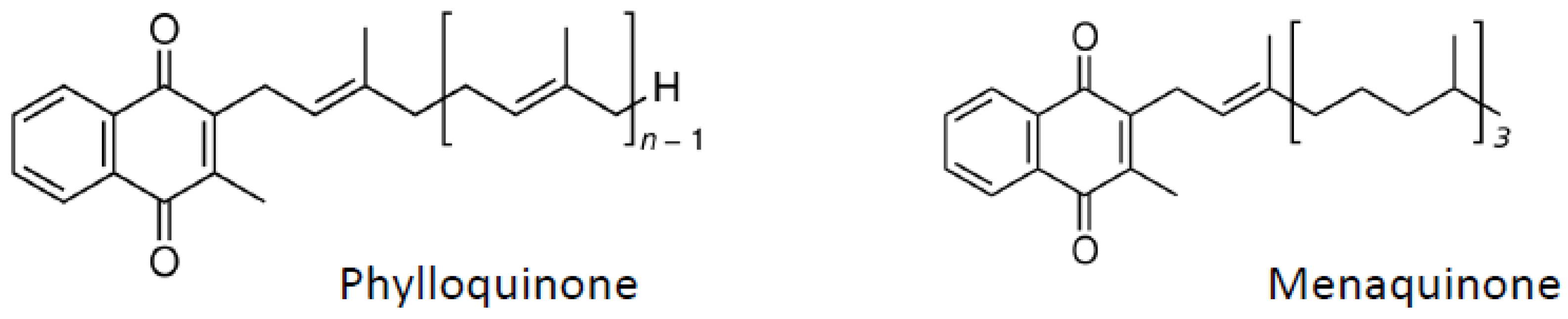
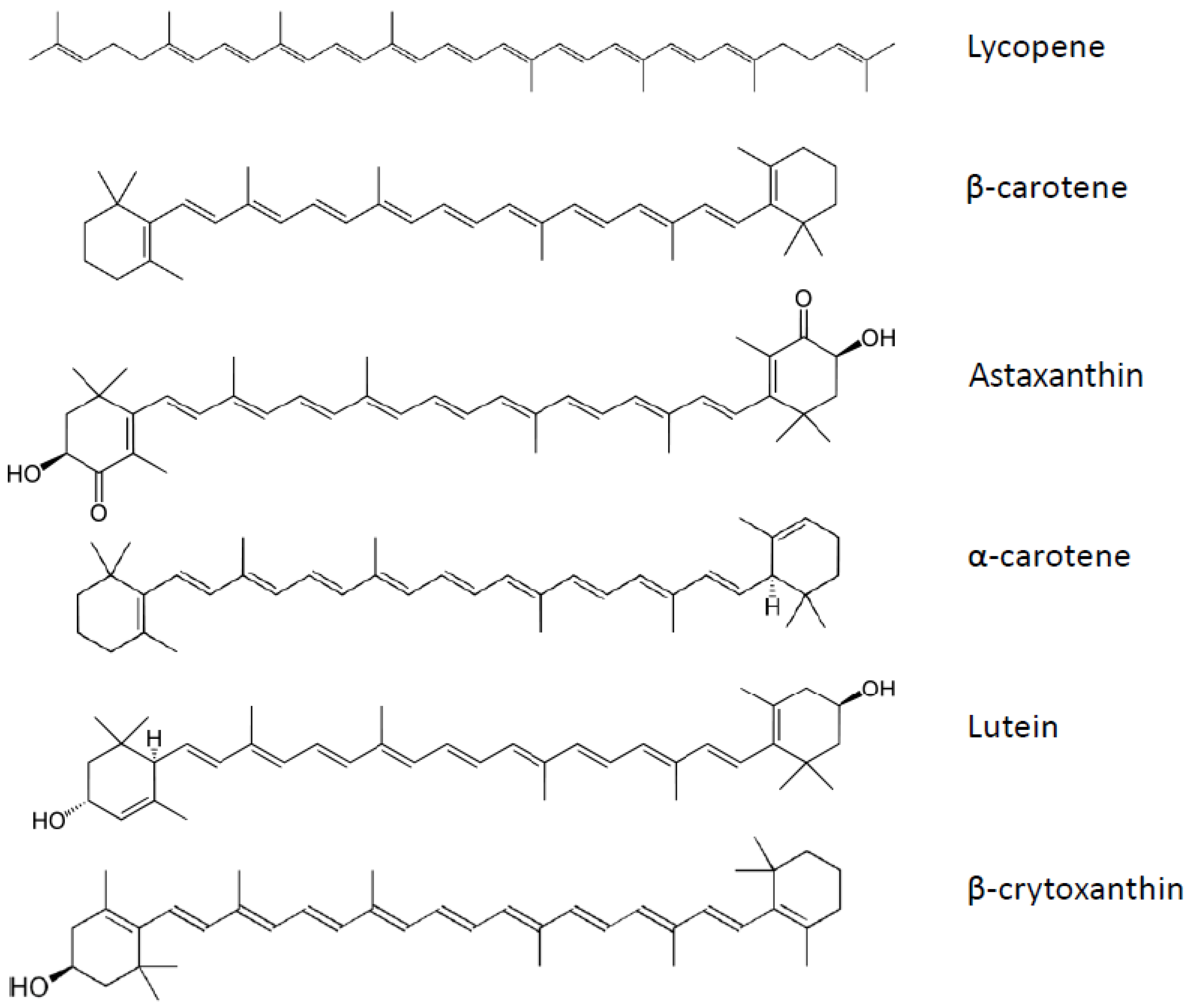
7. Carotenoids
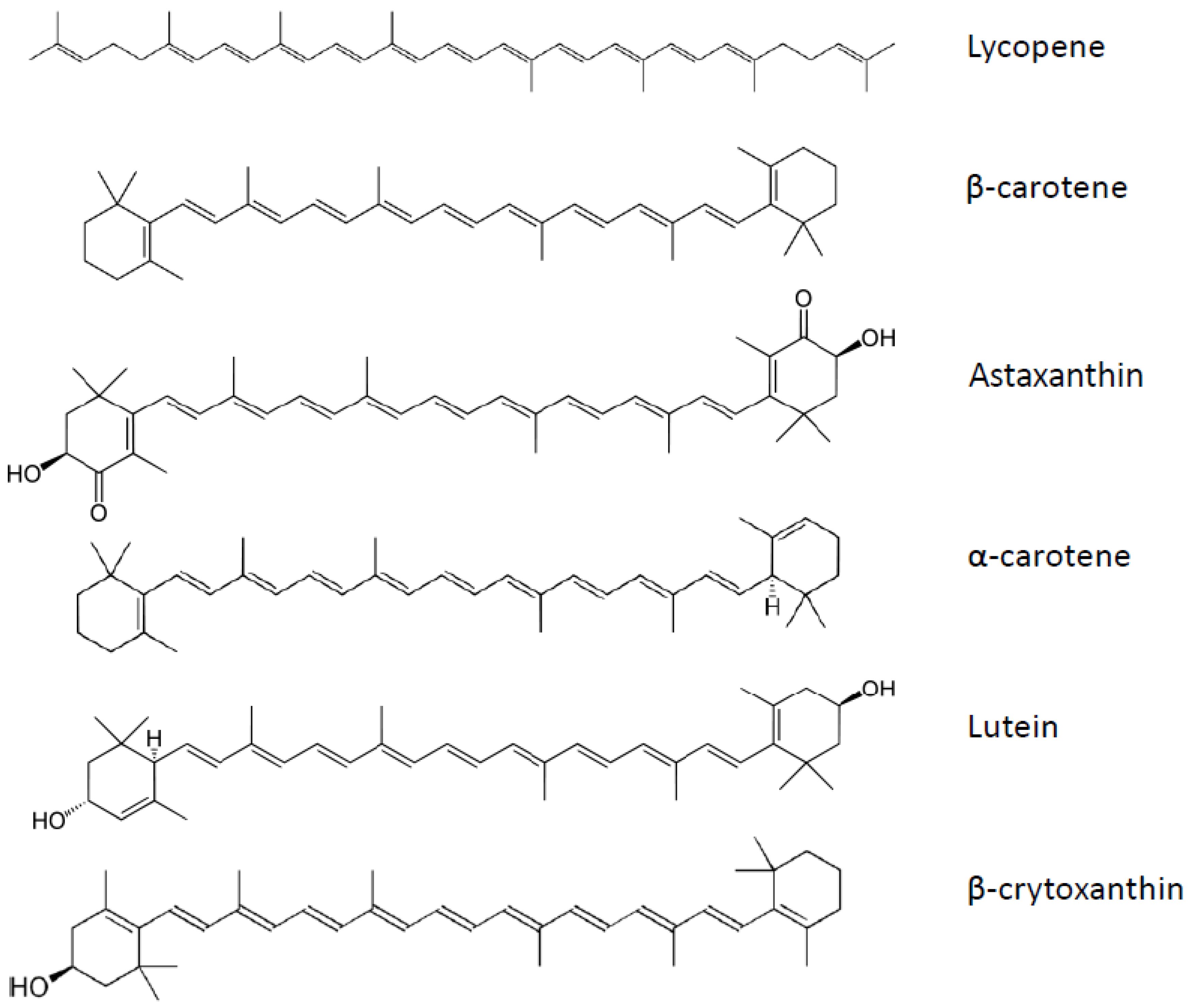
7.1. Adipogenesis
7.2. Inflammation
7.3. Metabolism
8. Conclusion
References
- Farmer, S.R. Transcriptional control of adipocyte formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef]
- Arner, P.; Spalding, K.L. Fat cell turnover in humans. Biochem. Biophys. Res. Commun. 2010, 396, 101–104. [Google Scholar] [CrossRef]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef]
- Olefsky, J.M.; Glass, C.K. Macrophages, inflammation, and insulin resistance. Annu. Rev. Physiol. 2010, 72, 219–246. [Google Scholar] [CrossRef]
- Bourlier, V.; Bouloumie, A. Role of macrophage tissue infiltration in obesity and insulin resistance. Diabetes Metab. 2009, 35, 251–260. [Google Scholar] [CrossRef]
- Cornier, M.A.; Dabelea, D.; Hernandez, T.L.; Lindstrom, R.C.; Steig, A.J.; Stob, N.R.; van Pelt, R.E.; Wang, H.; Eckel, R.H. The metabolic syndrome. Endocr. Rev. 2008, 29, 777–822. [Google Scholar] [CrossRef]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef]
- Lara-Castro, C.; Fu, Y.; Chung, B.H.; Garvey, W.T. Adiponectin and the metabolic syndrome: Mechanisms mediating risk for metabolic and cardiovascular disease. Curr. Opin. Lipidol. 2007, 18, 263–270. [Google Scholar] [CrossRef]
- Ahima, R.S.; Flier, J.S. Leptin. Annu. Rev. Physiol. 2000, 62, 413–437. [Google Scholar] [CrossRef]
- Antuna-Puente, B.; Feve, B.; Fellahi, S.; Bastard, J.P. Adipokines: The missing link between insulin resistance and obesity. Diabetes Metab. 2008, 34, 2–11. [Google Scholar] [CrossRef]
- Uysal, K.T.; Wiesbrock, S.M.; Marino, M.W.; Hotamisligil, G.S. Protection from obesity-induced insulin resistance in mice lacking TNF-alpha function. Nature 1997, 389, 610–614. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators linking adipose tissue, inflammation and immunity. Nat. Rev. Immunol. 2006, 6, 772–783. [Google Scholar] [CrossRef]
- Yang, Q.; Graham, T.E.; Mody, N.; Preitner, F.; Peroni, O.D.; Zabolotny, J.M.; Kotani, K.; Quadro, L.; Kahn, B.B. Serum retinol binding protein 4 contributes to insulin resistance in obesity and type 2 diabetes. Nature 2005, 436, 356–362. [Google Scholar]
- Kotnik, P.; Fischer-Posovszky, P.; Wabitsch, M. RBP4: A controversial adipokine. Eur. J. Endocrinol. 2011, 165, 703–711. [Google Scholar] [CrossRef]
- Erikstrup, C.; Mortensen, O.H.; Nielsen, A.R.; Fischer, C.P.; Plomgaard, P.; Petersen, A.M.; Krogh-Madsen, R.; Lindegaard, B.; Erhardt, J.G.; Ullum, H.; et al. RBP-to-retinol ratio, but not total RBP, is elevated in patients with type 2 diabetes. Diabetes Obes. Metab. 2009, 11, 204–212. [Google Scholar] [CrossRef]
- Mills, J.P.; Furr, H.C.; Tanumihardjo, S.A. Retinol to retinol-binding protein (RBP) is low in obese adults due to elevated apo-RBP. Exp. Biol. Med. (Maywood) 2008, 233, 1255–1261. [Google Scholar] [CrossRef]
- Berry, D.C.; Jin, H.; Majumdar, A.; Noy, N. Signaling by vitamin A and retinol-binding protein regulates gene expression to inhibit insulin responses. Proc. Natl. Acad. Sci. USA 2011, 108, 4340–4345. [Google Scholar]
- Norseen, J.; Hosooka, T.; Hammarstedt, A.; Yore, M.M.; Kant, S.; Aryal, P.; Kiernan, U.A.; Phillips, D.A.; Maruyama, H.; Kraus, B.J.; et al. Retinol-binding protein 4 inhibits insulin signaling in adipocytes by inducing proinflammatory cytokines in macrophages through a c-Jun N-terminal kinase- and toll-like receptor 4-dependent and retinol-independent mechanism. Mol. Cell. Biol. 2012, 32, 2010–2019. [Google Scholar] [CrossRef]
- Meijer, K.; de Vries, M.; Al-Lahham, S.; Bruinenberg, M.; Weening, D.; Dijkstra, M.; Kloosterhuis, N.; van der Leij, R.J.; van der Want, H.; Kroesen, B.J.; et al. Human primary adipocytes exhibit immune cell function: Adipocytes prime inflammation independent of macrophages. PLoS One 2010, 6, e17154. [Google Scholar]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K.; et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Invest. 2006, 116, 1494–1505. [Google Scholar]
- Kimmons, J.E.; Blanck, H.M.; Tohill, B.C.; Zhang, J.; Khan, L.K. Associations between body mass index and the prevalence of low micronutrient levels among US adults. MedGenMed 2006, 8, 59. [Google Scholar]
- Garcia, O.P.; Long, K.Z.; Rosado, J.L. Impact of micronutrient deficiencies on obesity. Nutr. Rev. 2009, 67, 559–572. [Google Scholar] [CrossRef]
- Calder, P.C.; Ahluwalia, N.; Brouns, F.; Buetler, T.; Clement, K.; Cunningham, K.; Esposito, K.; Jonsson, L.S.; Kolb, H.; Lansink, M.; et al. Dietary factors and low-grade inflammation in relation to overweight and obesity. Br. J. Nutr. 2011, 106, S5–S78. [Google Scholar] [CrossRef]
- Pittas, A.G.; Dawson-Hughes, B. Vitamin D and diabetes. J. Steroid Biochem. Mol. Biol. 2010, 121, 425–429. [Google Scholar] [CrossRef]
- Beydoun, M.A.; Shroff, M.R.; Chen, X.; Beydoun, H.A.; Wang, Y.; Zonderman, A.B. Serum antioxidant status is associated with metabolic syndrome among U.S. adults in recent national surveys. J. Nutr. 2011, 141, 903–913. [Google Scholar] [CrossRef]
- D’Ambrosio, D.N.; Clugston, R.D.; Blaner, W.S. Vitamin A metabolism: An update. Nutrients 2011, 3, 63–103. [Google Scholar] [CrossRef]
- Tsutsumi, C.; Okuno, M.; Tannous, L.; Piantedosi, R.; Allan, M.; Goodman, D.S.; Blaner, W.S. Retinoids and retinoid-binding protein expression in rat adipocytes. J. Biol. Chem. 1992, 267, 1805–1810. [Google Scholar]
- Kane, M.A.; Folias, A.E.; Napoli, J.L. HPLC/UV quantitation of retinal, retinol, and retinyl esters in serum and tissues. Anal. Biochem. 2008, 378, 71–79. [Google Scholar] [CrossRef]
- Sima, A.; Manolescu, D.C.; Bhat, P. Retinoids and retinoid-metabolic gene expression in mouse adipose tissues. Biochem. Cell Biol. 2011, 89, 578–584. [Google Scholar] [CrossRef]
- Ziouzenkova, O.; Orasanu, G.; Sharlach, M.; Akiyama, T.E.; Berger, J.P.; Viereck, J.; Hamilton, J.A.; Tang, G.; Dolnikowski, G.G.; Vogel, S.; et al. Retinaldehyde represses adipogenesis and diet-induced obesity. Nat. Med. 2007, 13, 695–702. [Google Scholar] [CrossRef]
- Kane, M.A.; Folias, A.E.; Wang, C.; Napoli, J.L. Quantitative profiling of endogenous retinoic acid in vivo and in vitro by tandem mass spectrometry. Anal. Chem. 2008, 80, 1702–1708. [Google Scholar] [CrossRef]
- O’Byrne, S.M.; Wongsiriroj, N.; Libien, J.; Vogel, S.; Goldberg, I.J.; Baehr, W.; Palczewski, K.; Blaner, W.S. Retinoid absorption and storage is impaired in mice lacking lecithin:retinol acyltransferase (LRAT). J. Biol. Chem. 2005, 280, 35647–35657. [Google Scholar]
- Kane, M.A. Analysis, occurrence, and function of 9-cis-retinoic acid. Biochim. Biophys. Acta 2012, 1821, 10–20. [Google Scholar] [CrossRef]
- Germain, P.; Chambon, P.; Eichele, G.; Evans, R.M.; Lazar, M.A.; Leid, M.; de Lera, A.R.; Lotan, R.; Mangelsdorf, D.J.; Gronemeyer, H. International union of pharmacology. LXIII. Retinoid X receptors. Pharmacol. Rev. 2006, 58, 760–772. [Google Scholar] [CrossRef]
- Germain, P.; Chambon, P.; Eichele, G.; Evans, R.M.; Lazar, M.A.; Leid, M.; de Lera, A.R.; Lotan, R.; Mangelsdorf, D.J.; Gronemeyer, H. International union of pharmacology. LX. Retinoic acid receptors. Pharmacol. Rev. 2006, 58, 712–725. [Google Scholar] [CrossRef]
- Yasmeen, R.; Jeyakumar, S.M.; Reichert, B.; Yang, F.; Ziouzenkova, O. The contribution of vitamin A to autocrine regulation of fat depots. Biochim. Biophys. Acta 2012, 1821, 190–197. [Google Scholar]
- Haq, R.; Chytil, F. Expression of nuclear retinoic acid receptors in rat adipose tissue. Biochem. Biophys. Res. Commun. 1991, 176, 1539–1544. [Google Scholar] [CrossRef]
- Kamei, Y.; Kawada, T.; Kazuki, R.; Sugimoto, E. Retinoic acid receptor gamma 2 gene expression is up-regulated by retinoic acid in 3T3-L1 preadipocytes. Biochem. J. 1993, 293, 807–812. [Google Scholar]
- Villarroya, F.; Giralt, M.; Iglesias, R. Retinoids and adipose tissues: Metabolism, cell differentiation and gene expression. Int. J. Obes. Relat. Metab. Disord. 1999, 23, 1–6. [Google Scholar] [CrossRef]
- Safonova, I.; Darimont, C.; Amri, E.Z.; Grimaldi, P.; Ailhaud, G.; Reichert, U.; Shroot, B. Retinoids are positive effectors of adipose cell differentiation. Mol. Cell. Endocrinol. 1994, 104, 201–211. [Google Scholar] [CrossRef]
- Murray, T.; Russell, T.R. Inhibition of adipose conversion in 3T3-L2 cells by retinoic acid. J. Supramol. Struct. 1980, 14, 255–266. [Google Scholar] [CrossRef]
- Castro-Munozledo, F.; Marsch-Moreno, M.; Beltran-Langarica, A.; Kuri-Harcuch, W. Commitment of adipocyte differentiation in 3T3 cells is inhibited by retinoic acid, and the expression of lipogenic enzymes is modulated through cytoskeleton stabilization. Differentiation 1987, 36, 211–219. [Google Scholar] [CrossRef]
- Kawada, T.; Aoki, N.; Kamei, Y.; Maeshige, K.; Nishiu, S.; Sugimoto, E. Comparative investigation of vitamins and their analogues on terminal differentiation, from preadipocytes to adipocytes, of 3T3-L1 cells. Comp. Biochem. Physiol. A Comp. Physiol. 1990, 96, 323–326. [Google Scholar] [CrossRef]
- Kuri-Harcuch, W. Differentiation of 3T3-F442A cells into adipocytes is inhibited by retinoic acid. Differentiation 1982, 23, 164–169. [Google Scholar] [CrossRef]
- Pairault, J.; Lasnier, F. Control of the adipogenic differentiation of 3T3-F442A cells by retinoic acid, dexamethasone, and insulin: A topographic analysis. J. Cell. Physiol. 1987, 132, 279–286. [Google Scholar] [CrossRef]
- Pairault, J.; Quignard-Boulange, A.; Dugail, I.; Lasnier, F. Differential effects of retinoic acid upon early and late events in adipose conversion of 3T3 preadipocytes. Exp. Cell Res. 1988, 177, 27–36. [Google Scholar] [CrossRef]
- Stephens, J.M.; Butts, M.; Stone, R.; Pekala, P.H.; Bernlohr, D.A. Regulation of transcription factor MRNA accumulation during 3T3-L1 preadipocyte differentiation by antagonists of adipogenesis. Mol. Cell. Biochem. 1993, 123, 63–71. [Google Scholar] [CrossRef]
- Stone, R.L.; Bernlohr, D.A. The molecular basis for inhibition of adipose conversion of murine 3T3-L1 cells by retinoic acid. Differentiation 1990, 45, 119–127. [Google Scholar] [CrossRef]
- Suryawan, A.; Hu, C.Y. Effect of retinoic acid on differentiation of cultured pig preadipocytes. J. Anim. Sci. 1997, 75, 112–117. [Google Scholar]
- Xue, J.C.; Schwarz, E.J.; Chawla, A.; Lazar, M.A. Distinct stages in adipogenesis revealed by retinoid inhibition of differentiation after induction of PPARgamma. Mol. Cell. Biol. 1996, 16, 1567–1575. [Google Scholar]
- Schwarz, E.J.; Reginato, M.J.; Shao, D.; Krakow, S.L.; Lazar, M.A. Retinoic acid blocks adipogenesis by inhibiting C/EBPbeta-mediated transcription. Mol. Cell. Biol. 1997, 17, 1552–1561. [Google Scholar]
- Berry, D.C.; DeSantis, D.; Soltanian, H.; Croniger, C.M.; Noy, N. Retinoic acid upregulates preadipocyte genes to block adipogenesis and suppress diet-induced obesity. Diabetes 2012, 61, 1112–1121. [Google Scholar] [CrossRef]
- Kim, C.H. Retinoic acid, immunity, and inflammation. Vitam. Horm. 2011, 86, 83–101. [Google Scholar] [CrossRef]
- Antras, J.; Lasnier, F.; Pairault, J. Adipsin gene expression in 3T3-F442A adipocytes is posttranscriptionally down-regulated by retinoic acid. J. Biol. Chem. 1991, 266, 1157–1161. [Google Scholar]
- Felipe, F.; Bonet, M.L.; Ribot, J.; Palou, A. Modulation of resistin expression by retinoic acid and vitamin A status. Diabetes 2004, 53, 882–889. [Google Scholar] [CrossRef]
- Gouranton, E.; Thabuis, C.; Riollet, C.; Malezet-Desmoulins, C.; El Yazidi, C.; Amiot, M.J.; Borel, P.; Landrier, J.F. Lycopene inhibits proinflammatory cytokine and chemokine expression in adipose tissue. J. Nutr. Biochem. 2011, 22, 642–648. [Google Scholar] [CrossRef]
- Esteban-Pretel, G.; Marin, M.P.; Cabezuelo, F.; Moreno, V.; Renau-Piqueras, J.; Timoneda, J.; Barber, T. Vitamin A deficiency increases protein catabolism and induces urea cycle enzymes in rats. J. Nutr. 2010, 140, 792–798. [Google Scholar] [CrossRef]
- Ribot, J.; Felipe, F.; Bonet, M.L.; Palou, A. Changes of adiposity in response to vitamin A status correlate with changes of PPAR gamma 2 expression. Obes. Res. 2001, 9, 500–509. [Google Scholar] [CrossRef]
- Mercader, J.; Madsen, L.; Felipe, F.; Palou, A.; Kristiansen, K.; Bonet, M.L. All-trans retinoic acid increases oxidative metabolism in mature adipocytes. Cell. Physiol. Biochem. 2007, 20, 1061–1072. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, R.; Li, Y.; Chen, W.; Zhao, S.; Chen, G. Vitamin A status affects obesity development and hepatic expression of key genes for fuel metabolism in Zucker fatty rats. Biochem. Cell Biol. 2012, 90, 548–557. [Google Scholar] [CrossRef]
- Amengual, J.; Ribot, J.; Bonet, M.L.; Palou, A. Retinoic acid treatment enhances lipid oxidation and inhibits lipid biosynthesis capacities in the liver of mice. Cell. Physiol. Biochem. 2010, 25, 657–666. [Google Scholar] [CrossRef]
- Felipe, F.; Mercader, J.; Ribot, J.; Palou, A.; Bonet, M.L. Effects of retinoic acid administration and dietary vitamin A supplementation on leptin expression in mice: Lack of correlation with changes of adipose tissue mass and food intake. Biochim. Biophys. Acta 2005, 1740, 258–265. [Google Scholar] [CrossRef]
- Mercader, J.; Ribot, J.; Murano, I.; Felipe, F.; Cinti, S.; Bonet, M.L.; Palou, A. Remodeling of white adipose tissue after retinoic acid administration in mice. Endocrinology 2006, 147, 5325–5332. [Google Scholar]
- Manolescu, D.C.; Sima, A.; Bhat, P.V. All-trans retinoic acid lowers serum retinol-binding protein 4 concentrations and increases insulin sensitivity in diabetic mice. J. Nutr. 2010, 140, 311–316. [Google Scholar] [CrossRef]
- Jeyakumar, S.M.; Vajreswari, A.; Giridharan, N.V. Chronic dietary vitamin A supplementation regulates obesity in an obese mutant WNIN/Ob rat model. Obesity (Silver Spring) 2006, 14, 52–59. [Google Scholar] [CrossRef]
- Jeyakumar, S.M.; Vajreswari, A.; Giridharan, N.V. Vitamin A regulates obesity in WNIN/Ob obese rat; independent of stearoyl-CoA desaturase-1. Biochem. Biophys. Res. Commun. 2008, 370, 243–247. [Google Scholar] [CrossRef]
- Cisneros, F.J.; Gough, B.J.; Patton, R.E.; Ferguson, S.A. Serum levels of albumin, triglycerides, total protein and glucose in rats are altered after oral treatment with low doses of 13-cis-retinoic acid or all-trans-retinoic acid. J. Appl. Toxicol. 2005, 25, 470–478. [Google Scholar] [CrossRef]
- Sakamuri, V.P.; Ananthathmakula, P.; Veettil, G.N.; Ayyalasomayajula, V. Vitamin a decreases pre-receptor amplification of glucocorticoids in obesity: Study on the effect of vitamin A on 11beta-hydroxysteroid dehydrogenase type 1 activity in liver and visceral fat of WNIN/Ob obese rats. Nutr. J. 2011, 10, 70. [Google Scholar] [CrossRef]
- Hauner, H.; Schmid, P.; Pfeiffer, E.F. Glucocorticoids and insulin promote the differentiation of human adipocyte precursor cells into fat cells. J. Clin. Endocrinol. Metab. 1987, 64, 832–835. [Google Scholar] [CrossRef]
- Felipe, F.; Bonet, M.L.; Ribot, J.; Palou, A. Up-regulation of muscle uncoupling protein 3 gene expression in mice following high fat diet, dietary vitamin A supplementation and acute retinoic acid-treatment. Int. J. Obes. Relat. Metab. Disord. 2003, 27, 60–69. [Google Scholar] [CrossRef]
- Bairras, C.; Menard, L.; Redonnet, A.; Ferrand, C.; Delage, B.; Noel-Suberville, C.; Atgie, C.; Higueret, P. Effect of vitamin A content in cafeteria diet on the expression of nuclear receptors in rat subcutaneous adipose tissue. J. Physiol. Biochem. 2005, 61, 353–361. [Google Scholar] [CrossRef]
- Berry, D.C.; Noy, N. All-trans-retinoic acid represses obesity and insulin resistance by activating both peroxisome proliferation-activated receptor beta/delta and retinoic acid receptor. Mol. Cell. Biol. 2009, 29, 3286–3296. [Google Scholar] [CrossRef]
- Shaw, N.; Elholm, M.; Noy, N. Retinoic acid is a high affinity selective ligand for the peroxisome proliferator-activated receptor beta/delta. J. Biol. Chem. 2003, 278, 41589–41592. [Google Scholar]
- Kiefer, F.W.; Vernochet, C.; O’Brien, P.; Spoerl, S.; Brown, J.D.; Nallamshetty, S.; Zeyda, M.; Stulnig, T.M.; Cohen, D.E.; Kahn, C.R.; et al. Retinaldehyde dehydrogenase 1 regulates a thermogenic program in white adipose tissue. Nat. Med. 2012, 18, 918–925. [Google Scholar] [CrossRef]
- Zhang, M.; Hu, P.; Krois, C.R.; Kane, M.A.; Napoli, J.L. Altered vitamin A homeostasis and increased size and adiposity in the rdh1-null mouse. FASEB J. 2007, 21, 2886–2896. [Google Scholar] [CrossRef]
- Mercader, J.; Granados, N.; Bonet, M.L.; Palou, A. All-trans retinoic acid decreases murine adipose retinol binding protein 4 production. Cell. Physiol. Biochem. 2008, 22, 363–372. [Google Scholar] [CrossRef]
- Moise, A.R.; Kuksa, V.; Imanishi, Y.; Palczewski, K. Identification of all-trans-retinol: All-trans-13,14-dihydroretinol saturase. J. Biol. Chem. 2004, 279, 50230–50242. [Google Scholar]
- Moise, A.R.; Alvarez, S.; Dominguez, M.; Alvarez, R.; Golczak, M.; Lobo, G.P.; von Lintig, J.; de Lera, A.R.; Palczewski, K. Activation of retinoic acid receptors by dihydroretinoids. Mol. Pharmacol. 2009, 76, 1228–1237. [Google Scholar] [CrossRef]
- Moise, A.R.; Kuksa, V.; Blaner, W.S.; Baehr, W.; Palczewski, K. Metabolism and transactivation activity of 13,14-dihydroretinoic acid. J. Biol. Chem. 2005, 280, 27815–27825. [Google Scholar]
- Schupp, M.; Lefterova, M.I.; Janke, J.; Leitner, K.; Cristancho, A.G.; Mullican, S.E.; Qatanani, M.; Szwergold, N.; Steger, D.J.; Curtin, J.C.; et al. Retinol saturase promotes adipogenesis and is downregulated in obesity. Proc. Natl. Acad. Sci. USA 2009, 106, 1105–1110. [Google Scholar]
- Moise, A.R.; Lobo, G.P.; Erokwu, B.; Wilson, D.L.; Peck, D.; Alvarez, S.; Dominguez, M.; Alvarez, R.; Flask, C.A.; de Lera, A.R.; et al. Increased adiposity in the retinol saturase-knockout mouse. FASEB J. 2010, 24, 1261–1270. [Google Scholar]
- Frayn, K.N. Metabolic Regulation: A Human Perspective; John Wiley & Sons Ltd: Hoboken, NJ, USA, 2010. [Google Scholar]
- Puigserver, P.; Spiegelman, B.M. Peroxisome proliferator-activated receptor-gamma coactivator 1 alpha (PGC-1 alpha): Transcriptional coactivator and metabolic regulator. Endocr. Rev. 2003, 24, 78–90. [Google Scholar] [CrossRef]
- Alvarez, R.; Checa, M.; Brun, S.; Vinas, O.; Mampel, T.; Iglesias, R.; Giralt, M.; Villarroya, F. Both retinoic-acid-receptor- and retinoid-X-receptor-dependent signalling pathways mediate the induction of the brown-adipose-tissue-uncoupling-protein-1 gene by retinoids. Biochem. J. 2000, 345, 91–97. [Google Scholar]
- Teruel, T.; Hernandez, R.; Benito, M.; Lorenzo, M. Rosiglitazone and retinoic acid induce uncoupling protein-1 (UCP-1) in a p38 mitogen-activated protein kinase-dependent manner in fetal primary brown adipocytes. J. Biol. Chem. 2003, 278, 263–269. [Google Scholar]
- Carmona, M.C.; Valmaseda, A.; Iglesias, R.; Mampel, T.; Vinas, O.; Giralt, M.; Villarroya, F. 9-cis retinoic acid induces the expression of the uncoupling protein-2 gene in brown adipocytes. FEBS Lett. 1998, 441, 447–450. [Google Scholar] [CrossRef]
- Kumar, M.V.; Scarpace, P.J. Differential effects of retinoic acid on uncoupling protein-1 and leptin gene expression. J. Endocrinol. 1998, 157, 237–243. [Google Scholar] [CrossRef]
- Kumar, M.V.; Sunvold, G.D.; Scarpace, P.J. Dietary vitamin A supplementation in rats: Suppression of leptin and induction of UCP1 mRNA. J. Lipid Res. 1999, 40, 824–829. [Google Scholar]
- Mercader, J.; Palou, A.; Bonet, M. Induction of uncoupling protein-1 in mouse embryonic fibroblast-derived adipocytes by retinoic acid. Obesity (Silver Spring) 2010, 18, 655–662. [Google Scholar] [CrossRef]
- Bonet, M.L.; Oliver, J.; Pico, C.; Felipe, F.; Ribot, J.; Cinti, S.; Palou, A. Opposite effects of feeding a vitamin A-deficient diet and retinoic acid treatment on brown adipose tissue uncoupling protein 1 (UCP1), UCP2 and leptin expression. J. Endocrinol. 2000, 166, 511–517. [Google Scholar] [CrossRef]
- Valmaseda, A.; Carmona, M.C.; Barbera, M.J.; Vinas, O.; Mampel, T.; Iglesias, R.; Villarroya, F.; Giralt, M. Opposite regulation of PPAR-alpha and -gamma gene expression by both their ligands and retinoic acid in brown adipocytes. Mol. Cell. Endocrinol. 1999, 154, 101–109. [Google Scholar] [CrossRef]
- Zhang, Y.; Matheny, M.; Zolotukhin, S.; Tumer, N.; Scarpace, P.J. Regulation of adiponectin and leptin gene expression in white and brown adipose tissues: Influence of beta3-adrenergic agonists, retinoic acid, leptin and fasting. Biochim. Biophys. Acta 2002, 1584, 115–122. [Google Scholar] [CrossRef]
- Sutton, A.L.; MacDonald, P.N. Vitamin D: More than a “Bone-a-fide” Hormone. Mol. Endocrinol. 2003, 17, 777–791. [Google Scholar] [CrossRef]
- Haussler, M.R.; Jurutka, P.W.; Mizwicki, M.; Norman, A.W. Vitamin D receptor (VDR)-mediated actions of 1α,25(OH)2vitamin D3: Genomic and non-genomic mechanisms. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 543–559. [Google Scholar] [CrossRef]
- Heaney, R.P.; Horst, R.L.; Cullen, D.M.; Armas, L.A. Vitamin D3 distribution and status in the body. J. Am. Coll. Nutr. 2009, 28, 252–256. [Google Scholar]
- Rosenstreich, S.J.; Rich, C.; Volwiler, W. Deposition in and release of vitamin D3 from body fat: Evidence for a storage site in the rat. J. Clin. Invest. 1971, 50, 679–687. [Google Scholar] [CrossRef]
- Blum, M.; Dolnikowski, G.; Seyoum, E.; Harris, S.S.; Booth, S.L.; Peterson, J.; Saltzman, E.; Dawson-Hughes, B. Vitamin D3 in fat tissue. Endocrine 2008, 33, 90–94. [Google Scholar] [CrossRef]
- Pramyothin, P.; Biancuzzo, R.M.; Lu, Z.; Hess, D.T.; Apovian, C.M.; Holick, M.F. Vitamin D in adipose tissue and serum 25-hydroxyvitamin d after roux-en-Y gastric bypass. Obesity (Silver Spring) 2011, 19, 2228–2234. [Google Scholar] [CrossRef]
- Earthman, C.P.; Beckman, L.M.; Masodkar, K.; Sibley, S.D. The link between obesity and low circulating 25-hydroxyvitamin D concentrations: Considerations and implications. Int. J. Obes. (Lond.) 2011, 36, 387–396. [Google Scholar]
- Drincic, A.T.; Armas, L.A.; Van Diest, E.E.; Heaney, R.P. Volumetric dilution, rather than sequestration best explains the low vitamin D status of obesity. Obesity (Silver Spring) 2012, 20, 1444–1448. [Google Scholar] [CrossRef]
- Wamberg, L.; Christiansen, T.; Paulsen, S.K.; Fisker, S.; Rask, P.; Rejnmark, L.; Richelsen, B.; Pedersen, S.B. Expression of vitamin D-metabolizing enzymes in human adipose tissue-the effect of obesity and diet-induced weight loss. Int. J. Obes. (Lond.) 2012. [Google Scholar] [CrossRef]
- Ishida, Y.; Taniguchi, H.; Baba, S. Possible involvement of 1α,25-dihydroxyvitamin D3 in proliferation and differentiation of 3T3-L1 cells. Biochem. Biophys. Res. Commun. 1988, 151, 1122–1127. [Google Scholar] [CrossRef]
- Kamei, Y.; Kawada, T.; Kazuki, R.; Ono, T.; Kato, S.; Sugimoto, E. Vitamin D receptor gene expression is up-regulated by 1,25-dihydroxyvitamin D3 in 3T3-L1 preadipocytes. Biochem. Biophys. Res. Commun. 1993, 193, 948–955. [Google Scholar] [CrossRef]
- Sato, M.; Hiragun, A. Demonstration of 1α,25-dihydroxyvitamin D3 receptor-like molecule in ST 13 and 3T3 L1 preadipocytes and its inhibitory effects on preadipocyte differentiation. J. Cell. Physiol. 1988, 135, 545–550. [Google Scholar] [CrossRef]
- Hida, Y.; Kawada, T.; Kayahashi, S.; Ishihara, T.; Fushiki, T. Counteraction of retinoic acid and 1,25-dihydroxyvitamin D3 on up-regulation of adipocyte differentiation with PPARgamma ligand, an antidiabetic thiazolidinedione, in 3T3-L1 cells. Life Sci. 1998, 62, PL205–PL211. [Google Scholar] [CrossRef]
- Zhuang, H.; Lin, Y.; Yang, G. Effects of 1,25-dihydroxyvitamin D3 on proliferation and differentiation of porcine preadipocyte in vitro. Chem. Biol. Interact. 2007, 170, 114–123. [Google Scholar] [CrossRef]
- Sun, X.; Zemel, M.B. Role of uncoupling protein 2 (UCP2) expression and 1α,25-dihydroxyvitamin D3 in modulating adipocyte apoptosis. FASEB J. 2004, 18, 1430–1432. [Google Scholar]
- Kong, J.; Li, Y.C. Molecular mechanism of 1,25-dihydroxyvitamin D3 inhibition of adipogenesis in 3T3-L1 cells. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E916–E924. [Google Scholar] [CrossRef]
- Sun, X.; Zemel, M.B. Calcium and 1,25-dihydroxyvitamin D3 regulation of adipokine expression. Obesity (Silver Spring) 2007, 15, 340–348. [Google Scholar] [CrossRef]
- Sun, X.; Morris, K.L.; Zemel, M.B. Role of calcitriol and cortisol on human adipocyte proliferation and oxidative and inflammatory stress: A microarray study. J. Nutrigenet. Nutrigenomics 2008, 1, 30–48. [Google Scholar] [CrossRef]
- Zemel, M.B.; Sun, X. Calcitriol and energy metabolism. Nutr. Rev. 2008, 66, S139–S146. [Google Scholar] [CrossRef]
- Gao, D.; Trayhurn, P.; Bing, C. 1,25-dihydroxyvitamin D(3) inhibits the cytokine-induced secretion of MCP-1 and reduces monocyte recruitment by human preadipocytes. Int. J. Obes. (Lond.) 2012. [Google Scholar] [CrossRef]
- Lorente-Cebrian, S.; Eriksson, A.; Dunlop, T.; Mejhert, N.; Dahlman, I.; Astrom, G.; Sjolin, E.; Wahlen, K.; Carlberg, C.; Laurencikiene, J.; et al. Differential effects of 1α,25-dihydroxycholecalciferol on MCP-1 and adiponectin production in human white adipocytes. Eur. J. Nutr. 2011, 51, 335–342. [Google Scholar]
- Mutt, S.J.; Karhu, T.; Lehtonen, S.; Lehenkari, P.; Carlberg, C.; Saarnio, J.; Sebert, S.; Hypponen, E.; Jarvelin, M.R.; Herzig, K.H. Inhibition of cytokine secretion from adipocytes by 1,25-dihydroxyvitamin D3 via the NF-kappaB pathway. FASEBJ. 2012, 26, 4400–4407. [Google Scholar] [CrossRef]
- Marcotorchino, J.; Gouranton, E.; Romier, B.; Malezet, C.; Riollet, C.; Landrier, J.F. Vitamin D reduces inflammatory markers induction mediated by tnfa via induction of dusp10. Unpublished work, 2012. [Google Scholar]
- Narvaez, C.J.; Matthews, D.; Broun, E.; Chan, M.; Welsh, J. Lean phenotype and resistance to diet-induced obesity in vitamin D receptor knockout mice correlates with induction of uncoupling protein-1 in white adipose tissue. Endocrinology 2009, 150, 651–661. [Google Scholar]
- Wong, K.E.; Szeto, F.L.; Zhang, W.; Ye, H.; Kong, J.; Zhang, Z.; Sun, X.J.; Li, Y.C. Involvement of the vitamin D receptor in energy metabolism: Regulation of uncoupling proteins. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E820–E828. [Google Scholar] [CrossRef]
- Wong, K.E.; Kong, J.; Zhang, W.; Szeto, F.L.; Ye, H.; Deb, D.K.; Brady, M.J.; Li, Y.C. Targeted expression of human vitamin D receptor in adipocytes decreases energy expenditure and induces obesity in mice. J. Biol. Chem. 2011, 286, 33804–33810. [Google Scholar]
- Jensen, S.K.; Lauridsen, C. Alpha-tocopherol stereoisomers. Vitam. Horm. 2007, 76, 281–308. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Sundaram, C.; Prasad, S.; Kannappan, R. Tocotrienols, the vitamin E of the 21st century: Its potential against cancer and other chronic diseases. Biochem. Pharmacol. 2010, 80, 1613–1631. [Google Scholar] [CrossRef]
- Brigelius-Flohe, R. Vitamin E: The shrew waiting to be tamed. Free Radic. Biol. Med. 2009, 46, 543–554. [Google Scholar] [CrossRef]
- Azzi, A. Molecular mechanism of alpha-tocopherol action. Free Radic. Biol. Med. 2007, 43, 16–21. [Google Scholar] [CrossRef]
- Traber, M.G. Vitamin E regulatory mechanisms. Annu. Rev. Nutr. 2007, 27, 347–362. [Google Scholar] [CrossRef]
- Traber, M.G.; Kayden, H.J. Tocopherol distribution and intracellular localization in human adipose tissue. Am. J. Clin. Nutr. 1987, 46, 488–495. [Google Scholar]
- Burton, G.W.; Traber, M.G.; Acuff, R.V.; Walters, D.N.; Kayden, H.; Hughes, L.; Ingold, K.U. Human plasma and tissue alpha-tocopherol concentrations in response to supplementation with deuterated natural and synthetic vitamin E. Am. J. Clin. Nutr. 1998, 67, 669–684. [Google Scholar]
- Ikeda, S.; Toyoshima, K.; Yamashita, K. Dietary sesame seeds elevate alpha- and gamma-tocotrienol concentrations in skin and adipose tissue of rats fed the tocotrienol-rich fraction extracted from palm oil. J. Nutr. 2001, 131, 2892–2897. [Google Scholar]
- Uchida, T.; Abe, C.; Nomura, S.; Ichikawa, T.; Ikeda, S. Tissue distribution of alpha- and gamma-tocotrienol and gamma-tocopherol in rats and interference with their accumulation by alpha-tocopherol. Lipids 2012, 47, 129–139. [Google Scholar] [CrossRef]
- Handelman, G.J.; Epstein, W.L.; Peerson, J.; Spiegelman, D.; Machlin, L.J.; Dratz, E.A. Human adipose alpha-tocopherol and gamma-tocopherol kinetics during and after 1 y of alpha-tocopherol supplementation. Am. J. Clin. Nutr. 1994, 59, 1025–1032. [Google Scholar]
- Traber, M.G.; Leonard, S.W.; Traber, D.L.; Traber, L.D.; Gallagher, J.; Bobe, G.; Jeschke, M.G.; Finnerty, C.C.; Herndon, D. Alpha-tocopherol adipose tissue stores are depleted after burn injury in pediatric patients. Am. J. Clin. Nutr. 2010, 92, 1378–1384. [Google Scholar] [CrossRef]
- Landes, N.; Pfluger, P.; Kluth, D.; Birringer, M.; Ruhl, R.; Bol, G.F.; Glatt, H.; Brigelius-Flohe, R. Vitamin E activates gene expression via the pregnane X receptor. Biochem. Pharmacol. 2003, 65, 269–273. [Google Scholar]
- Ricciarelli, R.; Tasinato, A.; Clement, S.; Ozer, N.K.; Boscoboinik, D.; Azzi, A. alpha-Tocopherol specifically inactivates cellular protein kinase C alpha by changing its phosphorylation state. Biochem. J. 1998, 334, 243–249. [Google Scholar]
- Maggi-Capeyron, M.F.; Ceballos, P.; Cristol, J.P.; Delbosc, S.; Le Doucen, C.; Pons, M.; Leger, C.L.; Descomps, B. Wine phenolic antioxidants inhibit AP-1 transcriptional activity. J. Agric. Food Chem. 2001, 49, 5646–5652. [Google Scholar] [CrossRef]
- Landrier, J.F.; Gouranton, E.; El Yazidi, C.; Malezet, C.; Balaguer, P.; Borel, P.; Amiot, M.J. Adiponectin expression is induced by vitamin E via a peroxisome proliferator-activated receptor gamma-dependent mechanism. Endocrinology 2009, 150, 5318–5325. [Google Scholar] [CrossRef]
- Landrier, J.F.; Gouranton, E.; Reboul, E.; Cardinault, N.; El Yazidi, C.; Malezet-Desmoulins, C.; Andre, M.; Nowicki, M.; Souidi, M.; Borel, P. Vitamin E decreases endogenous cholesterol synthesis and apo-AI-mediated cholesterol secretion in Caco-2 cells. J. Nutr. Biochem. 2010, 21, 1207–1213. [Google Scholar] [CrossRef]
- Uto-Kondo, H.; Ohmori, R.; Kiyose, C.; Kishimoto, Y.; Saito, H.; Igarashi, O.; Kondo, K. Tocotrienol suppresses adipocyte differentiation and Akt phosphorylation in 3T3-L1 preadipocytes. J. Nutr. 2009, 139, 51–57. [Google Scholar]
- Matsunaga, T.; Shoji, A.; Gu, N.; Joo, E.; Li, S.; Adachi, T.; Yamazaki, H.; Yasuda, K.; Kondoh, T.; Tsuda, K. γ-Tocotrienol attenuates TNF-α-induced changes in secretion and gene expression of MCP-1, IL-6 and adiponectin in 3T3-L1 adipocytes. Mol. Med. Report 2012, 5, 905–909. [Google Scholar]
- Lira, F.S.; Rosa, J.C.; Cunha, C.A.; Ribeiro, E.B.; do Nascimento, C.O.; Oyama, L.M.; Mota, J.F. Supplementing alpha-tocopherol (vitamin E) and vitamin D3 in high fat diet decrease IL-6 production in murine epididymal adipose tissue and 3T3-L1 adipocytes following LPS stimulation. Lipids Health Dis. 2011, 10, 37. [Google Scholar] [CrossRef]
- Shen, X.H.; Tang, Q.Y.; Huang, J.; Cai, W. Vitamin E regulates adipocytokine expression in a rat model of dietary-induced obesity. Exp. Biol. Med. (Maywood) 2010, 235, 47–51. [Google Scholar] [CrossRef]
- Isermann, B.; Bierhaus, A.; Tritschler, H.; Ziegler, R.; Nawroth, P.P. Alpha-tocopherol induces leptin expression in healthy individuals and in vitro. Diabetes Care 1999, 22, 1227–1228. [Google Scholar]
- Tontonoz, P.; Spiegelman, B.M. Fat and beyond: The diverse biology of PPARgamma. Annu. Rev. Biochem. 2008, 77, 289–312. [Google Scholar] [CrossRef]
- Booth, S.L. Roles for vitamin K beyond coagulation. Annu. Rev. Nutr. 2009, 29, 89–110. [Google Scholar] [CrossRef]
- Shea, M.K.; Booth, S.L.; Gundberg, C.M.; Peterson, J.W.; Waddell, C.; Dawson-Hughes, B.; Saltzman, E. Adulthood obesity is positively associated with adipose tissue concentrations of vitamin K and inversely associated with circulating indicators of vitamin K status in men and women. J. Nutr. 2010, 140, 1029–1034. [Google Scholar] [CrossRef]
- Okano, T.; Shimomura, Y.; Yamane, M.; Suhara, Y.; Kamao, M.; Sugiura, M.; Nakagawa, K. Conversion of phylloquinone (vitamin K1) into menaquinone-4 (vitamin K2) in mice: Two possible routes for menaquinone-4 accumulation in cerebra of mice. J. Biol. Chem. 2008, 283, 11270–11279. [Google Scholar]
- Landes, N.; Birringer, M.; Brigelius-Flohe, R. Homologous metabolic and gene activating routes for vitamins E and K. Mol. Aspects Med. 2003, 24, 337–344. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Suzawa, M.; Fukumoto, S.; Fujita, T. Vitamin K(2) inhibits adipogenesis, osteoclastogenesis, and ODF/RANK ligand expression in murine bone marrow cell cultures. Bone 2000, 27, 769–776. [Google Scholar] [CrossRef]
- Garcia Palacios, V.; Morita, I.; Murota, S. Expression of adipogenesis markers in a murine stromal cell line treated with 15-deoxy delta(12,14)-prostaglandin J2, interleukin-11, 9-cis retinoic acid and vitamin K2. Prostaglandins Leukot. Essent. Fatty Acids 2001, 65, 215–221. [Google Scholar] [CrossRef]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Aspects Med. 2005, 26, 459–516. [Google Scholar] [CrossRef]
- Tourniaire, F.; Gouranton, E.; von Lintig, J.; Keijer, J.; Luisa Bonet, M.; Amengual, J.; Lietz, G.; Landrier, J.F. Beta-carotene conversion products and their effects on adipose tissue. Genes Nutr. 2009, 4, 179–187. [Google Scholar] [CrossRef]
- Lietz, G.; Oxley, A.; Boesch-Saadatmandi, C.; Kobayashi, D. Importance of beta,beta-carotene 15,15′-monooxygenase 1 (BCMO1) and beta,beta-carotene 9′,10′-dioxygenase 2 (BCDO2) in nutrition and health. Mol. Nutr. Food Res. 2012, 56, 241–250. [Google Scholar] [CrossRef]
- Virtanen, S.M.; van’t Veer, P.; Kok, F.; Kardinaal, A.F.; Aro, A. Predictors of adipose tissue carotenoid and retinol levels in nine countries. The euramic study. Am. J. Epidemiol. 1996, 144, 968–979. [Google Scholar] [CrossRef]
- Parker, R.S. Carotenoids in human blood and tissues. J. Nutr. 1989, 119, 101–104. [Google Scholar]
- Chung, H.Y.; Ferreira, A.L.; Epstein, S.; Paiva, S.A.; Castaneda-Sceppa, C.; Johnson, E.J. Site-specific concentrations of carotenoids in adipose tissue: Relations with dietary and serum carotenoid concentrations in healthy adults. Am. J. Clin. Nutr. 2009, 90, 533–539. [Google Scholar] [CrossRef]
- Sy, C.; Gleize, B.; Dangles, O.; Landrier, J.F.; Veyrat, C.C.; Borel, P. Effects of physicochemical properties of carotenoids on their bioaccessibility, intestinal cell uptake, and blood and tissue concentrations. Mol. Nutr. Food Res. 2012, 56, 1385–1397. [Google Scholar] [CrossRef]
- Moussa, M.; Gouranton, E.; Gleize, B.; Yazidi, C.E.; Niot, I.; Besnard, P.; Borel, P.; Landrier, J.F. CD36 is involved in lycopene and lutein uptake by adipocytes and adipose tissue cultures. Mol. Nutr. Food Res. 2011, 55, 578–584. [Google Scholar] [CrossRef]
- Gouranton, E.; Yazidi, C.E.; Cardinault, N.; Amiot, M.J.; Borel, P.; Landrier, J.F. Purified low-density lipoprotein and bovine serum albumin efficiency to internalise lycopene into adipocytes. Food Chem. Toxicol. 2008, 46, 3832–3836. [Google Scholar] [CrossRef]
- Rao, A.V.; Ray, M.R.; Rao, L.G. Lycopene. Adv. Food Nutr. Res. 2006, 51, 99–164. [Google Scholar]
- Ben-Dor, A.; Steiner, M.; Gheber, L.; Danilenko, M.; Dubi, N.; Linnewiel, K.; Zick, A.; Sharoni, Y.; Levy, J. Carotenoids activate the antioxidant response element transcription system. Mol. Cancer Ther. 2005, 4, 177–186. [Google Scholar]
- Landrier, J.F. Les Phytomicronutriments; Lavoisier: Paris, France, 2012. [Google Scholar]
- Kawada, T.; Kamei, Y.; Fujita, A.; Hida, Y.; Takahashi, N.; Sugimoto, E.; Fushiki, T. Carotenoids and retinoids as suppressors on adipocyte differentiation via nuclear receptors. Biofactors 2000, 13, 103–109. [Google Scholar] [CrossRef]
- Ziouzenkova, O.; Orasanu, G.; Sukhova, G.; Lau, E.; Berger, J.P.; Tang, G.; Krinsky, N.I.; Dolnikowski, G.G.; Plutzky, J. Asymmetric cleavage of beta-carotene yields a transcriptional repressor of retinoid X receptor and peroxisome proliferator-activated receptor responses. Mol. Endocrinol. 2007, 21, 77–88. [Google Scholar]
- Shirakura, Y.; Takayanagi, K.; Mukai, K.; Tanabe, H.; Inoue, M. Beta-cryptoxanthin suppresses the adipogenesis of 3T3-L1 cells via rar activation. J. Nutr. Sci. Vitaminol. (Tokyo) 2011, 57, 426–431. [Google Scholar] [CrossRef]
- Inoue, M.; Tanabe, H.; Matsumoto, A.; Takagi, M.; Umegaki, K.; Amagaya, S.; Takahashi, J. Astaxanthin functions differently as a selective peroxisome proliferator-activated receptor gamma modulator in adipocytes and macrophages. Biochem. Pharmacol. 2012, 84, 692–700. [Google Scholar] [CrossRef]
- Gouranton, E.; Aydemir, G.; Reynaud, E.; Marcotorchino, J.; Malezet, C.; Caris-Veyrat, C.; Blomhoff, R.; Landrier, J.F.; Ruhl, R. Apo-10′-lycopenoic acid impacts adipose tissue biology via the retinoic acid receptors. Biochim. Biophys. Acta 2011, 1811, 1105–1114. [Google Scholar] [CrossRef]
- Landrier, J.-F. Unpublished work, Institut National de Recherche Agronomique (INRA): Marseille, France, 2012.
- Kameji, H.; Mochizuki, K.; Miyoshi, N.; Goda, T. Beta-carotene accumulation in 3T3-L1 adipocytes inhibits the elevation of reactive oxygen species and the suppression of genes related to insulin sensitivity induced by tumor necrosis factor-alpha. Nutrition 2010, 26, 1151–1156. [Google Scholar] [CrossRef]
- Marcotorchino, J.; Romier, B.; Gouranton, E.; Riollet, C.; Gleize, B.; Malezet-Desmoulins, C.; Landrier, J.F. Lycopene attenuates LPS-induced TNF-alpha secretion in macrophages and inflammatory markers in adipocytes exposed to macrophage-conditioned media. Mol. Nutr. Food Res. 2012, 56, 725–732. [Google Scholar]
- Ikeuchi, M.; Koyama, T.; Takahashi, J.; Yazawa, K. Effects of astaxanthin in obese mice fed a high-fat diet. Biosci. Biotechnol. Biochem. 2007, 71, 893–899. [Google Scholar] [CrossRef]
- Arunkumar, E.; Bhuvaneswari, S.; Anuradha, C.V. An intervention study in obese mice with astaxanthin, a marine carotenoid-Effects on insulin signaling and pro-inflammatory cytokines. Food Funct. 2012, 3, 120–126. [Google Scholar] [CrossRef]
- Takayanagi, K.; Morimoto, S.; Shirakura, Y.; Mukai, K.; Sugiyama, T.; Tokuji, Y.; Ohnishi, M. Mechanism of visceral fat reduction in tsumura suzuki obese, diabetes (TSOD) mice orally administered beta-cryptoxanthin from satsuma mandarin oranges (Citrus unshiu Marc). J. Agric. Food Chem. 2011, 59, 12342–12351. [Google Scholar]
- Amengual, J.; Gouranton, E.; van Helden, Y.G.; Hessel, S.; Ribot, J.; Kramer, E.; Kiec-Wilk, B.; Razny, U.; Lietz, G.; Wyss, A.; et al. Beta-carotene reduces body adiposity of mice via BCMO1. PLoS One 2011, 6, e20644. [Google Scholar]
- van Helden, Y.G.; Godschalk, R.W.; von Lintig, J.; Lietz, G.; Landrier, J.F.; Luisa Bonet, M.; van Schooten, F.J.; Keijer, J. Gene expression response of mouse lung, liver and white adipose tissue to beta-carotene supplementation, knockout of Bcmo1 and sex. Mol. Nutr. Food Res. 2011, 55, 1466–1474. [Google Scholar] [CrossRef]
- Murano, I.; Morroni, M.; Zingaretti, M.C.; Oliver, P.; Sanchez, J.; Fuster, A.; Pico, C.; Palou, A.; Cinti, S. Morphology of ferret subcutaneous adipose tissue after 6-month daily supplementation with oral beta-carotene. Biochim. Biophys. Acta 2005, 1740, 305–312. [Google Scholar] [CrossRef]
- Lobo, G.P.; Amengual, J.; Li, H.N.; Golczak, M.; Bonet, M.L.; Palczewski, K.; von Lintig, J. Beta,beta-carotene decreases peroxisome proliferator receptor gamma activity and reduces lipid storage capacity of adipocytes in a beta,beta-carotene oxygenase 1-dependent manner. J. Biol. Chem. 2010, 285, 27891–27899. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Landrier, J.-F.; Marcotorchino, J.; Tourniaire, F. Lipophilic Micronutrients and Adipose Tissue Biology. Nutrients 2012, 4, 1622-1649. https://doi.org/10.3390/nu4111622
Landrier J-F, Marcotorchino J, Tourniaire F. Lipophilic Micronutrients and Adipose Tissue Biology. Nutrients. 2012; 4(11):1622-1649. https://doi.org/10.3390/nu4111622
Chicago/Turabian StyleLandrier, Jean-François, Julie Marcotorchino, and Franck Tourniaire. 2012. "Lipophilic Micronutrients and Adipose Tissue Biology" Nutrients 4, no. 11: 1622-1649. https://doi.org/10.3390/nu4111622