Human Folate Bioavailability

Abstract
:1. Introduction
2. Synopsis of Post-Absorptive Folate Metabolism
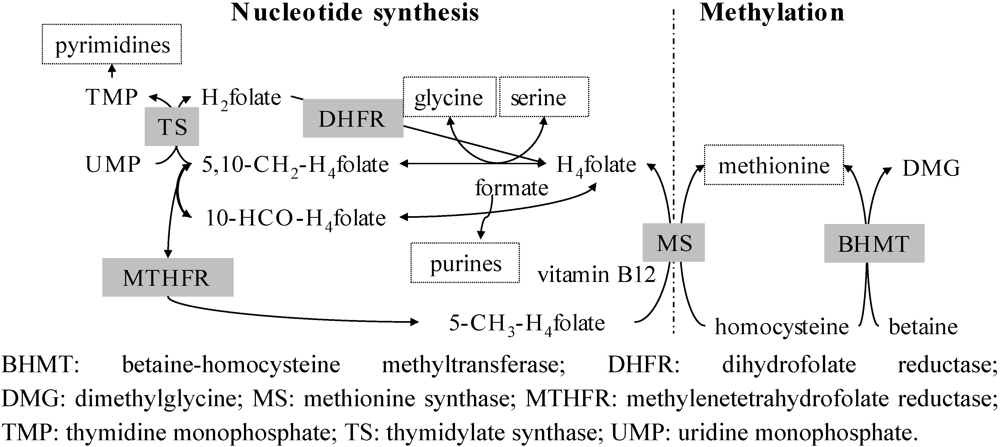
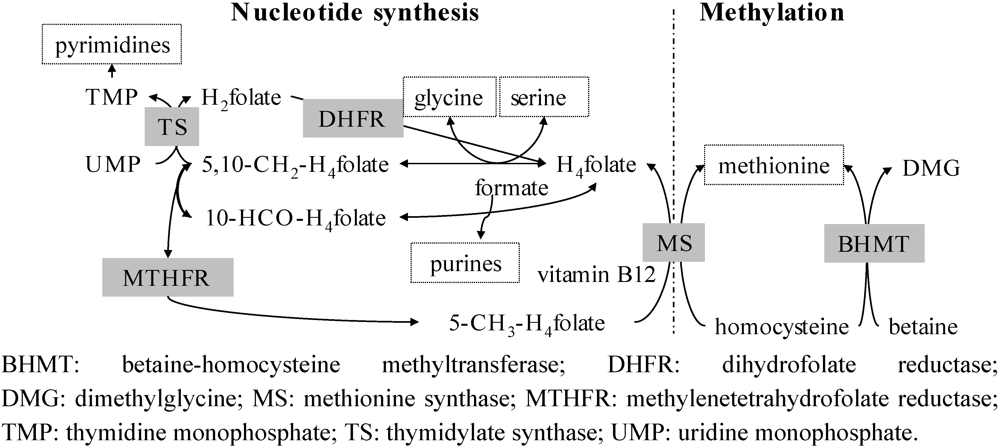
3. Estimation of Folate Bioavailability-Still a Challenge

{kind=link}
{kind=link}
| Intervention diet | Folate intake | W | N | Age | Erythrocyte folate | p/s-folate | tHcy (µmol/L) | Ref. | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dose (µg/day) (Q) 1 | Total 2(µg/day) | Baseline(nmol/L) | End(%) | Baseline(nmol/L) | End(%) | Baseline(nmol/L) | End(%) | |||||
| Control | 2 (C) 1 | 4 | 35, F | 41 ± 3 3 | 15 ± 9 3 | −15 | 11 ± 4 3 | −10 | [68] | |||
| 5-a-day fruit/veg | 43 (C) 1 | 4 | 36, F | 42 ± 4 3 | 17 ± 9 3 | −6 | 10 ± 3 3 | 0 | ||||
| Control | 0 (T) 1 | 186 | 4 | 18, M | 23-39 4 | 16 ± 9 3 | −7 | 12 ± 4 3 | 0 | [13] | ||
| Spinach | 200 (T) 1 | 384 | 4 | 18, M | 23-39 4 | 14 ± 5 3 | −7 | 12 ± 3 3 | 0 | |||
| Yeast | 200 (T) 1 | 411 | 4 | 19, M | 23-39 4 | 15 ± 7 3 | −17 | 12 ± 3 3 | −9 | |||
| Control | 23 (L) 1 | 210 | 12 | 9, F | 17-40 4 | 709 ± 157 3,5 | +3 5 | [66] | ||||
| High folate diet | 201 (L) 1 | 410 | 12 | 10, F | 17-40 4 | 797 ± 200 3,5 | +7 5 | |||||
| Intervention diet | Folate intake (µg/day) | W | N2 | Age | Erythrocyte folate | p/s-folate | tHcy | Ref. | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dose (Q) 1 | Total | Baseline (nmol/L) | End (%) | Baseline (nmol/L) | End (%) | Baseline (µmol/L) | End (%) | |||||
| Control | 0 | 236 | 16 | 43 | - | 26 ± 8 3 | −4 | 9 ± 3 3 | 0 | [69] | ||
| 5-a-day fruit/vegetables | 63 (C) | 306 | 16 | 41 | - | 26 ± 7 3 | +7 ♦ | 10 ± 4 3 | −11 | |||
| Low folate diet | 0 (L) | 131 | 4 | 23 | 50 ± 4 3 | 13 ± 6 3 | −8 | 14 ± 6 3 | −8 | [67] | ||
| 500 g fruit and vegetables | 97 (L) | 228 | 4 | 24 | 49 ± 6 3 | 16 ± 10 3 | 0 ♦ | 12 ± 4 3 | −9 ♦ | |||
| Control | 0 | 239 | 12 | 17, F | 48 (38, 57) 7 | 900 (647, 1079) 7 | −22 7 | 12 (11, 15) | 0 7 | 9 (7, 11) | +10 7 | [59] |
| 5 slices bread | 70 (T) | 379 | 12 | 17, F | 47 (40, 55) 7 | 855 (635, 992) 7 | −18 | 12 (9-16) | 0 | 9 (8, 11) | −21 * | |
| Breakfast (SNO [71]) | 125 (T) | 437 | 12 | 17, F | 49 (35, 53) 7 | 805 (742, 909) 7 | +12 ♦ | 12 (10, 16) | +8 | 7 (7, 9) | −23 *,♦ | |
| Folic acid-fortified bread | 188 (T) | 461 | 4 | 31 | 36 ± 13 3 | 606 ± 224 3 | +13 * | 12 ± 5 3 | +25 * | [60] | ||
| Rye and orange juice | 184 (T) | 461 | 4 | 33 | 36 ± 13 3 | 561 ± 190 3 | +15 * | 10 ± 3 3 | +23 * | |||
| Folic acid | 150 (T) | 221 4 | 4 | 15 | 19-48 5 | 12 ± 4 3 | +20 | 10 ± 2 3 | −11 | [61] | ||
| Vegetables, fruits, liver pate | 331 (T) | 427 4 | 4 | 29 | 19-41 5 | 12 ± 4 3 | +33 ♦ | 10 ± 3 3 | −11 | |||
| Placebo | 0 | ~360 9 | 16 | 43 | 46 ± 2 | 895 ±6 0 | +3 | 26 ±2 | −1 | 9 ± 0.4 | 0 | [19] |
| Food group | 200 (C) | ~305 9 | 16 | 38 | 45 ± 2 | 872 ± 61 | +14 ♦ | 25 ± 2 | +19 ♦ | 10 ± 1 | −6 ♦ | |
| 5-methyltetrahydrofolate caps | 200 (P) | ~295 9 | 16 | 42 | 45 ± 2 | 793 ± 42 | +30 ♦ | 22 ± 2 | +41 ♦ | 10 ± 1 | −14 ♦ | |
| Folic acid caps | 200 (P) | ~325 9 | 16 | 40 | 48 ± 2 | 833 ± 56 | +28 ♦ | 24 ± 2 | +44 ♦ | 9 ± 0.4 | −15 ♦ | |
| Control | 0 | 210 4 | 4 | 22 | 23 ± 8 3 | 347 ± 793 | −1 | 13 ± 3 3 | 0 | 10 ± 33 | +9 | [14] |
| Fruit and vegetables | 350 (T) | 560 4 | 4 | 23 | 23 ± 8 3 | 338 ± 813 | +16 ♦ | 14 ± 3 3 | +30 ♦ | 11 ± 53 | −10 ♦ | |
| Control | 0 (T) | 242 | 4 | 14 | 60 ± 15 3 | 18 (15-22) 6 | 0 6 | 14 (12-16) 6 | 0 6 | [62] | ||
| High folate diet | 355 (T) | 618 | 4 | 20 | 58 ± 18 3 | 18 (16-19) 6 | 28 6,♦ | 12 (11-13) 6 | −9 6,♦ | |||
| Control | 0 (T) | 227 | 12 | 15 | 36-71 8 | 539 ± 166 3 | 0 | 15 (12-18) 6 | −7 6 | 12 (11-14) 6 | 0 6 | [58] |
| High folate diet | ~350 (T) | 707 | 12 | 15 | 36-71 8 | 571 ± 162 3 | +11 | 15 (13-17) 6 | +32 6,♦ | 11 (9-12) 6 | −10 6 | |
| Pericarp flour bread | 223 (T) | 436 | 16 | 25 | 48-56 5 | 497 (414-581) 6 | +14 6 | 13 (11-15) 6 | +7 6 | 10 (8-11) 6 | −25 6 | [63] |
| Aleurone flour bread | 615 (T) | 836 | 16 | 25 | 46-54 5 | 509 (434-584) 6 | +34 6,♦ | 13 (10-16) 6 | +52 6,♦ | 9 (8-10) 6 | −29 6,♦ | |
| Dose nmol 1 | AUC or App Abs 2 | Sampling duration (nr/4 h) 3 | Females + males | Age 4 | Ref. | |
|---|---|---|---|---|---|---|
| Size of AUC (h∙nmol/L) from supplements and fortified foods | ||||||
| [13C6]-folic acid | 634 | 19 | 8 h (8) | 10 10 | 31 ± 1 | [17] |
| Folic acid 5 | 1134 | 146 | 10 h (6) | 0 + 13 | 26 ± 6 | [64] |
| Folic acid | 1134 | 40 | 10 h (6) | 6 + 6 | 36-69 | [45] |
| Folic acid | 1193 | 37, 49 7 | 7 h (3) | 8 + 8 | 20-50 | [65] |
| Folic acid | 907 | 62 | 10 h (4) | 10 + 10 | 27 ± 3 | [70] |
| [13C5]-folic acid in bread | 450 (D) | 28 | 12 h (7) | 5 + 3 | 39-66 | [46] |
| [13C5]-folic acid in breakfast meal | 450 (D) | 26 | 12 h (7) | 5 + 3 | 39-66 | [46] |
| (6S)-[13C6]-5-HCO-H4folate | 500 | 42 | 8 h (8) | 10 10 | 31 ± 1 | [17] |
| (6S)-5-CH3-H4folate 5 | 1088 | 142 | 10 h (6) | 0 + 13 | 26 ± 6 | [64] |
| (6S)-5-CH3-H4folate 5 | 830 6 | 44, 88 6 | 10 h (7) | 0 + 2 | 55, 77 | [40] |
| (6S)-5-CH3-H4folate i.m. 5 | 830 6 | 111, 144 6 | 10 h (7) | 0 + 2 | 55, 77 | [40] |
| (6S)-[13C5]-5-CH3-H4folate in bread | 450 (D) | 66 | 12 h (7) | 5 + 3 | 39-66 | [46] |
| Size of AUC (h∙nmol/L) from native food folates | ||||||
| Broccoli 5 | 440 (M) | 27, 41 6 | 10 h (7) | 0 + 2 | 55, 77 | [40] |
| Strawberries 5 | 450 (M) | 32, 41 6 | 10 h (7) | 0 + 2 | 55, 77 | [40] |
| Spinach, monoglutamate 8 | 820 (T) | 31 | 10 h (6) | 6 + 6 | 36-69 | [45] |
| Spinach, polyglutamate 9 | 990 (T) | 27 | 10 h (6) | 6 + 6 | 36-69 | [45] |
| Spinach | 544 (C) | 41 | 10 h (4) | 10 + 10 | 27 ± 3 | [70] |
| 1088 (C) | 71 | |||||
| Aleurone flour | 1167 (M) | 46, 38 7 | 7 h (3) | 8 + 8 | 20-50 | [65] |
| Wheat bran | 213 (M) | 8, 6 7 | 7 h (3) | 8 + 8 | 20-50 | [65] |
| Apparent absorption (%) from supplements, fortified foods and food folate | ||||||
| [13C6]-folic acid | 634 | 24% | 8 h (8) | 14 10 | 33 ± 2 | [18] |
| (6S)-[13C6]-5-HCO-H4folate | 500 | 38% | 8 h (8) | 14 10 | 33 ± 2 | [18] |
| Spinach [15N1-7]folate | 588 (M) | 44% | 8 h (8) | 14 10 | 33 ± 2 | [18] |
| Folic acid in bread 5 | 491 6 (M) | 74% | 10 h (7) | 1 + 8 | 51-79 | [44] |
| (6S)-5-CH3-H4folate in fermented milk 5 | 450 6 (M) | 86% | 10 h (7) | 1 + 8 | 51-79 | [44] |
| Yeast folate 5 | 155 6 (M) | 80% | 10 h (7) | 1 + 8 | 51-79 | [44] |
3.1. Intervention Trials
3.2. Short-Term Trials
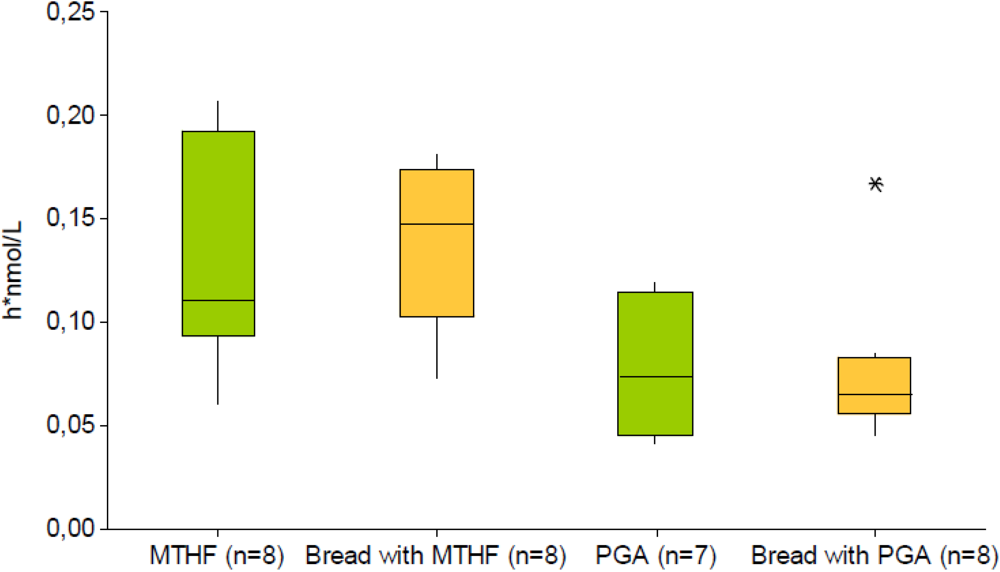
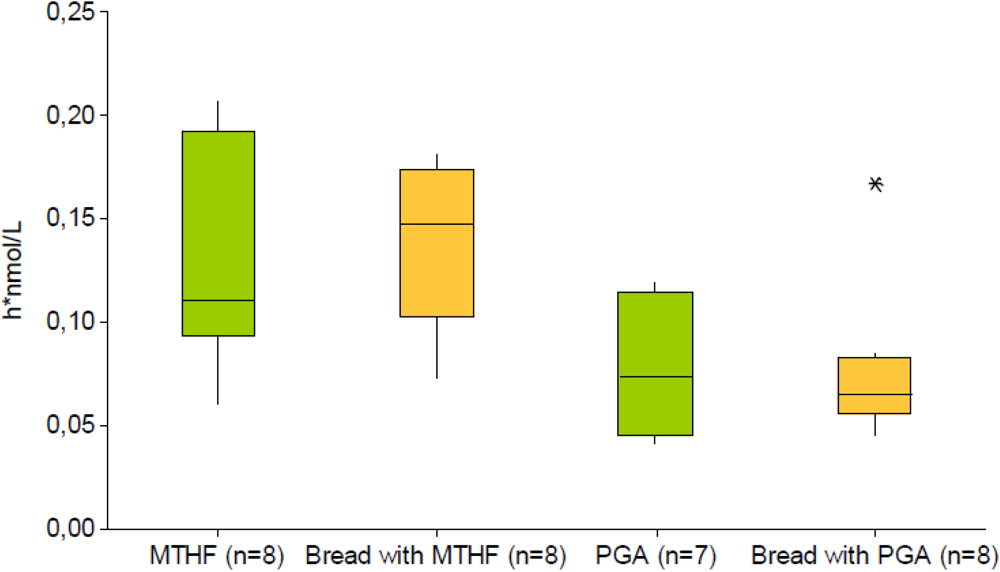
4. Conclusions
References
- Katan, M.B.; Boekschoten, M.V.; Connor, W.E.; Mensink, R.P.; Seidell, J.; Vessby, B.; Willett, W. Which Are the Greatest Recent Discoveries and the Greatest Future Challenges in Nutrition? Eur. J. Clin. Nutr. 2009, 63, 2–10. [Google Scholar]
- NNR, Nordic Nutrition Recommendations NNR 2004 Integrating Nutrition and Physical Activity; Nordic Council of Ministers: Stockholm, Sweden, 2005.
- Dhonukshe-Rutten, R.A.M.; de Vries, J.H.M.; de Bree, A.; van der Put, N.; van Staveren, W.A.; de Groot, L. Dietary Intake and Status of Folate and Vitamin B12 and Their Association with Homocysteine and Cardiovascular Disease in European Populations. Eur. J. Clin. Nutr. 2009, 63, 18–30. [Google Scholar]
- de Bree, A.; van Dusseldorp, M.; Brouwer, I.A.; van het Hof, K.H.; Steegers-Theunissen, R.P.M. Review Folate Intake in Europe: Recommended, Actual and Desired Intake. Eur. J. Clin. Nutr. 1997, 51, 643–660. [Google Scholar]
- Bergstrom, L. Nutrient Losses and Gains in the Preparation of Foods; National Food Administration: Sweden, Uppsala, 1994. [Google Scholar]
- Mitchell, L.E.; Adzick, N.S.; Melchionne, J.; Pasquariello, P.S.; Sutton, L.N.; Whitehead, A.S. Spina bifida. Lancet 2004, 364, 1885–1895. [Google Scholar]
- Gregory, J.F. Folate. In Food Chemistry; Fennema, O.R., Ed.; Marcel Dekker: New York, NY, USA, 1996; pp. 590–616. [Google Scholar]
- Mosley, B.S.; Cleves, M.A.; Siega-Riz, A.M.; Shaw, G.M.; Canfield, M.A.; Waller, D.K.; Werler, M.M.; Hobbs, C.A. Neural Tube Defects and Maternal Folate Intake among Pregnancies Conceived after Folic acid Fortification in the United States. Am. J. Epidemiol. 2009, 169, 9–17. [Google Scholar]
- Yang, Q.; Botto, L.D.; Erickson, J.D.; Berry, R.J.; Sambell, C.; Johansen, H.; Friedman, J.M. Improvement in Stroke Mortality in Canada and the United States, 1990 to 2002. Circulation 2006, 113, 1335–1343. [Google Scholar]
- Shirodaria, C.; Antoniades, C.; Lee, J.; Jackson, C.E.; Robson, M.D.; Francis, J.M.; Moat, S.J.; Ratnatunga, C.; Pillai, R.; Refsum, H.; Neubauer, S.; Channon, K.M. Global Improvement of Vascular Function and Redox State with Low-Dose Folic Acid. Circulation 2007, 115, 2262–2270. [Google Scholar]
- Sanderson, P.; McNulty, H.; Mastroiacovo, P.; McDowell, I.F.W.; Melse-Boonstra, A.; Finglas, P.M.; Gregory, J.F. Folate Bioavailability: UK Food Standards Agency Workshop Report. Br. J. Nutr. 2003, 90, 473–479. [Google Scholar]
- CDC. Water-Soluble Vitamins & Related Biochemical Compounds; Centers for Disease Control and Prevention: Atlanta, GA, USA. Available online: http://www.cdc.gov/nutritionreport/part_1.html (accessed on 13 July 2009).
- Hannon-Fletcher, M.P.; Armstrong, N.C.; Scott, J.M.; Pentieva, K.; Bradbury, I.; Ward, M.; Strain, J.J.; Dunn, A.A.; Molloy, A.M.; Kerr, M.A.; McNulty, H. Determining Bioavailability of Food Folates in a Controlled Intervention Study. Am. J. Clin. Nutr. 2004, 80, 911–918. [Google Scholar]
- Brouwer, I.A.; van Dusseldorp, M.; West, C.; Meyboom, S.; Thomas, C.M.G.; Duran, M.; van het Hof, K.H.; Eskes, T.K.A.B.; Hautvast, G.A.J.; Steegers-Theunissen, R.P.M. Dietary Folate from Vegetables and Citrus Fruit Decreases Plasma Homocysteine Concentrations in Humans in a Dietary Controlled trial. J. Nutr. 1999, 129, 1135–1139. [Google Scholar]
- Gregory, J.F. The Bioavailability of Folate. In Folate in Health and Disease; Bailey, L.B., Ed.; Marcel Dekker: New York, NY, USA, 1995; pp. 195–235. [Google Scholar]
- Melse-Boonstra, A.; Verhoef, P.; West, C. Quantifying Folate Bioavailability: A Critical Appraisal of Methods. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 539–545. [Google Scholar]
- Wright, A.J.A.; Finglas, P.M.; Dainty, J.R.; Hart, D.J.; Wolfe, C.A.; Southon, S.; Gregory, J.F. Single Oral Doses of C-13 Forms of Pteroylmonoglutamic Acid and 5-Formyltetrahydrofolic Acid Elicit Differences in Short-Term Kinetics of Labelled and Unlabelled Folates in Plasma: Potential Problems in Interpretation of Folate Bioavailability Studies. Br. J. Nutr. 2003, 90, 363–371. [Google Scholar]
- Wright, A.J.A.; Finglas, P.M.; Dainty, J.R.; Wolfe, C.A.; Hart, D.J.; Wright, D.M.; Gregory, J.F. Differential Kinetic Behavior and Distribution for Pteroylglutamic Acid and Reduced Folates: A Revised Hypothesis of the Primary Site of PteGlu Metabolism in Humans. J. Nutr. 2005, 135, 619–623. [Google Scholar]
- Wright, A.J.A.; King, M.J.; Wolfe, C.A.; Powers, H.J.; Finglas, P.M. Comparison of (6S)-5-methyltetrahydrofolic Acid v. Folic Acid as the Reference Folate in Longer-term Human Dietary Intervention Studies Assessing the Relative Bioavailability of Natural Food Folates: Comparative Changes in Folate Status Following a 16-week Placebo-controlled Study in Healthy Adults. Br. J. Nutr. 103, 724–729. [PubMed]
- Pfeiffer, C.M.; Fazili, Z.; Zhang, M. Folate Analytical Methodology. In Folate in Health and Disease, 2nd; Bailey, L.B., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 517–574. [Google Scholar]
- Finglas, P.M.; Scott, K.J.; Witthoft, C.M.; van den Berg, H.; de Froidmont-Gortz, I. The Certification of the Mass Fraction of Vitamins in Four Reference Materials: Wholemeal Flour (CRM 121), Milk powder (CRM 421), Lyophilised Mixed Vegetables (CRM 485) and Lyophilised Pig’s Liver (CRM 487); EUR-Report 18320; Office for Official Publications, Commission of the European Union: Luxembourg, Luxembourg, 1999. [Google Scholar]
- Vahteristo, L.T.; Finglas, P.M.; Witthoft, C.M.; Wigertz, K.; Seale, R.; De Froidmont-Gortz, I. Third EU MAT Intercomparison Study on Food Folate Analysis Using HPLC Procedures. Food Chem. 1996, 57, 109–111. [Google Scholar]
- Caudill, M.A. Folate Bioavailability: Implications for Establishing Dietary Recommendations and Optimizing status. Am. J. Clin. Nutr. 2010, 91, 1455S–1460S. [Google Scholar]
- Kim, Y.I. Folate and Colorectal Cancer: An Evidence-Based Critical Review. Mol. Nutr. Food Res. 2007, 51, 267–292. [Google Scholar]
- Mason, J.B.; Dickstein, A.; Jacques, P.F.; Haggarty, P.; Selhub, J.; Dallal, G.; Rosenberg, I.H. A Temporal Association between Folic Acid Fortification and an Increase in Colorectal Cancer Rates May Be Illuminating Important Biological Principles: A Hypothesis. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 1325–1329. [Google Scholar]
- Smith, A.D.; Kim, Y.-I.; Refsum, H. Is Folic Acid Good for Everyone? Am. J. Clin. Nutr. 2008, 87, 517–533. [Google Scholar] [PubMed]
- Halsted, C.H. testinal Absorption of Dietary Folates. In Folic Acid Metabolism in Health and Disease; Picciano, M.F., Gregory, J.F., Stokstad, E.L., Eds.; Wiley-Liss: New York, NY, USA, 1990; pp. 23–46. [Google Scholar]
- Zhao, R.; Matherly, L.H.; Goldman, I.D. Membrane Transporters and Folate Homeostasis: Intestinal Absorption and Transport into Systemic Compartments and Tissues. Expert Rev. Mol. Med. 2009, 11, e4. [Google Scholar] [CrossRef] [PubMed]
- Aufreiter, S.; Gregory, J.F., III; Pfeiffer, C.M.; Fazili, Z.; Kim, Y.-I.; Marcon, N.; Kamalaporn, P.; Pencharz, P.B.; O’Connor, D.L. Folate is Absorbed across the Colon of Adults: Evidence from Cecal Infusion of 13C-Labeled [6S]-5-formyltetrahydrofolic Acid. Am. J. Clin. Nutr. 2009, 90, 116–123. [Google Scholar] [PubMed]
- Camilo, E.; Zimmerman, J.; Mason, J.B.; Golner, B.; Russell, R.; Selhub, J.; Rosenberg, I.H. Folate Synthesized by Bacteria in the Human Upper Small Intestine is Assimilated by the Host. Gastroenterology 1996, 110, 991–998. [Google Scholar]
- Asrar, F.M.; O’Connor, D.L. Bacterially Synthesized Folate and Supplemental Folic Acid Are Absorbed across the Large Intestine of Piglets. J. Nutr. Biochem. 2005, 16, 587–593. [Google Scholar]
- Gregory, J.F.; Williamson, J.; Liao, J.F.; Bailey, L.B.; Toth, J.P. Kinetic Model of Folate Metabolism in Nonpregnant Women Consuming [H-2(2)]Folic Acid: Isotopic Labeling of Urinary Folate and the Catabolite Para-acetamidobenzoylglutamate Indicates Slow, Intake-Dependent, Turnover of Folate Pool. J. Nutr. 1998, 128, 1896–1906. [Google Scholar]
- Herbert, V. Recommended Dietary Intakes (RDI) of Folate in Humans. Am. J. Clin. Nutr. 1987, 45, 661–670. [Google Scholar]
- Lin, Y.; Dueker, S.R.; Follett, J.R.; Fadel, J.G.; Arjomand, A.; Schneider, P.D.; Miller, J.W.; Green, R.; Buchholz, B.A.; Vogel, J.S.; Phair, R.D.; Clifford, A.J. Quantitation ofin vivo Human Folate Metabolism. Am. J. Clin. Nutr. 2004, 80, 680–691. [Google Scholar]
- Wagner, C. Biochemical Role of Folate in Cellular Metabolism. In Folate in Health and Disease;Clinical Nutrition in Health and Disease 1; Bailey, L.B., Ed.; Marcel Dekker: New York, NY, USA, 1995; pp. 23–42. [Google Scholar]
- Tamura, T.; Picciano, M.F. Folate and Human Reproduction. Am. J. Clin. Nutr. 2006, 83, 993–1016. [Google Scholar]
- Reed, M.C.; Nijhout, H.F.; Neuhouser, M.L.; Gregory, J.E.; Shane, B.; James, S.J.; Boynton, A.; Ulrich, C.M. A Mathematical Model Gives Insights into Nutritional and Genetic Aspects of Folate-Mediated One-Carbon Metabolism. J. Nutr. 2006, 136, 2653–2661. [Google Scholar]
- Ohrvik, V. Folate Bioavailability. In-vitro Experiments and In-vivo Trials. Swedish University of Agricultural Sciences, Uppsala, Sweden, 2009. [Google Scholar]
- Gregory, J.F.; Caudill, M.A.; Opalko, F.J.; Bailey, L.B. Kinetics of Folate Turnover in Pregnant Women (Second Trimester) and Nonpregnant Controls during Folic Acid Supplementation: Stable-Isotopic Labeling of Plasma Folate, Urinary Folate and Folate Catabolites Shows Subtle Effects of Pregnancy on Turnover of Folate Pools. J. Nutr. 2001, 131, 1928–1937. [Google Scholar]
- Witthoft, C.; Stralsjo, L.; Berglund, G.; Lundin, E. A Human Model to Determine Folate Bioavailability from Food: A Pilot Study for Evaluation. Scand. J. Nutr. 2003, 47, 6–18. [Google Scholar]
- Pfeiffer, C.M.; Rogers, L.M.; Bailey, L.B.; Gregory, J.F. Absorption of Folate from Fortified Cereal-Grain Products and of Supplemental Folate Consumed with or Without Food Determined Using a Dual-Label Stable-Isotope Protocol. Am. J. Clin. Nutr. 1997, 66, 1388–1397. [Google Scholar]
- Rogers, L.M.; Pfeiffer, C.M.; Bailey, L.B.; Gregory, J.F. A Dual-Label Stable-Isotopic Protocol Is Suitable for Determination of Folate Bioavailability in Humans: Evaluation of Urinary Excretion and Plasma Folate Kinetics of Intravenous and Oral Doses of [C-13(5)] and [H-2(2)]Folic Acid. J. Nutr. 1997, 127, 2321–2327. [Google Scholar]
- Gregory, J.F.; Quinlivan, E.P. In vivo Kinetics of Folate Metabolism. Ann. Rev. Nutr. 2002, 22, 199–220. [Google Scholar]
- Witthoft, C.M.; Arkbage, K.; Johansson, M.; Lundin, E.; Berglund, G.; Zhang, J.X.; Lennernas, H.; Dainty, J.R. Folate Absorption from Folate-Fortified and Processed Foods Using a Human Ileostomy Model. Br. J. Nutr. 2006, 95, 181–187. [Google Scholar]
- Konings, E.J.M.; Troost, F.J.; Castenmiller, J.J.M.; Roomans, H.H.S.; van den Brandt, P.A.; Saris, W.H.M. Intestinal Absorption of Different Types of Folate in Healthy Subjects with an Ileostomy. Br. J. Nutr. 2002, 88, 235–242. [Google Scholar]
- Ohrvik, V.E.; Buttner, B.E.; Rychlik, M.; Lundin, E.; Witthoft, C.M. Folate Bioavailability from Breads and a Meal Assessed with a Human Stable-Isotope Area under the Curve and Ileostomy Model. Am. J. Clin. Nutr. 2010, 92, 532–538. [Google Scholar]
- Buttner, B.E.; Ohrvik, V.E.; Witthoft, C.M.; Rychlik, M. Quantification of Isotope-Labelled and Unlabelled Folates in Plasma, Ileostomy and Food Samples. Anal. Bioanal. Chem. 2011, 399, 429–439. [Google Scholar]
- Wilcken, B.; Bamforth, F.; Li, Z.; Zhu, H.; Ritvanen, A.; Redlund, M.; Stoll, C.; Alembik, Y.; Dott, B.; Czeizel, A.E.; et al. Geographical and Ethnic Variation of the 677C>T Allele of 5,10 Methylenetetrahydrofolate Reductase (MTHFR): Findings from over 7000 Newborns from 16 Areas World Wide. J. Med. Genet. 2003, 40, 619–625. [Google Scholar]
- Rozen, R. Folate and Genetics. J. Food Sci. 2004, 69, S65–S67. [Google Scholar]
- Gregory, J.F.; Quinlivan, E.P.; Davis, S.R. Integrating the Issues of Folate Bioavailability, Intake and Metabolism in the Era of Fortification. Trends Food Sci. Technol. 2005, 16, 229–240. [Google Scholar]
- Morin, I.; Devlin, A.M.; Leclerc, D.; Sabbaghian, N.; Halsted, C.H.; Finnell, R.; Rozen, R. Evaluation of Genetic Variants in the Reduced Folate Carrier and in Glutamate Carboxypeptidase II for Spina Bifida Risk. Mol. Genet. Metab. 2003, 79, 197–200. [Google Scholar]
- Tozer, T.; Rowland, M. Introduction to Pharmacokinetics and Pharmacodynamics; Lippincott Williams & Wilkins: Baltimore County, MD, USA, 2006. [Google Scholar]
- Gregory, J.F. Bioavailability of Folate. Eur. J. Clin. Nutr. 1997, 51, S54–S59. [Google Scholar]
- Bouckeart, K.P.; Slimani, N.; Nicolas, G.; Vignat, J.; Wright, A.J.A.; Roe, M.; Witthoft, C.M.; Finglas, P.M. Critical Evaluation of Folate Data in European and International Databases: Recommendations for Standardization in International Nutritional Studies. Mol. Nutr. Food Res. 2011, 55, 166–180. [Google Scholar]
- AACC International, Approved Methods of Analysis, 11th ed. Method 86-47.01. Total Folate in Cereal Products—Microbiological Assay Using Trienzyme Extraction; AACC International: St. Paul, MN, USA, 2000.
- DeVries, J.W.; Keagy, P.M.; Hudson, C.A.; Rader, J.I. AACC Collaborative Study of a Method for Determining Total Folate in Cereal Products-Microbiological Assay Using Trienzyme Extraction (AACC method 86-47). Cereal Food World 2001, 46, 216–219. [Google Scholar]
- Cho, S.; Choi, Y.; Lee, J.; Eitenmiller, R.R. Optimization of Enzyme Extractions for Total Folate in Cereals Using Response Surface Methodology. J. Agric. Food Chem. 2010, 58, 10781–10786. [Google Scholar]
- Riddell, L.J.; Chisholm, A.; Williams, S.; Mann, J.I. Dietary Strategies for Lowering Homocysteine Concentrations. Am. J. Clin. Nutr. 2000, 71, 1448–1454. [Google Scholar]
- Ohrvik, V.E.; Olsson, J.C.; Sundberg, B.E.; Witthoft, C.M. Effect of 2 pieces of nutritional advice on folate status in Swedish women: A randomized controlled trial. Am. J. Clin. Nutr. 2009, 89, 1053–1058. [Google Scholar]
- Vahteristo, L.; Kariluoto, S.; Barlund, S.; Karkkainen, M.; Lamberg-Allardt, C.; Salovaara, H.; Piironen, V. Functionality of Endogenous Folates from Rye and Orange Juice Using Human in vivo Model. Eur. J. Nutr. 2002, 41, 271–278. [Google Scholar]
- Winkels, R.M.; Brouwer, I.A.; Siebelink, E.; Katan, M.B.; Verhoef, P. Bioavailability of Food Folates is 80% of that of Folic Acid. Am. J. Clin. Nutr. 2007, 85, 465–473. [Google Scholar]
- Venn, B.J.; Mann, J.I.; Williams, S.M.; Riddell, L.J.; Chisholm, A.; Harper, M.J.; Aitken, W. Dietary Counseling to Increase Natural Folate Intake: A Randomized, Placebo-Controlled Trial in Free-living Subjects to Assess Effects on Serum Folate and Plasma Total Homocysteine. Am. J. Clin. Nutr. 2002, 76, 758–765. [Google Scholar]
- Fenech, M.; Noakes, M.; Clifton, P.; Topping, D. Aleurone Flour Increases Red-Cell Folate and Lowers Plasma Homocyst(E)Ine Substantially in Man. Br. J. Nutr. 2005, 93, 353–360. [Google Scholar]
- Pentieva, K.; McNulty, H.; Reichert, R.; Ward, M.; Strain, J.J.; McKillop, D.J.; McPartlin, J.M.; Connolly, E.; Molloy, A.; Kramer, K.; Scott, J.M. The Short-term Bioavailabilities of [6S]-5-Methyltetrahydrofolate and Folic Acid Are Equivalent in Men. J. Nutr. 2004, 134, 580–585. [Google Scholar]
- Fenech, M.; Noakes, M.; Clifton, P.; Topping, D. Aleurone Flour is a Rich Source of Bioavailable Folate in Humans. J. Nutr. 1999, 129, 1114–1119. [Google Scholar]
- Cuskelly, G.J.; McNulty, H.; Scott, J.M. Effect of Increasing Dietary Folate on Red-Cell Folate: Implications for Prevention of Neural Tube Defects. Lancet 1996, 347, 657–659. [Google Scholar]
- Broekmans, W.M.R.; Klopping-Ketelaars, I.A.A.; Schuurman, C.R.W.C.; Verhagen, H.; van den Berg, H.; Kok, F.J.; van Poppel, G. Fruits and Vegetables Increase Plasma Carotenoids and Vitamins and Decrease Homocysteine in Humans. J. Nutr. 2000, 130, 1578–1583. [Google Scholar]
- Bogers, R.P.; Dagnelie, P.C.; Bast, A.; van Leeuwen, M.; van Klaveren, J.D.; van den Brandt, P.A. Effect of Increased Vegetable and Fruit Consumption on Plasma Folate and Homocysteine Concentrations. Nutrition 2007, 23, 97–102. [Google Scholar]
- Ashfield-Watt, P.A.L.; Whiting, J.M.; Clark, Z.E.; Moat, S.J.; Newcombe, R.G.; Burr, M.L.; McDowell, I.F.W. A Comparison of the Effect of Advice to Eat Either “5-A-Day” Fruit and Vegetables or Folic Acid-Fortified Foods on Plasma Folate and Homocysteine. Eur. J. Clin. Nutr. 2003, 57, 316–323. [Google Scholar]
- Prinz-Langenohl, R.; Bronstrup, A.; Thorand, B.; Hages, M.; Pietrzik, K. Availability of Food Folate in Humans. J. Nutr. 1999, 129, 913–916. [Google Scholar]
- Enghardt-Barbieri, H.; Lindvall, C. Swedish Nutrition Recommendations Objectified (SNO)-Basis for General Advice on Food Consumption for Healthy Adults; National Food Administration: Sweden, Uppsala, 2005. [Google Scholar]
- Summers, C.M.; Mitchell, L.E.; Stanislawska-Sachadyn, A.; Baido, S.F.; Blair, I.A.; Von Feldt, J.M.; Whitehead, A.S. Genetic and Lifestyle Variables Associated with Homocysteine Concentrations and the Distribution of Folates Derivatives in Healthy Premenopausal Women. Birth Defects Res. A 2010, 88, 679–688. [Google Scholar]
- McKinley, M.C.; Strain, J.J.; McPartlin, J.; Scott, J.M.; McNulty, H. Plasma Homocysteine Is Not Subject to Seasonal Variation. Clin. Chem. 2001, 47, 1430–1436. [Google Scholar]
- Clarke, R.; Woodhouse, P.; Ulvik, A.; Frost, C.; Sherliker, P.; Refsum, H.; Ueland, P.M.; Khaw, K.-T. Variability and determinants of Total Homocysteine Concentrations in Plasma in an Elderly Population. Clin. Chem. 1998, 44, 102–107. [Google Scholar]
- Garg, U.C.; Zheng, Z.-J.; Folsom, A.R.; Moyer, Y.S.; Tsai, M.Y.; McGovern, P.; Eckfeldt, J.H. Short-Term and Long-Term Variability of Plasma Homocysteine Measurement. Clin. Chem. 1997, 43, 141–145. [Google Scholar]
- Antoniades, C.; Shirodaria, C.; Leeson, P.; Baarholm, O.A.; Van-Assche, T.; Stefanadis, C.; Refsum, H.; Channon, K.M. MTHFR 677 C>T Polymorphism Reveals Functional Importance for Methyltetrahydrofolate, Not Homocysteine, in Regulation of Vascular Redox State and Endothelial Function in Human Atherosclerosi. Circulation 2009, 119, 2507–2515. [Google Scholar]
- Tamura, T.; Stokstad, E.L. Availability of Food Folate in Man. Br. J. Haematol. 1973, 25, 513–532. [Google Scholar]
- Achon, M.; Arrate, A.; Alonso-Aperte, E.; Varela-Moreiras, G. Plasma Folate Concentrations after a Single Dose Ingestion of Whole and Skimmed Folic Acid Fortified Milks in Healthy Subjects. Eur. J. Nutr. 2011, 50, 119–125. [Google Scholar]
- Colman, N.; Green, R.; Metz, J. Prevention of Folate Deficiency by Food Fortification. II. Absorption of Folic Acid from Fortified Staple Foods. Am. J. Clin. Nutr. 1975, 28, 459–464. [Google Scholar] [PubMed]
- Finglas, P.M.; Witthoft, C.M.; Vahteristo, L.; Wright, A.J.A.; Southon, S.; Mellon, F.A.; Ridge, B.; Maunder, P. Use of an Oral/Intravenous Dual-Label Stable-Isotope Protocol to Determine Folic Acid Bioavailability from Fortified Cereal Grain Foods in Women. J. Nutr. 2002, 132, 936–939. [Google Scholar]
- Wei, M.M.; Bailey, L.B.; Toth, J.P.; Gregory, J.F. Bioavailability for Humans of Deuterium-Labeled Monoglutamyl and Polyglutamyl Folates Is Affected by Selected Foods. J. Nutr. 1996, 126, 3100–3108. [Google Scholar]
- Gregory, J.F.; Bhandari, S.D.; Bailey, L.B.; Toth, J.P.; Baumgartner, T.G.; Cerda, J.J. Relative Bioavailability of Deuterium-Labeled Monoglutamyl and Hexaglutamyl Folates in Human-Subjects. Am. J. Clin. Nutr. 1991, 53, 736–740. [Google Scholar]
- Buchholz, B.A.; Arjomand, A.; Dueker, S.R.; Schneider, P.D.; Clifford, A.J.; Vogel, J.S. Intrinsic Erythrocyte Labeling and Attomole Pharmacokinetic Tracing of 14C-Labeled Folic Acid with Accelerator Mass Spectrometry. Anal. Biochem. 1999, 269, 348–352. [Google Scholar]
- Wigertz, K. Milk Folates-Characterisation and Availability. Lund University, Lund, Sweden, 1997. [Google Scholar]
- Ashokkumar, B.; Mohammed, Z.M.; Vaziri, N.D.; Said, H.M. Effect of Folate Oversupplementation on Folate Uptake by Human Intestinal and Renal Epithelial Cells. Am. J. Clin. Nutr. 2007, 86, 159–166. [Google Scholar]
- Ohrvik, V.E.; Buttner, B.E.; Rychlik, M.; Lundin, E.; Witthoft, C.M. The Bioavailability of Folic Acid and Reduced Folate Bioavailability from Foods and Pharmaceutical Preparations Assessed With a Human Stable-Isotope Area under the Curve and Ileostomy Model. 2011, unpublished work. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ohrvik, V.E.; Witthoft, C.M. Human Folate Bioavailability. Nutrients 2011, 3, 475-490. https://doi.org/10.3390/nu3040475
Ohrvik VE, Witthoft CM. Human Folate Bioavailability. Nutrients. 2011; 3(4):475-490. https://doi.org/10.3390/nu3040475
Chicago/Turabian StyleOhrvik, Veronica E., and Cornelia M. Witthoft. 2011. "Human Folate Bioavailability" Nutrients 3, no. 4: 475-490. https://doi.org/10.3390/nu3040475
APA StyleOhrvik, V. E., & Witthoft, C. M. (2011). Human Folate Bioavailability. Nutrients, 3(4), 475-490. https://doi.org/10.3390/nu3040475