Effects of Beta-Alanine on Muscle Carnosine and Exercise Performance: A Review of the Current Literature


Abstract
:1. Introduction
2. Carnosine

3. Beta-Alanine
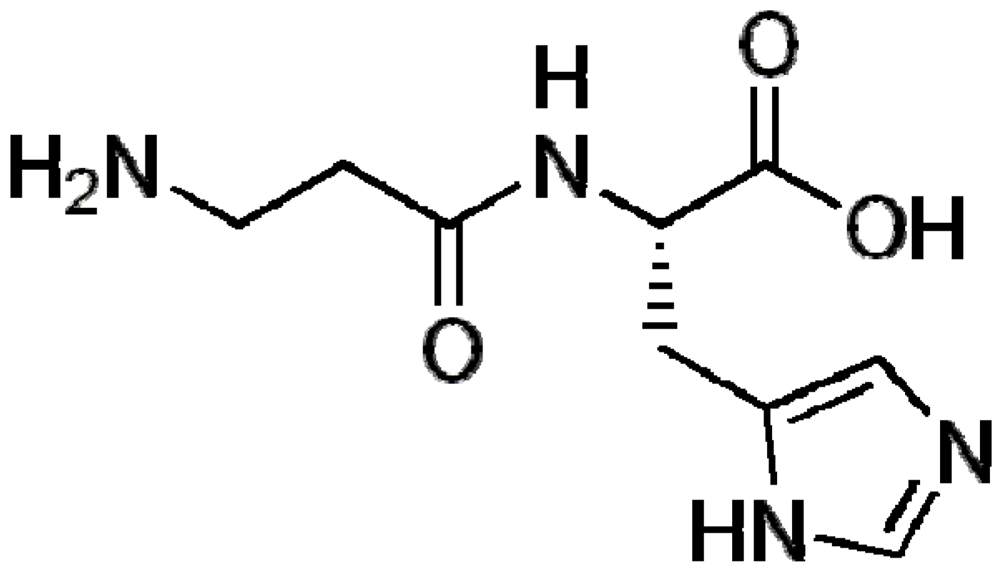
3.1. Beta-Alanine and Carnosine

{kind=link}
| Authors | Population | Supplementation Protocol | Muscle Carnosine Concentration Effects | Performance Results |
|---|---|---|---|---|
| Baguet et al., 2009 [56] | 20 physically active males | 5-6 weeks of β-ALA or placebo (maltodextrin) | In soleus, carnosine increased 30% (p=0.003) with β-ALA and remained stable with placebo (p=0.867) | None measured |
| 2.4 g/day - first 2 days | In tibialis anterior, carnosine increased 27% (p=0.005) with β-ALA and decreased 17% (p=0.05) with placebo | |||
| 3.6 g/day - days 3-4 | In gastrocnemius, carnosine increased 23% (p=0.038) and did not change with placebo (p=0.740). | |||
| 4.8 g/day to end of study | Carnosine elimination was measured at 3 and 9 weeks after supplementation period | |||
| At 3 weeks, only 26.1% (in the soleus), 20.1% (in tibialis anterior) and 44.7% (in the gastrocnemius) of the increase had disappeared. There was no difference between β-ALA and placebo at this point (p=0.431) | ||||
| At 9 weeks, carnosine levels in all 3 muscles returned to initial values | ||||
| Harris et al., 2006 [7] | Study 3:21 physically active males Ages 26.1 ± 5.6 yrs | 4 weeks, 4 groups (I - IV): I) 800mg β-ALA x 4 daily (avg. 3.2g daily and 89.6g 4wk total)II) 8 daily doses of either 400 or 800mg β-ALA (avg. 6.4g daily and 145.6g 4wk total)III) 8 daily doses of 1000 or 2000 mg L-carnosine (364g 4wk total L-carnosine, corresponding to 143.3g β-ALA) IV) Placebo of maltodextrin at doses to match groups II and III | Increase in carnosine concentration greatest with carnosine supplementation, followed by group II, then group II β-ALA protocols. | None measured |
| Mean increase over 4 weeks (mmol·kg-1dm) | ||||
| I) 7.80 ± .36 (p < .05) | ||||
| II) 11.04 ± 2.68 (p < .05) | ||||
| III) 16.37 ± 3.03 (p < .05) | ||||
| IV) 1.87 ± 1.73 (p>.05) | ||||
| Derave et al., 2007 [5] | 15 male track athletes (sprinters) 18-24 yrs | 4-5 weeks β-ALA or placebo (maltodextrin) | Soleus: | No difference between groups for 400m running performance |
| 2.4g/day - first 4 days | ↑ 47% with β-ALA | |||
| 3.6g/day - days 5-8 | No change with placebo | |||
| 4.8g/day to end of study | Gastrocnemius: | |||
| ↑ 37% with β-ALA | ||||
| No change with placebo | ||||
| Hill et al., 2007 [6] | 25 physically active males | 10 weeks β-ALA: | β-ALA group, ↑ from 19.0 to 30.1 mmol/kg (58.8%) at 4 weeks and up to 34.7 mmol/kg (80.1%) at 10 weeks | No effect on body mass |
| 4g/day - wk 1 | Not significant between weeks 4 and 10 | ↑cycling capacity time at 110% with β-ALA | ||
| 4.8g/day - wk 2 | ||||
| 5.6g/day - wk 3 | ||||
| 6.4g/day - wk 4-10 |
3.2. Beta-Alanine and Exercise Performance
| Authors | Population | Supplementation Protocol | Exercise Testing Protocol | Performance Results |
|---|---|---|---|---|
| Baguet et al., 2009 [60] | 14 physically active males | 4 weeks of β-ALA or placebo (maltodextrin) | Maximal ramp exercise test on cycle ergometer to determine VO2peak, VT and gas exchange threshold | Exercise-induced acidosis was 19% lower with β-ALA |
| 2.4 g/day - first 2 days | Pre and Post supplementation: 3 x 6min cycle exercise bouts at 50% ∆ power output | No difference in VO2 throughout exercise before or after supplementation in either group | ||
| 3.6 g/day - days 3-4 | Time delay in the fast component was significantly shorter with β-ALA than placebo | |||
| 4.8 g/day to end of study | Does not support a role for acidosis in O2 deficit or the slow component of VO2 kinetics | |||
| Stout et al., 2006 [4] | 51 males | 4 groups: | PWCMFT test with EMG measurements on a cycle ergometer | β-ALA may delay the onset of neuromuscular fatigue, but no additive effects of creatine |
| Placebo - 34 g dextrose | Significant increase in PWCFT with β-ALA (14.5%) and creatine plus β-ALA (11%) compared to placebo | |||
| Creatine - 5.25 g creatine monohydrate and 34 g dextrose | ||||
| β-ALA - 1.6 g β-ALA plus 34 g dextrose | ||||
| β-ALA+Creatine - 5.35 g creatine monohydrate, 1.6 g β-ALA and 34 g dextrose | ||||
| 28 days of supplementation: | ||||
| 4 doses/day - days 1-6 | ||||
| 2 doses/day - days 7-28 | ||||
| Stout et al., 2007 [8] | 22 females | 4 weeks β-ALA or placebo | Continuous graded exercise test on cycle ergometer for VO2max, ventilatory threshold , PWCFT and TTE | β-ALA delays onset of NMF during incremental cycle ergometry (↑ PWCFT, ↑VT, ↑TTE) |
| Ages: | 4 divided doses/day for totals of: | |||
| 28.9 ± 8.1 yrs (β-ALA) | 3.2g/day - days 1-7 | |||
| 25.8 ± 4.0 yrs (placebo) | 6.4g/day - days 8-28 | |||
| Stout et al., 2008 [61] | 26 elderly males and females | 90 days supplementation with β-ALA or placebo (microcrystalline cellulose) 3 doses/day of: 2.4 g β-ALA or 2.4 g placebo | Continuous graded exercise test on cycle ergometer for PWCFT with EMG measurements | 28,5% increase in PWCFT after 90 days of β-ALA |
| Sweeney et al., 2009 [62] | 19 physically active college-aged males | 5 weeks β-ALA or placebo (rice flour) | 2 sets of 5x5-sec sprints with 45- sec recovery between sprints and 2 min between sets performed on non-motorized treadmill at 15% body weight as resistance | No between group difference for peak or mean horizontal power |
| 4g/day - week 1 | No difference in % fatigue | |||
| 6g/day - weeks 2-5 | No difference in blood lactate pre- and post-testing between groups | |||
| Van Thienen et al., 2009 [57] | 17 healthy young males | 8 weeks β-ALA or placebo (maltodextrin) | Simulated road race of 110 minutes intermittent endurance with intensity between 50% and 90% of the maximal lactate steady state (MLSS) in 10 minute stages. Immediately after this, they started a 10 minute time trial at 100% MLSS with voluntary increase of intensity at each minute. | β-ALA enhanced sprint power output at the end of the endurance race compared to placebo |
| 2 g/day - weeks 1-2 | ||||
| 3 g/day - weeks 3-4 | ||||
| 4 g/day - weeks 5-8 | ||||
| Zoeller et al., 2007 [10] | 55 males ages 24.5 ± 5.3 yrs | 4 weeks, 4 groups (4 doses/day for first 6 days, then 2 doses/day | Continuous graded exercise test on cycle ergometer | ↑ in 5 cardio-respiratory endurance variables with creatine + β-ALA |
| Placebo - 34g dextrose | Combined supplementation may delay the onset of VT and lactate threshold during incremental cycle exercise | |||
| Creatine - 5.25g creatine monohydrate and 34g dextrose | ||||
| β-ALA - 1.6g β-alanine and 34g dextrose | ||||
| β-ALA plus Creatine - 5.25g creatine monohydrate, 1.6g β-ALA and 34g dextrose |
3.3. Beta-Alanine and Exercise Training
| Authors | Population | Supplementation Protocol | Exercise Protocol | Muscle Carnosine Concentration Effects | Performance Results |
|---|---|---|---|---|---|
| Hoffman et al., 2006 [12] | 33 male strength power athletes | 10 weeks | Resistance training program 4 days/week for 10 weeks | Not measured | ↓ fatigue rate in CA |
| Creatine β-ALA (CA) - 10.5g/day creatine monohydrate and 3.2g/day β-ALA | ↑ ∆ lean body mass and % body fat | ||||
| Creatine (C) - 10.5g/day | No change in power measures | ||||
| Placebo (P) - 10.5g/day dextrose | ↑ training volume in CA | ||||
| Kendrick et al., 2008 [9] | 26 healthy males, 19-24 yrs | 800mg x 8/day for 4 weeks of β-ALA or placebo (maltodextrin) | Resistance training 4days/wk for 10 weeks | β-ALA - 23.96± 5.94 to 36.77± 8.26 (p < 0.0001) | No difference in whole body strength or isokinetic force |
| Placebo - 29.17± 9.82 to 27.29± 9.52 (p > 0.05) | |||||
| Kendrick et al., 2009 [65] | 14 Vietnamese college aged students | 4 weeks β-ALA or placebo (maltodextrin) 800mg x 8/day | Single legged isokinetic training | Carnosine ↑ in both trained and untrained legs with β-ALA | None measured. |
| 3 sessions - weeks 1-2 | Training alone had no effect on carnosine levels | ||||
| 4 sessions - weeks 3-4 | |||||
| 10 × 10 maximal 90° extension and flexion contractions at 180°/sec on Kin-Com | |||||
| Smith et al., 2009 [59] | 46 recreationally active young males | 6g/day for 3 weeks, then 3g/day for 2nd 3 weeks of β-ALA or placebo (dextrose) | High intensity interval training | Not measured | Training increased EMGFT, no additive effect with β-ALA |
| Smith et al., 2009 [58] | 46 recreationally active young males | 6g/day for 3 weeks, then 3g/day for 2nd 3 weeks of β-ALA or placebo (dextrose) | High intensity interval training | Not measured | ↑ VO2peak and time to reach VO2peak with β-ALA |
| ↑ lean body mass with β-ALA |
3.4. Beta-Alanine and Muscular Fatigue
3.5. Beta-Alanine and Creatine Supplementation
3.6. Summary of Beta-Alanine Supplementation
4. Future Directions
5. Conclusion
References
- Hultman, E.; Sahlin, K. Acid-base balance during exercise. Exerc. Sports Sci. Rev. 1980, 8, 41–128. [Google Scholar]
- MacClaren, D.P.; Gibson, H.; Parry-Billings, M.; Edwards, R.H.T. A review of metabolic and physiological factors in fatigue. Exerc. Sports Sci. Rev. 1989, 17, 29–68. [Google Scholar]
- Taylor, A.D.; Bronks, R.; Bryant, A.L. The relationship between electromyography and work intensity revisited: A brief review with references to lactic acidosis and hyperammonia. Electromyo. Clin. Neurophysiol. 1997, 37, 387–398. [Google Scholar]
- Stout, J.R.; Cramer, J.T.; Mielke, M.; O'Kroy, J.A.; Torok, D.; Zoeller, R.F. Effects of twenty-eight days of beta-alanine and creatine monohydrate supplementation on the physical working capacity at neuromuscular fatigue threshold. J. Strength Cond. Res. 2006, 20, 928–931. [Google Scholar] [PubMed]
- Derave, W.; Ozdemir, M.S.; Harris, R.C.; Pottier, A.; Reyngoudt, H.; Koppo, K.; Wise, J.A.; Achten, E. Beta-alanine supplementation augments muscle carnosine content and attenuates fatigue during repeated isokinetic contraction bouts in trained sprinters. J. Appl. Physiol. 2007, 103, 1736–1743. [Google Scholar]
- Hill, C.A.; Harris, R.C.; Kim, H.J.; Harris, B.D.; Sale, C.; Boobis, L.H.; Kim, C.K.; Wise, J.A. Influence of beta-alanine supplementation on skeletal muscle carnosine concentrations and high intensity cycling capacity. Amino Acids 2007, 32, 225–233. [Google Scholar]
- Harris, R.C.; Tallon, M.J.; Dunnett, M.; Boobis, L.; Coakley, J.; Kim, H.J.; Fallowfield, J.L.; Hill, C.A.; Sale, C.; Wise, J.A. The absorption of orally supplied beta-alanine and its effect on muscle carnosine synthesis in human vastus lateralis. Amino Acids 2006, 30, 279–289. [Google Scholar]
- Stout, J.R.; Cramer, J.T.; Zoeller, R.F.; Torok, D.; Costa, P.; Hoffman, J.R.; Harris, R.C.; O'Kroy, J. Effects of beta-alanine supplementation on the onset of neuromuscular fatigue and ventilatory threshold in women. Amino Acids 2007, 32, 381–386. [Google Scholar]
- Kendrick, I.; Harris, R.; Kim, J.J.; Kim, C.; Dang, V.; Lam, T.; Bui, T.; Smith, M.; Wise, J. The effects of 10 weeks of resistance training combined with beta-alanine supplementation on whole body strength, force production, muscular endurance and body composition. Amino Acids 2008, 34, 546–554. [Google Scholar]
- Zoeller, R.F.; Stout, J.R.; O'Kroy, J.A.; Torok, D.J.; Mielke, M. Effects of 28 days of beta-alanine and creatine monohydrate supplementation on aerobic power, ventilatory and lactate thresholds, and time to exhaustion. Amino Acids 2007, 33, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Kreider, R.B. Effects of creatine supplementation on performance and training adaptations. Mol. Cell. Biochem. 2003, 244, 89–94. [Google Scholar]
- Hoffman, J.; Ratamess, N.; Kang, J.; Mangine, G.; Faigenbaum, A.; Stout, J. Effect of creatine and beta-alanine supplementation on performance and endocrine responses in strength/power athletes. Int. J. Sport Nutr. Exerc. Metab. 2006, 16, 430–446. [Google Scholar]
- Davey, C. The significance of carnosine and anserine in striated skeletal muscle. Arch. Biochem. Biophysiol. 1960, 89, 303–308. [Google Scholar]
- Dunnett, M.; Harris, R. Influence of oral beta-alanine and L-histidine supplementation on the carnosine content of the gluteus medius. Equine Vet. J. 1999, 30, 499–504. [Google Scholar]
- Chasovnikova, L.V.; Formazuyk, V.E.; Sergienko, V.I.; Boldyrev, A.A.; Severin, S.E. The anti-oxidative propertie of carnosine and other drugs. Biochem. Int. 1990, 20, 1097–1103. [Google Scholar]
- MacFarlane, N.; McMurray, J.; O'Dowd, J.J.; Dargie, H.J.; Miller, D.J. Synergism of histidyl dipeptides as antioxidants. J. Molec. Cell. Cardiol. 1991, 23, 1205–1207. [Google Scholar]
- Johnson, P.; Aldstadt, J. Effects of carnosine and anserine on muscle and non-muscle phosphorylases. Comp. Biochem. Physiol. B 1984, 78, 331–333. [Google Scholar]
- Batrukova, M.A.; Rubtsov, A.M. Histidine-containing dipeptides as endogenous regulators of the activity of sarcoplasmic reticulum Ca-release channels. BBA Biomembranes 1997, 1324, 142–150. [Google Scholar]
- Boldyrev, A.A.; Severin, S.E. The histidine-containing dipeptides, carnosine and anserine: distribution, properties and biological significance. Adv. Enzyme Reg. 1990, 30, 175–194. [Google Scholar] [CrossRef]
- Quinn, P.J.; Boldyrev, A.A.; Formazuyk, V.E. Carnosine: its properties, functions and potential therapeutic applications. Mol. Aspects Med. 1992, 13, 379–444. [Google Scholar]
- Harris, R.C.; Marlin, D.J.; Dunnett, M.; Snow, D.H.; Hultman, E. Muscle buffering capacity and dipeptide content in the thoroughbred horse, greyhound dog and man. Comp. Biochem. Physiol. A Comp. Physiol. 1990, 97, 249–251. [Google Scholar]
- Suyama, Y.; Suzuki, T.; Maruyama, M.; Saito, K. Determination of carnosine, anserine and beta-alanine in the muscle of animals. Bull. Jpn. Soc. Sci. Fish 1970, 36, 1048–1053. [Google Scholar]
- Parkhouse, W.S.; McKenzie, D.C.; Hochachka, P.W.; Ovalle, W.K. Buffering capacity of deproteinized human vastus lateralis muscle. J. Appl. Physiol. 1985, 58, 14–17. [Google Scholar]
- Hipkiss, A.; Brownson, C.; Bertani, M.; Ruiz, E.; Ferro, A. Reaction of carnosine with aged proteins: another protective process? Ann. N.Y. Acad. Sci. 2002, 959, 285–294. [Google Scholar] [PubMed]
- Tallon, M.J.; Harris, R.C.; Boobis, L.H.; Fallowfield, J.L.; Wise, J.A. The carnosine content of vastus lateralis is elevated in resistance-trained bodybuilders. J. Strength Cond. Res. 2005, 19, 725–729. [Google Scholar]
- Gardner, M.L.; Illingworth, K.M.; Kelleher, J.; Wood, D. Intestinal absorption of the intact peptide carnosine in man, and comparison with intestinal permeability to lactulose. J. Physiol. 1991, 439, 411–422. [Google Scholar]
- Suzuki, T.; Ito, O.; Takahashi, H.; Takamatsu, K. The effect of sprint training on skeletal muscle carnosine in humans. Int. J. Sport Health Sci. 2004, 2, 105–110. [Google Scholar]
- Skulachev, V.P. Biological role of carnosine in the functioning of excitable tissues.Centenary of Gulewitsch's discovery. Biochem. (Mosc) 2000, 65, 749–750. [Google Scholar]
- Mannion, A.F.; Jakeman, P.M.; Dunnett, M.; Harris, R.C.; Willan, P.L. Carnosine and anserine concentrations in the quadriceps femoris muscle of healthy humans. Eur. J. Appl. Physiol. Occup. Physiol. 1992, 64, 47–50. [Google Scholar]
- Dunnett, M.; Harris, R.C.; Soliman, M.Z.; Suwar, A.A. Carnosine, anserine and taurine contents in individual fibres from the middle gluteal muscle of the camel. Res. Vet. Sci. 1997, 62, 213–216. [Google Scholar]
- Suzuki, Y.; Ito, O.; Mukai, N.; Takahashi, H.; Takamatsu, K. High level of skeletal muscle carnosine contributes to the latter half of exercise performance during 30-s maximal cycle ergometer sprinting. Jpn. J. Physiol. 2002, 52, 199–205. [Google Scholar]
- Damon, B.; Hsu, A.; Stark, H.; Dawson, M. The carnosine C-2 proton's chemical shift reports intracellular pH in oxidative and glycolytic muscle fibers. Magn. Reson. Med. 2003, 49, 233–240. [Google Scholar]
- Beaver, W.L.; Wasserman, K.; Whipp, B.J. Bicarbonate buffering of lactic acid generated during exercise. J. Appl. Physiol. 1986, 60, 472–478. [Google Scholar]
- Abe, H. Role of histidine-related compounds as intracellular proton buffering constituents in vertebrate muscle. Biochemistry 2000, 65, 757–765. [Google Scholar]
- Bump, K.; Lawrence, L.; Moser, L.; Miller-Graber, P.; Kurcz, E. Effect of breed of horse on muscle carnosine concentration. Comp. Biochem. Physiol. 1990, 195–197. [Google Scholar]
- Sewell, D.A.; Harris, R.C.; Marlin, D.J.; Dunnett, M. Estimation of the carnosine content of different fibre types in the middle gluteal muscle of the thoroughbred horse. J. Physiol. 1992, 455, 447–453. [Google Scholar]
- Kraemer, W.J.; Gordon, S.E.; Lynch, J.M.; Pop, M.E.; Clark, K.L. Effects of multibuffer supplementation on acid-base balance and 2,3-diphosphoglycerate following repetitive anaerobic exercise. Int. J. Sport Nutr. 1995, 5, 300–314. [Google Scholar]
- Begum, G.; Cunliffe, A.; Leveritt, M. Physiological role of carnosine in contracting muscle. Int.l J. Sport Nutr. Exerc. Metabol. 2005, 15, 493–514. [Google Scholar]
- Tamaki, N.; Tsunemori, F.; Wakabayashi, M.; Hama, T. Effect of histidine-free and -excess diets on anserine and carnosine contents in rat gastrocnemius muscle. J. Nutr. Sci. Vitaminol. (Tokyo) 1977, 23, 331–340. [Google Scholar] [PubMed]
- Severin, S.E.; Boldyrev, A.A.; Stvolinskii, S.L.; Bordiukov, M.M.; Goncharenko, E.N.; Deev, L.I.; Malinina, I.E.; Kudriashov Iu, B. The radiation modifying properties of carnosine. Radiobiologiia 1990, 30, 765–768. [Google Scholar] [PubMed]
- Lamb, G.D.; Recupero, E.; Stephenson, D.G. Effect of myoplasmic pH on excitation-contraction coupling in skeletal muscle fibres of the toad. J. Physiol. 1992, 448, 211–224. [Google Scholar]
- Suzuki, Y.; Nakao, T.; Maemura, H.; Sato, M.; Kamahara, K.; Morimatsu, F.; Takamatsu, K. Carnosine and anserine ingestion enhances contribution of nonbicarbonate buffering. Med. Sci. Sports Exerc. 2006, 38, 334–338. [Google Scholar]
- Tamaki, N.; Funatsuka, A.; Fujimoto, S.; Hama, T. The utilization of carnosine in rats fed on a histidine-free diet and its effect on the levels of tissue histidine and carnosine. J. Nutr. Sci. Vitaminol. 1984, 30, 541–551. [Google Scholar]
- Boldyrev, A.A.; Dupin, A.; Ya Bunin, A.; Babizhaev, M.; Severin, S. The antioxidative properties of carnosine, a natural histidine containing dipeptide. Biochem. Int. 1987, 15, 1105–1113. [Google Scholar]
- Mastaloudis, A.; Leonard, S.W.; Traber, M.G. Oxidative stress in athletes during extreme endurance exercise. Free Radic. Biol. Med. 2001, 31, 911–922. [Google Scholar]
- Ebbeling, C.B.; Clarkson, P.M. Exercise-induced muscle damage and adaptation. Sports Med. 1989, 7, 207–234. [Google Scholar]
- Venditti, P.; DiMeo, S. Effect of training on antioxidant capacity, tissue damage, and endurance of adult male rat. Int. J. Sports Med. 1997, 18, 497–502. [Google Scholar]
- Bailey, D.M.; Davies, B.; Young, I.S.; Hullin, D.A.; Seddon, P.S. A potential role for free radical mediated skeletal muscle soreness in the pathophysiology of acute mountain sickness. Aviat. Space Environ. Med. 2001, 72, 513–521. [Google Scholar]
- Parker, C.J.J.; Ring, E. A comparative study of the effect of carnosine on myofibrilar-ATPase activity on vertebrate and invertebrate muscles. Comp. Biochem. Physiol. 1970, 37, 413–419. [Google Scholar]
- Lamont, C.; Miller, D.J. Calcium sensitizing action of carnosine and other endogenous imidaxoles in chemically skinned striated muscle. J. Physiol. 1992, 454, 421–434. [Google Scholar]
- Dutka, T.L.; Lamb, G.D. Effect of carnosine on excitation-contraction coupling in mechanically-skinned rat skeletal muscle. J. Muscle Res. Cell Motil. 2004, 25, 203–213. [Google Scholar]
- Bakardijiev, A.; Bauer, K. Transport of beta-alanine and biosynthesis of carnosine by skeletal muscle cells in primary culture. Eur. J. Biochem. 1994, 225, 617–623. [Google Scholar]
- Bauer, K.; Schulz, M. Biosynthesis of carnosine and related peptides by skeletal muscle cells in primary culture. Eur. J. Biochem. 1994, 219, 43–47. [Google Scholar]
- Guiotto, A.; Calderan, A.; Ruzza, P.; Borin, G. Carnosine and carnosine-related antioxidants: a review. Curr. Med. Chem. 2005, 12, 2293–2315. [Google Scholar]
- Kendrick, I.P.; Harris, R.C.; Kim, C.K.; Kim, H.J.; Viet, D.H.; Thanh, L.Q.; Toai, B.T.; Wise, J.A. The effect of beta-alanine (Carnosyn) supplementation on muscle carnosine synthesis during 4 weeks using a one-leg training model (Abstract). J. Int. Soc. Sports Nutr. 2006, 3, S8. [Google Scholar]
- Baguet, A.; Reyngoudt, H.; Pottier, A.; Everaert, I.; Callens, S.; Achten, E.; Derave, W. Carnosine loading and washout in human skeletal muscles. J. Appl. Physiol. 2009, 106, 837–842. [Google Scholar]
- Van Thienen, R.; Van Proeyen, K.; Vanden Eynde, B.; Puypo, J.; Lefere, T.; Hespel, P. Beta-alanine improves sprint performance in endurance cycling. Med. Sci. Sports Exerc. 2009, 41, 898–903. [Google Scholar]
- Smith, A.E.; Walter, A.A.; Graef, J.L.; Kendall, K.L.; Moon, J.R.; Lockwood, C.M.; Fukuda, D.H.; Beck, T.W.; Cramer, J.T.; Stout, J.R. ffects of beta-alanine supplementation and high-intensity interval training on endurance performance and body composition in men; a double-blind trial. J. Int. Soc. Sports Nutr. 2009, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.E.; Moon, J.R.; Kendall, K.L.; Graef, J.L.; Lockwood, C.M.; Walter, A.A.; Beck, T.W.; Cramer, J.T.; Stout, J.R. The effects of beta-alanine supplementation and high-intensity interval training on neuromuscular fatigue and muscle function. Eur. J. Appl. Physiol. 2009, 105, 357–363. [Google Scholar]
- Baguet, A.; Koppo, K.; Pottier, A.; Derave, W. Beta-alanine supplementation reduces acidosis but not oxygen uptake response during high-intensity cycling exercise. Eur. J. Appl. Physiol. 2009, October 16. Epub ahead of print. [Google Scholar]
- Stout, J.R.; Graves, B.S.; Smith, A.E.; Hartman, M.J.; Cramer, J.T.; Beck, T.W.; Harris, R.C. The effect of beta-alanine supplementation on neuromuscular fatigue in elderly (55-92 years): a double-blind randomized study. J. Int. Soc. Sports Nutr. 2008, 5, 21. [Google Scholar]
- Sweeney, K.M.; Wright, G.A.; Brice, A.G.; Doberstein, S.T. The effect of beta-alanine supplementation on power performance during repeated sprint activity. J. Strength Cond. Res. 2009. In Press. [Google Scholar]
- Schott, J.; McCully, K.; Rutherford, O.M. The role of metabolites in strength training. II. Short versus long isometric contractions. Eur. J. Appl. Physiol. Occup. Physiol. 1995, 71, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Edge, J.; Hill-Haas, S.; Goodman, C.; Bishop, D. Effects of resistance training on H+ regulation, buffer capacity, and repeated sprint. Med. Sci. Sports Exerc. 2006, 38, 2004–2011. [Google Scholar]
- Kendrick, I.P.; Kim, H.J.; Harris, R.C.; Kim, C.K.; Dang, V.H.; Lam, T.Q.; Bui, T.T.; Wise, J.A. The effect of 4 weeks beta-alanine supplementation and isokinetic training on carnosine concentrations in type I and II human skeletal muscle fibers. Eur. J. Appl. Physiol. 2009, 106, 131–138. [Google Scholar]
- Greenhaff, P.L.; Casey, A.; Short, A.H.; Harris, R.C.; Soderlund, K.; Hultman, E. Influence of oral creatine supplementation on muscle torque during repeated bouts of maximal voluntary exercise in man. Clin. Sci. 1993, 84, 565–571. [Google Scholar]
- deVries, H.A.; Tichy, M.W.; Housh, T.J.; Smyth, K.D.; Tichy, A.M.; Housh, D.J. A method for estimating physical working capacity at the fatigue threshold (PWCFT). Ergonomics 1987, 30, 1195–1204. [Google Scholar]
- Matsumoto, T.; Ito, K.; Moritani, T. The relationship between anaerobic threshold and electromyographic fatigue threshold in college women. Eur. J. Appl. Physiol. Occup. Physiol. 1990, 63, 1–5. [Google Scholar]
- Moritani, T.; Takaishi, T.; Matsumoto, T. Determination of maximal power output at neuromuscular fatigue threshold. J. Appl. Physiol. 1993, 74, 1729–1734. [Google Scholar]
- McKenna, M.J. The roles of ionic processes in muscular fatigue during intense exercise. Sports Med. 1992, 13, 134–145. [Google Scholar]
- deVries, H.A.; Moritani, T.; Nagata, A.; Magnussen, K. The relation between critical power and neuromuscular fatigue as estimated from electromyographic data. Ergonomics 1982, 25, 783–791. [Google Scholar]
- Heymsfield, S.B.; Arteaga, C.; McManus, C. Measurement of muscle mass in humans: validity of the 24-hour urinary creatinine method. Am. J. Clin. Nutr. 1983, 37, 478–494. [Google Scholar]
- McArdle, W.D.; Katch, F.I.; Katch, V.L. Exercise Physiology: energy, nutrition and human performance; Lea & Febiger: Philadelphia, NY, USA, 1991. [Google Scholar]
- Bessman, S.; Savabi, F. The role of phosphocreatine energy shuttle in exercise and muscle hypertrophy. In Creatine and Creatine Phosphate: Scientific and Clinical Perspectives; Conway M. A.;, Clark, J., F., Eds.; Academic Press: San Diego, CA, USA, 1988. [Google Scholar]
- Wallimann, T.; Dolder, M.; Schlattner, U.; Eder, M.; Hornemann, T.; O'Gorman, E.; Ruck, A.; Brdiczka, D. Some new aspects of creatine kinase (CK): Compartmentation, structure, function and regulation for cellular and mitochondrial bioenergetics and physiolog. Biofactors 1998, 8, 229–234. [Google Scholar]
- Hultman, E.; Bergstrom, J.; McLennan-Anderson, N. Breakdown and resynthesis of phosphorylcreatine and adenosine-triphosphate in connection with muscular work in man. Scand. J. Clin. Lab. Invest. 1967, 19, 56–66. [Google Scholar]
- Spriet, L.L.; Soderlund, K.; Bergstrom, M. Anaerobic energy release in skeletal muscle during electrical stimulation in men. J. Appl. Physiol. 1987, 62, 611–615. [Google Scholar]
- Greenhaff, P.L.; Bodin, K.; Soderlund, K.; Hultman, E. Effect of oral creatine supplementation on skeletal muscle phosphocreatine resynthesis. Am. J. Physiol. 1994, 266, E725–E730. [Google Scholar]
- Bemben, M.G.; Bemben, D.A.; Loftiss, D.D.; Khehans, A.W. Creatine supplementation during resistance training in college football players. Med. Sci. Sports Exerc. 2001, 33, 1667–1673. [Google Scholar]
- Kreider, R.B.; Ferreira, M.; Wilson, M.; Grindstaff, P.; Plisk, S.; Reinardy, J.; Cantler, E.; Almada, A.L. Effects of creatine supplementation on body composition, strength, and sprint performance. Med. Sci. Sports Exerc. 1998, 30, 73–82. [Google Scholar] [PubMed]
- Vandenberghe, K.; Goris, M.; Van Hecke, P.; Van Leemputte, M.; Vangerven, L.; Hespel, P. Long-term creatine intake is beneficial to muscle performance during resistance training. J. Appl. Physiol. 1997, 83, 2055–2063. [Google Scholar]
- Kreider, R.B.; Ferreira, M.; Wilson, M.; Grindstaff, P.; Plisk, S.; Reinardy, J.; Cantler, E.; Almada, A.L. Effects of creatine supplementation, strength and sprint performance. Med. Sci. Sports Exerc. 1998, 30, 73–82. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Culbertson, J.Y.; Kreider, R.B.; Greenwood, M.; Cooke, M. Effects of Beta-Alanine on Muscle Carnosine and Exercise Performance: A Review of the Current Literature. Nutrients 2010, 2, 75-98. https://doi.org/10.3390/nu2010075
Culbertson JY, Kreider RB, Greenwood M, Cooke M. Effects of Beta-Alanine on Muscle Carnosine and Exercise Performance: A Review of the Current Literature. Nutrients. 2010; 2(1):75-98. https://doi.org/10.3390/nu2010075
Chicago/Turabian StyleCulbertson, Julie Y., Richard B. Kreider, Mike Greenwood, and Matthew Cooke. 2010. "Effects of Beta-Alanine on Muscle Carnosine and Exercise Performance: A Review of the Current Literature" Nutrients 2, no. 1: 75-98. https://doi.org/10.3390/nu2010075
APA StyleCulbertson, J. Y., Kreider, R. B., Greenwood, M., & Cooke, M. (2010). Effects of Beta-Alanine on Muscle Carnosine and Exercise Performance: A Review of the Current Literature. Nutrients, 2(1), 75-98. https://doi.org/10.3390/nu2010075