Administration of Momordica charantia Enhances the Neuroprotection and Reduces the Side Effects of LiCl in the Treatment of Alzheimer’s Disease

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Wild Bitter Gourd Powder
2.3. Animal Experimental Design
2.4. Y Maze
2.5. Morris Water Maze (MWM)
2.6. Immunohistochemistry
2.7. Preparation of Liver Samples
2.8. Western Blot Analysis
2.9. Data Analysis
3. Results
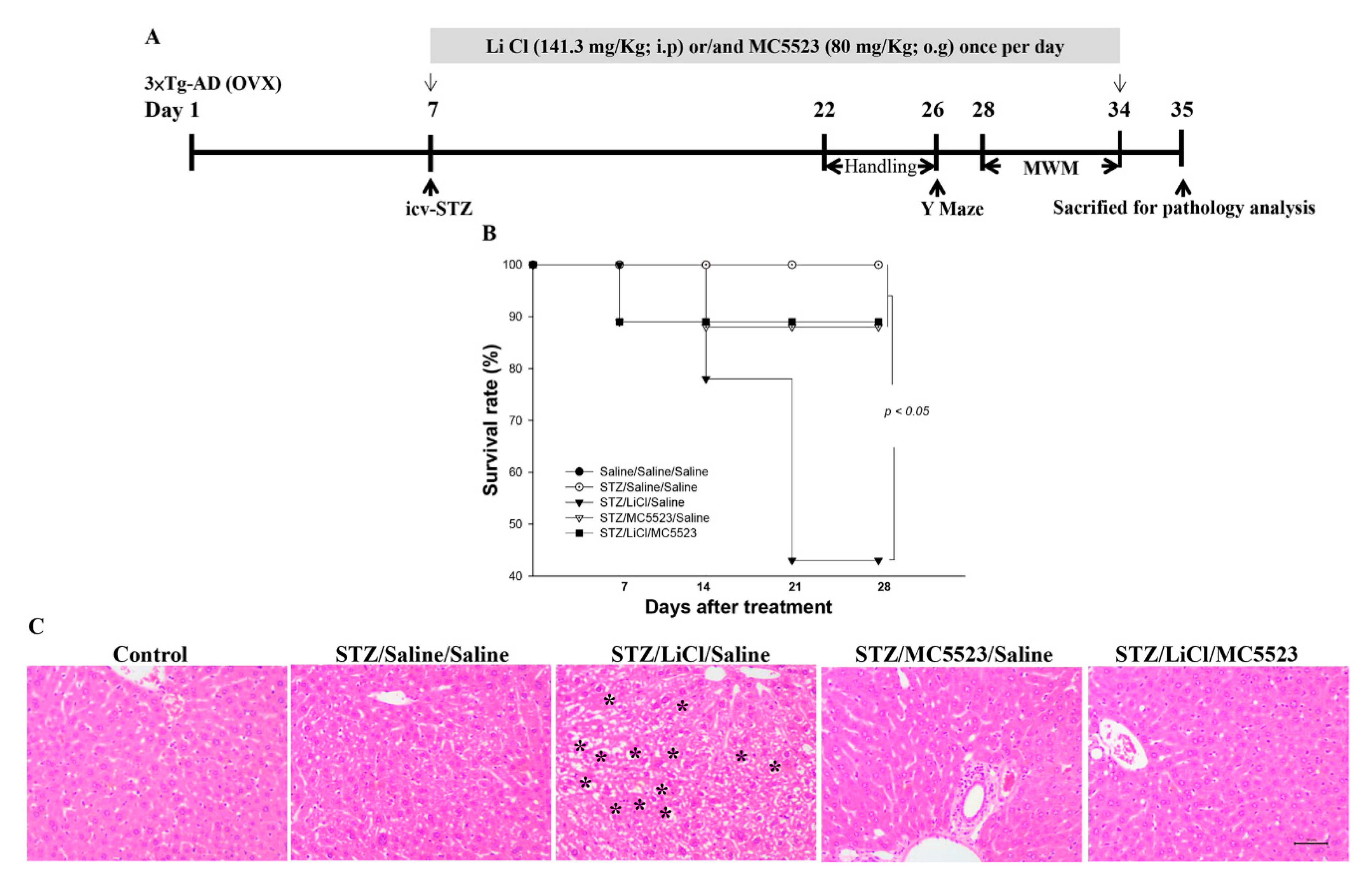
3.1. Administration of MC5523 Enhances Neuroprotection and Reduces Hepatotoxicity Induced by LiCl in icv-STZ OVX 3×Tg-AD Mice
3.2. Cotreatment with MC5523 and LiCl Greatly Prevents Neuronal Loss in icv-STZ OVX 3×Tg-AD Mice
3.3. Cotreatment with MC5523 and LiCl Greatly Reduces Gliosis in icv-STZ OVX 3×Tg-AD Mice
3.4. Combination of MC5523 and LiCl Shows More Beneficial Effects on Short-Term Memory in icv-STZ B6 Mice
3.5. Combination of MC5523 and LiCl Improves Spatial Cognition in icv-STZ B6 Mice
3.6. Combination of MC5523 and LiCl Prevents the Loss of Cholinergic and Noradrenergic Neurons in icv-STZ B6 Mice
3.7. Combination of MC5523 and LiCl Greatly Increases PSD95 and MAP2 Expression and the NR2A/NR2B Ratio in icv-STZ B6 Mice
3.8. Combination of MC5523 and LiCl Greatly Alleviates Gliosis in icv-STZ B6 Mice
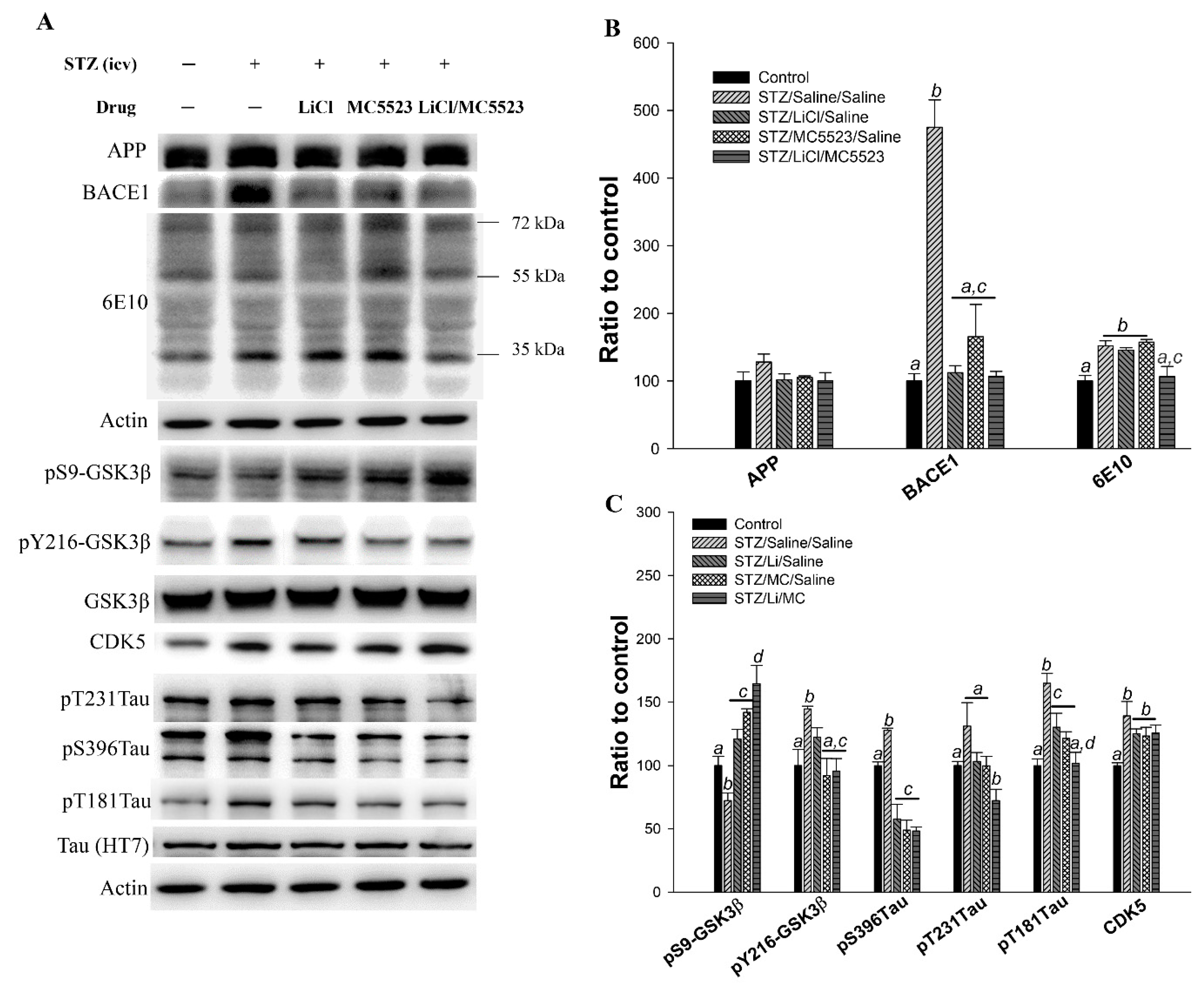
3.9. Combination of MC5523 and LiCl Decreases Oligomer Aβ and Tau Phosphorylation Levels by Increasing pS9-GSK3β in icv-STZ B6 Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mesulam, M. The cholinergic lesion of Alzheimer’s disease: Pivotal factor or side show? Learn Mem. 2004, 11, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Barage, S.H.; Sonawane, K.D. Amyloid cascade hypothesis: Pathogenesis and therapeutic strategies in Alzheimer’s disease. Neuropeptides 2015, 52, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Tabner, B.J.; Mayes, J.; Allsop, D. Hypothesis: Soluble abeta oligomers in association with redox-active metal ions are the optimal generators of reactive oxygen species in Alzheimer’s disease. Int. J. Alzheimers Dis. 2010, 2011, 546380. [Google Scholar] [CrossRef] [PubMed]
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s disease. N. Engl. J. Med. 2010, 362. [Google Scholar] [CrossRef] [PubMed]
- Kroner, Z. The relationship between Alzheimer’s disease and diabetes: Type 3 diabetes? Altern. Med. Rev. 2009, 14, 373–379. [Google Scholar] [PubMed]
- Lontchi-Yimagou, E.; Sobngwi, E.; Matsha, T.E.; Kengne, A.P. Diabetes mellitus and inflammation. Curr. Diab. Rep. 2013, 13, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Morales, I.; Farias, G.; Maccioni, R.B. Neuroimmunomodulation in the pathogenesis of Alzheimer’s disease. Neuroimmunomodulation 2010, 17. [Google Scholar] [CrossRef]
- Salkovic-Petrisic, M.; Hoyer, S. Central insulin resistance as a trigger for sporadic Alzheimer-like pathology: An experimental approach. J. Neural Transm. Suppl. 2007, 217–233. [Google Scholar]
- Salkovic-Petrisic, M.; Osmanovic, J.; Grunblatt, E.; Riederer, P.; Hoyer, S. Modeling sporadic Alzheimer's disease: The insulin resistant brain state generates multiple long-term morphobiological abnormalities including hyperphosphorylated tau protein and amyloid-beta. J. Alzheimers Dis. 2009, 18, 729–750. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liang, Z.; Tian, Z.; Blanchard, J.; Dai, C.L.; Chalbot, S.; Iqbal, K.; Liu, F.; Gong, C.X. Intracerebroventricular streptozotocin exacerbates Alzheimer-like changes of 3xTg-AD mice. Mol. Neurobiol. 2014, 49, 547–562. [Google Scholar] [CrossRef]
- Kim, J.; Moon, Y.; Hong, S. Identification of lead small molecule inhibitors of glycogen synthase kinase-3 beta using a fragment-linking strategy. Bioorg. Med. Chem. Lett. 2016, 26, 5669–5673. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Latypova, X.; Wilson, C.M.; Magnaudeix, A.; Perrin, M.L.; Yardin, C.; Terro, F. Tau protein kinases: Involvement in Alzheimer’s disease. Ageing Res. Rev. 2013, 12, 289–309. [Google Scholar] [CrossRef] [PubMed]
- Takashima, A.; Noguchi, K.; Michel, G.; Mercken, M.; Hoshi, M.; Ishiguro, K.; Imahori, K. Exposure of rat hippocampal neurons to amyloid beta peptide (25–35) induces the inactivation of phosphatidyl inositol-3 kinase and the activation of tau protein kinase I/glycogen synthase kinase-3 beta. Neurosci. Lett. 1996, 203, 33–36. [Google Scholar] [CrossRef]
- Himmelstein, D.S.; Ward, S.M.; Lancia, J.K.; Patterson, K.R.; Binder, L.I. Tau as a therapeutic target in neurodegenerative disease. Pharmacol. Ther. 2012, 136, 8–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, K.R.; Remmers, C.; Fu, Y.; Brooker, S.; Kanaan, N.M.; Vana, L.; Ward, S.; Reyes, J.F.; Philibert, K.; Glucksman, M.J.; Binder, L.I. Characterization of prefibrillar Tau oligomers in vitro and in Alzheimer disease. J. Biol. Chem. 2011, 286, 23063–23076. [Google Scholar] [CrossRef] [PubMed]
- Lasagna-Reeves, C.A.; Castillo-Carranza, D.L.; Sengupta, U.; Sarmiento, J.; Troncoso, J.; Jackson, G.R.; Kayed, R. Identification of oligomers at early stages of tau aggregation in Alzheimer’s disease. FASEB J. 2012, 26, 1946–1959. [Google Scholar] [CrossRef] [PubMed]
- Tariot, P.N.; Aisen, P.S. Can lithium or valproate untie tangles in Alzheimer’s disease? J. Clin. Psychiatry 2009, 70, 919–921. [Google Scholar] [CrossRef]
- Kessing, L.V.; Forman, J.L.; Andersen, P.K. Does lithium protect against dementia? Bipolar Disord. 2010, 12, 87–94. [Google Scholar] [CrossRef]
- Forlenza, O.V.; Diniz, B.S.; Radanovic, M.; Santos, F.S.; Talib, L.L.; Gattaz, W.F. Disease-modifying properties of long-term lithium treatment for amnestic mild cognitive impairment: Randomized controlled trial. Br. J. Psychiatry 2011, 198, 351–356. [Google Scholar] [CrossRef]
- Kessing, L.V.; Sondergard, L.; Forman, J.L.; Andersen, P.K. Lithium treatment and risk of dementia. Arch. Gen. Psychiatry 2008, 65, 1331–1335. [Google Scholar] [CrossRef]
- Gould, T.D.; Picchini, A.M.; Einat, H.; Manji, H.K. Targeting glycogen synthase kinase-3 in the CNS: Implications for the development of new treatments for mood disorders. Curr. Drug Targets 2006, 7, 1399–1409. [Google Scholar] [CrossRef] [PubMed]
- Jope, R.S.; Roh, M.S. Glycogen synthase kinase-3 (GSK3) in psychiatric diseases and therapeutic interventions. Curr. Drug Targets 2006, 7, 1421–1434. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Sintes, R.; Lucas, J.J. NFAT/Fas signaling mediates the neuronal apoptosis and motor side effects of GSK-3 inhibition in a mouse model of lithium therapy. J. Clin. Investig. 2010, 120, 2432–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaldubina, A.; Agam, G.; Belmaker, R.H. The mechanism of lithium action: State of the art, ten years later. Prog. Neuropsychopharmacol. Biol. Psychiatry 2001, 25, 855–866. [Google Scholar] [CrossRef]
- Giulietti, A.; Vignini, A.; Nanetti, L.; Mazzanti, L.; Di Primio, R.; Salvolini, E. Alzheimer’s disease risk and progression: The role of nutritional supplements and their effect on drug therapy outcome. Curr. Neuropharmacol. 2015, 14, 177–190. [Google Scholar] [CrossRef]
- Huang, M.; Hu, M.; Song, Q.; Song, H.; Huang, J.; Gu, X.; Wang, X.; Chen, J.; Kang, T.; Feng, X.; et al. GM1-modified lipoprotein-like nanoparticle: Multifunctional nanoplatform for the combination therapy of Alzheimer’s disease. ACS Nano 2015. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.Y.; Sung, P.J.; Wang, W.H.; Kuo, Y.H. Anti-inflammatory effect of Momordica charantia in sepsis mice. Molecules 2014, 19, 12777–12788. [Google Scholar] [CrossRef]
- Jiang, B.; Ji, M.; Liu, W.; Chen, L.; Cai, Z.; Zhao, Y.; Bi, X. Antidiabetic activities of a cucurbitanetype triterpenoid compound from Momordica charantia in alloxaninduced diabetic mice. Mol. Med. Rep. 2016, 14, 4865–4872. [Google Scholar] [CrossRef]
- Wang, S.; Li, Z.; Yang, G.; Ho, C.T.; Li, S. Momordica charantia: A popular health-promoting vegetable with multifunctionality. Food Funct. 2017, 8, 1749–1762. [Google Scholar] [CrossRef]
- Malik, Z.A.; Singh, M.; Sharma, P.L. Neuroprotective effect of Momordica charantia in global cerebral ischemia and reperfusion induced neuronal damage in diabetic mice. J. Ethnopharmacol. 2011, 133, 729–734. [Google Scholar] [CrossRef]
- Chen, Q.; Mo, R.; Wu, N.; Zou, X.; Shi, C.; Gong, J.; Li, J.; Fang, K.; Wang, D.; Yang, D.; et al. Berberine ameliorates diabetes-associated cognitive decline through modulation of aberrant inflammation response and insulin signaling pathway in DM rats. Front. Pharmacol. 2017, 8, 334. [Google Scholar] [CrossRef] [PubMed]
- Mardani, S.; Nasri, H.; Hajian, S.; Ahmadi, A.; Kazemi, R.; Rafieian-Kopaei, M. Impact of Momordica charantia extract on kidney function and structure in mice. J. Nephropathol. 2014, 3, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Sagkan, R.I. An in vitro study on the risk of non-allergic type-I like hypersensitivity to Momordica charantia. BMC Complement. Altern. Med. 2013, 13, 284. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.H.; Tseng, H.C.; Liu, C.T.; Huang, C.J.; Chyuan, J.H.; Sheen, L.Y. Wild bitter gourd protects against alcoholic fatty liver in mice by attenuating oxidative stress and inflammatory responses. Food Funct. 2014, 5, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.N.; Lu, K.N.; Pai, Y.P.; Chin, H.; Huang, C.J. Role of GLP-1 in the hypoglycemic effects of wild bitter gourd. Evid. Based Complement. Alternat. Med. 2013, 2013, 625892. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Chen, E.C.; Tsay, H.S.; Huang, C.J. Wild bitter gourd improves metabolic syndrome: A preliminary dietary supplementation trial. Nutr. J. 2012, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Ravelli, K.G.; Rosario, B.D.; Camarini, R.; Hernandes, M.S.; Britto, L.R. Intracerebroventricular streptozotocin as a model of alzheimer’s disease: Neurochemical and behavioral characterization in mice. Neurotox. Res. 2016. [Google Scholar] [CrossRef]
- Huang, H.J.; Huang, H.Y.; Hsieh-Li, H.M. MGCD0103, a selective histone deacetylase inhibitor, coameliorates oligomeric Abeta25-35 -induced anxiety and cognitive deficits in a mouse model. CNS Neurosci. Ther. 2018. [Google Scholar] [CrossRef]
- Huang, H.J.; Chen, S.L.; Hsieh-Li, H.M. Administration of NaHS attenuates footshock-induced pathologies and emotional and cognitive dysfunction in triple transgenic alzheimer’s mice. Front. Behav. Neurosci. 2015, 9, 312. [Google Scholar] [CrossRef]
- Huang, H.J.; Chen, Y.H.; Liang, K.C.; Jheng, Y.S.; Jhao, J.J.; Su, M.T.; Lee-Chen, G.J.; Hsieh-Li, H.M. Exendin-4 protected against cognitive dysfunction in hyperglycemic mice receiving an intrahippocampal lipopolysaccharide injection. PLoS ONE 2012, 7, e39656. [Google Scholar] [CrossRef]
- Janus, C.; Westaway, D. Transgenic mouse models of Alzheimer’s disease. Physiol. Behav. 2001, 73, 873–886. [Google Scholar] [CrossRef]
- Bronzuoli, M.R.; Iacomino, A.; Steardo, L.; Scuderi, C. Targeting neuroinflammation in Alzheimer’s disease. J. Inflamm. Res. 2016, 9, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tian, Z.; Liang, Z.; Sun, S.; Dai, C.L.; Lee, M.H.; LaFerla, F.M.; Grundke-Iqbal, I.; Iqbal, K.; Liu, F.; et al. Brain gene expression of a sporadic (icv-STZ Mouse) and a familial mouse model (3xTg-AD mouse) of Alzheimer’s disease. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Mehla, J.; Pahuja, M.; Gupta, Y.K. Streptozotocin-induced sporadic Alzheimer’s disease: Selection of appropriate dose. J. Alzheimers Dis. 2013, 33, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, Y.H.; Nam, G.H.; Choe, S.H.; Lee, S.R.; Kim, S.U.; Kim, J.S.; Sim, B.W.; Song, B.S.; Jeong, K.J.; et al. Quantitative expression analysis of APP pathway and tau phosphorylation-related genes in the ICV STZ-induced non-human primate model of sporadic Alzheimer’s disease. Int. J. Mol. Sci. 2015, 16, 2386–2402. [Google Scholar] [CrossRef] [PubMed]
- Rai, S.; Kamat, P.K.; Nath, C.; Shukla, R. Glial activation and post-synaptic neurotoxicity: The key events in Streptozotocin (ICV) induced memory impairment in rats. Pharmacol. Biochem. Behav. 2014, 117, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Zhang, Y.; Chuang, D.M. Lithium reduces BACE1 overexpression, beta amyloid accumulation, and spatial learning deficits in mice with traumatic braininjury. J. Neurotrauma 2012, 29, 2342–2351. [Google Scholar] [CrossRef]
- Schwartz, M.; Kipnis, J.; Rivest, S.; Prat, A. How do immune cells support and shape the brain in health, disease, and aging? J. Neurosci. 2013, 33, 17587–17596. [Google Scholar] [CrossRef]
- Tajes, M.; Gutierrez-Cuesta, J.; Folch, J.; Ferrer, I.; Caballero, B.; Smith, M.A.; Casadesus, G.; Camins, A.; Pallas, M. Lithium treatment decreases activities of tau kinases in a murine model of senescence. J. Neuropathol. Exp. Neurol. 2008, 67, 612–623. [Google Scholar] [CrossRef]
- Feng, H.L.; Leng, Y.; Ma, C.H.; Zhang, J.; Ren, M.; Chuang, D.M. Combined lithium and valproate treatment delays disease onset, reduces neurological deficitsand prolongs survival in an amyotrophic lateral sclerosis mouse model. Neuroscience 2008, 155, 567–572. [Google Scholar] [CrossRef]
- Xia, Y.; Rao, J.; Yao, A.; Zhang, F.; Li, G.; Wang, X.; Lu, L. Lithium exacerbates hepatic ischemia/reperfusion injury by inhibiting GSK-3beta/NF-kappaB-mediated protective signaling in mice. Eur. J. Pharmacol. 2012, 697, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Lamari, F.N.; Papasotiropoulos, V.; Tsiris, D.; Bariamis, S.E.; Sotirakis, K.; Pitsi, E.; Vogiatzoglou, A.P.; Iatrou, G. Phytochemical and genetic characterization of styles of wild Crocus species from the island of Crete, Greece and comparison to those of cultivated C. sativus. Fitoterapia 2018, 130, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Kurze, E.; Lo Scalzo, R.; Campanelli, G.; Schwab, W. Effect of tomato variety, cultivation, climate and processing on Sola l 4, an allergen from Solanum lycopersicum. PLoS ONE 2018, 13, e0197971. [Google Scholar] [CrossRef] [PubMed]
- Nerurkar, P.V.; Johns, L.M.; Buesa, L.M.; Kipyakwai, G.; Volper, E.; Sato, R.; Shah, P.; Feher, D.; Williams, P.G.; Nerurkar, V.R. Momordica charantia (bitter melon) attenuates high-fat diet-associated oxidative stress and neuroinflammation. J. Neuroinflammation 2011, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Sagor, A.T.; Chowdhury, M.R.; Tabassum, N.; Hossain, H.; Rahman, M.M.; Alam, M.A. Supplementation of fresh ucche (Momordica charantia L. var. muricata Willd) prevented oxidative stress, fibrosis and hepatic damage in CCl4 treated rats. BMC Complement. Altern. Med. 2015, 15, 115. [Google Scholar] [CrossRef]
- Katsouri, L.; Lim, Y.M.; Blondrath, K.; Eleftheriadou, I.; Lombardero, L.; Birch, A.M.; Mirzaei, N.; Irvine, E.E.; Mazarakis, N.D.; Sastre, M. PPARgamma-coactivator-1alpha gene transfer reduces neuronal loss and amyloid-beta generation by reducing beta-secretase in an Alzheimer’s disease model. Proc. Natl. Acad. Sci. USA 2016, 113, 12292–12297. [Google Scholar] [CrossRef] [PubMed]
- Durairajan, S.S.; Liu, L.F.; Lu, J.H.; Chen, L.L.; Yuan, Q.; Chung, S.K.; Huang, L.; Li, X.S.; Huang, J.D.; Li, M. Berberine ameliorates beta-amyloid pathology, gliosis, and cognitive impairment in an Alzheimer’s disease transgenic mouse model. Neurobiol. Aging. 2012, 33, 2903–2919. [Google Scholar] [CrossRef] [PubMed]
- Ly, P.T.; Wu, Y.; Zou, H.; Wang, R.; Zhou, W.; Kinoshita, A.; Zhang, M.; Yang, Y.; Cai, F.; Woodgett, J.; et al. Inhibition of GSK3beta-mediated BACE1 expression reduces Alzheimer-associated phenotypes. J. Clin. Investig. 2013, 123, 224–235. [Google Scholar] [CrossRef]
- Xu, J.; de Winter, F.; Farrokhi, C.; Rockenstein, E.; Mante, M.; Adame, A.; Cook, J.; Jin, X.; Masliah, E.; Lee, K.F. Neuregulin 1 improves cognitive deficits and neuropathology in an Alzheimer’s disease model. Sci. Rep. 2016, 6, 31692. [Google Scholar] [CrossRef] [Green Version]
- Angelo, M.; Plattner, F.; Giese, K.P. Cyclin-dependent kinase 5 in synaptic plasticity, learning and memory. J. Neurochem. 2006, 99, 353–370. [Google Scholar] [CrossRef] [Green Version]
- Ayala, R.; Shu, T.; Tsai, L.H. Trekking across the brain: The journey of neuronal migration. Cell 2007, 128, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Ip, N.Y. Cdk5: A new player at synapses. Neurosignals 2003, 12, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Evans, G.J.; Cousin, M.A. Activity-dependent control of slow synaptic vesicle endocytosis by cyclin-dependent kinase 5. J. Neurosci. 2007, 27, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Sananbenesi, F.; Spiess, J.; Radulovic, J. Cdk5: A novel role in learning and memory. Neurosignals 2003, 12, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, H.; Ortiz-Riano, E.; Krause, B.; Barriga, A.; Medina, F.; Pando, M.E.; Alberti, C.; Kettlun, A.M.; Collados, L.; Garcia, L.; et al. Microtubule proteins and their post-translational forms in the cerebrospinal fluid of patients with paraparesis associated with HTLV-I infection and in SH-SY5Y cells: An in vitro model of HTLV-I-induced disease. Biol. Res. 2008, 41, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Lazzara, C.A.; Riley, R.R.; Rane, A.; Andersen, J.K.; Kim, Y.H. The combination of lithium and l-Dopa/Carbidopa reduces MPTP-induced abnormal involuntarymovements (AIMs) via calpain-1 inhibition in a mouse model: Relevance for Parkinsons disease therapy. Brain Res. 2015, 1622, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Basu, S. Multi-targeting Strategies for Alzheimer’s Disease Therapeutics: Pros and Cons. Curr. Top Med. Chem. 2017, 17, 3017–3061. [Google Scholar] [CrossRef] [PubMed]
- Jalili-Baleh, L.; Babaei, E.; Abdpour, S.; Nasir Abbas Bukhari, S.; Foroumadi, A.; Ramazani, A.; Sharifzadeh, M.; Abdollahi, M.; Khoobi, M. A review on flavonoid-based scaffolds as multi-target-directed ligands (MTDLs) for Alzheimer’s disease. Eur. J. Med. Chem. 2018, 152, 570–589. [Google Scholar] [CrossRef]
- Wenzel, T.J.; Klegeris, A. Novel multi-target directed ligand-based strategies for reducing neuroinflammation in Alzheimer’s disease. Life Sci. 2018, 207, 314–322. [Google Scholar] [CrossRef]
- Anastasio, T.J. Computational identification of potential multi-drug combinations for reduction of microglial inflammation in Alzheimer disease. Front Pharmacol. 2015, 6, 116. [Google Scholar] [CrossRef]
- Purschke, B.; Scheibelberger, R.; Axmann, S.; Adler, A.; Jager, H. Impact of substrate contamination with mycotoxins, heavy metals and pesticides on the growth performance and composition of black soldier fly larvae (Hermetia illucens) for use in the feed and food value chain. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2017, 34, 1410–1420. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Connolly, L.; Frizzell, C.; Elliott, C.T. Challenging conventional risk assessment with respect to human exposure to multiple food contaminants in food: A case study using maize. Toxicol Lett. 2015, 238, 54–64. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Species | Supplier | WB Dilution | IHC Dilution | IF Dilution |
|---|---|---|---|---|---|
| APP | Rabbit | Sigma-Aldrich | 1:1000 | - | |
| BACE1 | Rabbit | Cell Signaling | 1:1000 | - | |
| 6E10 | Mouse | COVANCE | 1:1000 | ||
| 5-HT | Rat | Millipore | - | 1:200 | |
| ChAT | Rabbit | Millipore | - | 1:1000 | |
| TH | Rabbit | Millipore | - | 1:1000 | |
| NF-κB | Rabbit | Cell Signaling | 1:1000 | - | |
| IL-6 | Goat | Santa Cruz | 1:1000 | ||
| TNF-α | Goat | Santa Cruz | 1:1000 | ||
| IL-1β | Rabbit | Santa Cruz | 1:1000 | ||
| MnSOD | Rabbit | Millipore | 1:1000 | - | |
| GFAP | Mouse | Millipore | - | 1:1000 | |
| Iba-1 | Rabbit | Wako | - | 1:1000 | |
| CDK5 | Mouse | Millipore | 1:1000 | - | |
| pS9-GSK3β | Rabbit | Cell Signaling | 1:1000 | - | |
| GSK3β | Rabbit | Cell Signaling | 1:1000 | - | |
| PY216-GSK3β | Mouse | Millipore | 1:1,000 | ||
| pT181Tau | Rabbit | Millipore | 1:1000 | - | |
| pS396Tau | Rabbit | Invitrogen | 1:1000 | - | |
| pT231Tau | Rabbit | Invitrogen | 1:1000 | - | |
| Total tau (HT7) | Mouse | Thermo | 1:500 | ||
| NR2A | Rabbit | Millipore | 1:1000 | ||
| NR2B | Rabbit | Millipore | 1:1000 | - | |
| PSD95 | Goat | Santa Cruz | 1:1000 | - | |
| MAP2 | Rabbit | Millipore | 1:1000 | - | 1:1000 |
| Neu N | Mouse | Millipore | 1:1000 | 1:1000 | |
| β-Actin | Mouse | Millipore | 1:2000 | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.-J.; Chen, S.-L.; Chang, Y.-T.; Chyuan, J.-H.; Hsieh-Li, H.M. Administration of Momordica charantia Enhances the Neuroprotection and Reduces the Side Effects of LiCl in the Treatment of Alzheimer’s Disease. Nutrients 2018, 10, 1888. https://doi.org/10.3390/nu10121888
Huang H-J, Chen S-L, Chang Y-T, Chyuan J-H, Hsieh-Li HM. Administration of Momordica charantia Enhances the Neuroprotection and Reduces the Side Effects of LiCl in the Treatment of Alzheimer’s Disease. Nutrients. 2018; 10(12):1888. https://doi.org/10.3390/nu10121888
Chicago/Turabian StyleHuang, Hei-Jen, Shu-Ling Chen, Yen-Ting Chang, Jong-Ho Chyuan, and Hsiu Mei Hsieh-Li. 2018. "Administration of Momordica charantia Enhances the Neuroprotection and Reduces the Side Effects of LiCl in the Treatment of Alzheimer’s Disease" Nutrients 10, no. 12: 1888. https://doi.org/10.3390/nu10121888