Nesfatin-130-59 Injected Intracerebroventricularly Increases Anxiety, Depression-Like Behavior, and Anhedonia in Normal Weight Rats

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diets
2.3. Intracerebroventricular Cannulation
2.4. Peptide and Intracerebroventricular Injection
2.5. Experimental Design and Procedures
2.5.1. Sucrose Preference Test
2.5.2. Novelty-Induced Hypophagia
2.5.3. Open Field Test
2.5.4. Elevated Zero Maze
2.5.5. Light/Dark Box
2.6. Statistical Analysis
3. Results
3.1. Nefatin-130-59 Injected Intracerebroventricularly Induced Anhedonic and Depression-Like Behavior in Normal Weight but Not DIO Rats
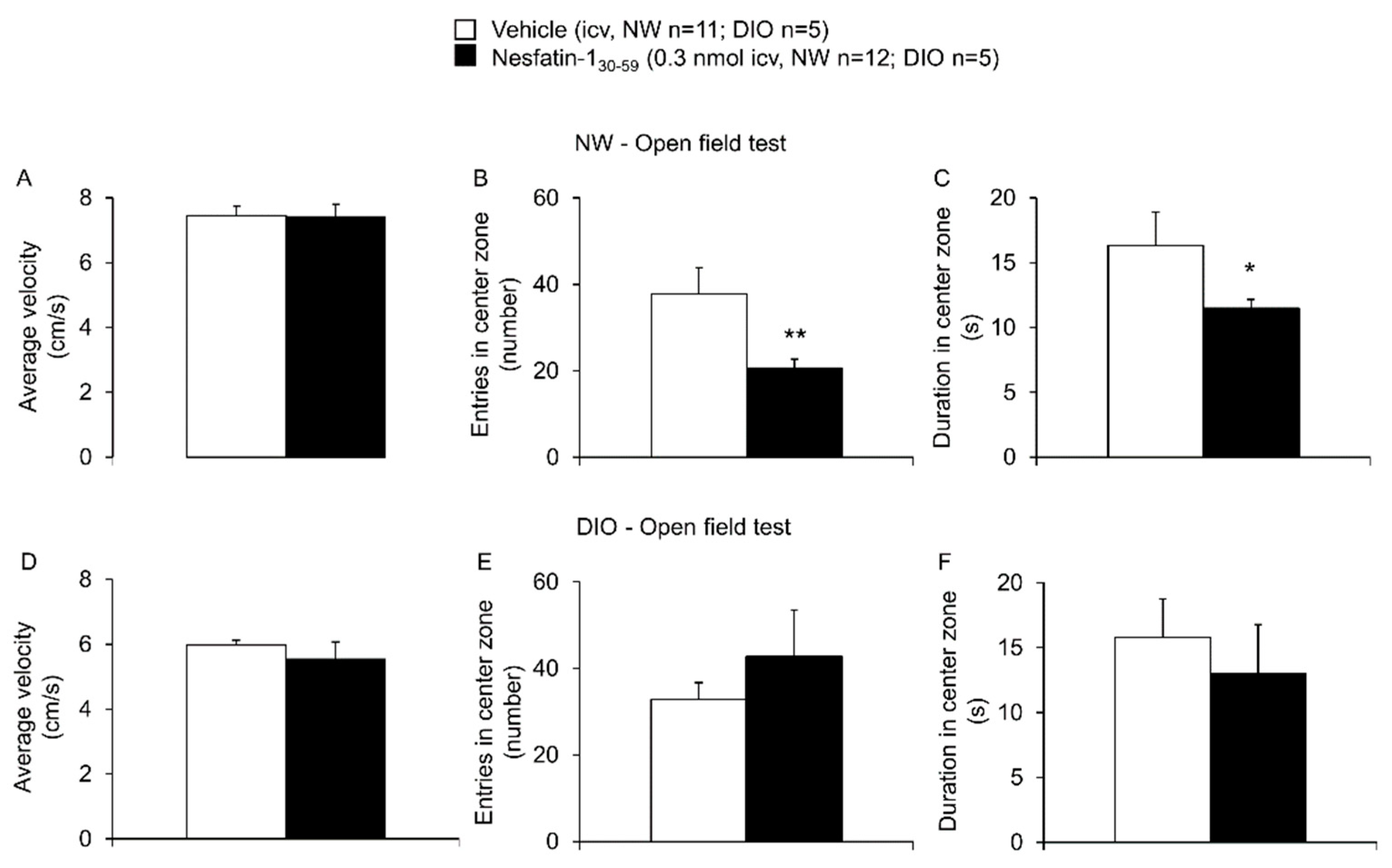
3.2. Nesfatin-130-59 Injected Intracerebroventricularly Induced Anxious Behavior in Normal Weight but Not DIO Rats
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oh-I, S.; Shimizu, H.; Satoh, T.; Okada, S.; Adachi, S.; Inoue, K.; Eguchi, H.; Yamamoto, M.; Imaki, T.; Hashimoto, K.; et al. Identification of nesfatin-1 as a satiety molecule in the hypothalamus. Nature 2006, 443, 709–712. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Oh-I, S.; Hashimoto, K.; Nakata, M.; Yamamoto, S.; Yoshida, N.; Eguchi, H.; Kato, I.; Inoue, K.; Satoh, T.; et al. Peripheral administration of nesfatin-1 reduces food intake in mice: The leptin-independent mechanism. Endocrinology 2009, 150, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Goebel, M.; Stengel, A.; Wang, L.; Lambrecht, N.W.; Taché, Y. Nesfatin-1 immunoreactivity in rat brain and spinal cord autonomic nuclei. Neurosci. Lett. 2009, 452, 241–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stengel, A.; Goebel, M.; Wang, L.; Rivier, J.; Kobelt, P.; Monnikes, H.; Lambrecht, N.W.; Taché, Y. Central nesfatin-1 reduces dark-phase food intake and gastric emptying in rats: Differential role of corticotropin-releasing factor2 receptor. Endocrinology 2009, 150, 4911–4919. [Google Scholar] [CrossRef] [PubMed]
- Prinz, P.; Teuffel, P.; Lembke, V.; Kobelt, P.; Goebel-Stengel, M.; Hofmann, T.; Rose, M.; Klapp, B.F.; Stengel, A. Nesfatin-1(30−59) injected intracerebroventricularly differentially affects food intake microstructure in rats under normal weight and diet-induced obese conditions. Front. Neurosci. 2015, 9, 422. [Google Scholar] [CrossRef] [PubMed]
- Ramanjaneya, M.; Chen, J.; Brown, J.E.; Tripathi, G.; Hallschmid, M.; Patel, S.; Kern, W.; Hillhouse, E.W.; Lehnert, H.; Tan, B.K.; et al. Identification of nesfatin-1 in human and murine adipose tissue: A novel depot-specific adipokine with increased levels in obesity. Endocrinology 2010, 151, 3169–3180. [Google Scholar] [CrossRef] [PubMed]
- Foo, K.S.; Brauner, H.; Ostenson, C.G.; Broberger, C. Nucleobindin-2/nesfatin in the endocrine pancreas: Distribution and relationship to glycaemic state. J. Endocrinol. 2010, 204, 255–263. [Google Scholar] [CrossRef] [PubMed]
- García-Galiano, D.; Pineda, R.; Ilhan, T.; Castellano, J.M.; Ruiz-Pino, F.; Sánchez-Garrido, M.A.; Vazquez, M.J.; Sangiao-Alvarellos, S.; Romero-Ruiz, A.; Pinilla, L.; et al. Cellular distribution, regulated expression, and functional role of the anorexigenic peptide, NUCB2/nesfatin-1, in the testis. Endocrinology 2012, 153, 1959–1971. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Goebel, M.; Yakubov, I.; Wang, L.; Witcher, D.; Coskun, T.; Taché, Y.; Sachs, G.; Lambrecht, N.W. Identification and characterization of nesfatin-1 immunoreactivity in endocrine cell types of the rat gastric oxyntic mucosa. Endocrinology 2009, 150, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Hofmann, T.; Goebel-Stengel, M.; Lembke, V.; Ahnis, A.; Elbelt, U.; Lambrecht, N.W.; Ordemann, J.; Klapp, B.F.; Kobelt, P. Ghrelin and NUCB2/nesfatin-1 are expressed in the same gastric cell and differentially correlated with body mass index in obese subjects. Histochem. Cell Biol. 2013, 139, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Price, T.O.; Samson, W.K.; Niehoff, M.L.; Banks, W.A. Permeability of the blood-brain barrier to a novel satiety molecule nesfatin-1. Peptides 2007, 28, 2372–2381. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Hsuchou, H.; Kastin, A.J. Nesfatin-1 crosses the blood-brain barrier without saturation. Peptides 2007, 28, 2223–2228. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Zhang, J.; Tang, Y.; Bi, F.; Liu, J.N. The novel function of nesfatin-1: Anti-hyperglycemia. Biochem. Biophys. Res. Commun. 2010, 391, 1039–1042. [Google Scholar] [CrossRef] [PubMed]
- Özsavcí, D.; Erşahin, M.; Şener, A.; Özakpinar, Ö.B.; Toklu, H.Z.; Akakin, D.; Şener, G.; Yegen, B.C. The novel function of nesfatin-1 as an anti-inflammatory and antiapoptotic peptide in subarachnoid hemorrhage-induced oxidative brain damage in rats. Neurosurgery 2011, 68, 1699–1708. [Google Scholar] [CrossRef] [PubMed]
- Yamawaki, H.; Takahashi, M.; Mukohda, M.; Morita, T.; Okada, M.; Hara, Y. A novel adipocytokine, nesfatin-1 modulates peripheral arterial contractility and blood pressure in rats. Biochem. Biophys. Res. Commun. 2012, 418, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Prinz, P.; Goebel-Stengel, M.; Teuffel, P.; Rose, M.; Klapp, B.F.; Stengel, A. Peripheral and central localization of the nesfatin-1 receptor using autoradiography in rats. Biochem. Biophys. Res. Commun. 2016, 470, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Weibert, E.; Stengel, A. The X/A-like cell revisited—Spotlight on the peripheral effects of NUCB2/nesfatin-1 and ghrelin. J. Physiol. Pharmacol. 2017, 68, 497–520. [Google Scholar] [PubMed]
- Merali, Z.; McIntosh, J.; Kent, P.; Michaud, D.; Anisman, H. Aversive and appetitive events evoke the release of corticotropin-releasing hormone and bombesin-like peptides at the central nucleus of the amygdala. J. Neurosci. 1998, 18, 4758–4766. [Google Scholar] [CrossRef] [PubMed]
- Merali, Z.; Hayley, S.; Kent, P.; McIntosh, J.; Bédard, T.; Anisman, H. Impact of repeated stressor exposure on the release of corticotropin-releasing hormone, arginine-vasopressin and bombesin-like peptides at the anterior pituitary. Behav. Brain Res. 2009, 198, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Upton, K.R.; Riley, L.G. Acute stress inhibits food intake and alters ghrelin signaling in the brain of tilapia (Oreochromis mossambicus). Domest. Anim. Endocrinol. 2013, 44, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, N.; Maejima, Y.; Sedbazar, U.; Ando, A.; Kurita, H.; Damdindorj, B.; Takano, E.; Gantulga, D.; Iwasaki, Y.; Kurashina, T.; et al. Stressor-responsive central nesfatin-1 activates corticotropin-releasing hormone, noradrenaline and serotonin neurons and evokes hypothalamic-pituitary-adrenal axis. Aging (Albany NY) 2010, 2, 775–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.Y.; Ge, J.F.; Qin, G.; Peng, Y.N.; Zhang, C.F.; Liu, X.R.; Liang, L.C.; Wang, Z.Z.; Chen, F.H.; Li, J. Acute, but not chronic, stress increased the plasma concentration and hypothalamic mRNA expression of NUCB2/nesfatin-1 in rats. Neuropeptides 2015, 54, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Goebel-Stengel, M.; Jawien, J.; Kobelt, P.; Taché, Y.; Lambrecht, N.W. Lipopolysaccharide increases gastric and circulating NUCB2/nesfatin-1 concentrations in rats. Peptides 2011, 32, 1942–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goebel, M.; Stengel, A.; Wang, L.; Taché, Y. Restraint stress activates nesfatin-1-immunoreactive brain nuclei in rats. Brain Res. 2009, 1300, 114–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stengel, A.; Goebel, M.; Wang, L.; Taché, Y. Abdominal surgery activates nesfatin-1 immunoreactive brain nuclei in rats. Peptides 2010, 31, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, M.S.; Pecchi, E.; Trouslard, J.; Jean, A.; Dallaporta, M.; Troadec, J.D. Central nesfatin-1-expressing neurons are sensitive to peripheral inflammatory stimulus. J. Neuroinflamm. 2009, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Algul, S.; Ozcelik, O. Evaluating the levels of nesfatin-1 and ghrelin hormones in patients with moderate and severe major depressive disorders. Psychiatry Investig. 2018, 15, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.; Stengel, A.; Ahnis, A.; Buße, P.; Elbelt, U.; Klapp, B.F. NUCB2/nesfatin-1 is associated with elevated scores of anxiety in female obese patients. Psychoneuroendocrinology 2013, 38, 2502–2510. [Google Scholar] [CrossRef] [PubMed]
- Merali, Z.; Cayer, C.; Kent, P.; Anisman, H. Nesfatin-1 increases anxiety- and fear-related behaviors in the rat. Psychopharmacology (Berl) 2008, 201, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.F.; Xu, Y.Y.; Qin, G.; Pan, X.Y.; Cheng, J.Q.; Chen, F.H. Nesfatin-1, a potent anorexic agent, decreases exploration and induces anxiety-like behavior in rats without altering learning or memory. Brain Res. 2015, 1629, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Davidowa, H.; Li, Y.; Plagemann, A. Altered neuronal responses to feeding-relevant peptides as sign of developmental plasticity in the hypothalamic regulatory system of body weight. Zhurnal Vyss. Nervn. Deiatelnosti Im. IP Pavlov. 2003, 53, 663–670. [Google Scholar]
- Schalla, M.; Prinz, P.; Friedrich, T.; Scharner, S.; Kobelt, P.; Goebel-Stengel, M.; Rose, M.; Stengel, A. Phoenixin-14 injected intracerebroventricularly but not intraperitoneally stimulates food intake in rats. Peptides 2017, 96, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Elsevier: London, UK, 2007. [Google Scholar]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R. Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology (Berl) 1987, 93, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Teuffel, P.; Wang, L.; Prinz, P.; Goebel-Stengel, M.; Scharner, S.; Kobelt, P.; Hofmann, T.; Rose, M.; Klapp, B.F.; Reeve, J.R., Jr.; et al. Treatment with the ghrelin-O-acyltransferase (GOAT) inhibitor GO-CoA-Tat reduces food intake by reducing meal frequency in rats. J. Physiol. Pharmacol. 2015, 66, 493–503. [Google Scholar] [PubMed]
- Goebel-Stengel, M.; Stengel, A.; Wang, L.; Ohning, G.; Taché, Y.; Reeve, J.R., Jr. CCK-8 and CCK-58 differ in their effects on nocturnal solid meal pattern in undisturbed rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R850–R860. [Google Scholar] [CrossRef] [PubMed]
- Brenes Sáenz, J.C.; Villagra, O.R.; Fornaguera Trías, J. Factor analysis of forced Swimming test, sucrose preference test and open field test on enriched, social and isolated reared rats. Behav. Brain Res. 2006, 169, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Dulawa, S.C.; Hen, R. Recent advances in animal models of chronic antidepressant effects: The novelty-induced hypophagia test. Neurosci. Biobehav. Rev. 2005, 29, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Shepherd, J.K.; Grewal, S.S.; Fletcher, A.; Bill, D.J.; Dourish, C.T. Behavioural and pharmacological characterisation of the elevated “zero-maze” as an animal model of anxiety. Psychopharmacology (Berl) 1994, 116, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Braun, A.A.; Skelton, M.R.; Vorhees, C.V.; Williams, M.T. Comparison of the elevated plus and elevated zero mazes in treated and untreated male sprague-dawley rats: Effects of anxiolytic and anxiogenic agents. Pharmacol. Biochem. Behav. 2011, 97, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Könczöl, K.; Pintér, O.; Ferenczi, S.; Varga, J.; Kovács, K.; Palkovits, M.; Zelena, D.; Tóth, Z.E. Nesfatin-1 exerts long-term effect on food intake and body temperature. Int. J. Obes. (Lond) (2005) 2012, 36, 1514–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourin, M.; Hascoët, M. The mouse light/dark box test. Eur. J. Pharmacol. 2003, 463, 55–65. [Google Scholar] [CrossRef]
- Costall, B.; Jones, B.J.; Kelly, M.E.; Naylor, R.J.; Tomkins, D.M. Exploration of mice in a black and white test box: Validation as a model of anxiety. Pharmacol. Biochem. Behav. 1989, 32, 777–785. [Google Scholar] [CrossRef]
- Whishaw, I.Q. The laboratory rat, the Pied Piper of twentieth century neuroscience. Brain Res. Bull. 1999, 50, 411. [Google Scholar] [CrossRef]
- Marques, M.D.; Waterhouse, J.M. Masking and the evolution of circadian rhythmicity. Chronobiol. Int. 1994, 11, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Bodnoff, S.R.; Suranyi-Cadotte, B.; Aitken, D.H.; Quirion, R.; Meaney, M.J. The effects of chronic antidepressant treatment in an animal model of anxiety. Psychopharmacology (Berl) 1988, 95, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Yin, C.Y.; Zhu, L.J.; Zhu, X.H.; Xu, C.; Luo, C.X.; Chen, H.; Zhu, D.Y.; Zhou, Q.G. Sucrose preference test for measurement of stress-induced anhedonia in mice. Nat. Protoc. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ari, M.; Ozturk, O.H.; Bez, Y.; Oktar, S.; Erduran, D. High plasma nesfatin-1 level in patients with major depressive disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.; Elbelt, U.; Ahnis, A.; Rose, M.; Klapp, B.F.; Stengel, A. Sex-specific regulation of NUCB2/nesfatin-1: Differential implication in anxiety in obese men and women. Psychoneuroendocrinology 2015, 60, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.F.; Xu, Y.Y.; Qin, G.; Peng, Y.N.; Zhang, C.F.; Liu, X.R.; Liang, L.C.; Wang, Z.Z.; Chen, F.H. Depression-like behavior induced by nesfatin-1 in rats: Involvement of increased immune activation and imbalance of synaptic vesicle proteins. Front. Neurosci. 2015, 9, 429. [Google Scholar] [CrossRef] [PubMed]
- Catak, Z.; Aydin, S.; Sahin, I.; Kuloglu, T.; Aksoy, A.; Dagli, A.F. Regulatory neuropeptides (ghrelin, obestatin and nesfatin-1) levels in serum and reproductive tissues of female and male rats with fructose-induced metabolic syndrome. Neuropeptides 2014, 48, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.K.; Hallschmid, M.; Kern, W.; Lehnert, H.; Randeva, H.S. Decreased cerebrospinal fluid/plasma ratio of the novel satiety molecule, nesfatin-1/NUCB-2, in obese humans: Evidence of nesfatin-1/NUCB-2 resistance and implications for obesity treatment. J. Clin. Endocrinol. Metab. 2011, 96, E669–E673. [Google Scholar] [CrossRef] [PubMed]
- Tanida, M.; Gotoh, H.; Yamamoto, N.; Wang, M.; Kuda, Y.; Kurata, Y.; Mori, M.; Shibamoto, T. Hypothalamic nesfatin-1 stimulates sympathetic nerve activity via hypothalamic ERK signaling. Diabetes 2015, 64, 3725–3736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Xu, Y.Y.; Ge, J.F.; Chen, F.H. CRHR1 mediates the up-regulation of synapsin I induced by nesfatin-1 through ERK 1/2 signaling in SH-SY5Y cells. Cell. Mol. Neurobiol. 2018, 38, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, K.; Masaki, T.; Chiba, S.; Ando, H.; Fujiwara, K.; Shimasaki, T.; Mitsutomi, K.; Katsuragi, I.; Kakuma, T.; Sakata, T.; et al. Brain-derived neurotrophic factor, corticotropin-releasing factor, and hypothalamic neuronal histamine interact to regulate feeding behavior. J. Neurochem. 2013, 125, 588–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.J.; Liu, M.Y.; Li, H.; Liu, X.; Chen, C.; Han, Z.; Wu, H.Y.; Jing, X.; Zhou, H.H.; Suh, H.; et al. The different roles of glucocorticoids in the hippocampus and hypothalamus in chronic stress-induced HPA axis hyperactivity. PLoS ONE 2014, 9, e97689. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kühne, S.G.; Schalla, M.A.; Friedrich, T.; Kobelt, P.; Goebel-Stengel, M.; Long, M.; Rivalan, M.; Winter, Y.; Rose, M.; Stengel, A. Nesfatin-130-59 Injected Intracerebroventricularly Increases Anxiety, Depression-Like Behavior, and Anhedonia in Normal Weight Rats. Nutrients 2018, 10, 1889. https://doi.org/10.3390/nu10121889
Kühne SG, Schalla MA, Friedrich T, Kobelt P, Goebel-Stengel M, Long M, Rivalan M, Winter Y, Rose M, Stengel A. Nesfatin-130-59 Injected Intracerebroventricularly Increases Anxiety, Depression-Like Behavior, and Anhedonia in Normal Weight Rats. Nutrients. 2018; 10(12):1889. https://doi.org/10.3390/nu10121889
Chicago/Turabian StyleKühne, Stephanie Gladys, Martha Anna Schalla, Tiemo Friedrich, Peter Kobelt, Miriam Goebel-Stengel, Melissa Long, Marion Rivalan, York Winter, Matthias Rose, and Andreas Stengel. 2018. "Nesfatin-130-59 Injected Intracerebroventricularly Increases Anxiety, Depression-Like Behavior, and Anhedonia in Normal Weight Rats" Nutrients 10, no. 12: 1889. https://doi.org/10.3390/nu10121889