Recent Progress in Perennial Buckwheat Development


Research Center of Buckwheat Industry Technology, Guizhou Normal University, Baoshan Beilu 116, Guiyang 550001, Guizhou, China
*
Author to whom correspondence should be addressed.
Sustainability 2018, 10(2), 536; https://doi.org/10.3390/su10020536
Submission received: 8 January 2018
/
Revised: 9 February 2018
/
Accepted: 13 February 2018
/
Published: 17 February 2018
(This article belongs to the Special Issue Strategies, Advances and Challenges of Breeding Perennial Grain Crops)
Abstract
:Grains in the genus Fagopyrum have benefits to human health and are an excellent gluten-free raw material. Of all cereal foods, this genus has the highest total content of amino-acid nutrients necessary for humans; nutrients that are resistant to digestion (protein and starch) resulting in their sustained release; higher dietary fiber content than key cereals, and is rich in a special healthy ingredient (flavonoids). Fagopyrum includes 24 species of which five are perennial. Among them, golden buckwheat (F.cymosum complex) is the most important perennial buckwheat, which is not only used in Chinese medicine, but also has great potential in healthy food crop. In order to provide some clues for perennial crop studies and their industry development, this paper presents the state of perennial buckwheat research in terms of taxonomy; natural chemical products and pharmacological and health functions; genetics and evolution; breeding; and product development and utilization. The great advances such as successful interspecific crossing and its subsequent new perennial buckwheat varieties will speed up the development of the perennial buckwheat industry.
1. Introduction
Grains in the genus Fagopyrum are an excellent gluten-free raw material and contain a range of nutrients including bioactive carbohydrates, essential amino-acid, proteins, phytosterols, vitamins, carotenoids, and minerals. Buckwheat, compared to other cereals, has both the highest content and distribution of essential amino-acid for humans [1,2,3], resulting in balanced nutrition without excesses or deficiencies. Furthermore, buckwheat has a higher content of albumin protein and starches resistant to digestion, resulting in the sustained release of nutrients, which is very important for people with diabetes and other diseases. Most importantly, buckwheat has the highest content of rutin with multiple phenolic hydroxyl groups in cereals [4], which can provide protection from diseases such as cardiovascular disease and cancer. In a traditional agricultural production model, the establishment of annual crops has both economic and agronomic implications such as high seed and nutrient inputs, ploughing, and may involve a number of sowings each year. Perennial crops have an important property, that is, “plant once and harvest always”, indicating a lower seeding cost and workload. Agriculture based on perennial crops may provide a model with high output and with lower inputs [5].
There are many perennial crop examples in vegetable and fruit crops, but few in grain and oilseed crops. There are some perennial species in the genus Fagopyrum, which have been studied for perennial food crop use for the last decade [1,5,6,7]. This paper will present the current state of perennial buckwheat development in China and may provide some clues for studies on other perennial food crops.
2. Taxonomy
Gross (1913) first classified the species of the Polygonaceae native to Asia including buckwheat [8]. Steward (1930) later classified ten buckwheat species in the Polygonaceae native to Asia [1]. Among them, F.suffruticosum Schm. is native to Sakhalin and the rest are all native to southwest China. Miyabe and Miyake (1915) [1] suggested that F.suffruticosum is the same species as F.tataricum in their book (Flora of Sakhalin). Ohnishi’s research group collected cultivated and wild buckwheat resources native to China and the Himalaya region beginning in 1988 [9,10] and suggested one big-achene species (F.homotropicum Ohnishi = F.esculentum var. homotropicum) and five new small-achene species (F.callianthum Ohnishi, F.pleioramosum Ohnishi, F.capillatum Ohnishi, F.rubifolium Ohsako et Ohnishi, F.macrocarpum Ohsako et Ohnishi). Chen (1999) suggested three new big-achene species, tetraploid annual F.zuogongense Q.F. Chen, diploid perennial F.megaspartanium Q.F. Chen and F.pilus Q.F. Chen [6,7]. Krotov (1973) [11] made a new annual species (F.giganteum) by crossing the tartary buckwheat and F.cymosum complex. Due to the similarity in morphology, the three large-achene perennial buckwheat species are normally called the F.cymosum complex. Liu et al. (2008) [12,13] reported two new species (F.crispatifolium Liu, F.densiovillosum Liu) in the small-achene group. Chen (2016) [14] made a new perennial species (F.tatari-cymosum Q.F. Chen) by crossing annual autotetraploid tartary buckwheat and perennial tetraploid F.cymosum.
So far, there are about 24 buckwheat species (Table 1), including two groupings: the big-achene group (namely F.cymosum group) consisting of eight; and the small-achene group (namely F.urophyllum group) consisting of sixteen species. The species in the big-achene group have much longer seeds than their persistent perianths, and bigger seeds than those in the small-achene group. Among all buckwheat species, only common buckwheat (F.esculentum) and tartary buckwheat (F.tataricum) are cultivated for grain food production. Common buckwheat is distributed in almost all grain crop countries including Asia, Europe, America, and Austria and tartary buckwheat is largely distributed only in Asia. Many aspects of the natural species in the big-achene group have been studied and proved to be biological species with reproduction isolation by means of interspecific hybridization [1,6,7]. There have been few studies on the species in the small-achene group as they have less agronomic importance.
In the big-achene group, there are three perennial species (diploid F.megaspartanium, diploid F.pilus, and allotetraploid F.cymosum) and a man-made perennial allotetraploid F.tatari-cymosum (perennial tartary buckwheat). In the small-achene group, there are two perennial species (F.urophyllum and F.statice).
3. Genetics and Evolution
Some early reports on buckwheat chromosome number [2,16,17] identified diploid F. esculentum (2n = 2x = 16), diploid F. tataricum (2n = 2x = 16) and tetraploid F. cymosum (2n = 4x = 32).
The phylogeny of the genus Fagopyrum including perennial buckwheat and the origins of cultivated buckwheat species have been studied using morphology [6,7,8], cytology [6,7,18,19], isozymes [6,7,20,21,22,23,24], seed protein content and subunit composition [25,26], molecular genetic markers such as AFLP/RAPD [27,28], and interspecific reproduction [6,7]. These works have established the F.cymosum complex and promoted the hypothesis that diploid perennial F.megaspartanium and diploid perennial F.pilus maybe the ancestral species of common buckwheat (F. esculentum) and tartary buckwheat (F.tataricum), respectively. At present, the genomes (Table 1) of most of buckwheat species are known or hypothesized [6,7,18,19,29].
There are some reports regarding golden buckwheat molecular genetics. Zhang et al. [30] studied the genetic diversity of 92 individuals from eight golden buckwheat populations using ISSR marker, and found high genetic diversity with the majority of the genetic variation occurring within the populations. Wu et al. [31] showed the sequence variation of the intergenic transcribed spacer region between the 16SrRNA and 23S rRNA genes (ITS).
Since the leaves of F. cymosum have high contents of flavonoid and high antioxidant activity than those of cultivated buckwheat [4], it is significant to study their genes. Li et al. [32,33] identified some genes from the F.cymosum complex using transcriptome sequencing, including Ocopherol cyclase (TC) and Flavanone 3′5′-hydroxylase. Meng et al. [34] obtained the chalcone synthase gene (1650bp, 395 amino acids) from the F.cymosum complex by homology cloning, genome-walking and RT-PCR. Using similar technology, Bu et al. [35] and Li et al. [36] cloned the anthocyanin synthase gene and phenylalanine ammonialyase gene FdPAL, respectively, in the F.cymosum complex. Liang et al. [37] studied the MAPK gene sequence and its variation among buckwheat species.
Li et al. [15] established the fluorescent chromosome in situ PCR technology for buckwheat and located 16S rDNA, 4.5S rDNA and psbAcpDNA genetic markers which were found with different abundances and physical distributions in the nuclear genomes of the seven buckwheat species including golden buckwheat.
With the development of transcriptomics and genomics, related technologies are entering the buckwheat field. Li et al. (2015) [38] used transcriptomics data of young tartary buckwheat seeds to develop EST-SSR. Yasuo et al. (2016) [39] established a common buckwheat genome database (BGDB) and generated a draft assembly of the common buckwheat genome using short reads obtained by next generation sequencing (NGS). Zhang et al. [40] reported a high-quality, chromosome-scale tartary buckwheat genome sequence of 489.3 Mb that is assembled by combining whole-genome shotgun sequencing of both Illumina short reads and single-molecule real-time long reads; sequence tags of a large DNA insert fosmid library; Hi-C sequencing data; BioNanogenome maps, and annotated 33,366 high-confidence protein-coding genes based on expression evidence.
4. Natural Chemical Products and Pharmacological and Health Functions
Studies on the chemical properties of perennial buckwheat have mainly focused on golden buckwheat (F.cymosum complex) because it is an important Chinese medicine. To date, many monomer compounds have been identified by infrared spectrometry (IR), ultraviolet spectrometry (US), the nuclear magnetic resonance (NMR) method and mass spectrometry (MS) and spectrum analysis [41].
Polyphenols are the most important active components in golden buckwheat, which are mainly a kind of mixture of primary anthocyanine tannins [42] and some flavonoids. Liu et al. (1980) [43] isolated the dimeric procyanidin from golden buckwheat, that is, the C4–C8 dimer of 5,7,3,4-four hydroxyl flavanols. Zhang et al. (1994) [44] reported that the extract medicine “Wei Manning” from the golden buckwheat included 3,4-dihydroxybenzoic acid, gallic acid, (-)epicatechin, (-)epicatechin-3-0 gallic acid ester, procyanidin B-2, procyanidin C-1, of which procyanidin B-2 is the main ingredient with the concentration of 0.19%. Shao et al. (2005) [45,46] discovered protocatechuic acid, trans-p-hydroxycinnamic methyl ester, 3,4-dihydroxybenzamide, and protocatechuic acid methyl ester. Wang et al. (2005) [47] obtained diboside A and lapathoside A from a golden buckwheat water acetone extract. Golden buckwheat also contains luteolin [45,46], quercitrin and rutin [48]. Wu et al. (2008) [49] first discovered pratol and luteolin-7′4′-dimethylether and rhamnetin, 3,6,3′,4′-tetrahydroxy-7-methoxyflavonetc flavonoids. Golden buckwheat contains some steroids such as Hecogenin and β-sitosterol [43] and some triterpenes such as glutinone and glutinol [45,46].
Golden buckwheat also contains glycerol monopalmitate, n-butyl-β-d-fructopyronoside [45,46], emodin, and β-daucosterol [49]. Bai et al. (2007) [50] isolated and identified thirteen hydrocarbons and thirty hydrocarbon oxides from golden buckwheat volatile components. They mainly contained palmitic acid, (Z,Z)9,12-eighteen diolefinic acid,1,4,4a,5,6,7,8,8a-eight hydrogen-2,5,5,8a-four methyl-1-naphthalene ene methanol; camphor and naphthalene; [lar(la,α,4a,α,7.β,7α.β,7b.α)] -dehydrogenation-1,1,7-three methyl-4-methylene-1H-Ciprofloxacin [e] azulene-7-alcohol; n-pelargonic aldehyde, and linalool, etc.
As stated above, golden buckwheat (F.cymosum complex) is rich in natural chemical products. Many scientists have studied golden buckwheat’s health promoting and curative functions with respect to human health. Modern pharmacological studies have found that golden buckwheat can inhibit cancer cells, enhance human immunity, reduce the level of blood sugar and blood lipid, and can be clinically used to cure cancer, diabetes, hyperlipidemia and aid in the treatment of rheumatism. Therefore, it has high medicinal value and health care function [51,52,53,54].
5. Breeding of Perennial Buckwheat—the F.cymosum Complex
Buckwheat breeding is focused on seed yield and the quality of cultivated common and tartary buckwheat. Cultivated types of buckwheat are annual. At present, the average yields of common and tartary buckwheat are about 1050kg/ha (with the highest yield of 4200kg/ha) and 1800kg/ha (with the highest yield of 4800kg/ha) respectively.
All natural perennial buckwheat are wild type; they have many negative traits including shattering, high sensitivity to light and temperature, indeterminate flower and fruit development, low fertility in spring and summer seasons, non-compact plant, low yield, strong seed dormancy and uneven germination, etc.
It is clear that perennial buckwheat is not fit for harvesting seeds, and most buckwheat has not yet been cultivated, except the F.cymosum complex. The traditional utility of the F.cymosum complex is mainly found in its perennial caudex which has medicinal applications in China. The F.cymosum complex in the big-achene group contains perennial buckwheat, has big seeds, higher yield potential, and much stronger adaptability and regenerative capacity than annual common and tartary buckwheat and the perennial buckwheat in the small-achene group. Consequently, Russia, Japan and China are trying to improve annual cultivated buckwheat by wide crossing with the F.cymosum complex.
The interspecific hybridization has proved to be very difficult [7,55,56,57,58] for producing normal hybrid seeds; the reasons for this maybe the lack of an ideal female parent with high crossability and compatibility. In order to improve the yield of buckwheat varieties, many hybridizations between buckwheat species have been attempted, but most of the interspecific crosses have not been successful in obtaining interspecific hybrid progenies as yet, with the exception of the following crosses: autotetraploid F.esculentum × tetraploid F. zuogongense (Chen [7]) and tetraploid F. tataricum × tetraploid F. cymosum [11,16,59,60,61,62]. Among them, Krotov and Dranenko’s (1973) wide hybridization involved the perennial buckwheat parent of the F.cymosum complex and produced an annual man-made new species of buckwheat (F.giganteum) with white hetero-style flowers [11]. Furthermore, the new species has not been cultivated because of reduced size seeds and much more shell than common and tartary buckwheat.
Chen (1999) [6,7] started to carry out perennial buckwheat breeding, including the perennial buckwheat, by selective breeding, cross breeding and interspecific hybridization breeding.
One way to improve perennial wild buckwheat is by selective breeding. There are many mutations that have occurred in natural populations of perennial buckwheat. In order to develop many varieties with a high flavonoid content in the leaves and flower of the F.cymosum complex, we collected 360 accessions nationwide in China and estimated their flavonoid content in leaves and flowers including F.megaspartanium, F.pilus, and F.cymosum. The first perennial variety from natural perennial species is Gui Jinqiaomai No.1 developed in 2005. It is from diploid (2x = 16) perennial F.megaspartanium populations native to Guiyang by single selection. Because it has high flavonoid content of up to 10.2% in leaves, much higher than the average 4.5%, it was used for harvesting leaves and making leaf tea products. Seed harvest is difficult due to shattering therefore it is reproduced via cutting from branches. Adaptational difficults due to high temperature and the sensitivity to light intensity and length of day limits it range.
A second method is to double the chromosome number. A 0.2% colchicines solution was used on the stem tips of Gui Jinqiaomai No.1 (Figure 2) plants for one week. As the new branch became thick, cuttings were used to reproduce a new variety Gui Ai Jinqiaomai No.1 (Figure 3) with dwarf stems, dark green thick leaves and high flavonoid content in leaves, which can be used for leaf tea products and as a vegetables.
In order to improve its seed set and shattering, a tetraploid variety “Hongxin Jinqiao” was developed in the autumn of 2007 by crossing among F. cymosum accessions with less shattering. “Hongxin Jinqiao” has a leaf flavonoid content of about 8% and better seed set in autumn but low seed yield due to some light shattering. We can reproduce the variety by both seed production and shoot cuttings. The variety can be used for vegetable and tea production.
The above perennial buckwheat varieties all have the same key problems: (1) strong shattering resulting in a difficulty in seed harvest; (2) strong sensitivity to light and temperature leading to narrow adaptability, sterility in the spring. (3) out-breeding and low seed set leading to low seed yield and genetic instability. (4) variability in flowering and seeding giving rise to maturity differences.
Cultivated tartary varieties normally can avoid the above problems. It is clear that cultivated tartary buckwheat varieties are one of best parents for improving perennial golden buckwheat by means of wide hybridization.
In order to increase the possibility getting fertile interspecific hybrids, an autotetraploid variety Daku No.1 bred by ourselves was used as female parent and crossed to tetraploid perennial buckwheat variety Hongxin Jinqiaomai on a large scale.
In spring of 2014, we identified a hybrid plant (Figure 4) with good set seed by self-pollination, little shattering, and larger full achenes (1000 grain weight = 38g).
Populations are segregating for shattering, seed size, plant height, style type, flower colour, stem regeneration and perennial habit etc. We currently got 20 F6 lines from the hybrid with three growth seasons a year. These new lines (Figure 5, Figure 6 and Figure 7, Chen, 2016, 2017) [5,14] are different from F. giganteum (Figure 8) on following traits: (1) high plant, 1.0–1.6 m; (2) woody base stem with strong regenerating ability and certain overwintering; (3) larger achenes without groove, 1000 grain weight of up to 30 g or more; (4) non-shattering, up to the yield of 2250 kg/ha or more.
Here, we name a new species, F.tatari-cymosum QF Chen nsp., which may become a new cultivated buckwheat species and form a new production model for buckwheat grain with the property of “plant once and harvest always”. There are currently three perennial tartary buckwheat varieties which have already been spread for next year: Gui Duoku 003, Gui Duoku 60, and Gui Duoku 74, with the yield to be about 3000 kg/ha or more.
The above success of wide hybridization between tartary buckwheat and perennial buckwheat indicated good crossability and compatibility of tartary buckwheat with perennial golden buckwheat, which stimulates us to attempt wider hybridizations. By autumn 2014, we had additional F1 hybrids, including: F.giganteum × F.cymosum Hongxin Jingqiao, F.giganteum × F.megaspartanium (Figure 9), F.giganteum × rice tartary buckwheat, rice tartary buckwheat × F.cymosum Hongxin Jingqiao (Figure 10), F.giganteum × common buckwheat, and wing rice tartary buckwheat rice × F.cymosum.
It is interesting that all hybrids show complete sterile and perennial properties except two crosses (rice tartary buckwheat × F.cymosum Hongxin Jingqiao, and wing rice tartary buckwheat rice × F.cymosum) with annual properties and a high seed-setting percentage (but of reduced size and lacking germinating capacity). All hybrids were crossed with common buckwheat, tartary buckwheat, rice tartary buckwheat, F.megaspartanium accession and F.cymosum Hongxing Jinqiao. However, we could not obtain any fertile seeds except the backcross F.giganteum/F.megaspartanium //F.megaspartanium (Figure 11).
We now have some progenies of the backcross, which show strong perennial properties similar to F.megaspartanium. The above advances will provide a strong support for the buckwheat industry.
6. Product Development and Utilization
6.1. Food Products from Perennial Buckwheat Seeds
The reports regarding buckwheat rice and flour products all concern common buckwheat and tartary buckwheat. Because of shattering and low seed yields, not all perennial buckwheat have industry applications; perennial golden buckwheat has been used for its caudex in Chinese medicine.
Chen (2016, 2017) [5,14] reported a new type of perennial tartary buckwheat—a food crop type. It is an allotetraploid, bred by wide hybridization between annual autotetraploid tartary buckwheat and perennial tetraploid F.cymosum. The flour can be used in a similar fashion as wheat, common buckwheat and tartary buckwheat, etc. There are not, however, any food products on the market, because perennial tartary buckwheat has not yet been produced on a large scale. However, next year, there will be many foods from perennial tartary buckwheat seeds in Guizhou, China.
6.2. Perennial Buckwheat Drink
Common and tartary buckwheat seeds or powder can be used as raw and auxiliary materials in common beverages such as milk beverage, protein beverage, and tea beverage. Due to the higher medicine value and reports on high flavonoid content [63,64,65,66,67,68,69,70], golden buckwheat leaves and caudex has been studied for use such as leaf green tea, leaf red tea, and root slices [54,71,72,73,74]. Huang and Tang (2015) [70] reported the preparation techniques on the golden buckwheat flower tea and its optimal steeping conditions (the rate of tea to water = 1:50, steeping at 90 °C for 30 min, repeated three times).
6.3. Golden Buckwheat Caudex Slices and Chewable Tablets
Golden buckwheat caudex slices are prescribed as Chinese Medicine, for fever reduction and toxin removed, removing abscess and stasis, as a cough expectorant, used in acute lung abscess, acute or chronic bronchitis, bronchial asthma and bacillary dysentery [75]. They are common products in medicinal supermarkets and used as a common tea drink.
A technique for the extraction of the medicinal component(s) from fermented golden buckwheat has allowed for the development of a chewable tablet [76]. This advancement has allowed for greater ease in used of golden buckwheat by public.
6.4. Commercial Medicines from Golden Buckwheat
Commercial medicinal products are manufactured from the underground caudex of golden buckwheat, including syrups, capsules and water aqua. The largest demand for golden buckwheat is for “Acute Bronchitis Syrup”, made by Chongqing Taiji Industry Co. Ltd. (Chongqing, China), a key medicine for acute bronchitis treatment.
“Golden buckwheat capsule” is a traditional Chinese medicine made from Cheng Yun Long’s secret recipe handed down from ancestors via the traditional Chinese medicine hospital of Nantong in Jiangsu. It has been used for 40 years, has unique curative effects on lung abscesses and good curative effects on bronchitis, pneumonia, bacillary dysentery, cholecystitis, and pelvic inflammatory disease, etc. Recently, it has been found to have anticancer activity and obvious clinical effects [77].
“Wei manning” capsule is a new drug in Group II of traditional Chinese medicines, made from extraction and purification of Golden buckwheat rhizome. It has significant curative effects on lung cancer (such as non-small cell lung cancer (NSCLC) at the middle and late period) and decreases the adverse reactions to radiation and chemotherapy [78,79,80].
Tao et al. (2008) [81] reported that the effect of “Golden buckwheat water aqua” from the Golden buckwheat caudex on the cytokines and lung function of chronic obstructive pulmonary disease was better than that of ambroxol; they also discovered the key reason for the improvement of weas and inflammation.
Buckwheat seed sprout, buckwheat leaves and young stem tips from cultivated buckwheat such as common buckwheat and tartary buckwheat are common vegetables in China. They are crisp, tender, and healthy; they contain rich nutrition and healthcare ingredients (such as flavonoids) and few farm chemicals; they are used to make dishes such as vegetable soups, hot foods, fried dish, and cold vegetable dishes in sauce. He et al. (2016) [82] reported the cultivation technique of golden buckwheat to achieve high vegetable quality and yield. The higher vegetable quality of Golden buckwheat suggests great potential for market penetration.
6.5. Forage
Vegetables used for human consumption are often used as forage. High quality vegetative golden buckwheat is an excellent forage for animal production. Golden buckwheat can produce a high quality silage [83] with crude protein content of 21.46%, amino content of 19.2%, ash of 10.41%, calcium of 1.72%, and phosphorus 0.49%, however the low sugar content of 0.6% indicates its best use model mixed with rice (by 30%) or corn (by 40%). Xu et al (2016) [84] discovered that the silage quality of gold buckwheat can be improved by adding lactobacillus and sucrose. Zhang et al (2017) [85] reported that fresh leaves of Golden buckwheat fed to Zangxiang pigs, had greater gains and quality. Golden buckwheat can also replace cabbage as forage for swine.
7. Problems and Prospect
At present, there remain some key unknown problems in the perennial buckwheat field: (1) The inheritance basis with regard to buckwheat perennial properties. (2) Shattering. (3) Low seedset in spring or summer. (4) Key factors in terms of overwintering. (5) Key medicinal compounds and their inheritance; (6) Differential flowering and seed set of branches; (7) The ecosystem properties.
Research on perennial buckwheat has focused on the breeding and quality of the plant materials. Future work will include the use of transcriptomics and genomics to aid in both the breeding of and in the identification and production of medicinal compounds in perennial buckwheat. The growth of the perennial buckwheat industry will be enhanced as these issues are addressed and overcome.
Acknowledgments
The author is grateful to Natural Science Foundation of China (31471562), Earmarked Fund for China Agriculture Research System (CARS-07-A5), Speical Project of Research Center of Buckwheat Engineering and Technology in Guizhou (QianKeHeNong G Zi (2015) 4003), High-level Innovative Talents Training Project in Guizhou (QianKeHeRenCai [2015]4020), and Key Project of Guizhou Agricultural Science Research (QianKeHeNong (2017)005) for providing funds and to the editors for proofing the English.
Author Contributions
Qing-Fu Chen wrote this paper; Xiao-Yan Huang, Hong-You Li, Li-Juan Yang, and Ya-Song Cui prepared the Parts 4 and 6. Authorship must be limited to those who have contributed substantially to the work reported.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chen, Q.F. (Ed.) Plant Sciences on Genus Fagopyrum; China Science Press: Beijing, China, 2012. [Google Scholar]
- Lin, R.F. (Ed.) Buckwheat in China; China Agricultural Science and Technology Press: Beijing, China, 1994. [Google Scholar]
- Liu, D.S.; Xu, R.Y.; Wang, Q.Q. Analysis of protein content and amino acid composition in buckwheat. Crop Germplasm Resour. 1997, 2, 26–28. [Google Scholar]
- Komori, A.; Naoto, I.; Kaori, F.; Shinya, K.; Asakazu, H. Measurement of rutin and quercetin in tartary buckwheat flour by ultraviolet-induced fluorescence. In Proceedings of the 10th International Symposium on Buckwheat, Yang Lin, China, 13–19 August 2007; pp. 403–409. [Google Scholar]
- Chen, Q.F. A study of development strategy about the agricultural production model of modern food crop. J. Mt. Agric. Biol. 2017, 36, 001–005. [Google Scholar]
- Chen, Q.F. A study of resources of Fagopyrum (Polygonaceae) native to China. Bot. J. Linn. Soc. 1999, 130, 53–64. [Google Scholar] [CrossRef]
- Chen, Q.F. Wide hybridization among Fagopyrum (Polygonaceae) species native to China. Bot. J. Linn. Soc. 1999, 131, 177–185. [Google Scholar] [CrossRef]
- Ohnishi, O.; Matsuoka, Y. Search for the wild ancestor of buckwheat. II. Taxonomy of Fagopyrum (Polygonaceae) species based on morphology, isozymes and cpDNA variability. Genes Genet. Syst. 1996, 71, 383–390. [Google Scholar] [CrossRef]
- Ohnishi, O. Discovery of the wild ancestor of common buckwheat. Fagopyrum 1991, 11, 5–10. [Google Scholar]
- Ohnishi, O. Search for the wild ancestor of buckwheat. I. Description of new Fagopyrum (polygonaceae) species and their distribution in China and the Himalayan hills. Fagopyrum 1998, 15, 18–28. [Google Scholar]
- Krotov, A.S.; Dranenko, E.T. An amphidiploid buckwheat, F. giganteum Krotov sp. nova. Byulletenl’ Vsesoyuznogo Ordena Lenina Instituta Rastenievodstva Imeni N.I. Vavilova 1973, 30, 41–45. [Google Scholar]
- Liu, J.N.; Tang, Y.; Xia, M.Z.; Shao, Q.R.; Cai, G.Z.; Luo, Q.; Sun, J.X. A new species of Chinese buckwheat (Polygonaceae)-Fagopyrum densovillosum Liu. Bull. Bot. Res. 2008, 28, 530–533. [Google Scholar]
- Liu, J.N.; Tang, Y.; Xia, M.Z.; Shao, Q.R.; Cai, G.Z.; Luo, Q.; Sun, J.X. A new species of Sichuan polygonaceae buckwheat in China-Fagopyrum crispatifolium Liu. J. Syst. Evol. 2008, 46, 929–932. [Google Scholar]
- Chen, Q.F. Recent progresses on interspecific crossbreeding of genus Fagopyrum Mill. In Proceedings of the 13th International Symposium on Buckwheat (ISB), Cheongju, Korea, 9–11 September 2016. [Google Scholar]
- Li, F.L.; Zeller, F.J.; Huang, K.F.; Shi, T.X.; Chen, Q.F. Improvement of fluorescent chromosome in situ PCR and its application in the phylogeny of the genus Fagopyrum Mill. using nuclear genes of chloroplast origin (cpDNA). Plant Syst. Evol. 2013, 299, 1679–1691. [Google Scholar] [CrossRef]
- Morris, M.R. Cytogenetic studies on buckwheat. J. Hered. 1951, 42, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.S.; Lin, R.F.; Li, Y.Q.; Niu, D.K. Studies of chromosome on different types of buckwheat. Chin. J. Cell Biol. 1984, 6, 130–131. [Google Scholar]
- Chen, Q.F. Karyotype analysis of five buckwheat species (Fagopyrum) native to China. Guihaia 2001, 21, 107–110. [Google Scholar]
- Chen, Q.F.; Hsam, S.L.K.; Zeller, F. A study of cytology, isozyme and interspecific hybridization on the big-achene group of buckwheat species (Fagopyrum, Polygonaceae). Crop Sci. 2004, 44, 1511–1518. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Chen, Q.F. The study of peroxidase isozyme in three leaf stages of genus Fagopyrum. J. Wuhan Bot. Res. 2008, 26, 213–217. [Google Scholar]
- Zhang, Y.Z.; Chen, Q.F. Study on the esterase isozyme of leaf of genus Fagopyrum. J. Wuhan Bot. Res. 2008, 26, 428–432. [Google Scholar]
- Zhang, Y.Z.; Chen, Q.F. The study of the isozyme of the glutamatic transaminase on the germplasm resources of genus Fagopyrum. Seeds 2008, 27, 39–46. [Google Scholar]
- Zhang, Y.Z.; Chen, Q.F. The study of seed peroxidase isozyme of germplasm resources of genus Fagopyrum. Guihaia 2008, 28, 553–557. [Google Scholar]
- Zhang, Y.Z.; Chen, Q.F. Studies on esterase isozyme of young leaves in wild buckwheat plants. Seeds 2009, 28, 41–43. [Google Scholar]
- Guo, Y.Z.; Chen, Q.F.; Yang, L.Y.; Huang, Y.H. Analyses of the seed protein contents on the cultivated and wild buckwheat resources. Genet. Resour. Crop Evol. 2007, 54, 1465–1472. [Google Scholar] [CrossRef]
- Li, J.H.; Chen, Q.F.; Zeller, F.J. Variation in seed protein subunits among species of the genus Fagopyrum Mill. Plant Syst. Evol. 2008, 273, 192–202. [Google Scholar] [CrossRef]
- Ohsako, T.; Ohnishi, O. Intra- and interspecific phylogeny of wild Fagopyrum (Polygonaceae) species based on nucleotide sequences of noncoding regions in chloroplast DNA. Am. J. Bot. 2000, 87, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.J.; Chen, Q.F. Study on RAPD of genus Fagopyrumresources. Seed 2009, 28, 37–48. [Google Scholar]
- Yang, X.Y.; Wu, Z.F.; Chen, H.; Shao, J.-R.; Wu, Q. Karyotype and genetics relationship based on RAPD markers of six wild buckwheat species (Fagopyrum ssp.) from southwest of China. Genet. Resour. Crop Evol. 2010, 57, 649–656. [Google Scholar] [CrossRef]
- Zhang, C.P.; He, P.; He, J.X.; Lei, S.T.; Hu, S.J. ISSR Analysis on genetic diveristy of Fagopyrum cymosum collected from eight wild populations. Chin. Tradit. Herb. Drugs 2010, 41, 1519–1523. [Google Scholar]
- Wu, C.S.; Wang, Y.N.; Liu, S.M.; Chaw, S.M. Chloroplast genome (cpDNA) of Cycastaitungensis and 56 cp protein-coding genes of gnetumparvifolium: Insights into cpDNA evolution and phylogeny of extant seed slants. Mol. Biol. Evol. 2007, 24, 1366–1379. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yu, S.; Chen, Q.F. Sequence analysis of F 3′ 5′ H from Fagopyrum cymosum. Seed 2014, 33, 6–10. [Google Scholar]
- Li, G.; Yu, S.; Chen, Q.F. Molecular cloning and sequences analysis of TC from Fagopyrum cymosum. Seed 2015, 34, 1–4. [Google Scholar]
- Meng, H.; Li, C.L.; Wu, Q.; Shao, J.R.; Chen, H. Cloning and sequence analysis of the chalcone synthase gene (CHS) from Fagopyrum dibotrys. Acta Pratacult. Sin. 2010, 19, 162–169. [Google Scholar]
- Bu, X.X.; Luo, X.P.; Bai, Y.C.; Li, C.L.; Chen, H.; Wu, Q. Gene cloning of anthocyanin synthase in Fagopyrumdibotrys and correlation between its expression level and anthocyan in content. Chin. Tradit. Herb. Drugs 2014, 45, 985–989. [Google Scholar]
- Li, C.L.; Feng, Z.Y.; Bai, Y.C.; Chen, H.; Zhao, H.X.; Wu, Q. Molecular cloning and prokaryotic expression of phenylalanine ammonialyase gene FdPAL from Fagopyrum dibotrys. China J. Chin. Mater. Med. 2011, 36, 3238–3243. [Google Scholar]
- Liang, C.G.; Chen, Q.Q.; Shi, T.X.; Chen, Q.J.; Chen, Q.F. Sequence analysis of MAPK gene fragment and phylogenetic relationship of genus Fagopyrum. Acta Agric. Zhejiangensis 2016, 28, 1631–1636. [Google Scholar]
- Li, R.Y.; Pan, F.; Chen, Q.F.; Shi, T.X. Excavation and polymorphism analysis of EST-SSR from transcriptorne of tartary buckwheat. J. Agric. Sci. Technol. 2015, 17, 42–52. [Google Scholar]
- Yasui, Y.; Hirakawa, H.; Matsui, K.; Katsube-Tanaka, T.; Yang, S.J.; Aii, J.; Sato, S.; Mori, M. Construction of buckwheat genome database (BGDB). In Proceedings of the 13th International Symposium on Buckwheat (ISB), Cheongju, Korea, 9–11 September 2016; pp. 205–206. [Google Scholar]
- Zhang, L.J.; Li, X.X.; Ma, B.; Gao, Q.; Du, H.; Han, Y.; Li, Y.; Cao, Y.; Qi, M.; Zhu, Y.; et al. The tartary buckwheat genome provides Insights into rutin biosynthesis and abiotic stress tolerance. Mol. Plant 2017, 10, 1224–1237. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.S.; Yi, F. A review of researches on chemical components and active substances of wild resource plant Fagopyrum cymosum (Trev.) Meism. J. Anhui Agric. Sci. 2013, 41, 3379–3381. [Google Scholar]
- Chen, X.F.; Gu, Z.L. Study of anti-tumor effect on Fagopyrum cymosum. Chin. Tradit. Herb. Drugs 2000, 31, 715–718. [Google Scholar]
- Liu, Y.L.; Fang, Q.N.; Zhang, X.Q.; Feng, X.X.; Zhang, L.F.; He, X.W. Studies on the active ingredients of Fagopyrum cymosum. Chin. Pharm. J. 1980, 15, 40–41. [Google Scholar]
- Zhang, W.J.; Li, X.C.; Liu, Y.Q.; Yao, R.C.; Nonaka, G.I. Phenolic constituents from Fagopyrum dibotrys. Acta Botanica Yunnanica. 1994, 16, 354–356. [Google Scholar]
- Shao, M.; Yang, Y.H.; Gao, H.Y.; Wu, B.; Wang, L.B.; Wu, L.J. Chemical constituents from Elsholtzia blanda. China J. Chin. Mater. Med. 2005, 30, 1591–1593. [Google Scholar]
- Shao, M.; Yang, Y.H.; Gao, H.Y.; Wu, B.; Wang, L.B. Study on the chemical constituents of Fagopyrum cymosum. J. Shenyang Pharm. Univ. 2005, 22, 100–102. [Google Scholar]
- Wang, K.J.; Zhang, Y.J.; Yang, C.R. Antioxidant phenolic constituents from Fagopyrum dibotrys. J. Ethnopharmacol. 2005, 99, 259. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Z.; Zhou, J.Y.; Pan, H.L. Study on bacteriostasis of Fagopyrum cymosum Meisn. Chin. J. Microecol. 2005, 8, 330–331. [Google Scholar]
- Wu, H.Z.; Zhou, J.Y.; Pan, H.L. Study on chemical constituents of Fagopyrum dibotrys (D.Don) Hara. Chin. Hosp. Pharm. J. 2008, 28, 1829–1831. [Google Scholar]
- Bai, Z.Z.; Sun, H.; Cao, F.; Mu, F.K. Analysis of volatile oil from Fagopyrum dibotrys (D.Don) Hara. by GC/MS. Chin. J. Pharm. Anal. 2007, 11, 1832–1835. [Google Scholar]
- Huang, X.Y.; Wang, J.Y.; Chen, Q.F. Golden buckwheat (Fagopyrum cymosum) leaf tea function and mechanism of resistance to type II diabetes. Lishizhen Med. Mater. Med. Res. 2014, 6, 1334–1337. [Google Scholar]
- Huang, X.Y.; Huang, S.; Chen, Q.F. Antineoplastic function and mechanism of golden buckwheat leaf fermented tea in the mice of H22 tumor. J. Anhui Agric. Univ. 2015, 42, 854–859. [Google Scholar]
- Huang, X.Y.; Huang, S.; Chen, Q.F. Antineoplastic function and mechanism of golden buckwheat leaf fermented tea. Mod. Tradit. Chin. Med. Mater. Med. World Sci. Technol. 2015, 5, 981–984. [Google Scholar]
- Huang, S.; Wang, J.Y.; Chen, Q.F.; Huang, X.Y. Antioxidant activity of the active components of the buckwheat leaf tea. Food Oil 2016, 2, 30–32. [Google Scholar]
- Minami, M.; Gomi, M.; Ujihara, A. Pollen tube growth in interspecific crosses in Fagopyrum. J. Fac. Agric. Shinshu Univ. 1992, 29, 129–135. [Google Scholar]
- Hirose, T.; Ujihara, A.; Kitayashi, H.; Minami, M. Interspecific cross-compatibility in Fagopyrum according to pollen tube growth. Breed. Sci. 1994, 44, 307–314. [Google Scholar] [CrossRef]
- Hirose, T.; Ujihara, A.; Kitayashi, H.; Minami, M. Pollen tube behaviour related to self-incompatibility in interspecific crosses of Fagopyrum. Breed. Sci. 1995, 45, 65–70. [Google Scholar]
- Kachonpadungkitti, Y.; Mangkita, W.; Romchatngoen, S.; Hasegawa, K.; Hisajima, S. Possibility of cross breeding of buckwheat (Fagopyrum esculentum Moench) in vitro. Shokubutsu Kojo Gakkaishi 2003, 15, 98–101. [Google Scholar] [CrossRef]
- Ujihara, A.Y.; Nakamura, Y.; Minami, M. Interspecific hybridization in genus Fagopyrum. Properties of hybrids (F. esculentum × F. cymosum) through ovule culture. Gamma Field Symp. 1990, 29, 33–45. [Google Scholar]
- Samimy, C. Barrier to interspecific crossing of Fagopyrum esculentum with F. tataricum. I. Site of pollen tube arrest. II. Organogenesis from immature embryos of F. tataricum. Euphytica 1991, 54, 215–219. [Google Scholar]
- Lee, B.S.; Ujihara, A.; Minami, M.; Hirose, T. Breeding of interspecific hybrids in genus Fagopyrum. 4. Production of interspecific hybrid ovules culture among F. esculentum, F. tataricum and F. cymosum. Breed. Sci. 1994, 44 (Suppl. 1), 183. [Google Scholar]
- Nóra, M.D.; Andrew, J.C.; Judit, D. Progress and prospects for interspecific hybridization in buckwheat and the genus Fagopyrum. Biotechnol. Adv. 2013, 31, 1768–1775. [Google Scholar]
- Feng, X.Y.; Chen, Q.F. Determination of total flavones content in different organs of F. megaspartanium. Guizhou Agric. Sci. 2007, 35, 15–16. [Google Scholar]
- Liu, N.; Zeller, F.J.; Chen, Q.F. The flavonoid content in leaves and inflorescences of the wild perennial Fagopyrum cymosum complex. Genet. Resour. Crop Evol. 2013, 60, 825–838. [Google Scholar] [CrossRef]
- Li, G.; Yu, S.; Zhou, Y.H.; Chen, Q.F. Spectrophotometric determination of flavonoids content in leaves of Fagopyrum cymosum complex. Asian J. Chem. 2013, 25, 7575–7578. [Google Scholar]
- Li, G.; Yu, S.; Zhou, Y.H.; Chen, Q.F. Analysis of the antioxidant related substances in leaves of the Fagopyrum cymosum complex and common buckwheat plants. Guangdong Agric. Sci. 2013, 40, 7–11. [Google Scholar]
- Li, G.; Yu, S.; Chen, Q.F. Research status of antioxidative substances from buckwheat. Ningxia J. Agric. For. Sci. Technol. 2014, 2, 87–89. [Google Scholar]
- Li, G.; Yu, S.; Deng, Y.; Zhou, Y.H.; Chen, Q.F. Study on extraction technology of flavonoids from Golden Buckwheat leaves. Jiangsu Agric. Sci. 2013, 41, 264–266. [Google Scholar]
- Shi, T.X.; Gu, L.L.; Chen, Z.L.; Chen, Q.F. Analysis of flavonoids, soluble protein and soluble sugar content in leaves of Fagopyrum tataricum. Jiangsu Agric. Sci. 2014, 42, 252–255. [Google Scholar]
- Huang, M.J.; Tang, H.M. The preliminary study of Fagopyrum dibotrys scented tea’s manufacture crafts. Food Res. Dev. 2015, 6, 53–56. [Google Scholar]
- Gu, L.L.; Chen, Q.F. A study on brewing condition of golden buckwheat functional leaf tea. Food Res. Dev. 2014, 3, 59–64. [Google Scholar]
- Gu, L.G.; Huang, X.Y.; Li, Y.; Chen, Q.F. Production technology optimization on fermented tea of golden buckwheat. Guizhou Agric. Sci. 2014, 3, 59–64. [Google Scholar]
- Gu, L.L.; Chen, Q.F. Variation of flavonoid and GABA content in leaves among different golden buckwheat accessions. Southwest China J. Agric. Sci. 2014, 27, 582–586. [Google Scholar]
- Liu, N.; Chen, Q.F. Research on the making technology of golden buckwheat green tea. J. Yunnan Agric. Univ. (Nat. Sci.) 2008, 23, 76–79. [Google Scholar]
- He, M.S.; Qian, B.H.; Wang, Z.L.; Wang, Z.Y.; Yan, Y.M. Quality standard for Jinqiaomai Tablets. Chin. Tradit. Pat. Med. 2010, 32, 779–782. [Google Scholar]
- Hu, T.; Huang, K.F.; Huang, X.Y.; Chen, Q.F. Preparation process of chewable tablet of a fermented tea from golden buckwheat (Fagopyrum cymosum) comeplex leaf. J. Anhui Agric. Sci. 2017, 44, 27–32. [Google Scholar]
- Xu, Y.; Tong, Y.H.; Qiu, H.G. Preparation of buckwheat capsule and quality standards. Lishizhen Med. Mater. Med. Res. 2004, 15, 753–754. [Google Scholar]
- Huang, J.W.; Zhao, D. Analysis of the efficacy of Weimaining capsule combined with chemotherapy in the advanced non-small cell lung cancer. Asia-Pac. Tradit. Med. 2015, 11, 125–126. [Google Scholar]
- Zhang, M.; Gao, Z.; Yang, Q.J.; Qian, Y.; Li, D.; Tan, G.F.; Zhang, Y. Clinical observation of Weimaining capsule combined with bronchial artery infusion chemotherapy in the treatment of advanced non-small cell lung cancer. Chin. J. Clin. Oncol. Rehabilit. 2013, 10, 1114–1116. [Google Scholar]
- Zhao, S.Q.; Xu, C.Q.; Chen, X.H. Effect of Weimaining capsule in treating non-small cell lung cancer and its effect on serum epidermal growth factor receptor. Chin. J. Gerontol. 2014, 18, 5070–5071. [Google Scholar]
- Tao, Z.Q.; Gao, X.; Tang, Y.F.; Sun, J.; Zhang, Y.; You, J.S.; Ni, W.B.; Yu, H.Z. Effect of buckwheat liquid on serum cytokines and lung function in patients with chronic obstructive pulmonary disease. Liaoning J. Tradit. Chin. Med. 2008, 35, 332–333. [Google Scholar]
- He, Y.K.; Zhao, W.; Ma, J.; Mian, J.Y.; Cheng, N.; Deng, L.S.; Zhao, M.Y. Optimization of high yield cultivation technique in Fagopyrum dibotrys. Guizhou Agric. Sci. 2016, 44, 52–55. [Google Scholar]
- Deng, R.; Zhang, D.H.; Wang, A.N.; Xiang, Q.H.; Luo, Q.H.; Zhang, L. Study on silage experiment of Golden buckwheat. Caoye Yu Xumu 2012, 9, 5–7. [Google Scholar]
- Xu, L.X.; Xiong, K.N.; Yang, S.M.; Guo, W.; Liu, K.X. The effect of quality of lactobacillus and sucrose on the silage of golden buckwheat. Anim. Husb. Vet. Med. 2016, 48, 54–59. [Google Scholar]
- Zhang, J.; Deng, R.; Xu, Z.H.; Li, L.N.; Zhang, D.H. Gaining effect of bean zangxiang with fresh stem leaves of Qian Fagopyrum dibotrys. Swine Prod. 2017, 3, 17–18. [Google Scholar]
- Ruan, Y.; Ji, X.Q.; Xia, X.L.; Wen, M.; Li, S.J.; Liu, Y.J.; Chen, J.Q. Effect of golden buckwheat on the immunity effect of duck plague seedlings. Heilongjiang Anim. Sci. Vet. Med. 2015, 10, 203–205. [Google Scholar]
- Ruan, Y.; Ji, X.Q.; Xia, X.L.; Wen, M.; Li, S.J.; Liu, T.J.; Chen, J.Q. Effects of Fago-con immune effects of avian influenza vaccine for ducks. Guangdong Agric. Sci. 2015, 13, 119–122. [Google Scholar]
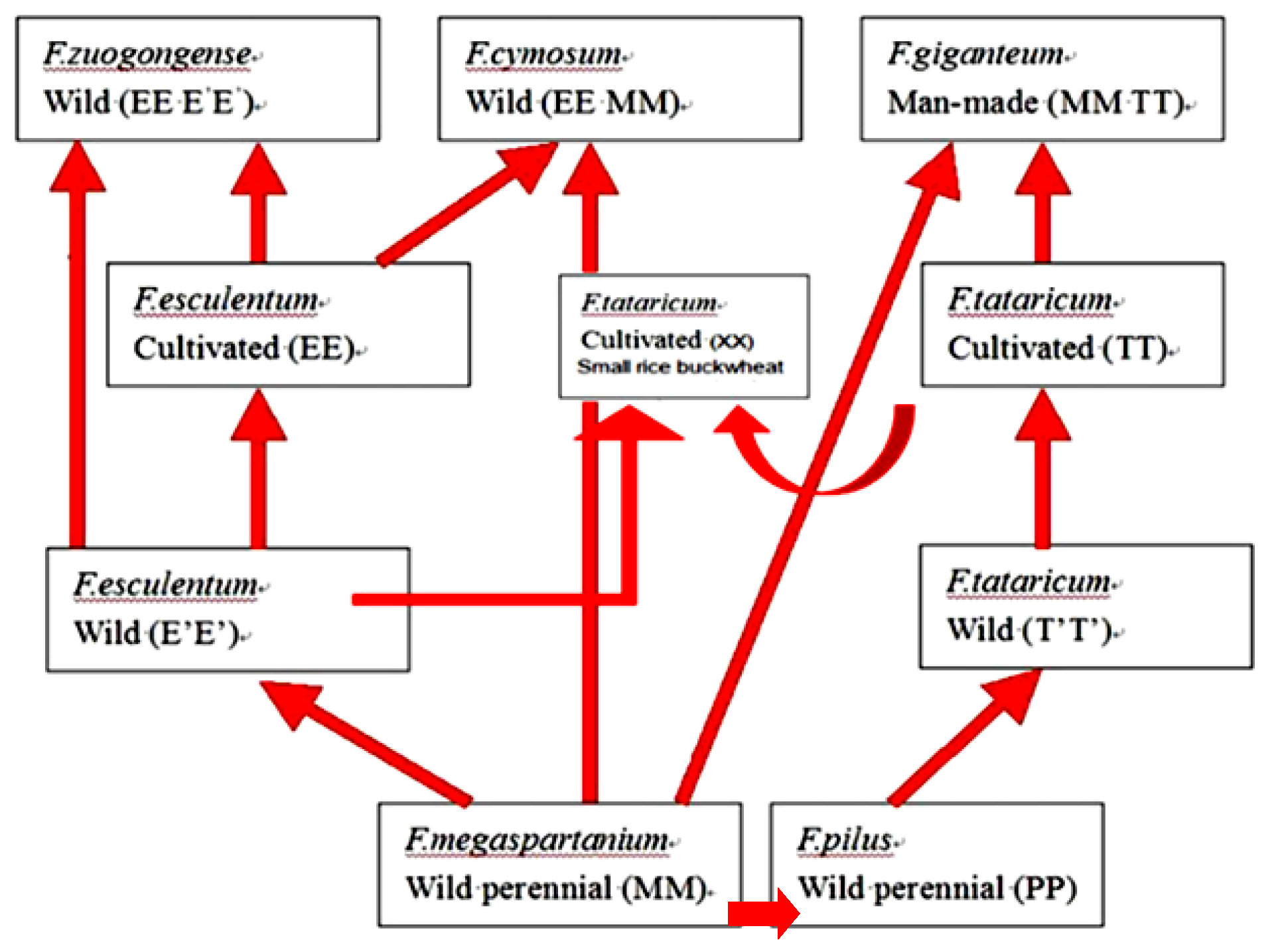
Figure 1.
A diagram of possible relationships among seven buckwheat species in the big-achene group (Li et al. [15]).
Figure 1.
A diagram of possible relationships among seven buckwheat species in the big-achene group (Li et al. [15]).
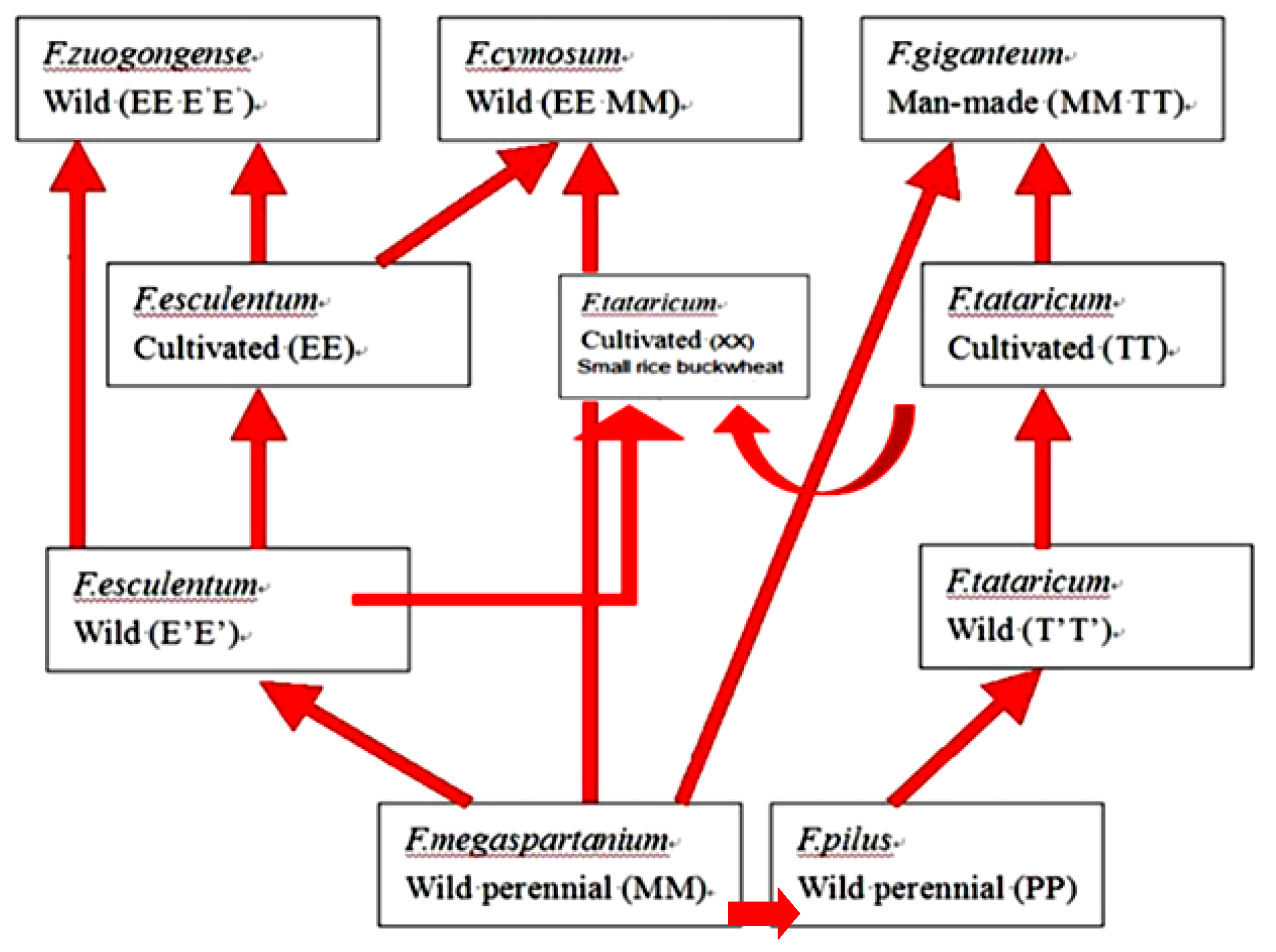
Figure 2.
Plant (a) and inflorescence (b) of Gui Jinqiaomai No.1, a perennial Golden buckwheat variety (F.megaspartanium).
Figure 2.
Plant (a) and inflorescence (b) of Gui Jinqiaomai No.1, a perennial Golden buckwheat variety (F.megaspartanium).

Figure 3.
Plant (a) and inflorescence (b) of Gui Ai JinqiaomaiNo.1, a dwarf perennial Golden buckwheat variety (F.megaspartanium).
Figure 3.
Plant (a) and inflorescence (b) of Gui Ai JinqiaomaiNo.1, a dwarf perennial Golden buckwheat variety (F.megaspartanium).

Figure 4.
The comparision of morphology among autotetraploid tartary buckwheat “Daku No.1”, tetraploid perennial Golden buckwheat variety HongxinJinqiaomai; and their hybrid. (a) tetraploid tartary buckwheat plant; (b) hybrid plant between tartary buckwheat and golden buckwheat; (c) golden buckwheat plant; (d) tetraploid tartary buckwheat inflorescence; (e) hybrid plant inflorescence between tartary buckwheat and golden buckwheat; (f) inflorescence of golden buckwheat.
Figure 4.
The comparision of morphology among autotetraploid tartary buckwheat “Daku No.1”, tetraploid perennial Golden buckwheat variety HongxinJinqiaomai; and their hybrid. (a) tetraploid tartary buckwheat plant; (b) hybrid plant between tartary buckwheat and golden buckwheat; (c) golden buckwheat plant; (d) tetraploid tartary buckwheat inflorescence; (e) hybrid plant inflorescence between tartary buckwheat and golden buckwheat; (f) inflorescence of golden buckwheat.

Figure 5.
A perennial tartary buckwheat variety—Guiduoku003. (A) plant; (B) inflorescence; (C) Fruit branch; (D) achenes [14].
Figure 5.
A perennial tartary buckwheat variety—Guiduoku003. (A) plant; (B) inflorescence; (C) Fruit branch; (D) achenes [14].

Figure 6.
A perennial tartary buckwheat variety—Gui Duoku60. (A) plant; (B) inflorescence; (C) Fruit branch; (D) achenes [14].
Figure 6.
A perennial tartary buckwheat variety—Gui Duoku60. (A) plant; (B) inflorescence; (C) Fruit branch; (D) achenes [14].

Figure 7.
Field of Gui Duoku No.003, a perennial tartary buckwheat variety.

Figure 8.
A comparison of perennial F.tatari-cymosum and annual F.giganteum plants [14]. (a) F.tatari-cymosum plant; (b) F.giganteum plant.
Figure 8.
A comparison of perennial F.tatari-cymosum and annual F.giganteum plants [14]. (a) F.tatari-cymosum plant; (b) F.giganteum plant.

Figure 9.
F1 perennial hybrid of the cross F.giganteum/F.megaspartanium [14].
Figure 9.
F1 perennial hybrid of the cross F.giganteum/F.megaspartanium [14].

Figure 10.
F1 annual hybrid plant of rice tartary buckwheat/F.cymosum and their inflorescence and shrink seeds [14]; (a) hybirid plant; (b) inflorescence; (c) shrink seeds.
Figure 10.
F1 annual hybrid plant of rice tartary buckwheat/F.cymosum and their inflorescence and shrink seeds [14]; (a) hybirid plant; (b) inflorescence; (c) shrink seeds.

Figure 11.
F1 perennial hybrid plants of the cross F.giganteum/F.megaspartanium//F.megaspartanium [14]; (a) plant; (b) inflorescence.
Figure 11.
F1 perennial hybrid plants of the cross F.giganteum/F.megaspartanium//F.megaspartanium [14]; (a) plant; (b) inflorescence.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The classification and key properties of the genus Fagopyrum (Chen, 2012) [1].
Table 1.
The classification and key properties of the genus Fagopyrum (Chen, 2012) [1].
| Section | Species | Key Distribution | Annual/Perennial | Ploidy | Genome | Properties |
|---|---|---|---|---|---|---|
| Big-achene group | 1. Common buckwheat, F.esculentum Moench | Worldwide | Annual | 2x = 16, 4x = 32 | 2x = 16, EE 4x = 32, EEEE | Big flower and achene, cross-pollination |
| 2. Tartary buckwheat, F.tataricum (Linnaeus) Gaertner | Southwest China | Annual | 2x = 16, 4x = 32 | 2x = 16, TT; 4x = 32, TTTT | Small green flower and bigger achene, self-pollination | |
| 3. F.megaspartanium Chen | South China | Perennial | 2x = 16 | 2x = 16, MM | F.cymosum complex. Big flower and achene, cross-pollination, bulbous caudex | |
| 4. F.pilus Chen | Tibet, China | Perennial | 2x = 16, | 2x = 16, PP | F.cymosum complex. Bigger flower and achene, cross-pollination, bulbous caudex | |
| 5. F.cymosum (Trev.) Meisner | Southwest China | Perennial | 4x = 32 | 4x = 32, MMXX | F.cymosum complex. Bigger flower and achene, cross-pollination, with subsurface transverse stem | |
| 6. F.zengongense QF Chen | Tibet, China | Annual | 4x = 32 | 4x = 32, EEE’E’ | Bigger flower and achene, self or cross-pollination | |
| 7. F.giganteum Krotov | Man-made | Annual | 4x = 32 | 4x = 32, TTXX | Bigger flower and big achene, cross or self-pollination | |
| 8. F.tatari-cymosum Chen | Man-made | Perennial | 4x = 32 | 4x = 32, TTXX | Bigger flower and big achene, self-pollination | |
| Small-achene group | 1. F.gracilipes (Hemsley) Dammer ex Diels | Southwest China | Annual | 4x = 32 | / | small flower and achene, self-pollination |
| 2. F.leptopodum (Diels) Hedberg | Southwest China | Annual | 2x = 16 | / | small flower and achene, cross or self-pollination | |
| 3. F.gilesii (Hemsley) Hedberg | Southwest China | Annual | 2x = 16 | / | small flower and achene, cross or self-pollination | |
| 4. F.lineare (Samuelsson) Haraldson | Yunnan, China | Annual | 2x = 16 | / | small flower and achene, cross or self-pollination | |
| 5. F.urophyllum (Bureau et Franch) Gross | Southwest China | Perennial | 2x = 16 | / | small flower and achene, cross pollination, perennial sub-shrubs | |
| 6. F.statice (Lev.) Gross | Southwest China | Perennial | 2x = 16 | / | small flower and achene, cross pollination, perennial herbs | |
| 7. F.caudatum (Samuelsson) Li | Southwest China | Annual | / | / | small flower and achene, cross or self-pollination | |
| 8. F.pleioramosum Ohnishi | Southwest China | Annual | 2x = 16 | / | small flower and achene, cross or self-pollination | |
| 9. F.capillatum Ohnishi | Southwest China | Annual | / | / | small flower and achene, cross or self-pollination | |
| 10. F.calliansum Ohnishi | Southwest China | Annual | / | / | small flower and achene, cross or self-pollination | |
| 11. F.macrocarpum Ohsako & Ohnishi | Southwest China | Annual | 2x = 16 | / | small flower and achene, cross or self-pollination | |
| 12. F.gracilipedoides Ohsako& Ohnishi | Southwest China | Annual | / | / | small flower and achene, cross or self-pollination | |
| 13. F.jinshaense Ohsako& Ohnishi | Southwest China | Annual | / | / | small flower and achene, cross or self-pollination | |
| 14. F.rubifolium Ohsako& Ohnishi | Southwest China | Annual | / | / | small flower and achene, cross or self-pollination | |
| 15. F.crispatifolium Liu | Southwest China | Annual | / | / | small flower and achene, cross or self-pollination | |
| 16. F.densiovillosumLiu | Southwest China | Annual | 16 | / | small flower and achene, cross or self-pollination |
Note: X is an unknown number.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, Q.-F.; Huang, X.-Y.; Li, H.-Y.; Yang, L.-J.; Cui, Y.-S. Recent Progress in Perennial Buckwheat Development. Sustainability 2018, 10, 536. https://doi.org/10.3390/su10020536
AMA Style
Chen Q-F, Huang X-Y, Li H-Y, Yang L-J, Cui Y-S. Recent Progress in Perennial Buckwheat Development. Sustainability. 2018; 10(2):536. https://doi.org/10.3390/su10020536
Chicago/Turabian StyleChen, Qing-Fu, Xiao-Yan Huang, Hong-You Li, Li-Juan Yang, and Ya-Song Cui. 2018. "Recent Progress in Perennial Buckwheat Development" Sustainability 10, no. 2: 536. https://doi.org/10.3390/su10020536
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.