
In Vitro Synthesized RNA Generated from cDNA Clones of Both Genomic Components of Cucurbit yellow stunting disorder virus Replicates in Cucumber Protoplasts


 and
and 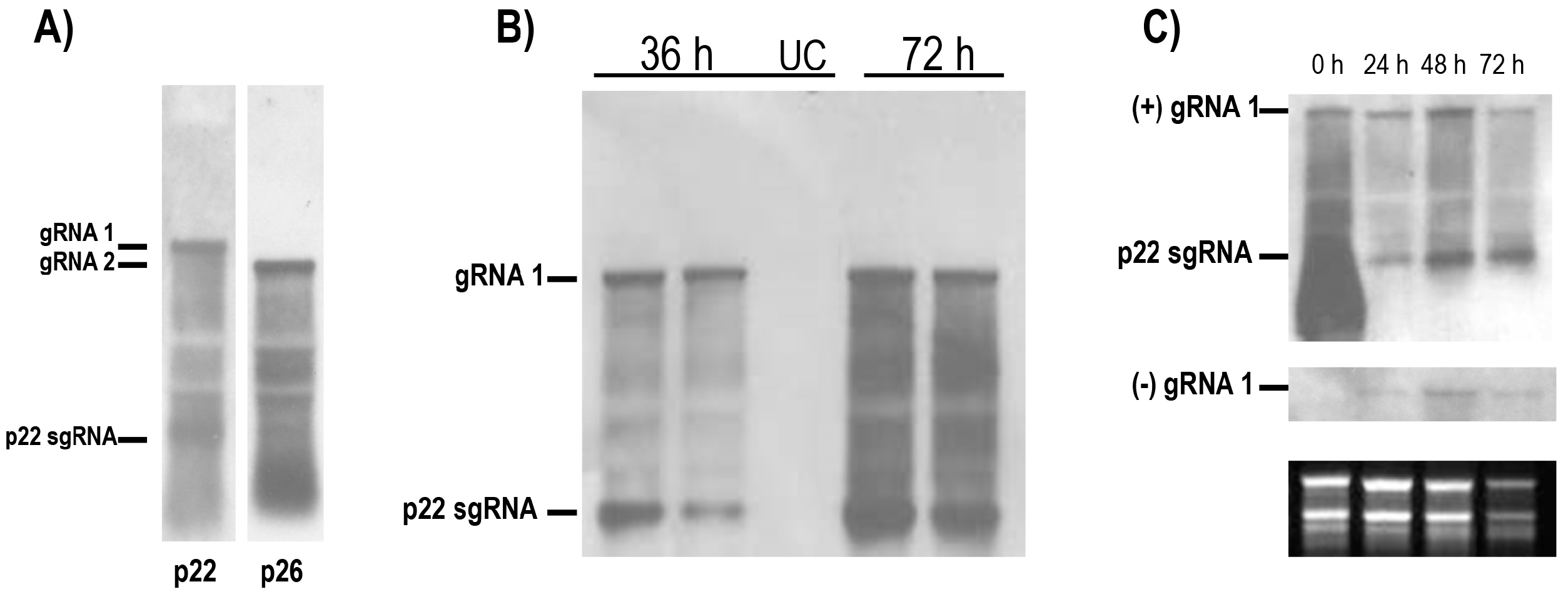{kind=link}
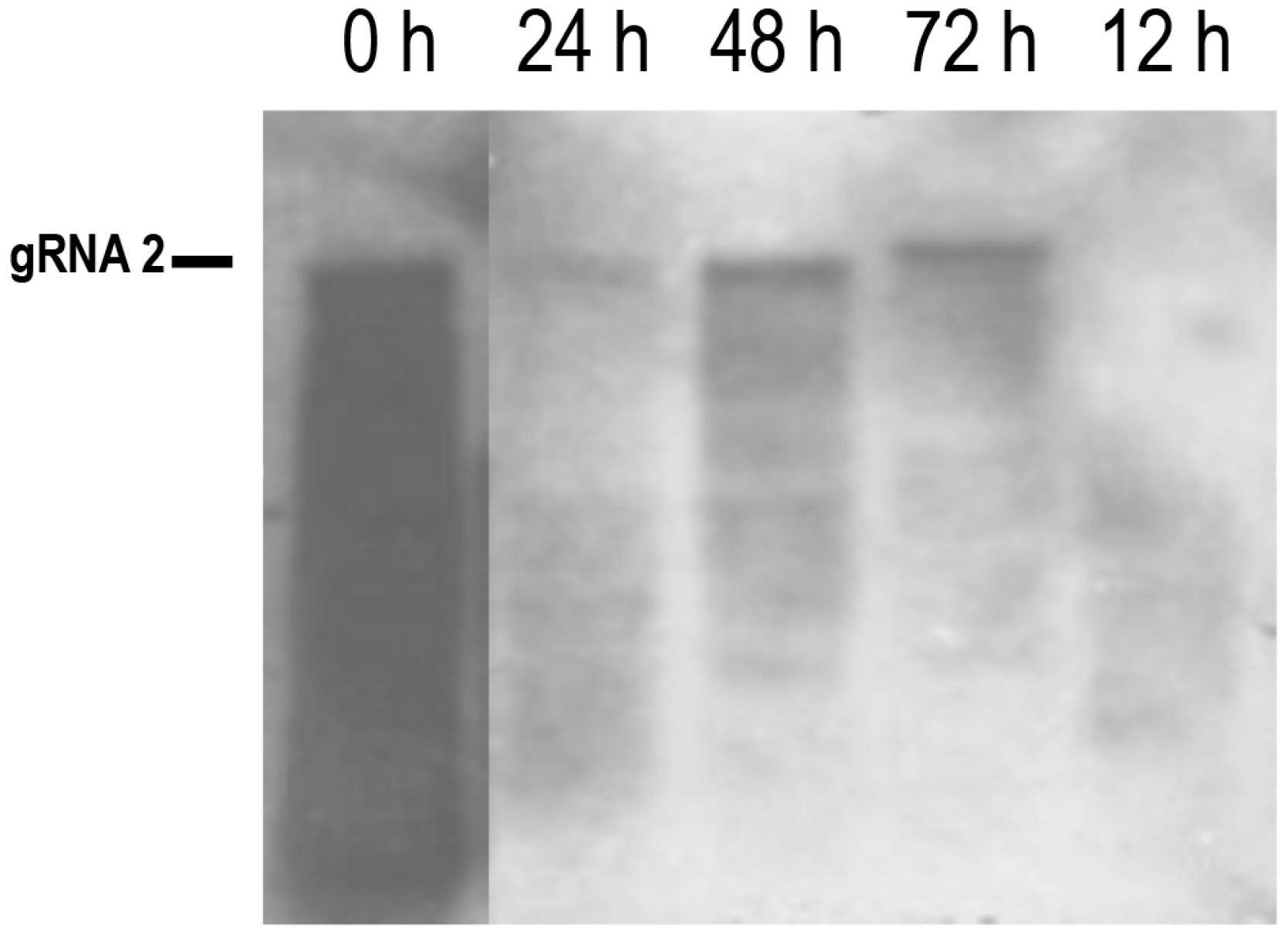{kind=link}
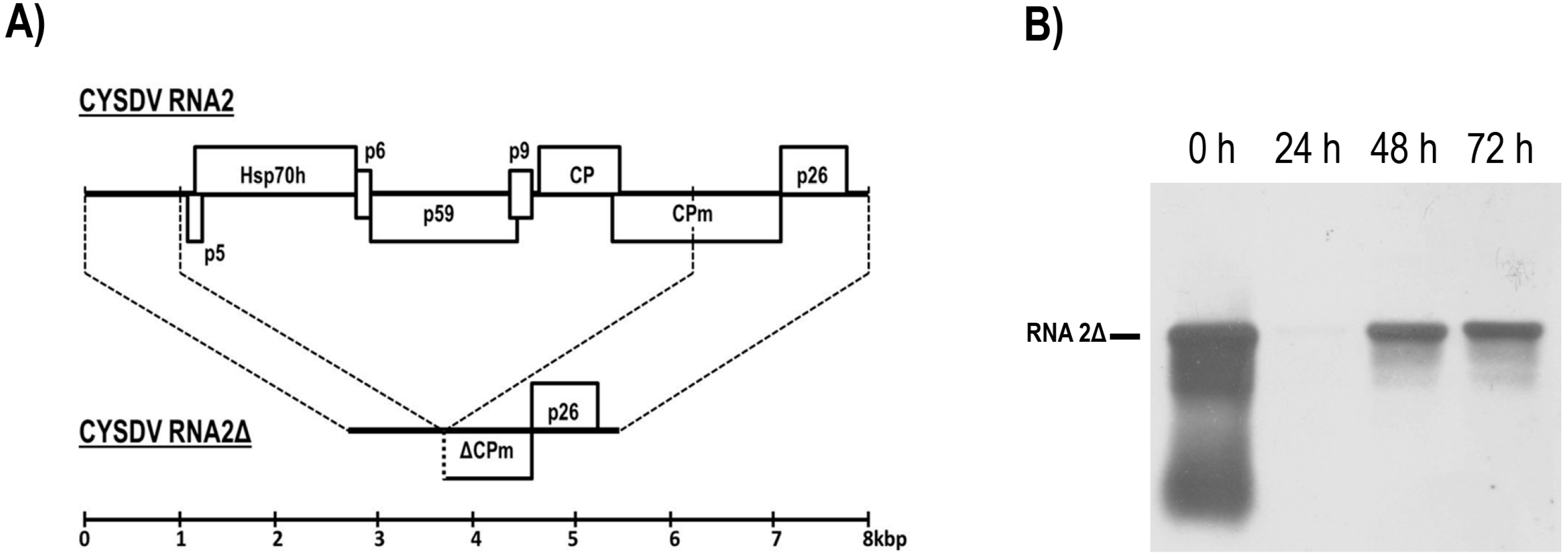{kind=link}
Abstract
:Supplementary Materials
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Navas-Castillo, J.; Lopez-Moya, J.J.; Aranda, M.A. Whitefly-transmitted RNA viruses that affect intensive vegetable production. Ann. Appl. Biol. 2014, 165, 155–171. [Google Scholar]
- Tzanetakis, I.E.; Martin, R.R.; Wintermantel, W.M. Epidemiology of criniviruses: An emerging problem in world agriculture. Front. Microbiol. 2013, 4, 193–207. [Google Scholar]
- Abrahamian, P.E.; Abou-Jawdah, Y. Whitefly transmitted criniviruses of cucurbits: Current status and future prospects. Virus Dis. 2014, 5, 26–38. [Google Scholar]
- Klaassen, V.A.; Mayhew, D.; Fisher, D.; Falk, B.W. In vitro transcripts from cloned cDNAs of the Lettuce infectious yellows Closterovirus bipartite genomic RNAs are competent for replication in Nicotiana benthamiana protoplasts. Virology 1996, 222, 169–175. [Google Scholar]
- Wang, J.; Turina, M.; Stewart, L.R.; Lindbo, J.A.; Falk, B.W. Agroinoculation of the Crinivirus Lettuce infectious yellows virus, for systemic plant infection. Virology 2009, 392, 131–136. [Google Scholar]
- Kiss, Z.A.; Medina, V.; Falk, B.W. Crinivirus replication and host interactions. Front. Microbiol. 2013, 4, 172–182. [Google Scholar]
- Yeh, H.H.; Tian, H.; Rubio, L.; Crawford, B.; Falk, B.W. Asynchronous accumulation of Lettuce infectious yellows virus RNAs 1 and 2 and identification of an RNA 1 trans enhancer of RNA 2 accumulation. J. Virol. 2000, 74, 5762–5768. [Google Scholar]
- Wang, J.; Stewart, L.R.; Kiss, Z.; Falk, B.W. Lettuce infectious yellows virus (LIYV) RNA 1-encoded P34 is an RNA-binding protein and exhibits perinuclear localization. Virology 2010, 403, 67–77. [Google Scholar]
- Tian, T.; Rubio, L.; Yeh, H.H.; Crawford, B.; Falk, B.W. Lettuce infectious yellows virus: In vitro acquisition analysis using partially purified virions and the whitefly Bemisia tabaci. J. Gen. Virol. 1999, 80, 1111–1117. [Google Scholar]
- Chen, A.Y.; Walker, G.P.; Carter, D.; Ng, J.C.K. A virus capsid component mediates virion retention and transmission by its insect vector. Proc. Natl. Acad. Sci. USA 2011, 108, 16777–16782. [Google Scholar]
- Stewart, L.R.; Medina, V.; Tian, T.; Turina, M.; Falk, B.W.; Ng, J.C. A mutation in the Lettuce infectious yellows virus minor coat protein disrupts whitefly transmission but not in planta systemic movement. J. Virol. 2010, 84, 12165–12173. [Google Scholar]
- Chen, A.Y.S.; Patrivin, A.; Ng, J.C.K. Agroinoculation of the cloned infectious cDNAs of Lettuce chlorosis virus results in systematic plant infection and production of whitefly transmissible virions. Virus Res. 2012, 169, 310–315. [Google Scholar]
- Mongkolsiriwattana, C.; Chen, A.Y.S.; Ng, J.C.K. Replication of Lettuce chlorosis virus (LCV), a crinivirus in the family Closteroviridae, is accompanied by the production of LCV RNA 1-derived novel RNAs. Virology 2011, 420, 89–97. [Google Scholar]
- Orilio, A.F.; Fortes, I.M.; Navas-Castillo, J. Infectious cDNA clones of the crinivirus Tomato chlorosis virus are competent for systemic plant infection and whitefly-transmission. Virology 2014, 464, 365–374. [Google Scholar]
- Mathioudakis, M.M.; Rodríguez-Moreno, L.; Sempere, R.N.; Aranda, M.A.; Livieratos, I. Multifaceted capsid proteins: Multiple interactions suggest multiple roles for Pepino mosaic virus capsid protein. Mol. Plant-Microbe Interact. 2014, 12, 1356–1369. [Google Scholar]
- Navas-Castillo, J.; Albiach-Martí, M.R.; Gowda, S.; Hilf, M.E.; Garnsey, S.M.; Dawson, W.O. Kinetics of accumulation of Citrus tristeza virus RNAs. Virology 1997, 228, 92–97. [Google Scholar]
- Kreuze, J.F.; Savenkov, E.I.; Cuellar, W.; Li, X.; Valkonen, J.P. Viral class 1 RNase III involved in suppression of RNA silencing. J. Virol. 2005, 79, 7227–7238. [Google Scholar]
- Cañizares, M.C.; Navas-Castillo, J.; Moriones, E. Multiple suppressors of RNA silencing encoded by both genomic RNAs of the crinivirus Tomato chlorosis virus. Virology 2008, 379, 168–174. [Google Scholar]
- Kataya, A.R.A.; Stavridou, E.; Farhan, K.; Livieratos, I.C. Nucleotide sequence analysis and detection of a Greek isolate of Tomato chlorosis virus. Plant Pathol. 2009, 57, 789–989. [Google Scholar]
- Cuellar, W.J.; Kreuze, J.F.; Rajamaki, M.L.; Cruzado, K.R.; Untiveros, M.; Valkonen, J.P. Elimination of antiviral defense by viral RNase III. Proc. Natl. Acad. Sci. USA 2009, 106, 10354–10358. [Google Scholar]
- Landeo-Ríos, Y.; Navas-Castillo, J.; Moriones, E.; Cañizares, M.C. The p22 RNA silencing suppressor of the crinivirus Tomato chlorosis virus preferentially binds long dsRNAs preventing them from cleavage. Virology 2015, 488, 129–136. [Google Scholar]
- Aguilar, J.M.; Franco, M.; Marco, C.F.; Berdiales, B.; Rodriguez-Cerezo, E.; Truniger, V.; Aranda, M.A. Further variability within the genus Crinivirus, as revealed by determination of the complete RNA genome sequence of Cucurbit yellow stunting disorder virus. J. Gen. Virol. 2003, 84, 2555–2564. [Google Scholar]
- Rubio, L.; Yeh, H.H.; Tian, T.; Falk, B.W. A heterogeneous population of defective RNAs is associated with Lettuce infectious yellows virus. Virology 2000, 271, 205–212. [Google Scholar]
- Rubio, L.; Ayllón, A.; Kong, P.; Fernández, A.; Polek, M.; Guerri, J.; Moreno, P.; Falk, B.W. Genetic variation of Citrus tristeza virus isolates from California and Spain: Evidence for mixed infections and recombination. J. Virol. 2001, 75, 8054–8062. [Google Scholar]
- Salem, N.M.; Chen, A.Y.S.; Tzanetakis, I.E.; Mongkolsiriwattana, C.; Ng, J.C.K. Further complexity of the genus Crinivirus revealed by the complete genome sequence of Lettuce chlorosis virus (LCV) and the similar temporal accumulation of LCV genomic RNAs 1 and 2. Virology 2009, 390, 45–55. [Google Scholar]
- Dreher, T.W. Functions of the 3-untranslated regions of positive strand RNA viral genomes. Annu. Rev. Phytopathol. 1999, 37, 151–174. [Google Scholar]
- Livieratos, I.C.; Eliasco, E.; Müller, G.; Olsthoorn, R.C.; Salazar, L.F.; Pleij, C.W.; Coutts, R.H. Analysis of the RNA of Potato yellow vein virus: Evidence for a tripartite genome and conserved 3′-terminal structures among members of the genus Crinivirus. J. Gen. Virol. 2004, 85, 2065–2075. [Google Scholar]
- Peremyslov, V.V.; Hagiwara, Y.; Dolja, V.V. Genes required for replication of the 15.5-kilobase RNA genome of a plant closterovirus. J. Virol. 1998, 72, 5870–5876. [Google Scholar]
- Satyanarayana, T.; Gowda, S.; Boyko, V.P.; Albiach-Marti, M.R.; Mawassi, M.; Navas-Castillo, J.; Karasev, A.V.; Dolja, V.; Hilf, M.E.; Lewandowski, D.J.; et al. An engineered closterovirus RNA replicon and analysis of heterologous terminal sequences for replication. Proc. Natl. Acad. Sci. USA 1999, 96, 7433–7438. [Google Scholar]
- Ng, J.C. A quantum dot-immunofluorescent labelling method to investigate the interactions between a Crinivirus and its whitefly vector. Front. Microbiol. 2013, 4, 183–192. [Google Scholar]
- Rodríguez-López, M.J.; Garzo, E.; Bonani, J.P.; Fereres, A.; Fernández-Muñoz, R.; Moriones, E. Whitefly resistance traits derived from the wild tomato Solanum pimpinellifolium affect the preference and feeding behavior of Bemisia tabaci and reduce the spread of Tomato yellow leaf curl virus. Phytopathology 2011, 101, 1191–1201. [Google Scholar]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Owen, C.A.; Moukarzel, R.; Huang, X.; Kassem, M.A.; Eliasco, E.; Aranda, M.A.; Coutts, R.H.A.; Livieratos, I.C. In Vitro Synthesized RNA Generated from cDNA Clones of Both Genomic Components of Cucurbit yellow stunting disorder virus Replicates in Cucumber Protoplasts. Viruses 2016, 8, 170. https://doi.org/10.3390/v8060170
Owen CA, Moukarzel R, Huang X, Kassem MA, Eliasco E, Aranda MA, Coutts RHA, Livieratos IC. In Vitro Synthesized RNA Generated from cDNA Clones of Both Genomic Components of Cucurbit yellow stunting disorder virus Replicates in Cucumber Protoplasts. Viruses. 2016; 8(6):170. https://doi.org/10.3390/v8060170
Chicago/Turabian StyleOwen, Carolyn A., Romy Moukarzel, Xiao Huang, Mona A. Kassem, Eleonora Eliasco, Miguel A. Aranda, Robert H. A. Coutts, and Ioannis C. Livieratos. 2016. "In Vitro Synthesized RNA Generated from cDNA Clones of Both Genomic Components of Cucurbit yellow stunting disorder virus Replicates in Cucumber Protoplasts" Viruses 8, no. 6: 170. https://doi.org/10.3390/v8060170
APA StyleOwen, C. A., Moukarzel, R., Huang, X., Kassem, M. A., Eliasco, E., Aranda, M. A., Coutts, R. H. A., & Livieratos, I. C. (2016). In Vitro Synthesized RNA Generated from cDNA Clones of Both Genomic Components of Cucurbit yellow stunting disorder virus Replicates in Cucumber Protoplasts. Viruses, 8(6), 170. https://doi.org/10.3390/v8060170