Regulated Transport into the Nucleus of Herpesviridae DNA Replication Core Proteins

 ,
, 
Abstract
:1. Introduction
2. Nuclear Import
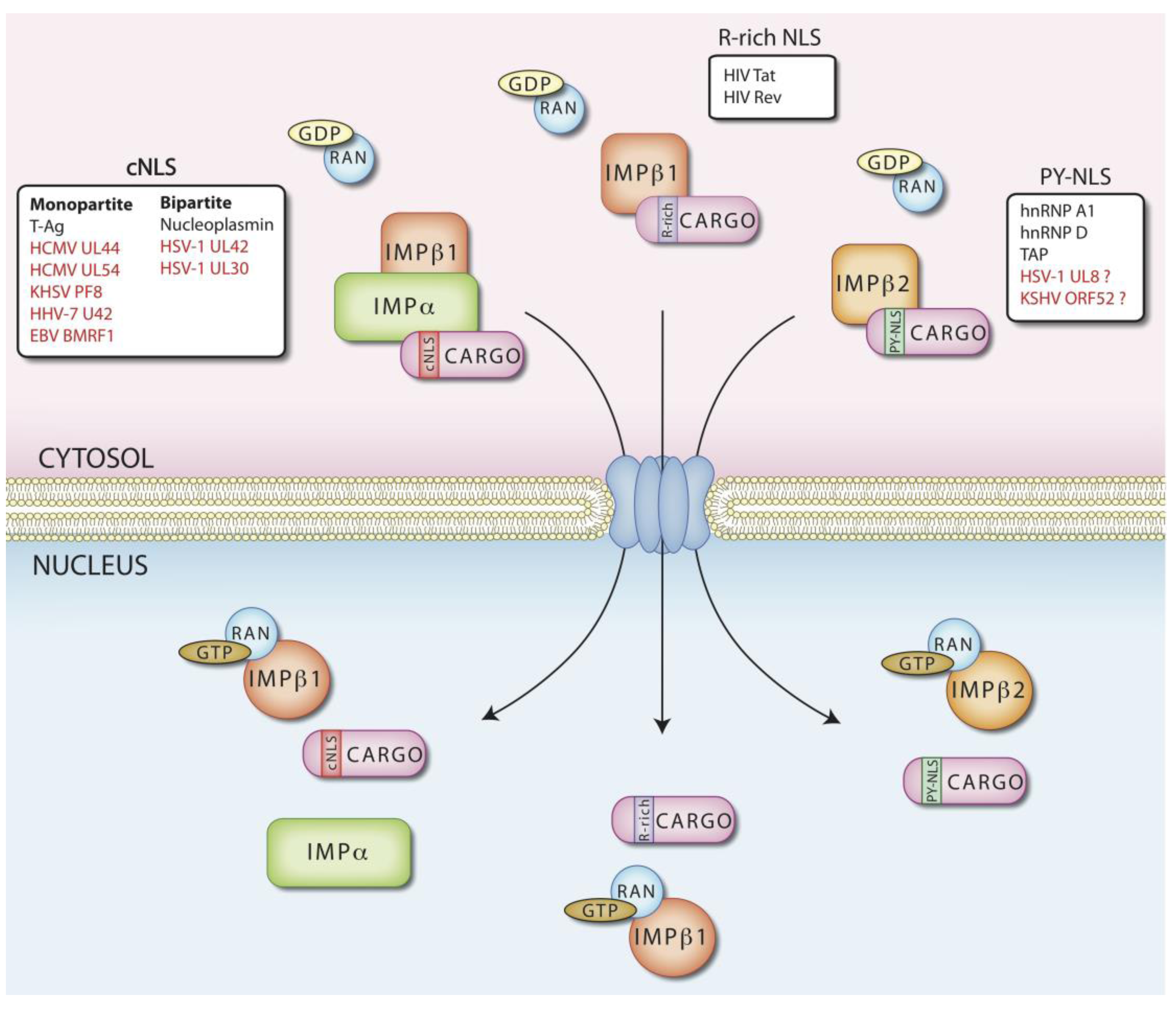
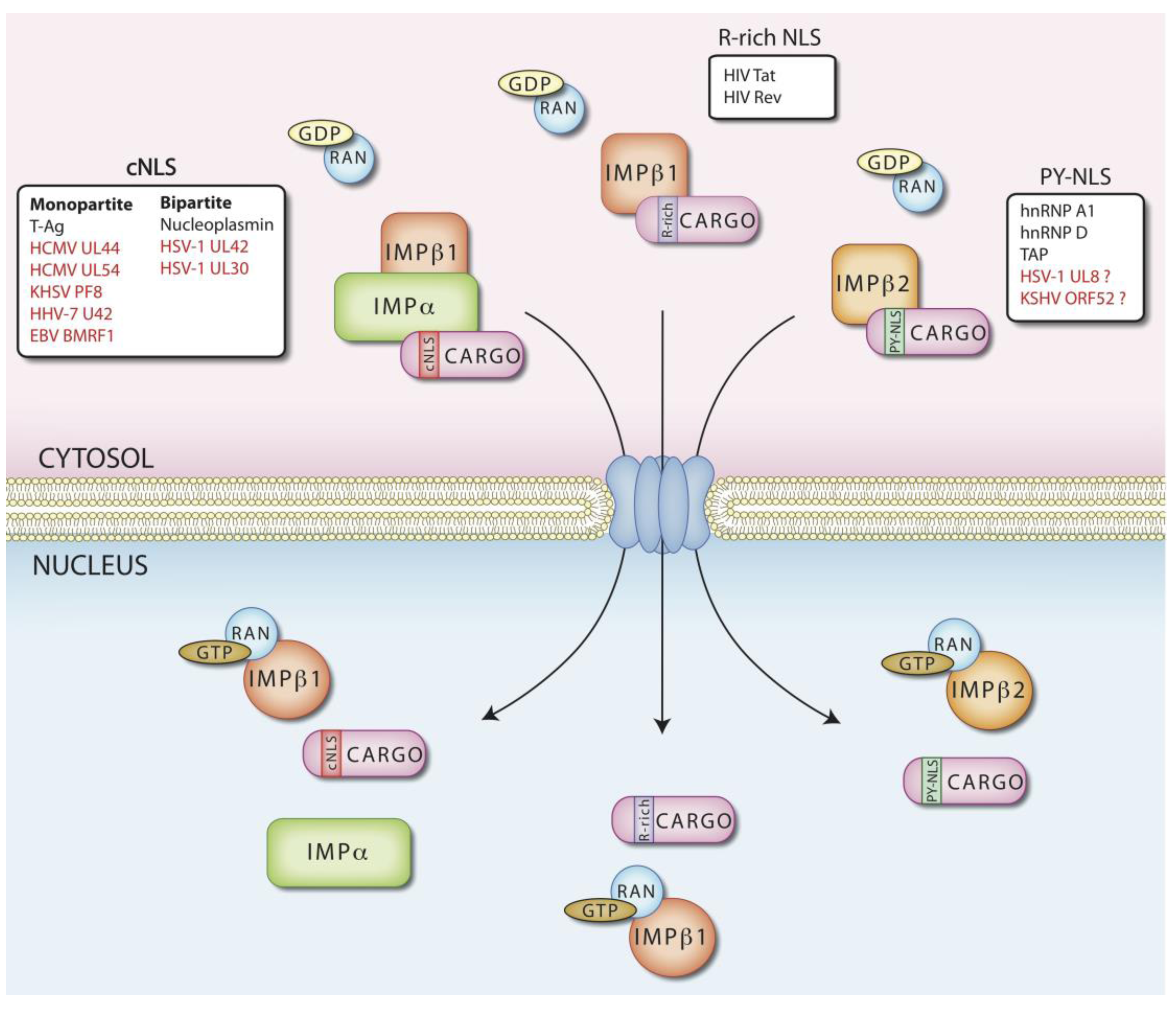
3. Regulation of Nuclear Transport
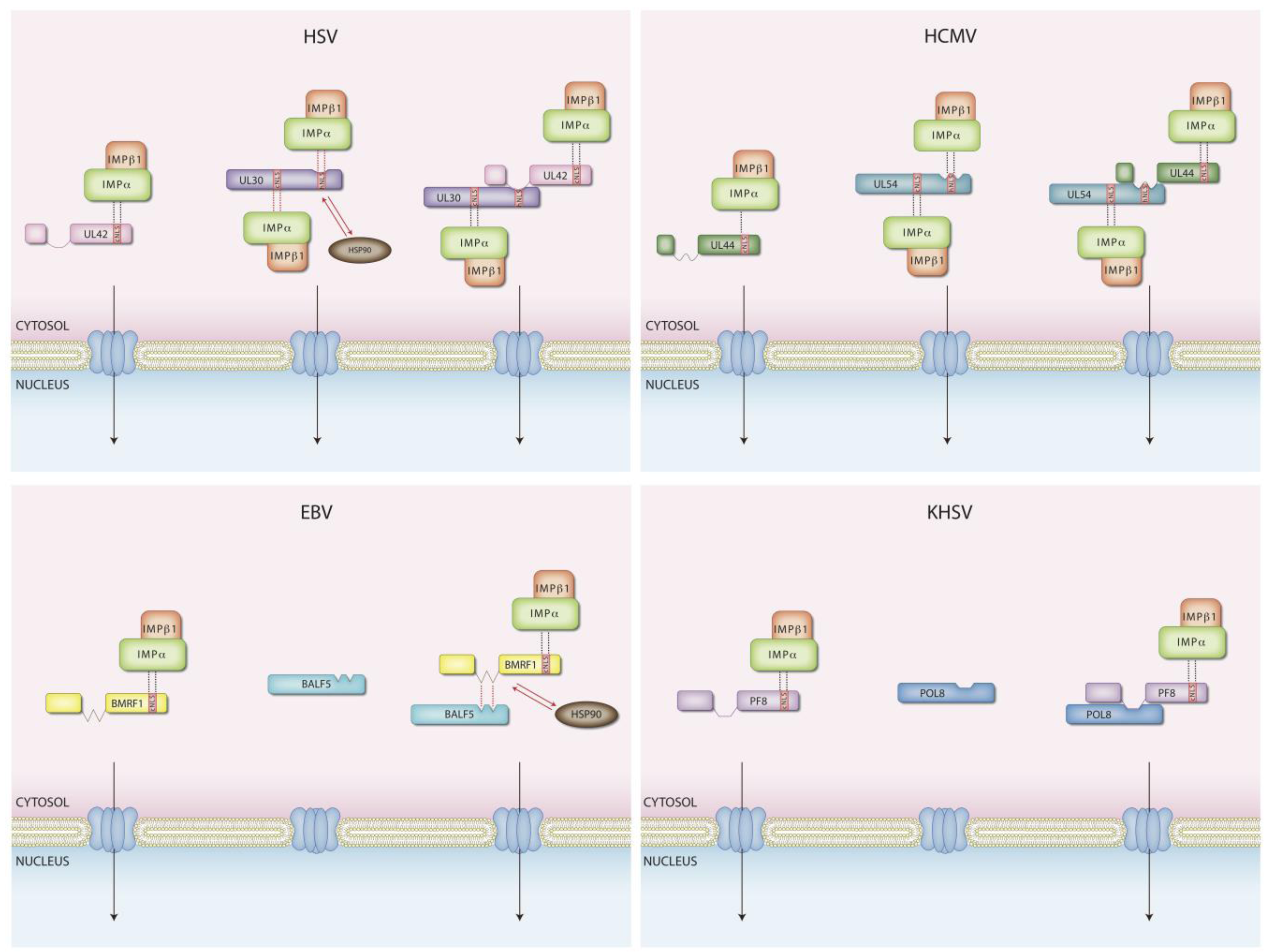
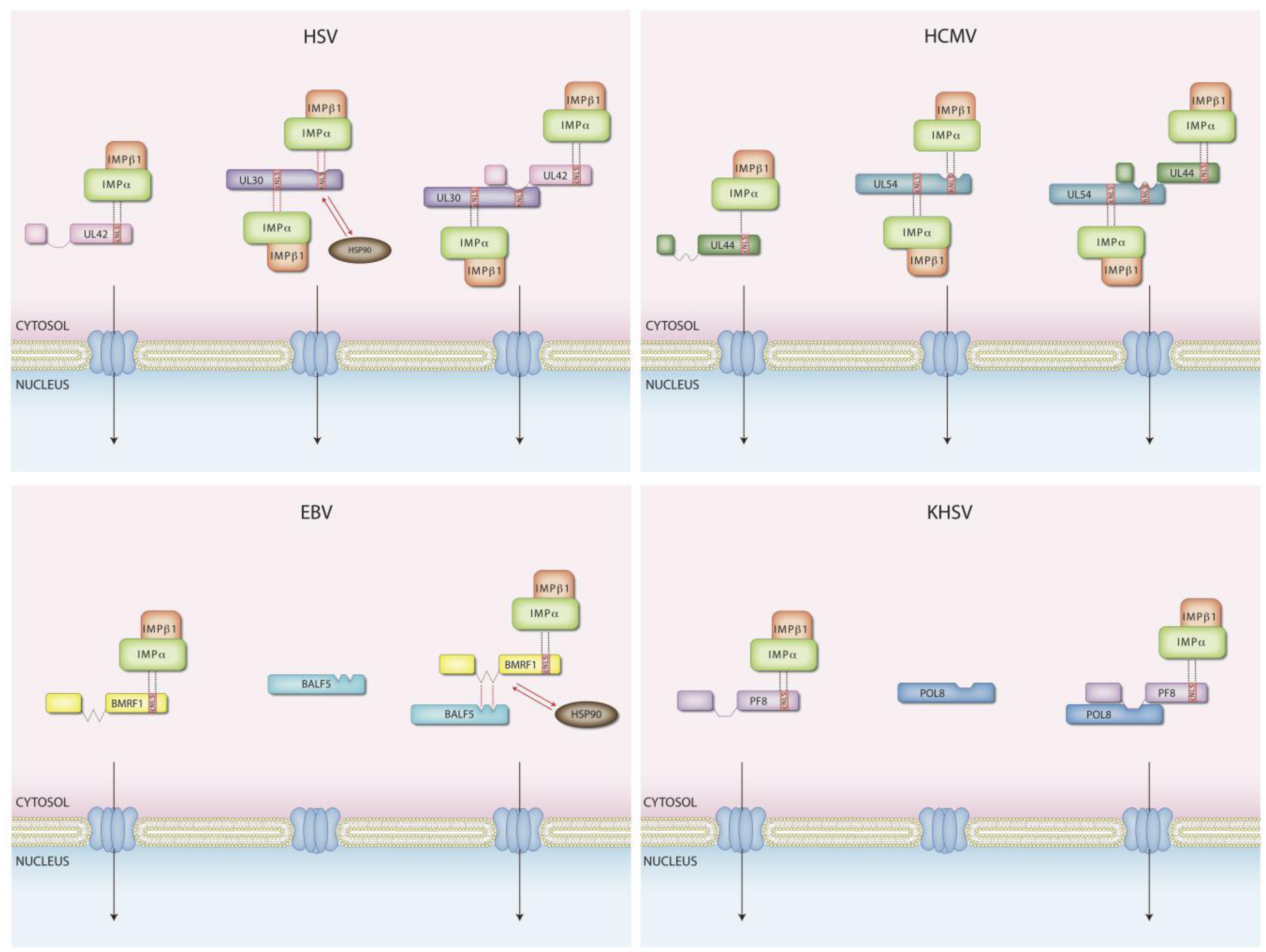
4. Nuclear Transport of Herpesviral DNA Replicating Enzymes

{kind=link}
{kind=link}
{kind=link}
| HSV-1 | HCMV | |||||||
|---|---|---|---|---|---|---|---|---|
| protein | name | NLS | IMPs | loc. alone | name | NLS | IMPs | loc. alone |
| processivity factor | UL42 | 1 cNLS bip [55] | a/β | N [55] | UL44 | 1 cNLS [52] | a/β | N [52] |
| DNA polymerase | UL30 | 1 cNLS bip [53] 1 NLS hyd [53,56] | a/β | N [53,56,87] | UL54 | 1 cNLS [54] 1 NLS hyd [56] | a/β | N [56] |
| primase | UL5 | C [63,64] | UL70 | C [52] | ||||
| helicase | UL52 | C [63,64] | UL105 | N in infected cells [110] | ||||
| primase-associated factor | UL8 | putative PY-NLS (see text for details) | C [63,64] | UL102 | ||||
| ssDNA binding protein | UL29 | ? | UL57 | |||||
| DNA polymerase holoenzyme nuclear import | Individual subunits and holoenzyme [55] | Individual subunits and holoenzyme [54] | ||||||
| PAP status | Monomer [76] | Dimer [77,106]; transported to the nucleus as dimer [105] | ||||||
| Helicase primase complex nuclear import | Transported as a trimeric complex C [63,64] | Not known, but UL70 can localize to the nucleus independently of the other subunits if the cellular chaperone DNAJB6-a is overexpressed [109] | ||||||
| EBV | KHSV | |||||||
| protein | name | NLS | IMPs | loc. alone | name | NLS | IMPs | loc. alone |
| processivity factor | BMRF1 | 1 cNLS [84] | N [84] | ORF59 | 1 cNLS [59] | N [59] | ||
| DNA polymerase | BALF5 | C [60] | ORF9 | C [59] | ||||
| primase | BSLF1 | C [61] | ORF44 | putative PY-NLS (see text for details) | C [59] | |||
| helicase | BBLF4 | C [61] | ORF56 | C [59] | ||||
| primase-associated factor | BBLF2/3 | C [61] | ORF40/41 | C [59] | ||||
| ssDNA binding protein | BALF2 | ORF6 | ||||||
| DNA polymerase holoenzyme nuclear import | Piggy back as mediated by PAP [60] | Piggy back as mediated by PAP [59] | ||||||
| PAP status | Multimer [78,114] | Dimer [79,115 ]; transported to the nucleus as dimer [116] | ||||||
| Helicase primase complex nuclear import | Imported as a complex; BBLF4 can be imported if expressed with the ZTA transactivator [ 61] | pol/PAP and all the helicase primase subunits need to be simultaneously expressed to achieve nuclear targeting [62] | ||||||
5. DNA Polymerase Holoenzyme Nuclear Transport
| Virus | Protein | Name | NLS type | Sequence | IMPs | Ran | ATP | Nuclear import | Notes | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| HSV-1 | pol | UL30 | hNLS | RRMLHR-1129 | ? | ? | ? | s | overlapping PAP binding domain | [56] |
| cNLS bip | PAKRPRETPSPADPPGGASKPRK-1136 | a/β | X | X | s + h | [53] | ||||
| PAP | UL42 | cNLS bip | PTTKRGRSGGEDARADALKKPK-413 | a/β | X | X | s + h | [55] | ||
| HCMV | pol | UL54 | hNLS | PRRLHL | ? | ? | ? | s | overlapping PAP binding domain | [56] |
| cNLS | PAKKRAR-1159 | a/β | X | X | s + h | [54] | ||||
| PAP | UL44 | cNLS | PNTKKQK-431 | a/β | X | X | s + h | regulated by phosphorylation | [52] | |
| HHV-7 | pol | U38 | / | / | / | / | / | / | ||
| PAP | U27 | cNLS | PNSKRQR-361 | ? | X | X | s | [57] | ||
| KHSV-1 | pol | pol-8 | no NLS | h | requires PF-8 for import | [59] | ||||
| PAP | PF-8 | cNLS | KRPHKRRSD-377 | ? | ? | ? | s + h | mediates translocation of holoenzyme | [59] | |
| EBV | pol | BALF1 | no NLS | h | requires BMRF1 for import | [60] | ||||
| PAP | BMRF1 | cNLS | KHPKK-396 | ? | ? | ? | s + h | mediates translocation of holoenzyme | [58] |
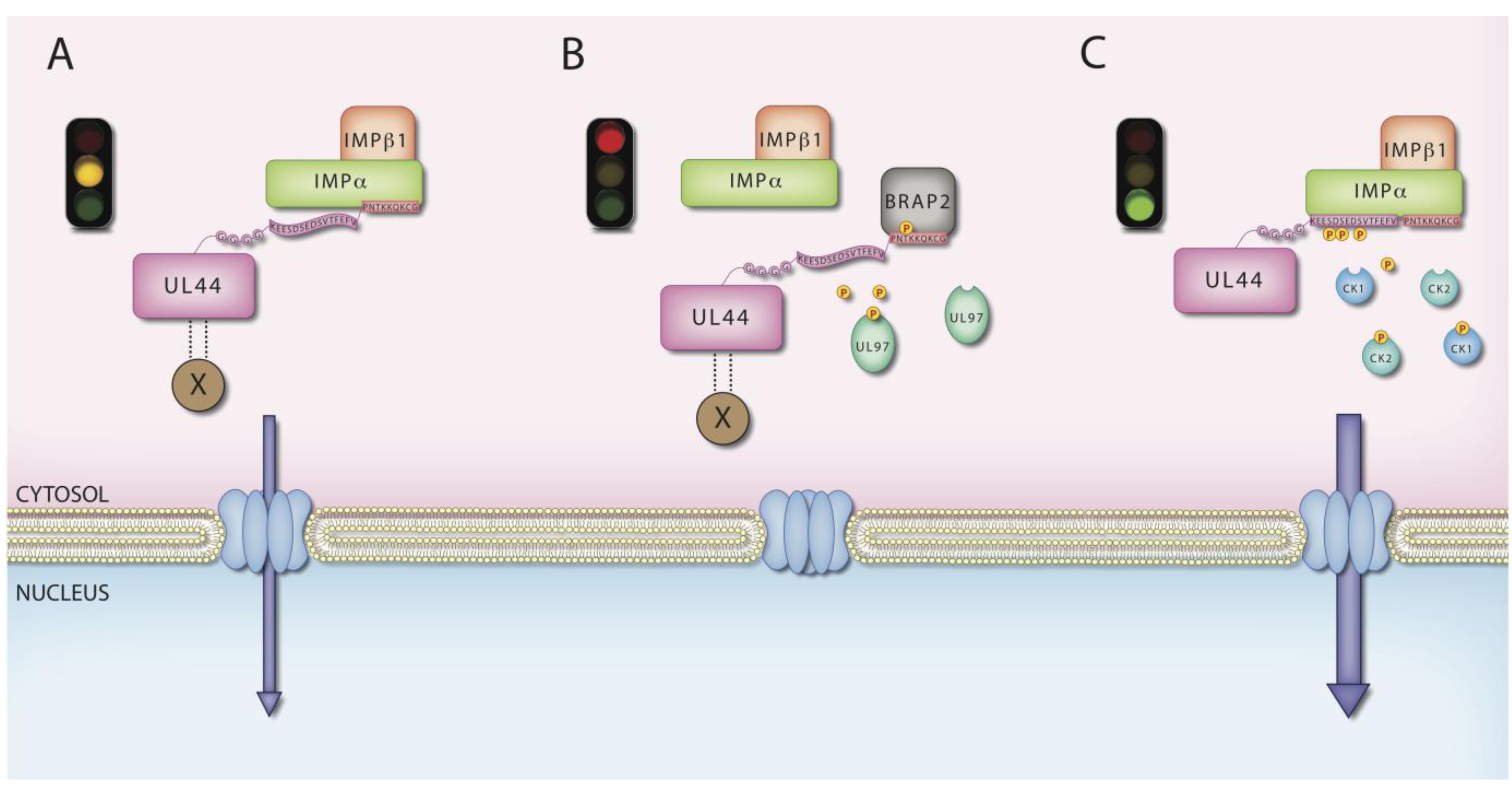
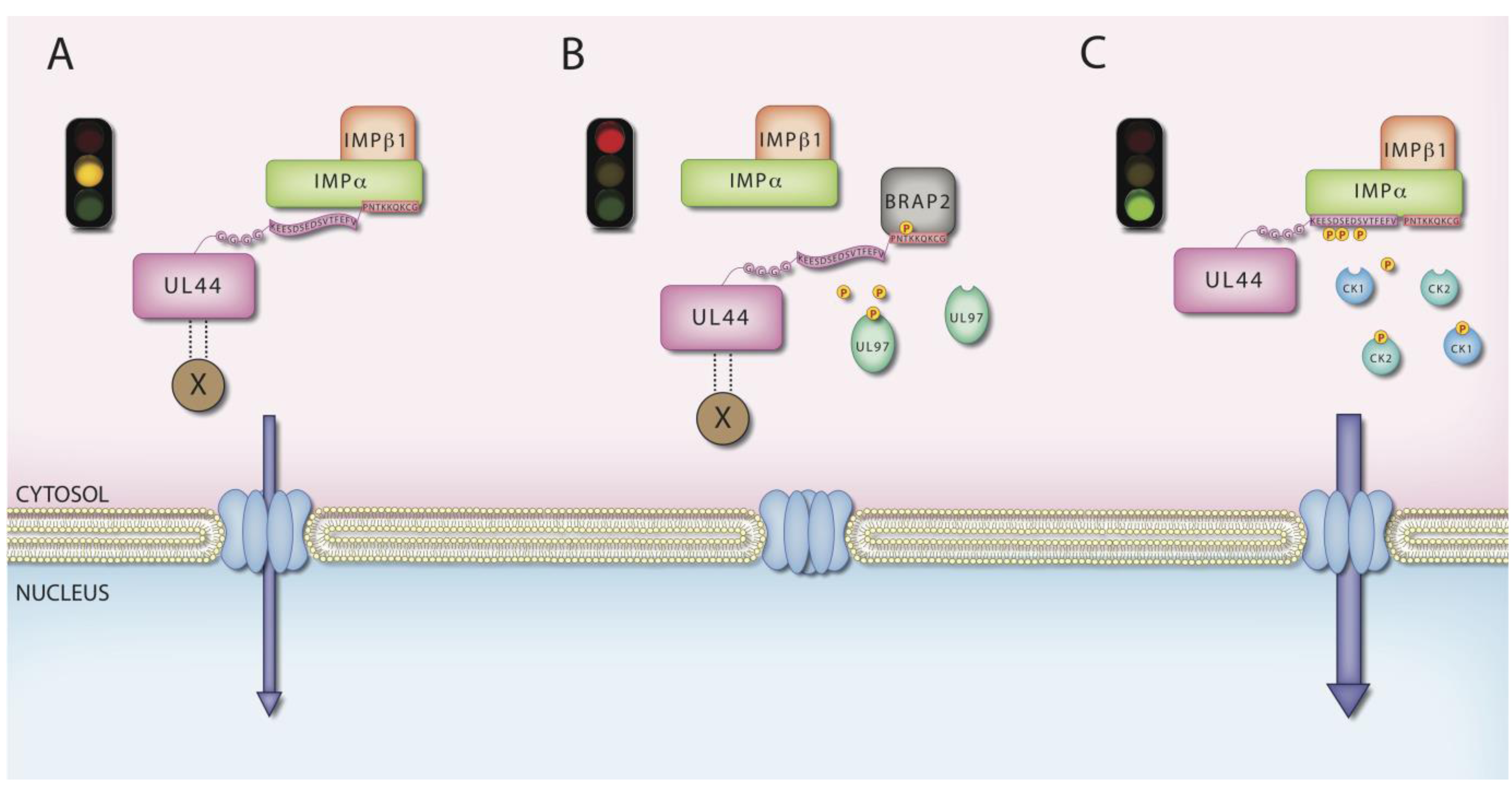
6. MCV UL44 by IMPs
7. Nuclear Import of DNA Primase Helicase Complexes and the Formation of Composite NLSs
8. An Emerging Role for Cellular Chaperones in Nuclear Import
9. Conclusions
Acknowledgments
Conflicts of Interest
References and Notes
- Roizman, B.; Pellett, P.E. The family Herpesviridae: A brief introduction. In Fields Virology, 4th ed.; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2001; Volume 2. [Google Scholar]
- Wilkinson, D.E.; Weller, S.K. The role of DNA recombination in herpes simplex virus DNA replication. IUBMB Life 2003, 55, 451–458. [Google Scholar] [CrossRef]
- Strang, B.L.; Boulant, S.; Coen, D.M. Nucleolin associates with the human cytomegalovirus DNA polymerase accessory subunit UL44 and is necessary for efficient viral replication. J. Virol. 2010, 84, 1771–1784. [Google Scholar] [CrossRef]
- Anders, D.G.; Kacica, M.A.; Pari, G.; Punturieri, S.M. Boundaries and structure of human cytomegalovirus oriLyt, a complex origin for lytic-phase DNA replication. J. Virol. 1992, 66, 3373–3384. [Google Scholar]
- Pari, G.S.; Anders, D.G. Eleven loci encoding trans-acting factors are required for transient complementation of human cytomegalovirus oriLyt-dependent DNA replication. J. Virol. 1993, 67, 6979–6988. [Google Scholar]
- Pari, G.S.; Kacica, M.A.; Anders, D.G. Open reading frames UL44, IRS1/TRS1, and UL36-38 are required for transient complementation of human cytomegalovirus oriLyt-dependent DNA synthesis. J. Virol. 1993, 67, 2575–2582. [Google Scholar]
- Anders, D.G.; McCue, L.A. The human cytomegalovirus genes and proteins required for DNA synthesis. Intervirology 1996, 39, 378–388. [Google Scholar]
- Tran, E.J.; Wente, S.R. Dynamic nuclear pore complexes: Life on the edge. Cell 2006, 125, 1041–1053. [Google Scholar] [CrossRef]
- Cardarelli, F.; Serresi, M.; Bizzarri, R.; Giacca, M.; Beltram, F. In vivo study of HIV-1 Tat arginine-rich motif unveils its transport properties. Mol. Ther. 2007, 15, 1313–1322. [Google Scholar] [CrossRef]
- Fahrenkrog, B.; Aebi, U. The nuclear pore complex: Nucleocytoplasmic transport and beyond. Nat. Rev. Mol. Cell Biol. 2003, 4, 757–766. [Google Scholar] [CrossRef]
- Katahira, J.; Yoneda, Y. Nucleocytoplasmic transport of microRNAs and related small RNAs. Traffic 2011, 12, 1468–1474. [Google Scholar] [CrossRef]
- Weis, K. Importins and exportins: How to get in and out of the nucleus. Trends Biochem. Sci. 1998, 23, 185–189. [Google Scholar] [CrossRef]
- Czernilofsky, A.P.; Levinson, A.D.; Varmus, H.E.; Bishop, J.M.; Tischer, E.; Goodman, H.M. Nucleotide sequence of an avian sarcoma virus oncogene (src) and proposed amino acid sequence for gene product. Nature 1980, 287, 198–203. [Google Scholar] [CrossRef]
- Balch, W.E.; McCaffery, J.M.; Plutner, H.; Farquhar, M.G. Vesicular stomatitis virus glycoprotein is sorted and concentrated during export from the endoplasmic reticulum. Cell 1994, 76, 841–852. [Google Scholar] [CrossRef]
- Kalderon, D.; Richardson, W.D.; Markham, A.F.; Smith, A.E. Sequence requirements for nuclear location of simian virus 40 large-T antigen. Nature 1984, 311, 33–38. [Google Scholar] [CrossRef]
- Kalderon, D.; Roberts, B.L.; Richardson, W.D.; Smith, A.E. A short amino acid sequence able to specify nuclear location. Cell 1984, 39, 499–509. [Google Scholar] [CrossRef]
- Lange, A.; Mills, R.E.; Lange, C.J.; Stewart, M.; Devine, S.E.; Corbett, A.H. Classical nuclear localization signals: Definition, function, and interaction with importin alpha. J. Biol. Chem. 2007, 282, 5101–5105. [Google Scholar]
- Conti, E.; Uy, M.; Leighton, L.; Blobel, G.; Kuriyan, J. Crystallographic analysis of the recognition of a nuclear localization signal by the nuclear import factor karyopherin alpha. Cell 1998, 94, 193–204. [Google Scholar] [CrossRef]
- Kobe, B. Autoinhibition by an internal nuclear localization signal revealed by the crystal structure of mammalian importin alpha. Nat. Struct. Biol. 1999, 6, 388–397. [Google Scholar] [CrossRef]
- Lee, S.J.; Matsuura, Y.; Liu, S.M.; Stewart, M. Structural basis for nuclear import complex dissociation by RanGTP. Nature 2005, 435, 693–696. [Google Scholar] [CrossRef]
- Scheffzek, K.; Klebe, C.; Fritz-Wolf, K.; Kabsch, W.; Wittinghofer, A. Crystal structure of the nuclear Ras-related protein Ran in its GDP-bound form. Nature 1995, 374, 378–381. [Google Scholar] [CrossRef]
- Vetter, I.R.; Nowak, C.; Nishimoto, T.; Kuhlmann, J.; Wittinghofer, A. Structure of a Ran-binding domain complexed with Ran bound to a GTP analogue: Implications for nuclear transport. Nature 1999, 398, 39–46. [Google Scholar] [CrossRef]
- Izaurralde, E.; Kutay, U.; von Kobbe, C.; Mattaj, I.W.; Gorlich, D. The asymmetric distribution of the constituents of the Ran system is essential for transport into and out of the nucleus. EMBO J. 1997, 16, 6535–6547. [Google Scholar] [CrossRef]
- Bischoff, F.R.; Klebe, C.; Kretschmer, J.; Wittinghofer, A.; Ponstingl, H. RanGAP1 induces GTPase activity of nuclear Ras-related Ran. Proc. Natl. Acad. Sci. USA 1994, 91, 2587–2591. [Google Scholar]
- Bischoff, F.R.; Ponstingl, H. Catalysis of guanine nucleotide exchange on Ran by the mitotic regulator RCC1. Nature 1991, 354, 80–82. [Google Scholar] [CrossRef]
- Ribbeck, K.; Lipowsky, G.; Kent, H.M.; Stewart, M.; Gorlich, D. NTF2 mediates nuclear import of Ran. EMBO J. 1998, 17, 6587–6598. [Google Scholar] [CrossRef]
- Kutay, U.; Bischoff, F.R.; Kostka, S.; Kraft, R.; Gorlich, D. Export of importin alpha from the nucleus is mediated by a specific nuclear transport factor. Cell 1997, 90, 1061–1071. [Google Scholar] [CrossRef]
- Hogarth, C.A.; Calanni, S.; Jans, D.A.; Loveland, K.L. Importin alpha mRNAs have distinct expression profiles during spermatogenesis. Dev. Dyn. 2006, 235, 253–262. [Google Scholar] [CrossRef]
- Huber, J.; Cronshagen, U.; Kadokura, M.; Marshallsay, C.; Wada, T.; Sekine, M.; Luhrmann, R. Snurportin1, an m3G-cap-specific nuclear import receptor with a novel domain structure. EMBO J. 1998, 17, 4114–4126. [Google Scholar] [CrossRef]
- Alvisi, G.; Poon, I.K.; Jans, D.A. Tumor-specific nuclear targeting: Promises for anti-cancer therapy? Drug Resist. Updates 2006, 9, 40–50. [Google Scholar] [CrossRef]
- Kuusisto, H.V.; Wagstaff, K.M.; Alvisi, G.; Jans, D.A. The C-terminus of apoptin represents a unique tumor cell-enhanced nuclear targeting module. Int. J. Cancer 2008, 123, 2965–2969. [Google Scholar] [CrossRef]
- Kuusisto, H.V.; Wagstaff, K.M.; Alvisi, G.; Roth, D.M.; Jans, D.A. Global enhancement of nuclear localization-dependent nuclear transport in transformed cells. FASEB J. 2012, 26, 1181–1193. [Google Scholar] [CrossRef]
- Truant, R.; Cullen, B.R. The arginine-rich domains present in human immunodeficiency virus type 1 Tat and Rev function as direct importin beta-dependent nuclear localization signals. Mol. Cell. Biol. 1999, 19, 1210–1217. [Google Scholar]
- Cherezova, L.; Burnside, K.L.; Rose, T.M. Conservation of complex nuclear localization signals utilizing classical and non-classical nuclear import pathways in LANA homologs of KSHV and RFHV. PLoS One 2011, 6, e18920. [Google Scholar] [CrossRef]
- Fontes, M.R.; Teh, T.; Kobe, B. Structural basis of recognition of monopartite and bipartite nuclear localization sequences by mammalian importin-alpha. J. Mol. Biol. 2000, 297, 1183–1194. [Google Scholar] [CrossRef]
- Chook, Y.M.; Suel, K.E. Nuclear import by karyopherin-betas: Recognition and inhibition. Biochim. Biophys. Acta 2011, 1813, 1593–1606. [Google Scholar]
- Pollard, V.W.; Michael, W.M.; Nakielny, S.; Siomi, M.C.; Wang, F.; Dreyfuss, G. A novel receptor-mediated nuclear protein import pathway. Cell 1996, 86, 985–994. [Google Scholar] [CrossRef]
- Kawamura, H.; Tomozoe, Y.; Akagi, T.; Kamei, D.; Ochiai, M.; Yamada, M. Identification of the nucleocytoplasmic shuttling sequence of heterogeneous nuclear ribonucleoprotein D-like protein JKTBP and its interaction with mRNA. J. Biol. Chem. 2002, 277, 2732–2739. [Google Scholar]
- Rebane, A.; Aab, A.; Steitz, J.A. Transportins 1 and 2 are redundant nuclear import factors for hnRNP A1 and HuR. RNA 2004, 10, 590–599. [Google Scholar] [CrossRef]
- Siomi, M.C.; Fromont, M.; Rain, J.C.; Wan, L.; Wang, F.; Legrain, P.; Dreyfuss, G. Functional conservation of the transportin nuclear import pathway in divergent organisms. Mol. Cell. Biol. 1998, 18, 4141–4148. [Google Scholar]
- Truant, R.; Kang, Y.; Cullen, B.R. The human tap nuclear RNA export factor contains a novel transportin-dependent nuclear localization signal that lacks nuclear export signal function. J. Biol. Chem. 1999, 274, 32167–32171. [Google Scholar] [CrossRef]
- Lee, B.J.; Cansizoglu, A.E.; Suel, K.E.; Louis, T.H.; Zhang, Z.; Chook, Y.M. Rules for nuclear localization sequence recognition by karyopherin beta 2. Cell 2006, 126, 543–558. [Google Scholar] [CrossRef]
- Imasaki, T.; Shimizu, T.; Hashimoto, H.; Hidaka, Y.; Kose, S.; Imamoto, N.; Yamada, M.; Sato, M. Structural basis for substrate recognition and dissociation by human transportin 1. Mol. Cell 2007, 28, 57–67. [Google Scholar] [CrossRef]
- Cansizoglu, A.E.; Lee, B.J.; Zhang, Z.C.; Fontoura, B.M.; Chook, Y.M. Structure-based design of a pathway-specific nuclear import inhibitor. Nat. Struct. Mol. Biol. 2007, 14, 452–454. [Google Scholar] [CrossRef]
- Okada, N.; Ishigami, Y.; Suzuki, T.; Kaneko, A.; Yasui, K.; Fukutomi, R.; Isemura, M. Importins and exportins in cellular differentiation. J. Cell. Mol. Med. 2008, 12, 1863–1871. [Google Scholar] [CrossRef]
- Jans, D.A.; Xiao, C.Y.; Lam, M.H. Nuclear targeting signal recognition: A key control point in nuclear transport? Bioessays 2000, 22, 532–544. [Google Scholar] [CrossRef]
- Terry, L.J.; Shows, E.B.; Wente, S.R. Crossing the nuclear envelope: Hierarchical regulation of nucleocytoplasmic transport. Science 2007, 318, 1412–1416. [Google Scholar] [CrossRef]
- Poon, I.K.; Jans, D.A. Regulation of nuclear transport: Central role in development and transformation? Traffic 2005, 6, 173–186. [Google Scholar] [CrossRef]
- Briggs, L.J.; Stein, D.; Goltz, J.; Corrigan, V.C.; Efthymiadis, A.; Hubner, S.; Jans, D.A. The cAMP-dependent protein kinase site (Ser312) enhances dorsal nuclear import through facilitating nuclear localization sequence/importin interaction. J. Biol. Chem. 1998, 273, 22745–22752. [Google Scholar]
- Hubner, S.; Xiao, C.Y.; Jans, D.A. The protein kinase CK2 site (Ser111/112) enhances recognition of the simian virus 40 large T-antigen nuclear localization sequence by importin. J. Biol. Chem. 1997, 272, 17191–17195. [Google Scholar] [CrossRef]
- Fulcher, A.J.; Roth, D.M.; Fatima, S.; Alvisi, G.; Jans, D.A. The BRCA-1 binding protein BRAP2 is a novel, negative regulator of nuclear import of viral proteins, dependent on phosphorylation flanking the nuclear localization signal. FASEB J. 2010, 24, 1454–1466. [Google Scholar] [CrossRef]
- Alvisi, G.; Jans, D.A.; Guo, J.; Pinna, L.A.; Ripalti, A. A protein kinase CK2 site flanking the nuclear targeting signal enhances nuclear transport of human cytomegalovirus ppUL44. Traffic 2005, 6, 1002–1013. [Google Scholar] [CrossRef]
- Alvisi, G.; Musiani, D.; Jans, D.A.; Ripalti, A. An importin alpha/beta-recognized bipartite nuclear localization signal mediates targeting of the human herpes simplex virus type 1 DNA polymerase catalytic subunit pUL30 to the nucleus. Biochemistry 2007, 46, 9155–9163. [Google Scholar] [CrossRef]
- Alvisi, G.; Ripalti, A.; Ngankeu, A.; Giannandrea, M.; Caraffi, S.G.; Dias, M.M.; Jans, D.A. Human cytomegalovirus DNA polymerase catalytic subunit pUL54 possesses independently acting nuclear localization and ppUL44 binding motifs. Traffic 2006, 7, 1322–1332. [Google Scholar] [CrossRef]
- Alvisi, G.; Avanzi, S.; Musiani, D.; Camozzi, D.; Leoni, V.; Ly-Huynh, J.D.; Ripalti, A. Nuclear import of HSV-1 DNA polymerase processivity factor UL42 is mediated by a C-terminally located bipartite nuclear localization signal. Biochemistry 2008, 47, 13764–13777. [Google Scholar] [CrossRef]
- Loregian, A.; Piaia, E.; Cancellotti, E.; Papini, E.; Marsden, H.S.; Palu, G. The catalytic subunit of herpes simplex virus type 1 DNA polymerase contains a nuclear localization signal in the UL42-binding region. Virology 2000, 273, 139–148. [Google Scholar] [CrossRef]
- Takeda, K.; Haque, M.; Nagoshi, E.; Takemoto, M.; Shimamoto, T.; Yoneda, Y.; Yamanishi, K. Characterization of human herpesvirus 7 U27 gene product and identification of its nuclear localization signal. Virology 2000, 272, 394–401. [Google Scholar] [CrossRef]
- Zhang, Q.; Holley-Guthrie, E.; Dorsky, D.; Kenney, S. Identification of transactivator and nuclear localization domains in the Epstein-Barr virus DNA polymerase accessory protein, BMRF1. J. Gen. Virol. 1999, 80, 69–74. [Google Scholar]
- Chen, Y.; Ciustea, M.; Ricciardi, R.P. Processivity factor of KSHV contains a nuclear localization signal and binding domains for transporting viral DNA polymerase into the nucleus. Virology 2005, 340, 183–191. [Google Scholar] [CrossRef]
- Kawashima, D.; Kanda, T.; Murata, T.; Saito, S.; Sugimoto, A.; Narita, Y.; Tsurumi, T. Nuclear transport of epstein-barr virus DNA polymerase is dependent on the BMRF1 polymerase processivity factor and molecular chaperone Hsp90. J. Virol. 2013, 87, 6482–6491. [Google Scholar] [CrossRef]
- Gao, Z.; Krithivas, A.; Finan, J.E.; Semmes, O.J.; Zhou, S.; Wang, Y.; Hayward, S.D. The Epstein-Barr virus lytic transactivator Zta interacts with the helicase-primase replication proteins. J. Virol. 1998, 72, 8559–8567. [Google Scholar]
- Wu, F.Y.; Ahn, J.H.; Alcendor, D.J.; Jang, W.J.; Xiao, J.; Hayward, S.D.; Hayward, G.S. Origin-independent assembly of Kaposi’s sarcoma-associated herpesvirus DNA replication compartments in transient cotransfection assays and association with the ORF-K8 protein and cellular PML. J. Virol. 2001, 75, 1487–1506. [Google Scholar] [CrossRef]
- Calder, J.M.; Stow, E.C.; Stow, N.D. On the cellular localization of the components of the herpes simplex virus type 1 helicase-primase complex and the viral origin-binding protein. J. Gen. Virol. 1992, 73, 531–538. [Google Scholar] [CrossRef]
- Barnard, E.C.; Brown, G.; Stow, N.D. Deletion mutants of the herpes simplex virus type 1 UL8 protein: Effect on DNA synthesis and ability to interact with and influence the intracellular localization of the UL5 and UL52 proteins. Virology 1997, 237, 97–106. [Google Scholar] [CrossRef]
- Ertl, P.F.; Powell, K.L. Physical and functional interaction of human cytomegalovirus DNA polymerase and its accessory protein (ICP36) expressed in insect cells. J. Virol. 1992, 66, 4126–4133. [Google Scholar]
- Gottlieb, J.; Marcy, A.I.; Coen, D.M.; Challberg, M.D. The herpes simplex virus type 1 UL42 gene product: A subunit of DNA polymerase that functions to increase processivity. J. Virol. 1990, 64, 5976–5987. [Google Scholar]
- Loregian, A.; Case, A.; Cancellotti, E.; Valente, C.; Marsden, H.S.; Palu, G. Cloning, expression, and functional characterization of the equine herpesvirus 1 DNA polymerase and its accessory subunit. J. Virol. 2006, 80, 6247–6258. [Google Scholar] [CrossRef]
- Stinski, M.F. Herpesviruses. In Herpesviruses; Roizman, B., Ed.; Plenum Press: New York, NY, USA, 1983; Volume 2, pp. 67–113. [Google Scholar]
- Alvisi, G.; Marin, O.; Pari, G.; Mancini, M.; Avanzi, S.; Loregian, A.; Jans, D.A.; Ripalti, A. Multiple phosphorylation sites at the C-terminus regulate nuclear import of HCMV DNA polymerase processivity factor ppUL44. Virology 2011, 417, 259–267. [Google Scholar] [CrossRef]
- Alvisi, G.; Rawlinson, S.M.; Ghildyal, R.; Ripalti, A.; Jans, D.A. Regulated nucleocytoplasmic trafficking of viral gene products: A therapeutic target? Biochim. Biophys. Acta 2008, 1784, 213–227. [Google Scholar]
- Alvisi, G.; Roth, D.M.; Camozzi, D.; Pari, G.S.; Loregian, A.; Ripalti, A.; Jans, D.A. The flexible loop of the human cytomegalovirus DNA polymerase processivity factor ppUL44 is required for efficient DNA binding and replication in cells. J. Virol. 2009, 83, 9567–9576. [Google Scholar] [CrossRef]
- Weiland, K.L.; Oien, N.L.; Homa, F.; Wathen, M.W. Functional analysis of human cytomegalovirus polymerase accessory protein. Virus Res. 1994, 34, 191–206. [Google Scholar] [CrossRef]
- Digard, P.; Chow, C.S.; Pirrit, L.; Coen, D.M. Functional analysis of the herpes simplex virus UL42 protein. J. Virol. 1993, 67, 1159–1168. [Google Scholar]
- Gallo, M.L.; Dorsky, D.I.; Crumpacker, C.S.; Parris, D.S. The essential 65-kilodalton DNA-binding protein of herpes simplex virus stimulates the virus-encoded DNA polymerase. J. Virol. 1989, 63, 5023–5029. [Google Scholar]
- Gallo, M.L.; Jackwood, D.H.; Murphy, M.; Marsden, H.S.; Parris, D.S. Purification of the herpes simplex virus type 1 65-kilodalton DNA-binding protein: Properties of the protein and evidence of its association with the virus-encoded DNA polymerase. J. Virol. 1988, 62, 2874–2883. [Google Scholar]
- Zuccola, H.J.; Filman, D.J.; Coen, D.M.; Hogle, J.M. The crystal structure of an unusual processivity factor, herpes simplex virus UL42, bound to the C terminus of its cognate polymerase. Mol. Cell 2000, 5, 267–278. [Google Scholar] [CrossRef]
- Appleton, B.A.; Loregian, A.; Filman, D.J.; Coen, D.M.; Hogle, J.M. The cytomegalovirus DNA polymerase subunit UL44 forms a C clamp-shaped dimer. Mol. Cell 2004, 15, 233–244. [Google Scholar] [CrossRef]
- Murayama, K.; Nakayama, S.; Kato-Murayama, M.; Akasaka, R.; Ohbayashi, N.; Kamewari-Hayami, Y.; Terada, T.; Shirouzu, M.; Tsurumi, T.; Yokoyama, S. Crystal structure of epstein-barr virus DNA polymerase processivity factor BMRF1. J. Biol. Chem. 2009, 284, 35896–35905. [Google Scholar] [CrossRef]
- Baltz, J.L.; Filman, D.J.; Ciustea, M.; Silverman, J.E.; Lautenschlager, C.L.; Coen, D.M.; Ricciardi, R.P.; Hogle, J.M. The crystal structure of PF-8, the DNA polymerase accessory subunit from Kaposi’s sarcoma-associated herpesvirus. J. Virol. 2009, 83, 12215–12228. [Google Scholar] [CrossRef]
- Krishna, T.S.; Kong, X.P.; Gary, S.; Burgers, P.M.; Kuriyan, J. Crystal structure of the eukaryotic DNA polymerase processivity factor PCNA. Cell 1994, 79, 1233–1243. [Google Scholar] [CrossRef]
- Stinski, M.F. Sequence of protein synthesis in cells infected by human cytomegalovirus: Early and late virus-induced polypeptides. J. Virol. 1978, 26, 686–701. [Google Scholar]
- Geballe, A.P.; Leach, F.S.; Mocarski, E.S. Regulation of cytomegalovirus late gene expression: Gamma genes are controlled by posttranscriptional events. J. Virol. 1986, 57, 864–874. [Google Scholar]
- Boccuni, M.C.; Campanini, F.; Battista, M.C.; Bergamini, G.; Dal Monte, P.; Ripalti, A.; Landini, M.P. Human cytomegalovirus product UL44 downregulates the transactivation of HIV-1 long terminal repeat. AIDS 1998, 12, 365–372. [Google Scholar] [CrossRef]
- Zhang, Q.; Hong, Y.; Dorsky, D.; Holley-Guthrie, E.; Zalani, S.; Elshiekh, N.A.; Kiehl, A.; Le, T.; Kenney, S. Functional and physical interactions between the Epstein-Barr virus (EBV) proteins BZLF1 and BMRF1: Effects on EBV transcription and lytic replication. J. Virol. 1996, 70, 5131–5142. [Google Scholar]
- Advani, S.J.; Weichselbaum, R.R.; Roizman, B. Herpes simplex virus 1 activates cdc2 to recruit topoisomerase II alpha for post-DNA synthesis expression of late genes. Proc. Natl. Acad. Sci. USA 2003, 100, 4825–4830. [Google Scholar] [CrossRef]
- Perry, R.B.; Fainzilber, M. Nuclear transport factors in neuronal function. Semin. Cell Dev. Biol. 2009, 20, 600–606. [Google Scholar] [CrossRef]
- Burch, A.D.; Weller, S.K. Herpes simplex virus type 1 DNA polymerase requires the mammalian chaperone hsp90 for proper localization to the nucleus. J. Virol. 2005, 79, 10740–10749. [Google Scholar] [CrossRef]
- Wagstaff, K.M.; Rawlinson, S.M.; Hearps, A.C.; Jans, D.A. An AlphaScreen(R)-based assay for high-throughput screening for specific inhibitors of nuclear import. J. Biomol. Screen 2011, 16, 192–200. [Google Scholar] [CrossRef]
- Wagstaff, K.M.; Sivakumaran, H.; Heaton, S.M.; Harrich, D.; Jans, D.A. Ivermectin is a specific inhibitor of importin alpha/beta-mediated nuclear import able to inhibit replication of HIV-1 and dengue virus. Biochem. J. 2012, 443, 851–856. [Google Scholar] [CrossRef]
- Tay, M.Y.; Fraser, J.E.; Chan, K.K.; Moreland, N.J.; Rathore, A.P.; Wang, C.; Vasudevan, S.G.; Jans, D.A. Nuclear localization of dengue virus (DENV) 1–4 non-structural protein 5; protection against all 4 DENV serotypes by the inhibitor Ivermectin. Antivir. Res. 2013, 99, 301–306. [Google Scholar] [CrossRef]
- Caly, L.; Wagstaff, K.M.; Jans, D.A. Nuclear trafficking of proteins from RNA viruses: Potential target for antivirals? Antivir. Res. 2012, 95, 202–206. [Google Scholar] [CrossRef]
- Haffar, O.; Dubrovsky, L.; Lowe, R.; Berro, R.; Kashanchi, F.; Godden, J.; Vanpouille, C.; Bajorath, J.; Bukrinsky, M. Oxadiazols: A new class of rationally designed anti-human immunodeficiency virus compounds targeting the nuclear localization signal of the viral matrix protein. J. Virol. 2005, 79, 13028–13036. [Google Scholar] [CrossRef]
- Kaiser, N.; Lischka, P.; Wagenknecht, N.; Stamminger, T. Inhibition of human cytomegalovirus replication via peptide aptamers directed against the nonconventional nuclear localization signal of the essential viral replication factor pUL84. J. Virol. 2009, 83, 11902–11913. [Google Scholar] [CrossRef]
- Gao, M.; DiTusa, S.F.; Cordingley, M.G. The C-terminal third of UL42, a HSV-1 DNA replication protein, is dispensable for viral growth. Virology 1993, 194, 647–653. [Google Scholar] [CrossRef]
- Silva, L.A.; Loregian, A.; Pari, G.S.; Strang, B.L.; Coen, D.M. The carboxy-terminal segment of the human cytomegalovirus DNA polymerase accessory subunit UL44 is crucial for viral replication. J. Virol. 2010, 84, 11563–11568. [Google Scholar] [CrossRef]
- Kim, Y.E.; Ahn, J.H. Role of the specific interaction of UL112-113 p84 with UL44 DNA polymerase processivity factor in promoting DNA replication of human cytomegalovirus. J. Virol. 2010, 84, 8409–8421. [Google Scholar] [CrossRef]
- Sinigalia, E.; Alvisi, G.; Segre, C.V.; Mercorelli, B.; Muratore, G.; Winkler, M.; Hsiao, H.H.; Urlaub, H.; Ripalti, A.; Chiocca, S.; et al. The human cytomegalovirus DNA polymerase processivity factor UL44 is modified by SUMO in a DNA-dependent manner. PLoS One 2012, 7, e49630. [Google Scholar]
- Mattoscio, D.; Segré, C.V.; Chiocca, S. Viral manipulation of cellular protein conjugation pathways: The SUMO lesson. World J. Virol. 2013, 2, 79–90. [Google Scholar] [CrossRef]
- Davies, R.G.; Wagstaff, K.M.; McLaughlin, E.; Loveland, K.A.; Jans, D.A. The BRCA1-binding protein BRAP2 can act as a cytoplasmic retention factor for nuclear and nuclear envelope-localizing testicular proteins. Biochim. Biophys. Acta 2013. [Google Scholar] [CrossRef]
- Hodel, M.R.; Corbett, A.H.; Hodel, A.E. Dissection of a nuclear localization signal. J. Biol. Chem. 2001, 276, 1317–1325. [Google Scholar] [CrossRef]
- Rihs, H.P.; Peters, R. Nuclear transport kinetics depend on phosphorylation-site-containing sequences flanking the karyophilic signal of the Simian virus 40 T-antigen. EMBO J. 1989, 8, 1479–1484. [Google Scholar]
- Fulcher, A.J.; Jans, D.A. Regulation of nucleocytoplasmic trafficking of viral proteins: An integral role in pathogenesis? Biochim. Biophys. Acta 2011, 1813, 2176–2190. [Google Scholar]
- Silva, L.A.; Strang, B.L.; Lin, E.W.; Kamil, J.P.; Coen, D.M. Sites and roles of phosphorylation of the human cytomegalovirus DNA polymerase subunit UL44. Virology 2011, 417, 268–280. [Google Scholar] [CrossRef]
- Prichard, M.N.; Lawlor, H.; Duke, G.M.; Mo, C.; Wang, Z.; Dixon, M.; Kemble, G.; Kern, E.R. Human cytomegalovirus uracil DNA glycosylase associates with ppUL44 and accelerates the accumulation of viral DNA. Virol. J. 2005, 2. [Google Scholar] [CrossRef] [Green Version]
- Alvisi, G.; Jans, D.A.; Ripalti, A. Human cytomegalovirus (HCMV) DNA polymerase processivity factor ppUL44 dimerizes in the cytosol before translocation to the nucleus. Biochemistry 2006, 45, 6866–6872. [Google Scholar] [CrossRef]
- Sinigalia, E.; Alvisi, G.; Mercorelli, B.; Coen, D.M.; Pari, G.S.; Jans, D.A.; Ripalti, A.; Palu, G.; Loregian, A. Role of homodimerization of human cytomegalovirus DNA polymerase accessory protein UL44 in origin-dependent DNA replication in cells. J. Virol. 2008, 82, 12574–12579. [Google Scholar] [CrossRef]
- Cozza, G.; Pinna, L.A.; Moro, S. Kinase CK2 inhibition: An update. Curr. Med. Chem. 2013, 20, 671–693. [Google Scholar] [CrossRef]
- Duncan, J.S.; Gyenis, L.; Lenehan, J.; Bretner, M.; Graves, L.M.; Haystead, T.A.; Litchfield, D.W. An unbiased evaluation of CK2 inhibitors by chemoproteomics: Characterization of inhibitor effects on CK2 and identification of novel inhibitor targets. Mol. Cell. Proteomics 2008, 7, 1077–1088. [Google Scholar] [CrossRef]
- Pei, Y.; Fu, W.; Yang, E.; Shen, A.; Chen, Y.C.; Gong, H.; Chen, J.; Huang, J.; Xiao, G.; Liu, F. A Hsp40 chaperone protein interacts with and modulates the cellular distribution of the primase protein of human cytomegalovirus. PLoS Pathog. 2012, 8, e1002968. [Google Scholar] [CrossRef]
- Smith, J.A.; Jairath, S.; Crute, J.J.; Pari, G.S. Characterization of the human cytomegalovirus UL105 gene and identification of the putative helicase protein. Virology 1996, 220, 251–255. [Google Scholar] [CrossRef]
- Stallings, C.L.; Silverstein, S. Dissection of a novel nuclear localization signal in open reading frame 29 of varicella-zoster virus. J. Virol. 2005, 79, 13070–13081. [Google Scholar] [CrossRef]
- Kyratsous, C.A.; Silverstein, S.J. BAG3, a host cochaperone, facilitates varicella-zoster virus replication. J. Virol. 2007, 81, 7491–7503. [Google Scholar] [CrossRef]
- Hanai, R.; Mashima, K. Characterization of two isoforms of a human DnaJ homologue, HSJ2. Mol. Biol. Rep. 2003, 30, 149–153. [Google Scholar] [CrossRef]
- Makhov, A.M.; Subramanian, D.; Holley-Guthrie, E.; Kenney, S.C.; Griffith, J.D. The Epstein-Barr virus polymerase accessory factor BMRF1 adopts a ring-shaped structure as visualized by electron microscopy. J. Biol. Chem. 2004, 279, 40358–40361. [Google Scholar]
- Chen, X.; Lin, K.; Ricciardi, R.P. Human Kaposi’s sarcoma herpesvirus processivity factor-8 functions as a dimer in DNA synthesis. J. Biol. Chem. 2004, 279, 28375–28386. [Google Scholar] [CrossRef]
- Zhou, X.; Liao, Q.; Ricciardi, R.P.; Peng, C.; Chen, X. Kaposi’s sarcoma-associated herpesvirus processivity factor-8 dimerizes in cytoplasm before being translocated to nucleus. Biochem. Biophys. Res. Commun. 2010, 397, 520–525. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Alvisi, G.; Jans, D.A.; Camozzi, D.; Avanzi, S.; Loregian, A.; Ripalti, A.; Palù, G. Regulated Transport into the Nucleus of Herpesviridae DNA Replication Core Proteins. Viruses 2013, 5, 2210-2234. https://doi.org/10.3390/v5092210
Alvisi G, Jans DA, Camozzi D, Avanzi S, Loregian A, Ripalti A, Palù G. Regulated Transport into the Nucleus of Herpesviridae DNA Replication Core Proteins. Viruses. 2013; 5(9):2210-2234. https://doi.org/10.3390/v5092210
Chicago/Turabian StyleAlvisi, Gualtiero, David A. Jans, Daria Camozzi, Simone Avanzi, Arianna Loregian, Alessandro Ripalti, and Giorgio Palù. 2013. "Regulated Transport into the Nucleus of Herpesviridae DNA Replication Core Proteins" Viruses 5, no. 9: 2210-2234. https://doi.org/10.3390/v5092210