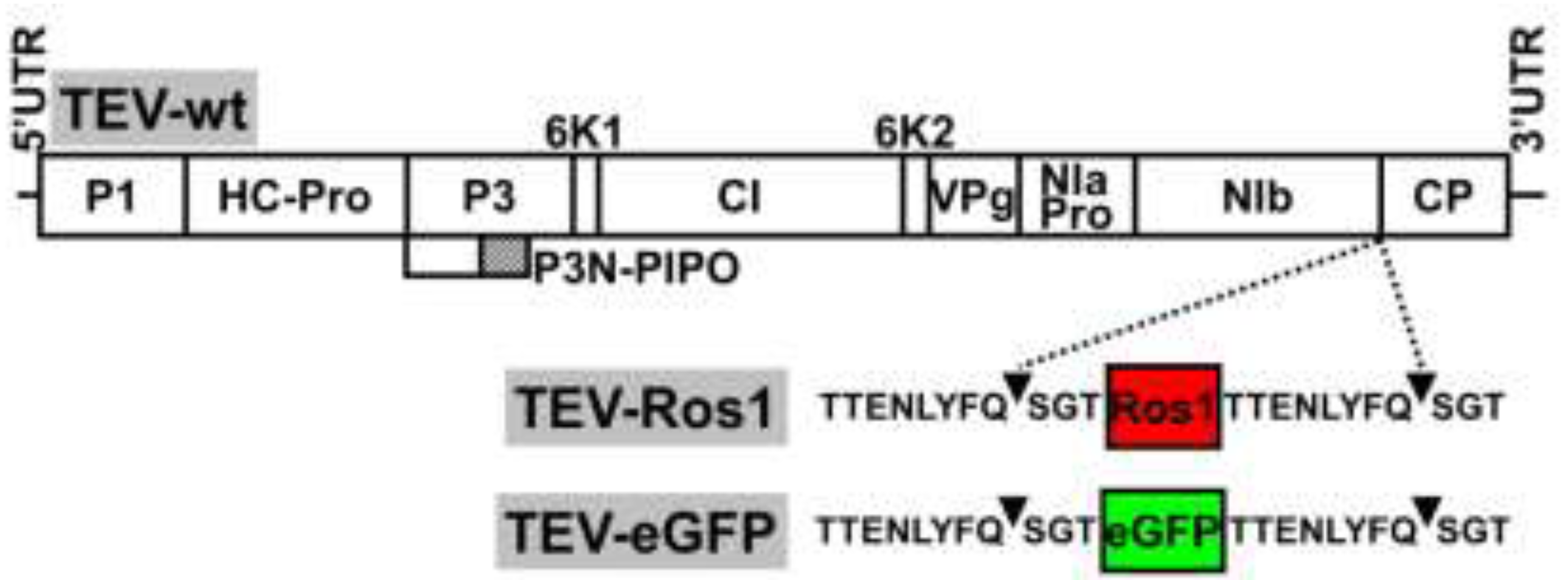
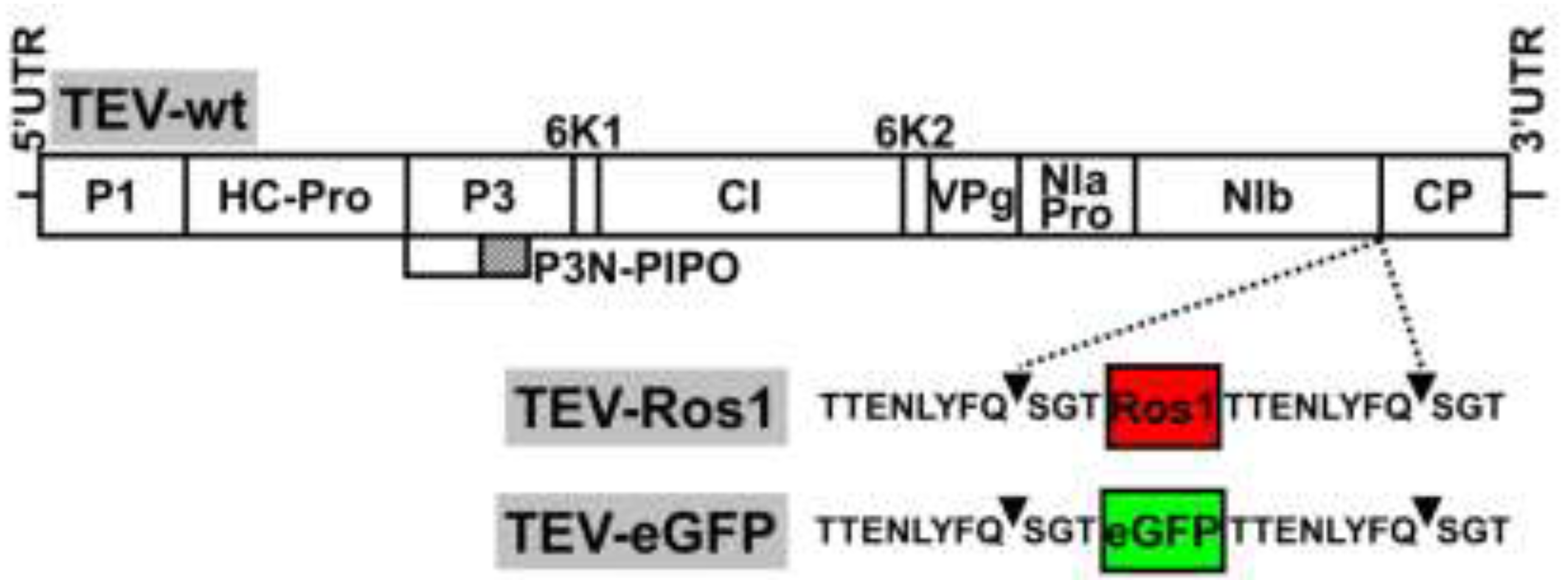
2.1. Virus Constructs
Our aim was to compare some of the properties of the recently developed Ros1 marker with the more classical eGFP as reporters of plant virus infection. We therefore constructed a TEV recombinant clone with an eGFP cDNA inserted between NIb and CP cistrons: TEV-eGFP (
Figure 1 and
Supplemental Figure 1). This new recombinant clone was absolutely parallel in sequence to our previously described TEV-Ros1 (NIb/CP) [
11] (referred to here as TEV-Ros1, for simplicity) except for the sequence of the reporter marker (
Figure 1 and
Supplemental Figure 1, see Electronic Supplementary Information). In both TEV-Ros1 and TEV-eGFP, the Ros1 and eGFP markers are released from the viral polyprotein by the activity of viral NIa proteinase on properly engineered cleavage sites flanking both markers. The sequences flanking both markers were exactly the same (
Supplemental Figure 1). After proteolytic cleavage, both markers are released with three (SGT) and eight (TTENLYFQ) additional amino acids at the amino and carboxyl termini, respectively. These additional peptides result from the engineered NIb/marker and marker/CP cleavage sites (
Figure 1).
Figure 1.
Schemes of wild-type TEV (TEV-wt) and recombinant TEV-Ros1 and TEV-eGFP. Lines represent the viral 5' and 3' untranslated regions (5'UTR and 3'UTR) and boxes represent the viral cistrons P1, HC-Pro, P3, P3N-PIPO, 6K1, CI, 6K2, VPg, NIaPro, NIb and CP, as indicated. The red and green boxes represent Ros1 and eGFP markers as indicated. Sequences of NIaPro cleavage sites flanking the markers in TEV-Ros1 and TEV-eGFP are given. Arrowheads point the exact cleavage position of the protease.
Figure 1.
Schemes of wild-type TEV (TEV-wt) and recombinant TEV-Ros1 and TEV-eGFP. Lines represent the viral 5' and 3' untranslated regions (5'UTR and 3'UTR) and boxes represent the viral cistrons P1, HC-Pro, P3, P3N-PIPO, 6K1, CI, 6K2, VPg, NIaPro, NIb and CP, as indicated. The red and green boxes represent Ros1 and eGFP markers as indicated. Sequences of NIaPro cleavage sites flanking the markers in TEV-Ros1 and TEV-eGFP are given. Arrowheads point the exact cleavage position of the protease.
2.2. Quantification of Primary Infection Foci
Nicotiana tabacum L. cv. Xanthi plants were initially agroinoculated with TEV-Ros1 and TEV-eGFP. Symptomatic tissues were collected at 7 dpi and aliquoted to more easily produce infectious extracts of both virus variants. When new
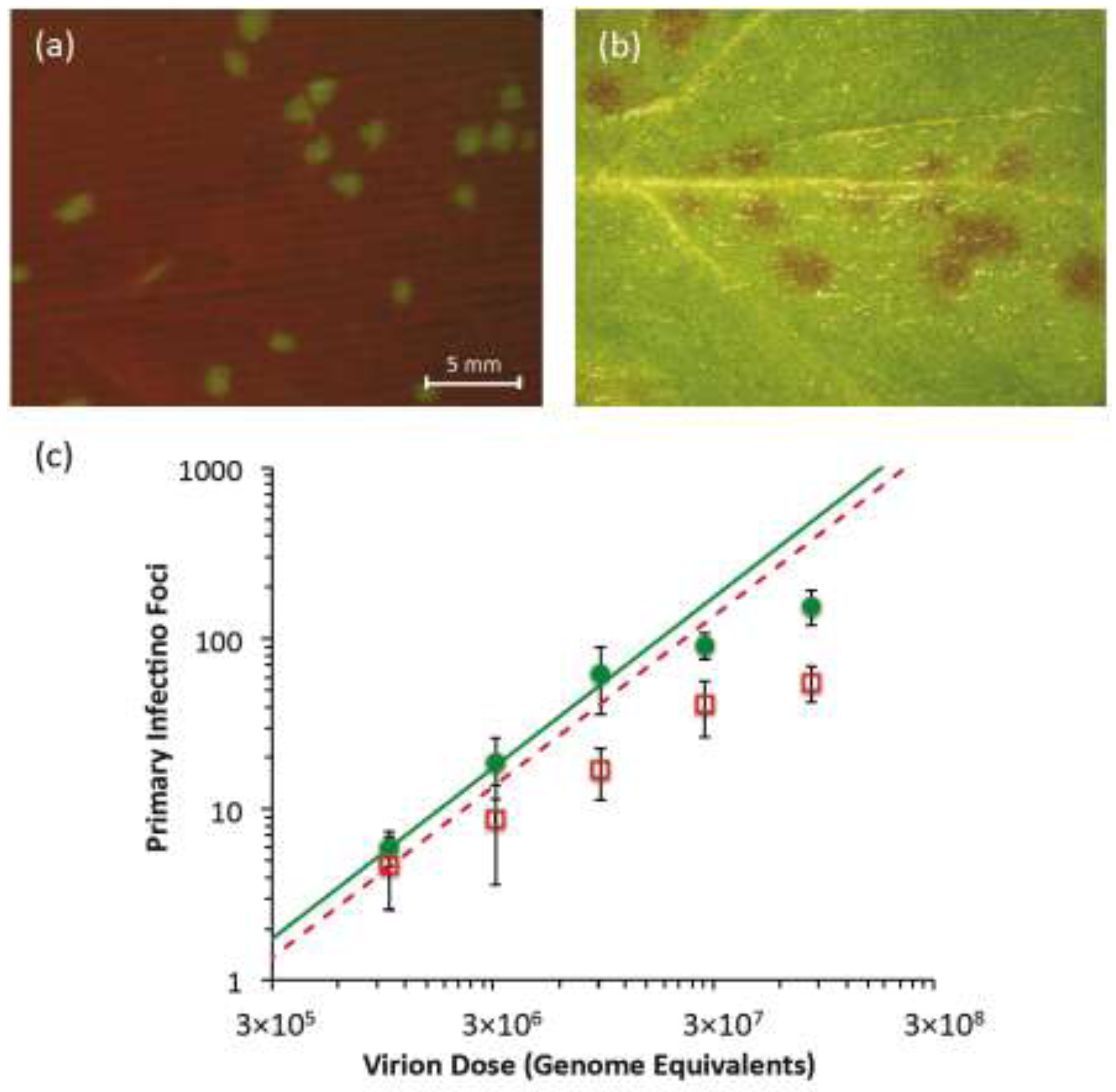
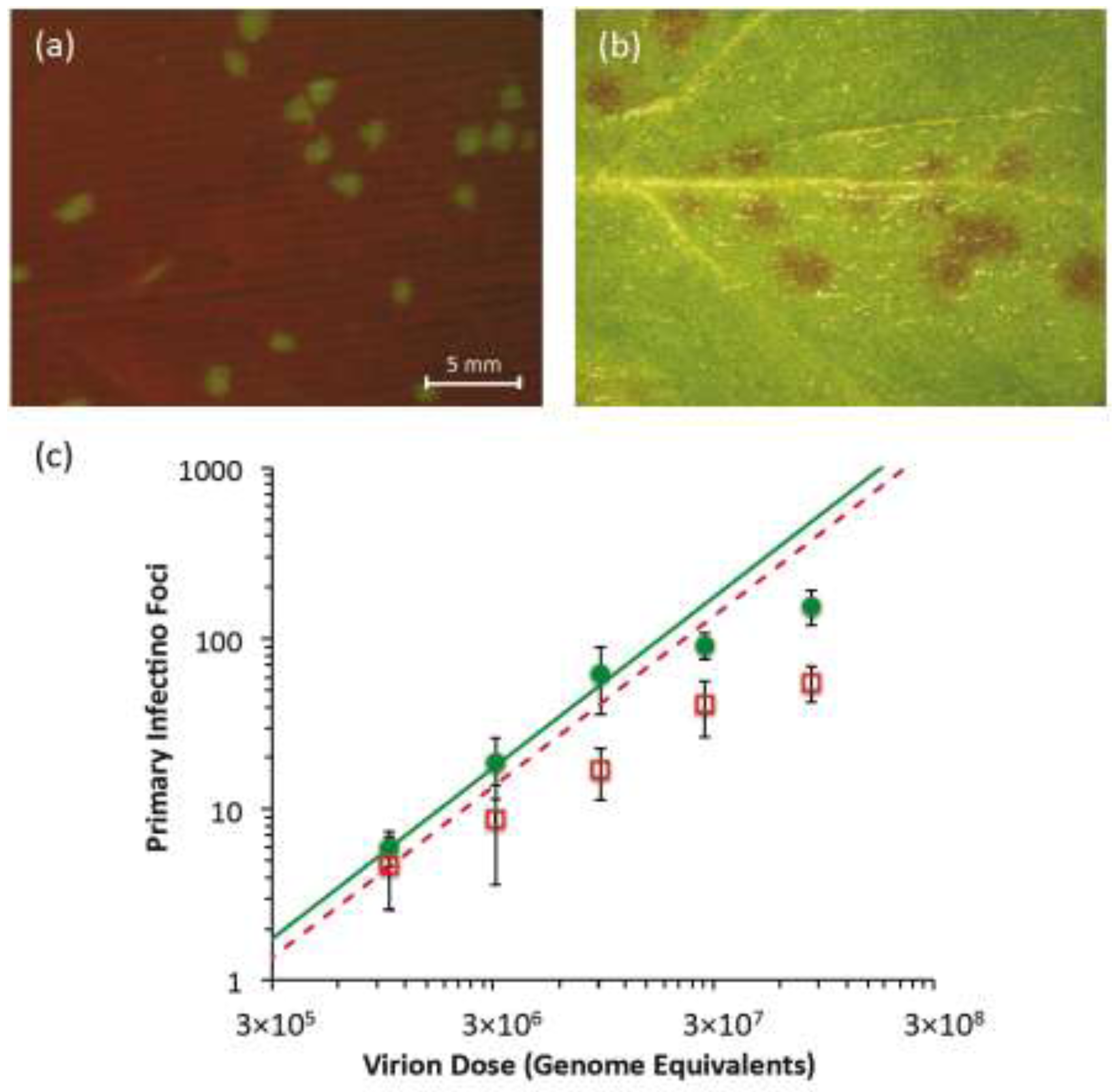
N. tabacum plants were mechanically inoculated with these infectious extracts, individual primary infection foci were detected in the inoculated leaves. In the case of TEV‑eGFP, infection foci were fluorescent and they were detected starting from 3 dpi using a fluorescent stereomicroscope (
Figure 2a). In the case of TEV-Ros1, red-colored primary infection foci were detected using a stereomicroscope under visible light conditions, although they were only visible at 5 dpi and were not as clearly defined as TEV-eGFP foci (
Figure 2b). When the dose
vs. foci relationship was plotted (
Figure 2c), the number of observed foci appears to start saturating immediately at the lowest dose for TEV-Ros1. For TEV-eGFP, saturation only occurs around 100 foci, whereas the response appears to be linear for lower doses. We then fitted the independent-action hypothesis (IAH) and dependent action (DA) infection models to the full data set using maximum-likelihood methods (
Section 3.4). The DA model was better supported for both viruses, as shown by model selection using the Akaike Information Criterion (AIC) (
Table 1). This result confirms that the data for both viruses deviate appreciably from the linear dose-foci relationship predicted by the IAH model. On the other hand, if only the lowest three doses were considered, the IAH model was better supported for TEV-eGFP than TEV-Ros1 (
Table 1). This result suggests that the dose-foci relationship is only linear for TEV-eGFP at low doses (
i.e., doses which result in <100 foci).
Figure 2.
Primary infection foci of TEV-eGFP and TEV-Ros1. In (
a), fluorescent stereomicroscope image of primary infection foci of TEV-eGFP at 3 dpi (dose = 3.09 × 10
6 genome equivalents). In (
b), visible stereomicroscope image of primary infection foci of TEV-Ros1 at 6 dpi (dose = 9.26 × 10
6 genome equivalents). In panel (
c), plot of the relationship between dose (abscissae) and the number of foci (ordinate) for TEV-eGFP (green circles) and TEV-Ros1 (red open squares). Error bars denote the standard deviation. To help interpret the data, we used the data from the lowest dose to calculate an infection probability (
Section 3.4), and used this value to plot the predicted dose-foci relationship (green line for TEV-eGFP, dotted red line for TEV-Ros1). Note that for the IAH model, the slope of this line is always the same, irrespective of the infection probability.
Figure 2.
Primary infection foci of TEV-eGFP and TEV-Ros1. In (
a), fluorescent stereomicroscope image of primary infection foci of TEV-eGFP at 3 dpi (dose = 3.09 × 10
6 genome equivalents). In (
b), visible stereomicroscope image of primary infection foci of TEV-Ros1 at 6 dpi (dose = 9.26 × 10
6 genome equivalents). In panel (
c), plot of the relationship between dose (abscissae) and the number of foci (ordinate) for TEV-eGFP (green circles) and TEV-Ros1 (red open squares). Error bars denote the standard deviation. To help interpret the data, we used the data from the lowest dose to calculate an infection probability (
Section 3.4), and used this value to plot the predicted dose-foci relationship (green line for TEV-eGFP, dotted red line for TEV-Ros1). Note that for the IAH model, the slope of this line is always the same, irrespective of the infection probability.
Table 1.
Infection models for dose-foci relationship. Model fitting and selection results are given for the dose-foci relationship (
Figure 2c) of TEV-eGFP and TEV-Ros1. Data indicates whether the full data set was used (all doses), or only the partial data set was used (low doses), which comprises only the lowest three doses. Model indicates whether the independent action hypothesis (IAH) or dependent action (DA) model was fitted. Estimated model parameters are the probability of infection per virion (
ρ) and, for the DA model, a constant that determines the strength of dose-dependent effects on
ρ (
κ). For a detailed description of the models see
Section 3.4. NLL is negative likelihood, a measure of model fit. AIC is Akaike Information Criterion, ΔAIC is the difference in AIC between the best-fitting model and other models, and AW is the Akaike Weight, the probability that the data provide most support to the model in question. For the full data sets of both viruses, the DA model is better supported than the IAH model. For the low-dose data, the IAH model is better supported for the TEV-eGFP data, indicating a linear response. On the other hand, for TEV-Ros1, the DA model is better supported even when only these data are considered.
Table 1.
Infection models for dose-foci relationship. Model fitting and selection results are given for the dose-foci relationship (Figure 2c) of TEV-eGFP and TEV-Ros1. Data indicates whether the full data set was used (all doses), or only the partial data set was used (low doses), which comprises only the lowest three doses. Model indicates whether the independent action hypothesis (IAH) or dependent action (DA) model was fitted. Estimated model parameters are the probability of infection per virion (ρ) and, for the DA model, a constant that determines the strength of dose-dependent effects on ρ (κ). For a detailed description of the models see Section 3.4. NLL is negative likelihood, a measure of model fit. AIC is Akaike Information Criterion, ΔAIC is the difference in AIC between the best-fitting model and other models, and AW is the Akaike Weight, the probability that the data provide most support to the model in question. For the full data sets of both viruses, the DA model is better supported than the IAH model. For the low-dose data, the IAH model is better supported for the TEV-eGFP data, indicating a linear response. On the other hand, for TEV-Ros1, the DA model is better supported even when only these data are considered.
| Virus | Data | Model | Parameter estimates | NLL | AIC | ΔAIC | AW |
|---|
| TEV-eGFP | All doses | IAH | ρ = 2.63 × 10−6 | 302.719 | 607.438 | 299.944 | 0 |
| | | DA | ρ = 2.95 × 10−3 ; κ = 0.60 | 151.747 | 397.494 | - | 1 |
| | Low doses | IAH | ρ = 6.46 × 10−6 | 61.456 | 124.912 | - | 0.601 |
| | | DA | ρ = 1.86 × 10−6 ; κ = 1.08 | 60.865 | 125.730 | 0.818 | 0.399 |
| TEV-Ros1 | All doses | IAH | ρ = 1.02 × 10−6 | 139.048 | 280.097 | 129.090 | 0 |
| | | DA | ρ = 2.69 × 10−3; κ = 0.55 | 73.504 | 151.007 | - | 1 |
| | Low doses | IAH | ρ = 2.29 × 10−6 | 39.012 | 80.024 | 10.508 | 0.005 |
| | | DA | ρ = 1.32 × 10−3; κ = 0.59 | 32.758 | 69.516 | - | 0.995 |
The number of primary infection foci renders an estimate of virus effective population size (
Ne) for virus infection of a plant [
8], because
Ne can be approximated as the smallest census size for rapidly expanding and contracting population [
15]. Here we found that TEV-Ros1 did not perform as well as TEV-eGFP for the quantification of primary infection foci. Infection foci were only visible starting from 5 dpi, at which point they were too diffuse to quantify well at high virus doses. Moreover, the dose-foci relationship appears to saturate much more quickly for TEV-Ros1 than for TEV-eGFP. In contrast to eGFP, the Ros1 marker may therefore be unsuitable for accurate quantification of primary infection foci. However, the appearance of primary infection foci may depend on exact inoculation and growing conditions, and on the virus in which the marker is inserted. For example, during inoculation of larger leaves in older plants, individual primary infection foci were more easily discriminated [
16]. Moreover, the marker may still be useful in experiments where a semi‑quantitative assessment of the primary infection foci is needed. The dose-foci relationship has been reported to saturate for TEV‑eGFP at high doses [
5]. Unlike TEV-Ros1, TEV-eGFP foci can be readily quantified when the number of foci <100 [
8,
9]. Rejection of the IAH model at high TEV-eGFP doses could be due to real deviations from the IAH model, such as a limited number of infectious sites in the inoculated leaf [
17]. On the other hand, it might simply be difficult to distinguish foci when high doses are used and there are a large number of foci.
2.3. Within-Host Competitive Fitness of TEV-Ros1 and TEV-eGFP
We quantified the within-host competitive fitness (
W; see
Section 3.5) of wild-type TEV (TEV-wt) [
11], TEV-Ros1 and TEV-eGFP in
N. tabacum using TEV-mCherry [
8] (a similar virus variant carrying the fluorescent protein mCherry in0 between the P1 and HC-Pro cistrons) as a common competitor (
Figure 3).
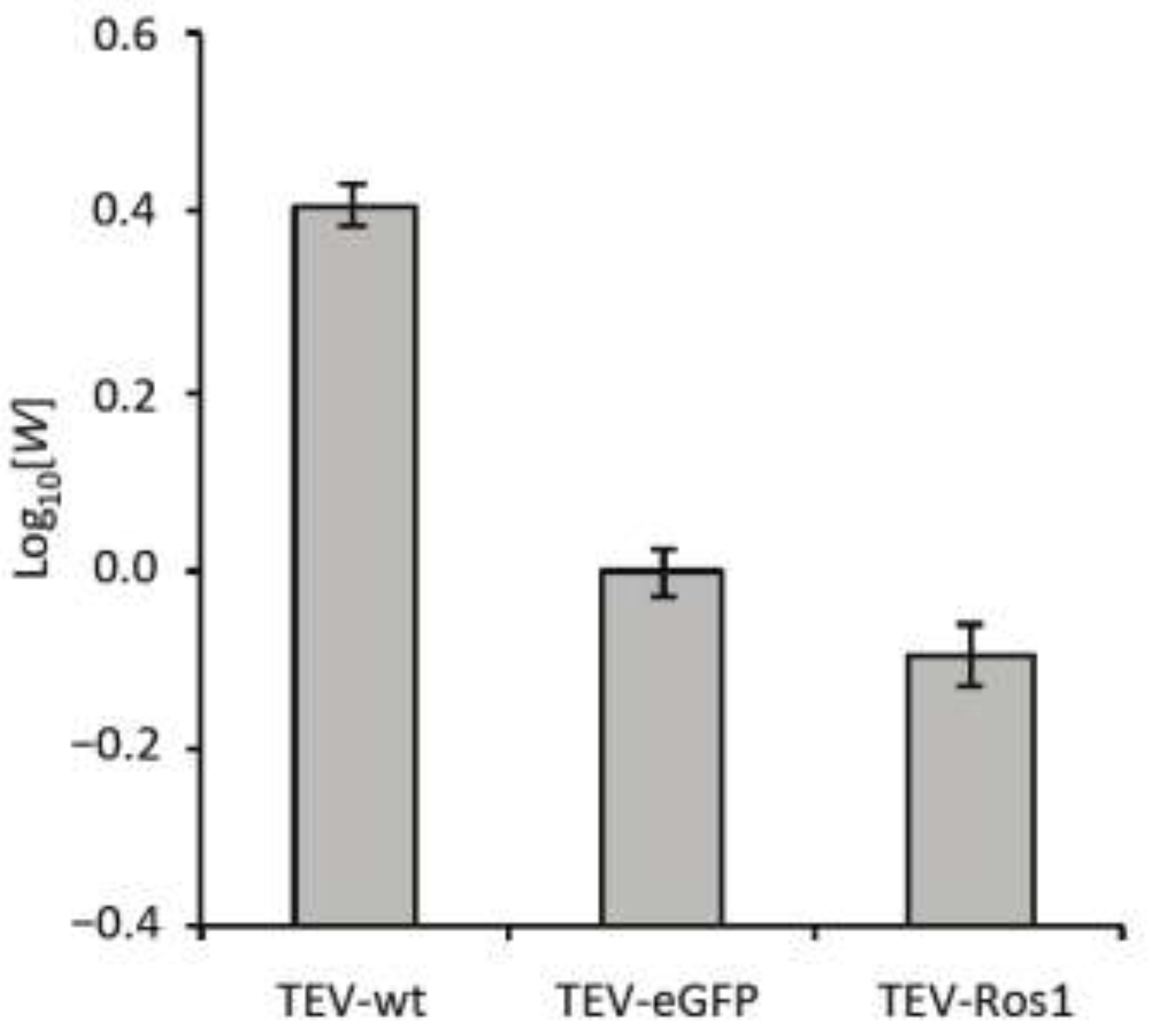
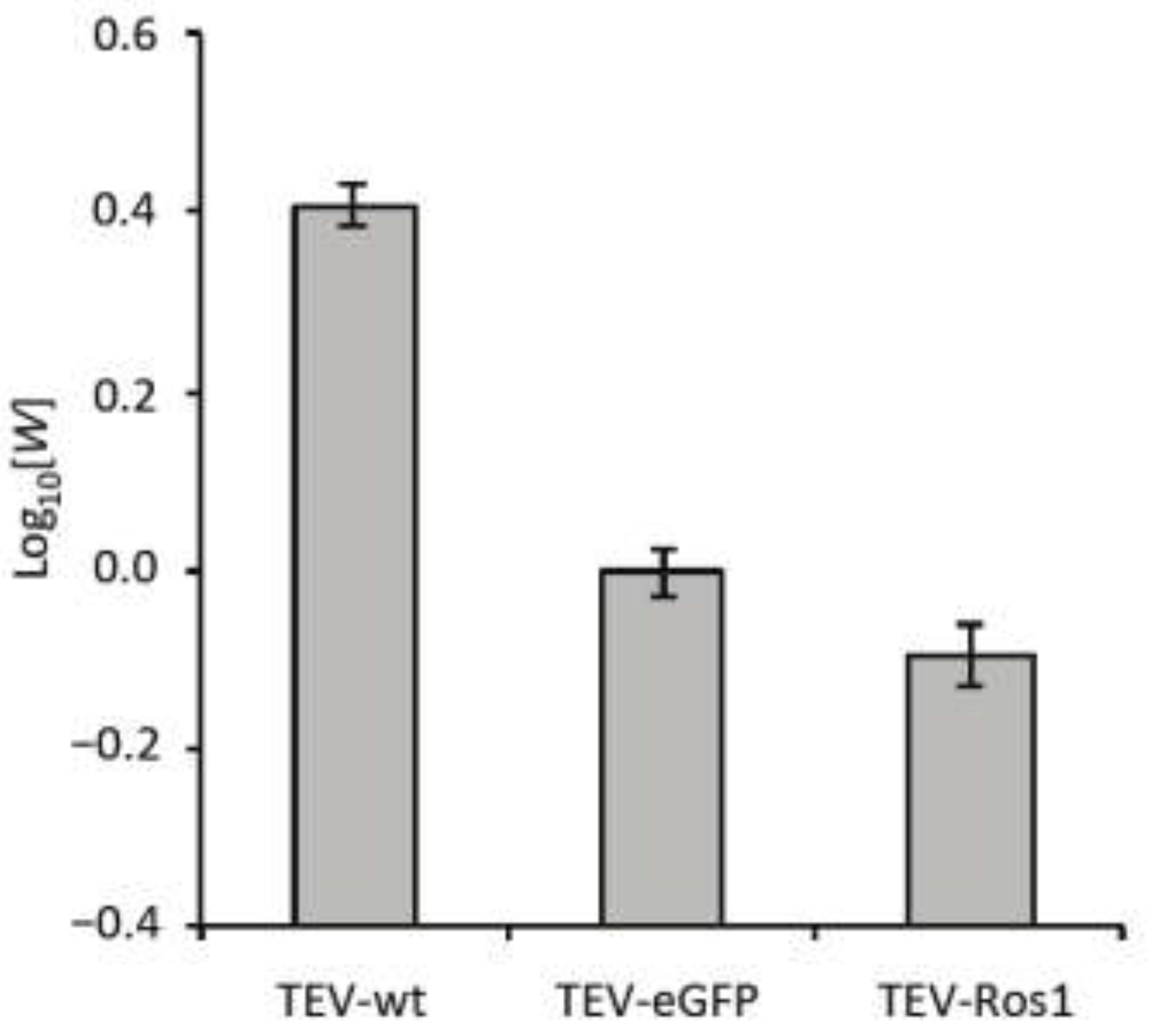
Figure 3.
Competitive within-host fitness of TEV-wt TEV-eGFP and TEV-Ros1. Plot of the log10-transformed within-host competitive fitness (W) for wild-type TEV (TEV-wt) and its two derived viruses (TEV-eGFP and TEV-Ros1) carrying markers, as measured by direct competitions with TEV-mCherry. Error bars indicate the standard deviation.
Figure 3.
Competitive within-host fitness of TEV-wt TEV-eGFP and TEV-Ros1. Plot of the log10-transformed within-host competitive fitness (W) for wild-type TEV (TEV-wt) and its two derived viruses (TEV-eGFP and TEV-Ros1) carrying markers, as measured by direct competitions with TEV-mCherry. Error bars indicate the standard deviation.
Overall, there were highly significant differences between the viruses in fitness (ANOVA: F2,12 = 432.381, P < 0.001). TEV-wt had a significantly higher within-host fitness than the TEV-eGFP (independent samples t-test with Holm-Bonferroni correction were used as a post hoc test: t = 25.213, 8 d.f., P < 0.001), whereas TEV-Ros1 had a significantly lower fitness than TEV-eGFP (t = −4.795, 8 d.f., P = 0.001). Therefore, even though TEV-Ros1 has a slightly shorter marker sequence than TEV‑eGFP, its fitness was significantly lower. The difference between TEV-eGFP and TEV-Ros1 is, however, small compared to differences between these two viruses and TEV-wt.
The within-host competitive fitness data have two ramifications that should be given consideration. First, infection dynamics of both marked viruses will probably not be exactly the same as the wild-type virus, given that there is a fitness cost associated to the marker. Second, the difference in fitness between TEV-Ros1 and a TEV-Ros1-derived deletion mutant can in principle be larger than for TEV‑eGFP, although this will depend on the exact deletion variant. Therefore, ceteris paribus a TEV-Ros1 population could show substantial losses of the marker before a TEV-eGFP population.
2.4. Marker Instability in TEV-Ros1 and TEV-eGFP
After seven weeks of infection in
N. tabacum, we assessed the integrity of the marker gene for TEV-Ros1 and TEV-eGFP, using RT-PCR with primers flanking the marker gene on RNA extracted from the 30th leaf (
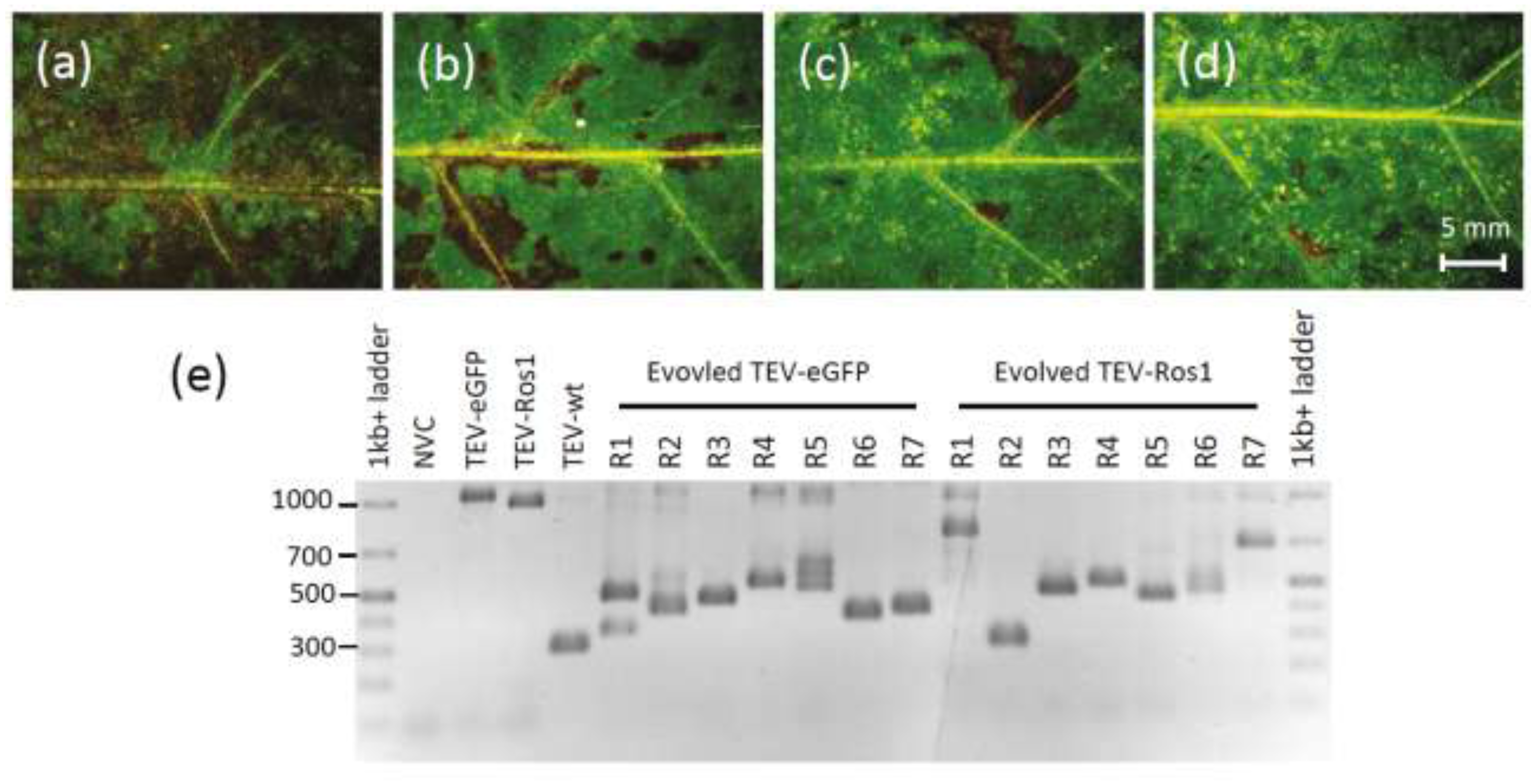
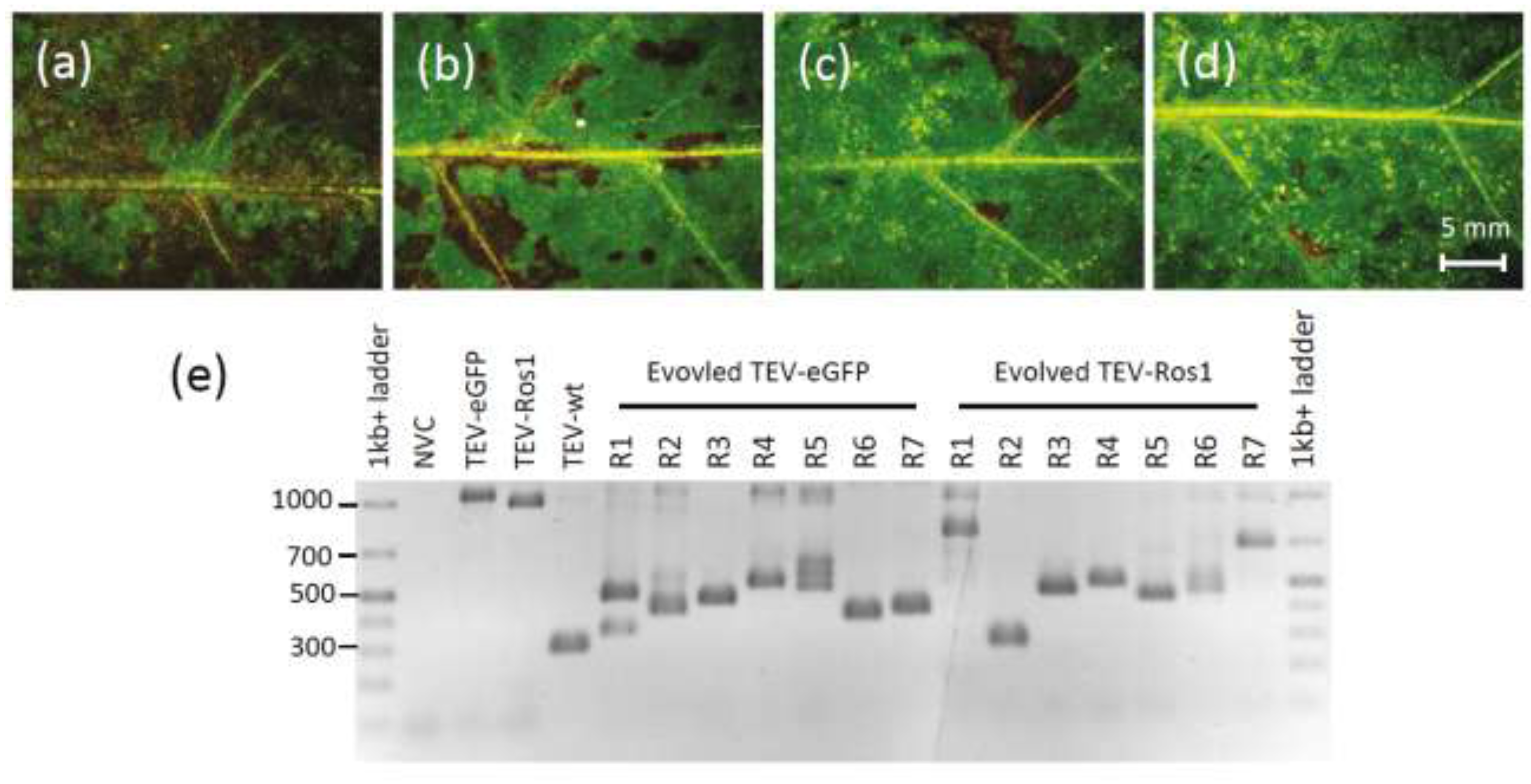
Section 3.6). Visual examination revealed that deletions of the marker gene had probably occurred in TEV-Ros1-infected plants (
Figure 4a–d). Note that direct visual examination of TEV-eGFP-infected plants is not informative. RT-PCR showed that most plants had accumulated deletions at this point for both marked viruses, and in some plants the ancestral viruses could no longer be detected (
Figure 4e). We then classified the plants based on the presence of the ancestral virus and did not find a significant difference between the TEV-Ros1 and TEV-eGFP (
Table 2). We also performed a second analysis comparing the evolved populations of TEV-Ros1 and TEV-eGFP, which considers both the length and relative intensity of PCR amplicons by means of a weighted mean (
Section 3.6). With this procedure we also found no significant differences between the two viruses (Mann-Whitney
U-test:
P = 0.865). Our data therefore suggest that the stability of TEV-Ros1 and TEV-eGFP are similar, despite the minor difference in within-host competitive fitness.
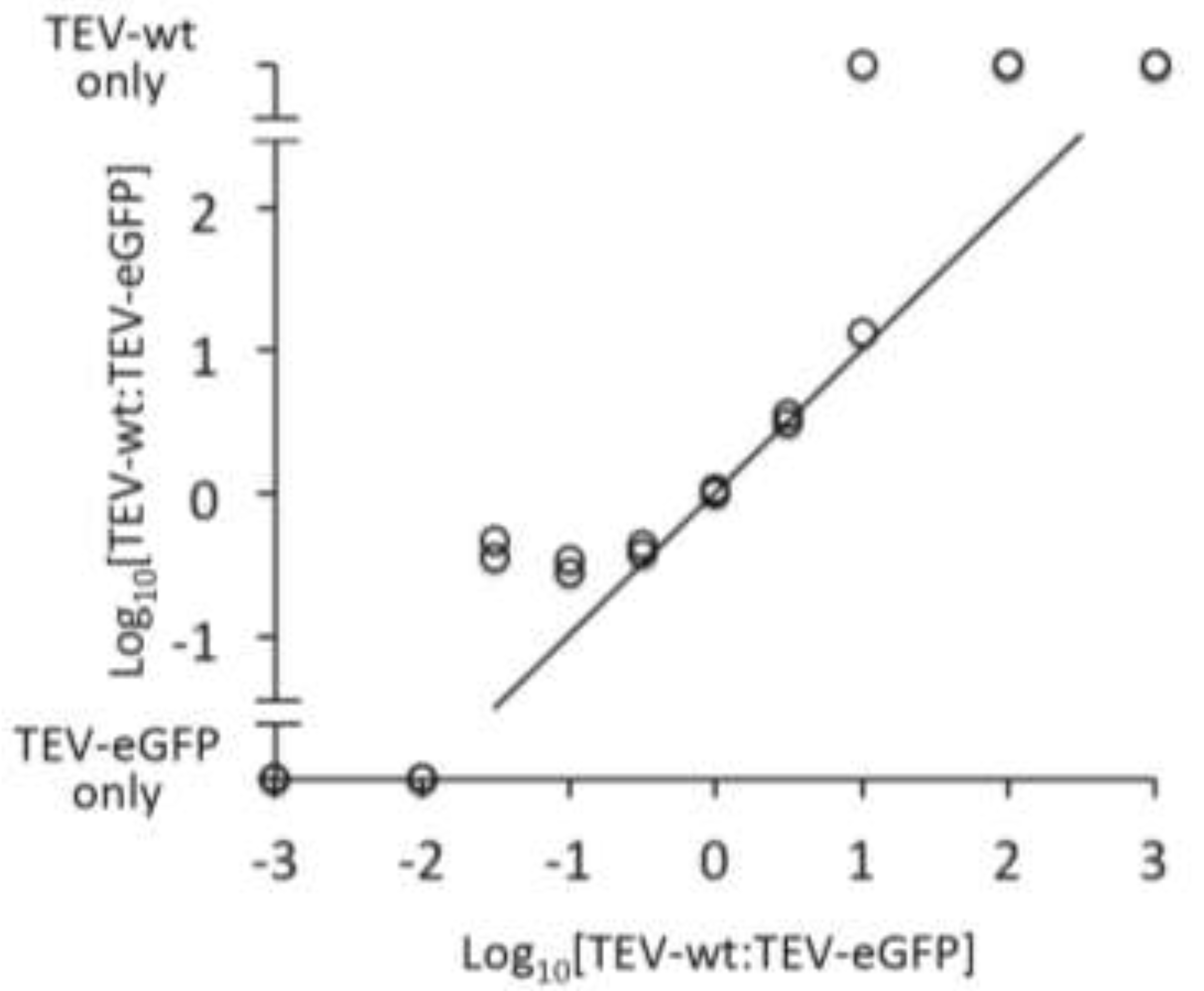
During a PCR, smaller products may be amplified with a greater efficiency than larger products [
18]. Selective amplification of smaller products could in turn bias our comparison of the weighted means of PCR amplicons towards smaller products, lowering the sensitivity of this test. We therefore performed a control experiment to ensure the quantification of the PCR amplicons did not strongly bias the results (
Section 3.6). In essence, we compared known input ratios to the output ratio generated by the assay. The results (
Figure 5) show that our PCR assay is not very sensitive: the rare variant was never detected with ratios TEV-wt:TEV-eGFP ≤1:100 or ≥100:1. However, for a range from 3:1 to 1:10 the input and output of the control were very similar. For the experimental RT-PCR stability data, the vast majority of samples had a high frequency of deletion variants, and low frequencies of the ancestral marker virus. These conditions therefore coincide well with the range in which the assay appears to work best, supporting the use of our test for differences in marker stability using weighted means. Note that we did not perform a correction for the size of PCR amplicons (
i.e., output is the raw ratio of the signal of the two amplicons on gel). Therefore, under the conditions used the assay appears to work well because the higher efficiency of PCR amplification for smaller products [
18] is counterbalanced by their lower intensity in an agarose gel stained with ethidium bromide.
Figure 4.
(a–d) Leaves 10, 15, 20 and 25, respectively, of a plant infected with TEV-Ros1 after seven weeks of infection. Although all the tissues shown are fully symptomatic, anthocyanin expression appears to decrease, being almost entirely lost in leaf 25. RT-qPCR confirmed the loss of the marker gene in this particular plant. (e) An agarose gel containing RT-PCR products with primers flanking the marker gene is shown. The GeneRuler 1 kb+ DNA Ladder (Thermo Scientific) was used and NVC is a non-virus control (mock‑inoculated plant). Note that in most of the 7-week-infected plants there are PCR products smaller than the ancestral virus visible, indicating the occurrence of deletions in the marker gene.
Figure 4.
(a–d) Leaves 10, 15, 20 and 25, respectively, of a plant infected with TEV-Ros1 after seven weeks of infection. Although all the tissues shown are fully symptomatic, anthocyanin expression appears to decrease, being almost entirely lost in leaf 25. RT-qPCR confirmed the loss of the marker gene in this particular plant. (e) An agarose gel containing RT-PCR products with primers flanking the marker gene is shown. The GeneRuler 1 kb+ DNA Ladder (Thermo Scientific) was used and NVC is a non-virus control (mock‑inoculated plant). Note that in most of the 7-week-infected plants there are PCR products smaller than the ancestral virus visible, indicating the occurrence of deletions in the marker gene.
Table 2.
Marker instability for TEV-Ros1 and TEV-eGFP. After seven weeks of infection in N. tabacum, RT-PCR was used to detect deletions in the marker gene in individual plants. We then categorized the evolved populations as either containing the ancestral virus (i.e., having a PCR amplicon corresponding to the ancestral virus) or not. There was not a significant difference between the two viruses (χ2 test: χ2 = 1.414, 1 d.f., P = 0.234). Note that for both viruses, in two plants we detected only ancestral virus and no deletion mutants (and hence here they are classified as having the ancestral virus present), an insignificant difference (Binomial test: P = 1).
Table 2.
Marker instability for TEV-Ros1 and TEV-eGFP. After seven weeks of infection in N. tabacum, RT-PCR was used to detect deletions in the marker gene in individual plants. We then categorized the evolved populations as either containing the ancestral virus (i.e., having a PCR amplicon corresponding to the ancestral virus) or not. There was not a significant difference between the two viruses (χ2 test: χ2 = 1.414, 1 d.f., P = 0.234). Note that for both viruses, in two plants we detected only ancestral virus and no deletion mutants (and hence here they are classified as having the ancestral virus present), an insignificant difference (Binomial test: P = 1).
| Virus | Detection of ancestral virus by RT-PCR |
|---|
| Present | Absent |
|---|
| TEV-Ros1 | 25 (67.6%) | 12 (32.4%) |
| TEV-eGFP | 16 (53.3%) | 14 (46.7%) |
Figure 5.
The results of a control experiment to determine whether smaller PCR amplicons were selectively amplified are given. The abscissa is the log10-transformed input ratio, whilst the ordinate is the log10-transformed output ratio. Note that samples containing only TEV-wt or TEV-eGFP are also depicted on the ordinate. Circles represent the data, and the 1:1 relationship between input and output is also given for reference.
Figure 5.
The results of a control experiment to determine whether smaller PCR amplicons were selectively amplified are given. The abscissa is the log10-transformed input ratio, whilst the ordinate is the log10-transformed output ratio. Note that samples containing only TEV-wt or TEV-eGFP are also depicted on the ordinate. Circles represent the data, and the 1:1 relationship between input and output is also given for reference.
Two tests of marker instability were performed: a χ2 test for the presence of the ancestral virus and a second procedure developed here, a Mann-Whitney U-test on the mean relative amplicon length for the genomic region containing the marker. The latter procedure may in principle be useful for detecting more subtle differences between evolved virus populations than simply the presence or absence of the ancestral virus (i.e., the χ2 test). For example, the amplicon-length-based procedure might still detect differences if (i) the frequency of the ancestral virus tends to be higher in the evolved populations of one virus type, but (ii) the number of lineages in which deletions of the marker gene has occurred is the same for both virus types. In practice, however, for both procedures we did not find a difference in instability between the two viruses, and there is therefore not yet any evidence that the amplicon-length-based procedure has greater power for detecting differences in instability.
After seven weeks of infection plants infected with TEV-Ros1 were significantly shorter than those infected with TEV-eGFP (Mann-Whitney U-test: P < 0.001), suggesting that Ros1 expression is adversely affecting plant growth. When the Ros1 maker was lost during virus expansion, it appeared as if plants grew faster in those regions of the plant in which virus expressing Ros1 were no longer present. However, this effect was not statistically significant for either virus; those plants in which we sampled only viruses that did not express the marker gene in the 30th leaf were not significantly larger than plants in which marker genes were detected (Mann-Whitney U-test; TEV-Ros1: P = 0.355, TEV‑eGFP: P = 0.181).
The
N. tabacum genome includes the
An2 gene, coding for an MYB transcription factor and a homolog of
A. majus Ros1 [
19]. During infection RNA silencing mechanisms might, therefore, potentially interfere either with the expression of the endogenous
An2 or the virally expressed Ros1. However, our experimental observations suggest that interference does not occur. One explanation for the lack of RNA silencing in infected tissues is the activity of TEV HC-Pro, a strong suppressor of RNA silencing. Nonetheless, subtle alterations in expression of the endogenous
An2 may contribute to the larger phenotypic effects induced by Ros1, in contrast to eGFP.
Finally, the distribution of mutational effects on viral fitness might be different for the eGFP and Ros1 markers. Single-nucleotide mutations in the eGFP marker are unlikely to have an effect on viral fitness, because fitness is independent of fluorophore activity. On the other hand, mutations in Ros1 leading to a loss of anthocyanin-biosynthesis induction may be beneficial for viral fitness. Nevertheless, TEV has a relatively low mutation rate [
20] and single-nucleotide mutations have been observed to accumulate slowly during experimental evolution [
21]. These observations suggest that in the vast majority of cases the marker protein will be inactivated by genomic deletions rather than single‑nucleotide mutations.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

 , where κ is constant that determines the strength of dose-dependent effects. When κ > 1 there are synergistic effects, when κ < 1 there are antagonistic effects, and when κ = 1 the DA model collapses to the IAH model without dose‑dependent effects on the probability of infection per virion. Both models were fitted to the data using a maximum-likelihood approach. Given that the distribution of primary infection foci for a given dose follows approximately a Poisson distribution for low and intermediate doses [8,9], the likelihood of any observed number of foci k is
, where κ is constant that determines the strength of dose-dependent effects. When κ > 1 there are synergistic effects, when κ < 1 there are antagonistic effects, and when κ = 1 the DA model collapses to the IAH model without dose‑dependent effects on the probability of infection per virion. Both models were fitted to the data using a maximum-likelihood approach. Given that the distribution of primary infection foci for a given dose follows approximately a Poisson distribution for low and intermediate doses [8,9], the likelihood of any observed number of foci k is  . The negative log likelihood was then minimized by grid searches using R 2.14 [29], rendering model parameter estimates and allowing for model selection using the AIC.
. The negative log likelihood was then minimized by grid searches using R 2.14 [29], rendering model parameter estimates and allowing for model selection using the AIC. , where nCP and nmCherry are the copy numbers of CP and mCherry cistrons, respectively, as measured using the RT-qPCR assay. We then use the approach of [26] to measure the replicative advantage (W) such that
, where nCP and nmCherry are the copy numbers of CP and mCherry cistrons, respectively, as measured using the RT-qPCR assay. We then use the approach of [26] to measure the replicative advantage (W) such that  . Here t is the time in days, Rt is the virus ratio at the end of the experiment, and R0 is the virus ratio at the start of the experiment. Note that we used a 3:1 mixture of the two viruses in the inoculum, instead of a 1:1 mixture, for methodological reasons. R can only be accurately measured when nmCherry levels are considerably lower (i.e., one-half fold) than nCP levels. We anticipated that the within-host competitive fitness of TEV-Ros1 might be low, and starting with a 3:1 ratio ensures nmCherry levels remain lower than nCP levels.
. Here t is the time in days, Rt is the virus ratio at the end of the experiment, and R0 is the virus ratio at the start of the experiment. Note that we used a 3:1 mixture of the two viruses in the inoculum, instead of a 1:1 mixture, for methodological reasons. R can only be accurately measured when nmCherry levels are considerably lower (i.e., one-half fold) than nCP levels. We anticipated that the within-host competitive fitness of TEV-Ros1 might be low, and starting with a 3:1 ratio ensures nmCherry levels remain lower than nCP levels. ). A Mann-Whitney U-test was then performed to test for significant differences in μ values for the two viruses.
). A Mann-Whitney U-test was then performed to test for significant differences in μ values for the two viruses.