Complete Nucleotide Sequence of Watermelon Chlorotic Stunt Virus Originating from Oman

Abstract
:1. Introduction
2. Results and Discussion
2.1. Cloning and Sequencing
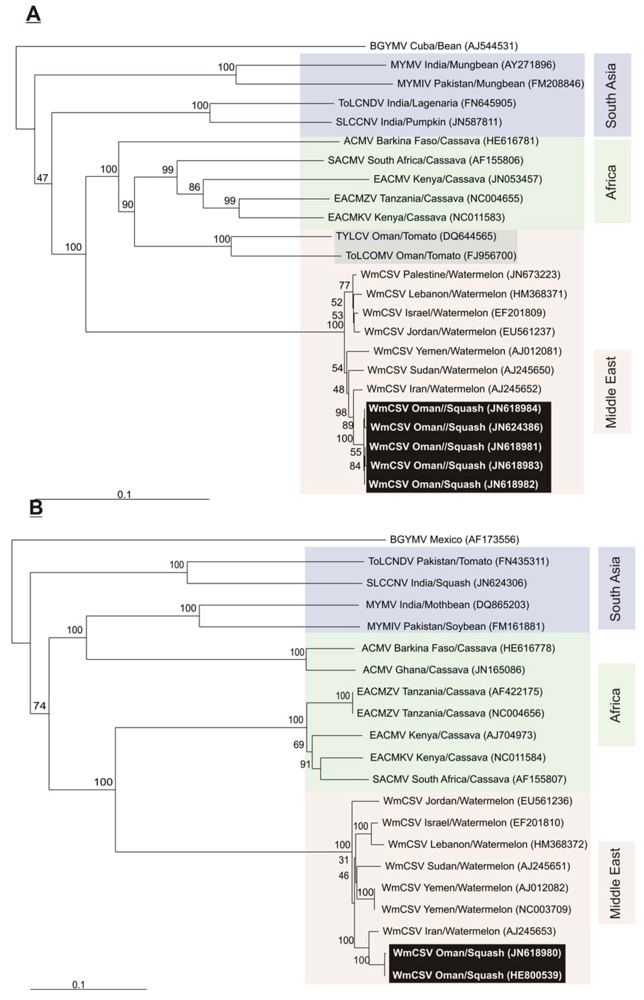
2.2. Sequence Comparisons and Analysis

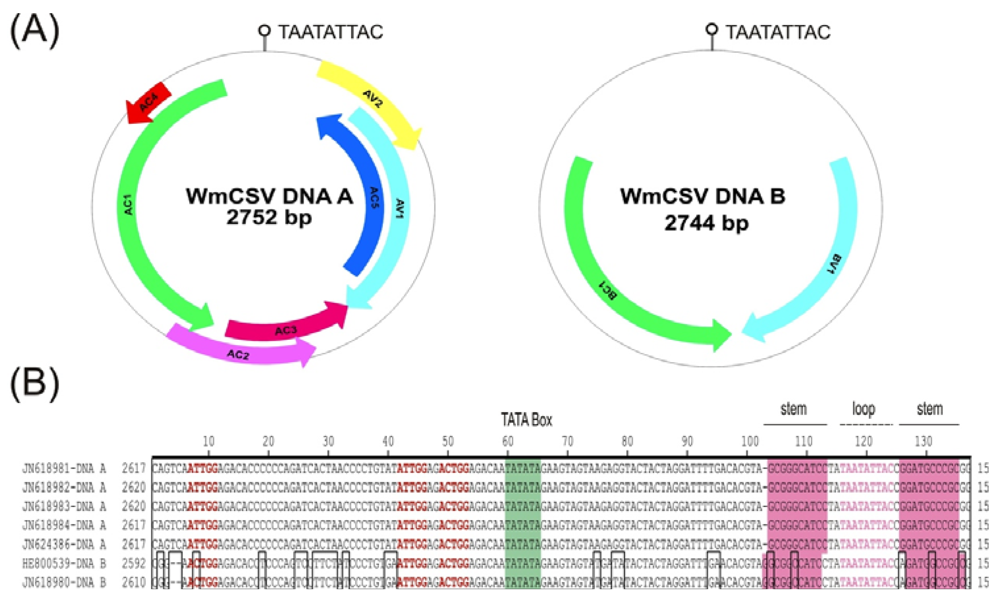{kind=link}
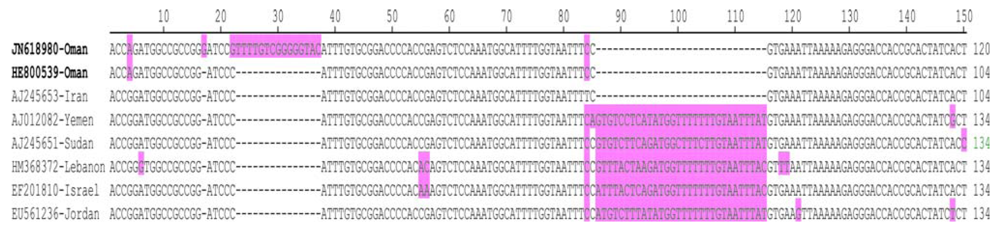{kind=link}
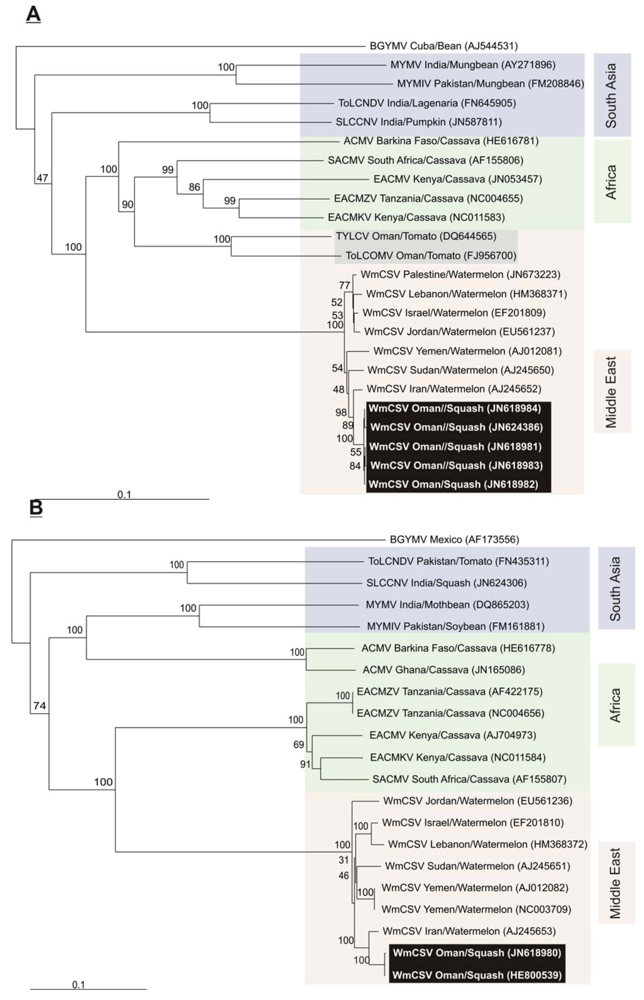{kind=link}
{kind=link}
| Isolate | Isolate Descriptor | DNA A | DNA B | |||||||||||
| Accession Number | Size (nt) | Position of Genes (Coordinates of Start/Stop Codons [Predicted Coding Capacity in kDa]) | Accession Number | Size (nt) | Position of Genes (Coordinates of Start/Stop Codons [Predicted Coding Capacity in kDa]) | |||||||||
| AV1 | AV2 | AC1 | AC2 | AC3 | AC4 | AC5 | BV1 | BC1 | ||||||
| Als-1 | [OM:Squ1:11] | JN618981 | 2752 | 315–1094 (28.6) | 155–514 (13.2) | 1540–2625 (39.8) | 1233–1640 (15.0) | 1088–1492 (14.9) | 2328–2471 (5.3) | 219–986 (28.2) | JN618980 | 2744 | 516–1271 (27.7) | 1308–2231 (33.9) |
| Als-2 | [OM:Squ2:11] | JN618982 | 2755 | 315–1094 (28.6) | 155–514 (13.2) | 1540–2628 (39.9) | 1233–1643 (15.1) | 1088–1492 (14.9) | 2331–2474 (5.4) | 219–986 (28.2) | HE800539 | 2726 | 500–1255 (27.7) | 1295–2215 (33.8) |
| Als-3 | [OM:Squ3:11] | JN618983 | 2755 | 315–1094 (28.6) | 155–514 (13.2) | 1540–2628 (39.9) | 1233–1643 (15.1) | 1088–1492 (14.9) | 2331–2474 (5.4) | 219–986 (28.2) | _ | _ | _ | _ |
| Als-4 | [OM:Squ4:11] | JN618984 | 2752 | 315–1094 (28.6) | 155–514 (13.2) | 1540–2625 (39.8) | 1233–1640 (15.0) | 1088–1492 (14.9) | 2328–2471 (5.3) | 219–986 (28.2) | _ | _ | _ | _ |
| Als-5 | [OM:Squ5:11] | JN624386 | 2752 | 315–1091 (28.5) | 155–514 (13.2) | 1540–2625 (39.8) | 1233–1640 (15.0) | 1088–1492 (14.9) | 2328–2471 (5.3) | 219–986 (28.2) | _ | _ | _ | _ |
| WmCSV Isolates from Oman | WmCSV isolates from the databases * (accession numbers of DNA A/DNA B) | |||||||
|---|---|---|---|---|---|---|---|---|
| Component | Isolate | Iran (AJ245652/AJ245653) | Sudan (AJ245650/AJ245651) | Yemen (AJ012081/AJ012082) | Jordan (EU561237/EU561236) | Israel (EF201809/EF201810) | Lebanon (HM368371/HM368372) | Palestine * (JN673223) |
| DNA A | Als-1 | 98.8 | 98.0 | 96.7 | 97.7 | 97.7 | 97.6 | 97.9 |
| Als-2 | 98.6 | 97.8 | 96.6 | 97.5 | 97.6 | 97.5 | 97.7 | |
| Als-3 | 98.5 | 97.7 | 96.5 | 97.4 | 97.5 | 97.4 | 97.5 | |
| Als-4 | 98.8 | 98.0 | 96.7 | 97.7 | 97.8 | 97.7 | 98.0 | |
| Als-5 | 98.7 | 98.0 | 96.6 | 97.6 | 97.7 | 97.6 | 98.0 | |
| DNA B | Als-1 | 96.9 | 92.2 | 93.4 | 91.9 | 92.5 | 91.8 | - |
| Als-2 | 97.4 | 92.5 | 93.3 | 93.4 | 92.9 | 92.2 | - | |


2.3. Infectivity of WmCSV

3. Experimental Section
3.1. Sample Collection and DNA Extraction
3.2. Amplification, Cloning and Sequencing of Virus Components
3.3. Sequence Analysis
3.4. Agrobacterium-Mediated Inoculation
3.5. Diagnostic PCR
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Brown, J.K.; Fauquet, C.M.; Briddon, R.W.; Zerbini, M.; Moriones, E.; Navas-Castillo, J. Geminiviridae. In Virus Taxonomy - Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Associated Press, Elsevier Inc.: London, UK, 2012; pp. 351–373. [Google Scholar]
- Moffat, A.S. Geminiviruses emerge as serious crop threat. Science 1999, 286, 1835. [Google Scholar] [CrossRef]
- Nawaz-ul-Rehman, M.S.; Fauquet, C.M. Evolution of geminiviruses and their satellites. FEBS Lett. 2009, 583, 1825–1832. [Google Scholar] [CrossRef]
- Briddon, R.W.; Stanley, J. Sub-viral agents associated with plant-infecting single-stranded DNA viruses. Virology 2006, 344, 198–210. [Google Scholar] [CrossRef]
- Mansoor, S.; Briddon, R.W.; Bull, S.E.; Bedford, I.D.; Bashir, A.; Hussain, M.; Saeed, M.; Zafar, M.Y.; Malik, K.A.; Fauquet, C.; et al. Cotton leaf curl disease is associated with multiple monopartite begomoviruses supported by single DNA β. Arch. Virol. 2003, 148, 1969–1986. [Google Scholar]
- Gutierrez, C. Strategies for geminivirus DNA replication and cell cycle interference. Physiol. Mol. Plant Pathol. 2002, 60, 219–230. [Google Scholar] [CrossRef]
- Kheyr-Pour, A.; Bananej, K.; Dafalla, G.A.; Caciagli, P.; Noris, E.; Ahoonmanesh, A.; Lecoq, H.; Gronenborn, B. Watermelon chlorotic stunt virus from Sudan and Iran: Sequence comparisons and identification of a whitefly-transmission determinant. Phytopathology 2000, 90, 629–635. [Google Scholar] [CrossRef]
- Laufs, J.; Jupin, I.; David, C.; Schumacher, S.; Heyraud-Nitschke, F.B.G. Geminivirus replication: Genetic and biochemical characterization of Rep protein function, a review. Biochimie 1995, 77, 765–773. [Google Scholar] [CrossRef]
- Orozco, B.M.; Hanley-Bowdoin, L. Conserved sequence and structural motifs contribute to the DNA binding and cleavage activities of a geminivirus replication protein. J. Biol. Chem. 1998, 273, 24448–24456. [Google Scholar]
- Harrison, B.D.; Robinson, D.J. Green shoots of geminivirology. Physiol. Mol. Plant Pathol. 2002, 60, 215–218. [Google Scholar] [CrossRef]
- Varma, A.; Malathi, V.G. Emerging geminivirus problems: A serious threat to crop production. Ann. Appl. Biol. 2003, 142, 145–164. [Google Scholar] [CrossRef]
- Jones, P.; Sattar, M.H.A.; Al Kaff, N. The incidence of virus disease in watermelon and sweetmelon crops in the Peoples Democratic Republic of Yemen and its impact on cropping policy. Ann. Appl. Biol. 1988, 17, 203–207. [Google Scholar]
- Al-Musa, A.; Anfoka, G.; Al-Abdulat, A.; Misbeh, S.; Haj Ahmed, F.; Otri, I. Watermelon chlorotic stunt virus (WmCSV): A serious disease threatening watermelon production in Jordan. Virus Genes 2011, 43, 79–89. [Google Scholar] [CrossRef]
- Bananej, K.; Dafalla, G.A.; Ahoonmanesh, A.; Kheyr-Pour, A. Host range of an Iranian isolate of Watermelon chlorotic stunt virus as determined by whitefly-mediated inoculation and agroinfection, and its geographical distribution. J. Phytopath. 2002, 150, 423–430. [Google Scholar]
- Haible, D.; Kober, S.; Jeske, H. Rolling circle amplification revolutionizes diagnosis and genomics of geminiviruses. J. Virol. Methods 2006, 135, 9–16. [Google Scholar] [CrossRef]
- Fauquet, C.; Briddon, R.; Brown, J.; Moriones, E.; Stanley, J.; Zerbini, M.; Zhou, X. Geminivirus strain demarcation and nomenclature. Arch. Virol. 2008, 153, 783–821. [Google Scholar]
- Raghavan, V.; Malik, P.S.; Choudhury, N.R.; Mukherjee, S.K. The DNA-A component of a plant geminivirus (Indian mung bean yellow mosaic virus) replicates in budding yeast cells. J. Virol. 2004, 78, 2405–2413. [Google Scholar]
- Laufs, J.; Schumacher, S.; Geisler, N.; Jupin, I.; Bronenborn, B. Identification of the nicking tyrosine of geminivirus Rep protein. FEBS Lett. 1995, 377, 258–262. [Google Scholar] [CrossRef]
- Stanley, J. Analysis of African cassava mosaic virus recombinants suggests strand nicking occurs within the conserved nonanucleotide motif during the initiation of rolling circle DNA replication. Virology 1995, 206, 707–712. [Google Scholar] [CrossRef]
- Settlage, S.B.; See, R.G.; Hanley-Bowdoin, L. Geminivirus C3 protein: Replication enhancement and protein interactions. J. Virol. 2005, 79, 9885–9895. [Google Scholar]
- Sunter, G.; Bisaro, D.M. Identification of a minimal sequence required for activation of the Tomato golden mosaic virus coat protein promoter in protoplasts. Virology 2003, 305, 452–462. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Settlage, S.B.; Orozco, B.M.; Nagar, S.; Robertson, D. Geminviruses: Models for plant DNA replication, transcription, and cell cycle regulation. Crit. Rev. Plant Sci. 1999, 18, 71–106. [Google Scholar] [CrossRef]
- Sanderfoot, A.A.; Ingham, D.J.; Lazarowitz, S.C. A viral movement protein as a nuclear shuttle. Plant Physiol. 1996, 11, 22–33. [Google Scholar]
- Ward, B.M.; Medville, R.; Lazarowitz, S.G.; Turgeon, R. The geminivirus BL1 movement protein is associated with edoplasmic reticulum-derived tubules in developing phloem cells. J. Virol. 1997, 71, 3726–3733. [Google Scholar]
- Abudy, A.; Sufrin-Ringwald, T.; Dayan-Glick, C.; Guenoune-Gelbart, D.; Livneh, O.; Zaccai, M.; Lapidot, M. Watermelon chlorotic stunt and Squash leaf curl begomoviruses—new threats to cucurbit crops in the Middle East. Isr. J. Plant Sci. 2010, 58, 33–42. [Google Scholar] [CrossRef]
- Argüello-Astorga, G.R.; Guevara-González, L.R.; Herrera-Estrella, L.R.; Rivera-Bustamante, R.F. Geminivirus replication origins have a group-specific organization of iterative elements: A model for replication. Virology 1994, 203, 90–100. [Google Scholar] [CrossRef]
- Chatterji, A.; Padidam, M.; Beachy, R.N.; Fauquet, C.M. Identification of replication specificity determinants in two strains of tomato leaf curl virus from New Delhi. J. Virol. 1999, 73, 5481–5489. [Google Scholar]
- Argüello-Astorga, G.R.; Ruiz. Medrano, R. An iteron-related domain is associated to motif 1 in the replication proteins of geminiviruses: Identification of potential interacting amino acid-base pairs by a comparartive approach. Arch. Virol. 2001, 146, 1465–1485. [Google Scholar] [CrossRef]
- Pennazio, S. Recovery. An enigmatic and neglected form of plant resistance to viruses. Riv. Biol. 2010, 103, 51–70. [Google Scholar]
- Khan, A.J.; Idris, A.M.; Al-Saady, N.A.; Al-Mahruki, M.S.; Al-Subhi, A.M.; Brown, J.K. A divergent isolate of Tomato yellow leaf curl virus from Oman with an associated DNA beta satellite: An evolutionary link between Asian and the Middle Eastern virus-satellite complexes. Virus Genes 2008, 36, 169–176. [Google Scholar] [CrossRef]
- Idris, A.M.; Shahid, M.S.; Briddon, R.W.; Khan, A.J.; Zhu, J.K.; Brown, J.K. An unusual alphasatellite associated with monopartite begomoviruses attenuates symptoms and reduces betasatellite accumulation. J. Gen. Virol. 2011, 92, 706–717. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. FOCUS 1990, 12, 13–15. [Google Scholar]
- Fire, A.; Xu, S.Q. Rolling replication of short DNA circles. Proc. Natl. Acad. Sci. USA 1995, 92, 4641–4645. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal_X windows interface; flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Page, R.D.M. Treeview: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Khan, A.J.; Akhtar, S.; Briddon, R.W.; Ammara, U.; Al-Matrooshi, A.M.; Mansoor, S. Complete Nucleotide Sequence of Watermelon Chlorotic Stunt Virus Originating from Oman. Viruses 2012, 4, 1169-1181. https://doi.org/10.3390/v4071169
Khan AJ, Akhtar S, Briddon RW, Ammara U, Al-Matrooshi AM, Mansoor S. Complete Nucleotide Sequence of Watermelon Chlorotic Stunt Virus Originating from Oman. Viruses. 2012; 4(7):1169-1181. https://doi.org/10.3390/v4071169
Chicago/Turabian StyleKhan, Akhtar J., Sohail Akhtar, Rob W. Briddon, Um Ammara, Abdulrahman M. Al-Matrooshi, and Shahid Mansoor. 2012. "Complete Nucleotide Sequence of Watermelon Chlorotic Stunt Virus Originating from Oman" Viruses 4, no. 7: 1169-1181. https://doi.org/10.3390/v4071169
APA StyleKhan, A. J., Akhtar, S., Briddon, R. W., Ammara, U., Al-Matrooshi, A. M., & Mansoor, S. (2012). Complete Nucleotide Sequence of Watermelon Chlorotic Stunt Virus Originating from Oman. Viruses, 4(7), 1169-1181. https://doi.org/10.3390/v4071169