Spread of the Emerging Viral Hemorrhagic Septicemia Virus Strain, Genotype IVb, in Michigan, USA

Abstract
:1. Introduction
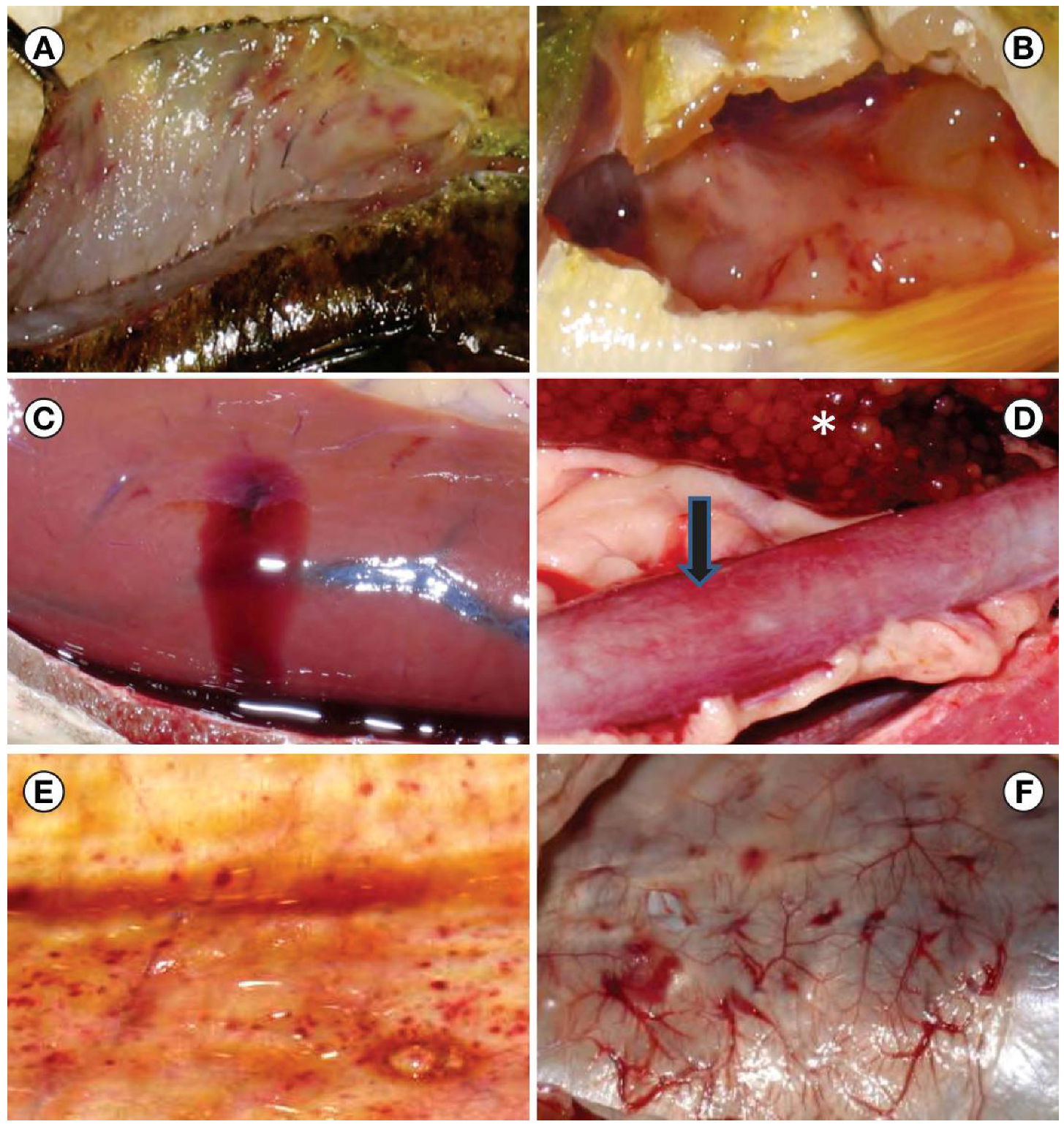
2. Results and Discussion
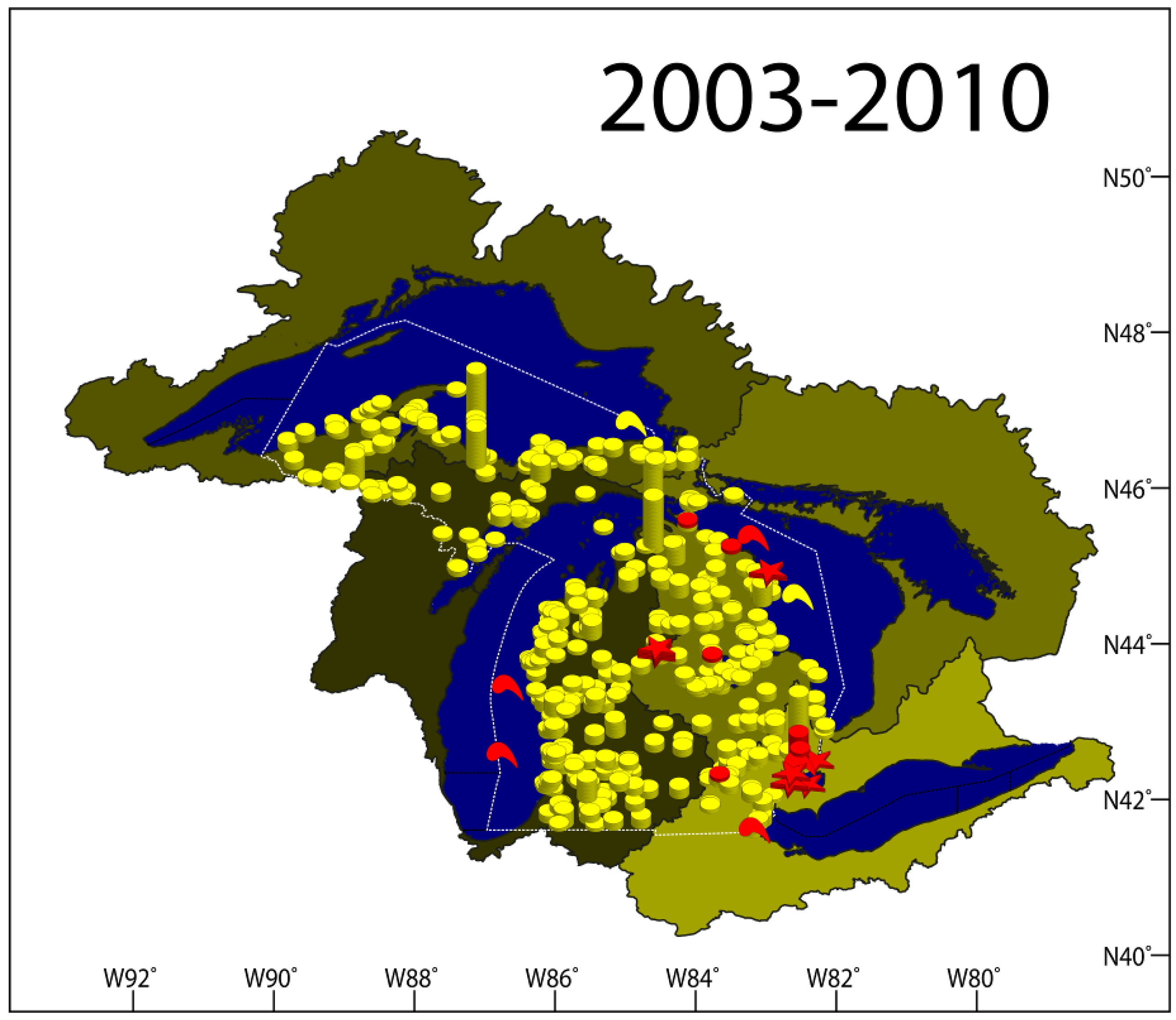
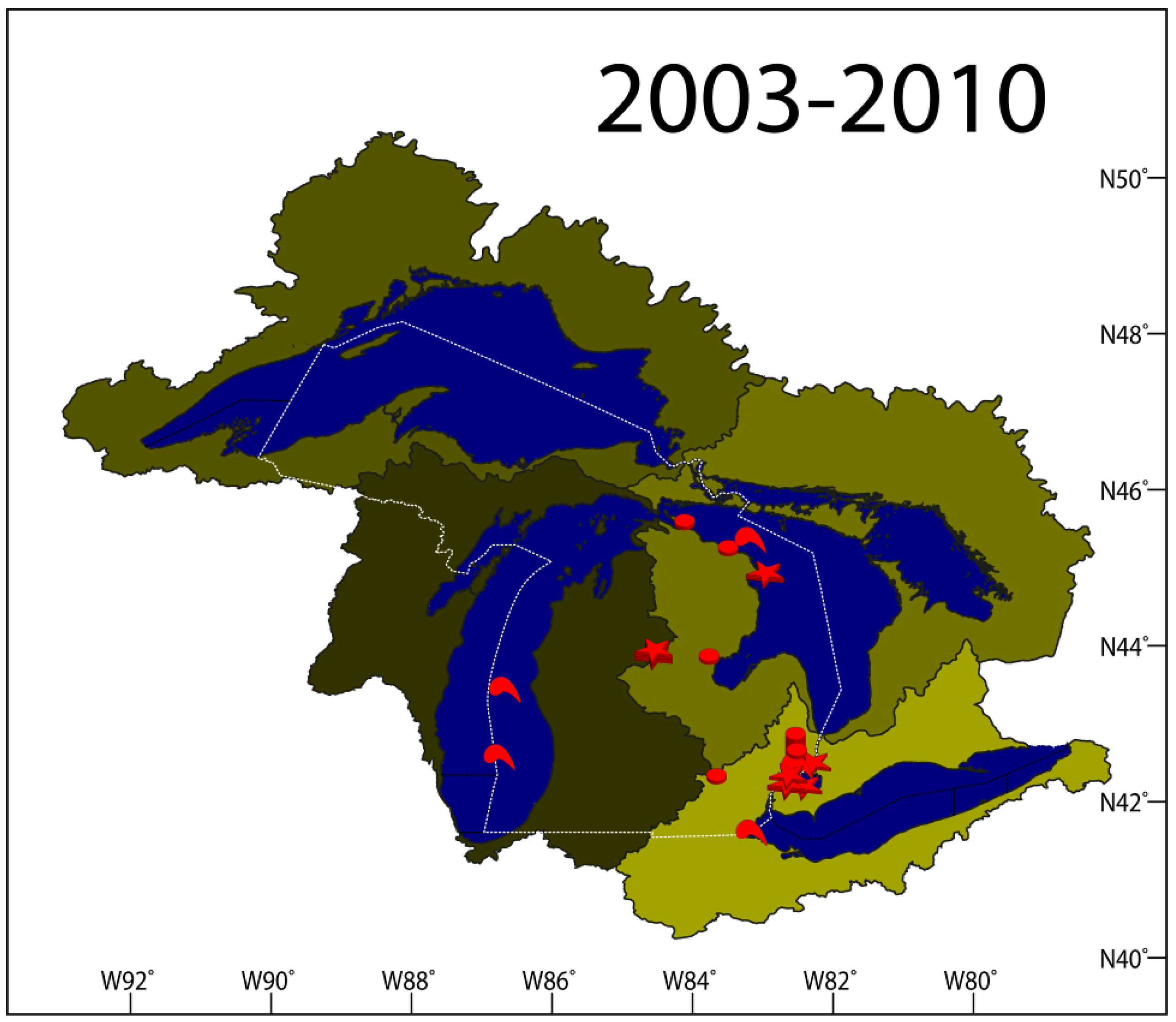
2.1. VHSV Spread in Michigan 2003–2010
2.1.1. 2003–2005
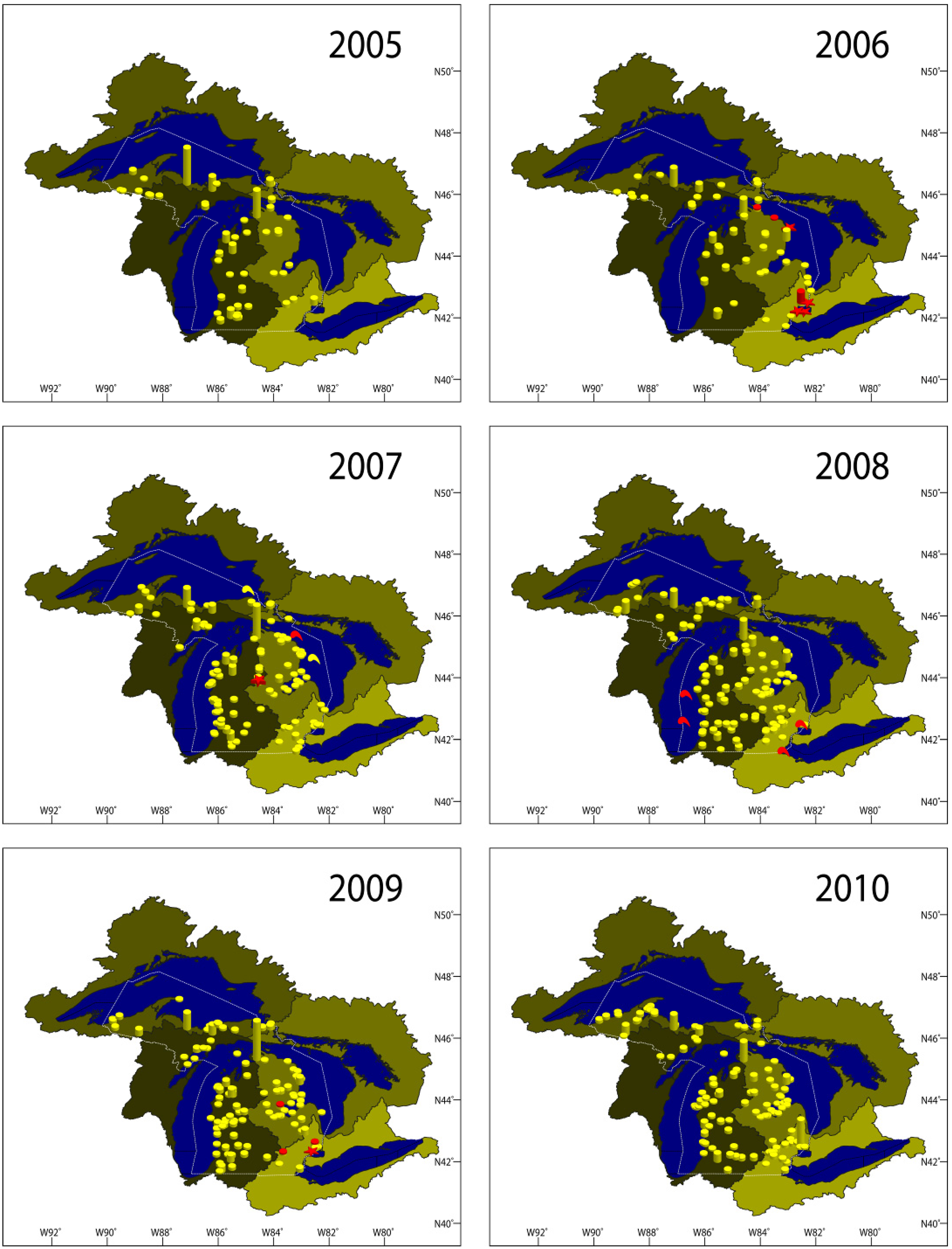
2.1.2. 2006

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date of sampling | Location | Wild species collected | Purpose of investigation | Latitude | Longitude |
|---|---|---|---|---|---|
| 3/14/06 | Lake St. Clair | gizzard shad | mortalities | 42.631389 | −82.5375 |
| 4/26/06 | Lake St. Clair | muskellunge | mortalities | 42.343 | −82.902 |
| yellow perch | mortalities | 42.343 | −82.902 | ||
| 5/17/06 | Lake St. Clair | northern pike | mortalities | 42.632 | −82.776 |
| muskellunge | mortalities | 42.632 | −82.776 | ||
| shorthead redhorse | mortalities | 42.632 | −82.776 | ||
| freshwater drum | mortalities | 42.632 | −82.776 | ||
| rockbass | mortalities | 42.632 | −82.776 | ||
| silver redhorse | mortalities | 42.632 | −82.776 | ||
| 5/22/06 | Lake St. Clair | freshwater drum | mortalities | 42.632 | −82.776 |
| rockbass | mortalities | 42.632 | −82.776 | ||
| northern pike | mortalities | 42.632 | −82.776 | ||
| muskellunge | mortalities | 42.632 | −82.776 | ||
| 5/24/06 | Lake St. Clair | freshwater drum | mortalities | 42.632 | −82.776 |
| muskellunge | mortalities | 42.632 | −82.776 | ||
| 7/10/06 | Lake St. Clair | muskellunge | mortalities | 42.333333 | −82.666667 |
| 8/18/06 | Lake Huron, Cheboygan | lake whitefish | research | 45.694798* | −84.355815 |
| 9/28/06 | Lake Huron, Swan River Weir | feral spawning chinook salmon | inspection | 45.402803 | −83.734867 |
| 10/6/06 | Lake Huron, Thunder Bay | lake whitefish | mortalities | 45.05 | −83.2 |
| walleye | mortalities | 45.05 | −83.2 | ||
| 12/13/06 | Lake St. Clair | spottail shiner | surveillance | 42.632* | −82.776 |
| emerald shiner | surveillance | 42.632* | −82.776 | ||
| 5/2/07 | Budd Lake, Clare County | black crappie | mortalities | 44.01585 | −84.788083 |
| bluegill | mortalities | 44.01585 | −84.788083 | ||
| pumpkinseed | mortalities | 44.01585 | −84.788083 | ||
| largemouth bass | mortalities | 44.01585 | −84.788083 | ||
| 5/11/09 | Baseline Lake, Livingston County | brown bullhead | surveillance | 42.42724 | −83.89926 |
| 5/18/09 | Lake St. Clair | spawning muskellunge | inspection | 42.6157* | −82.757 |
| 6/4/09 | Lake St. Clair | smallmouth bass | mortalities | 42.475507 | −82.879249 |
| 12/16/09 | Bait Facility, Bay County | spottail shiner | inspection | 43.932527 | −84.001776 |


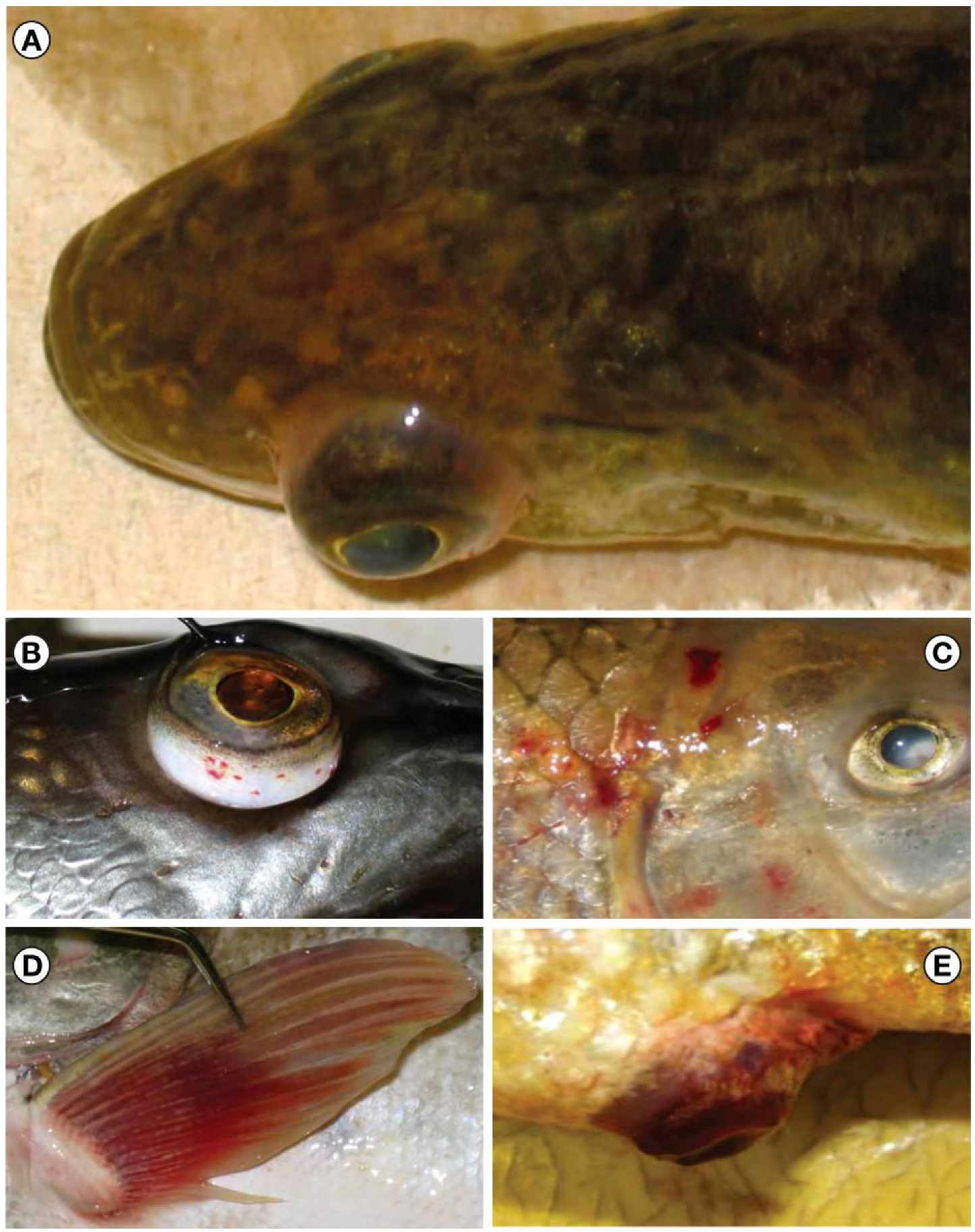
2.1.3. 2007
2.1.4. 2008
2.1.5. 2009
2.1.6. 2010
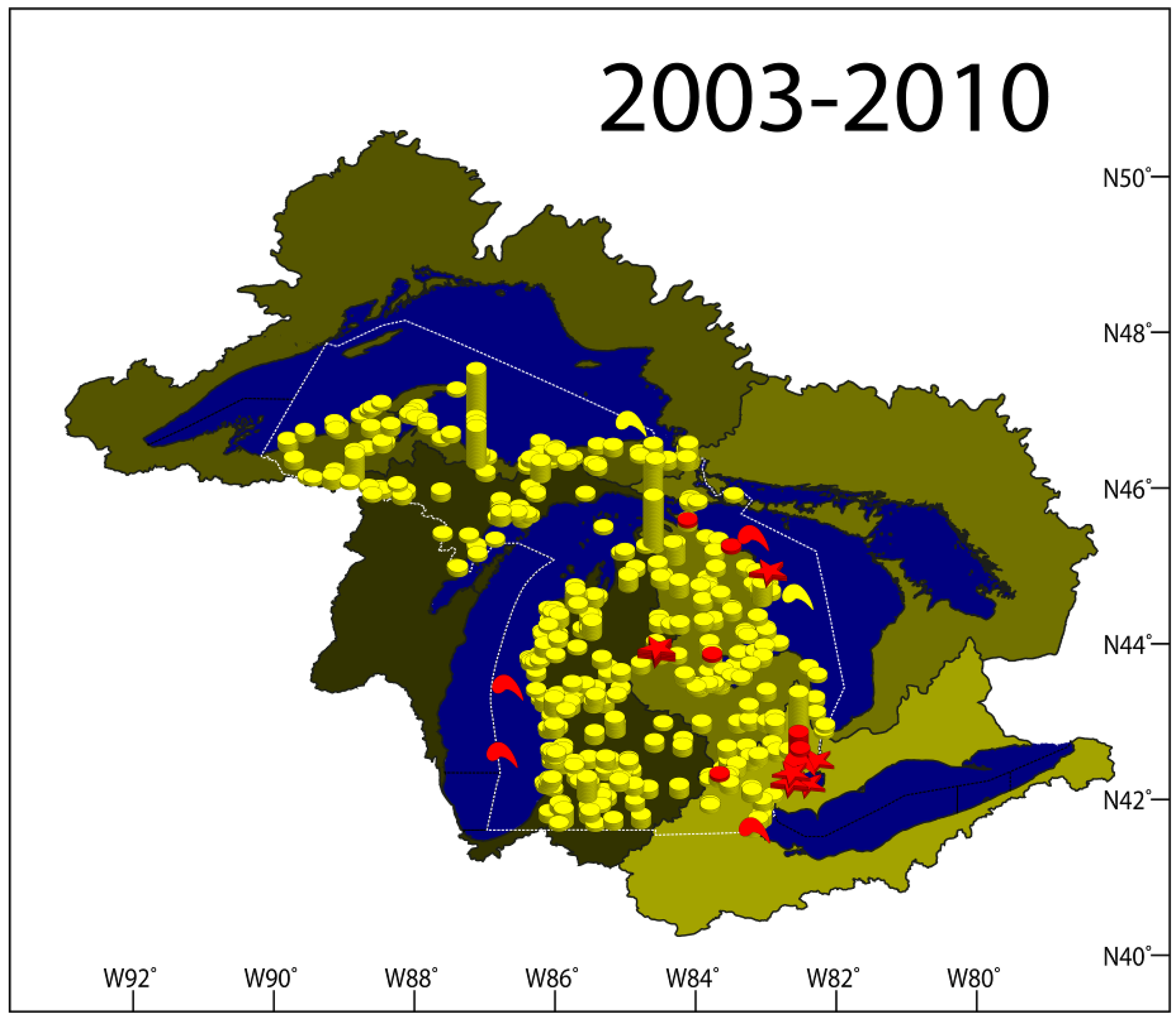
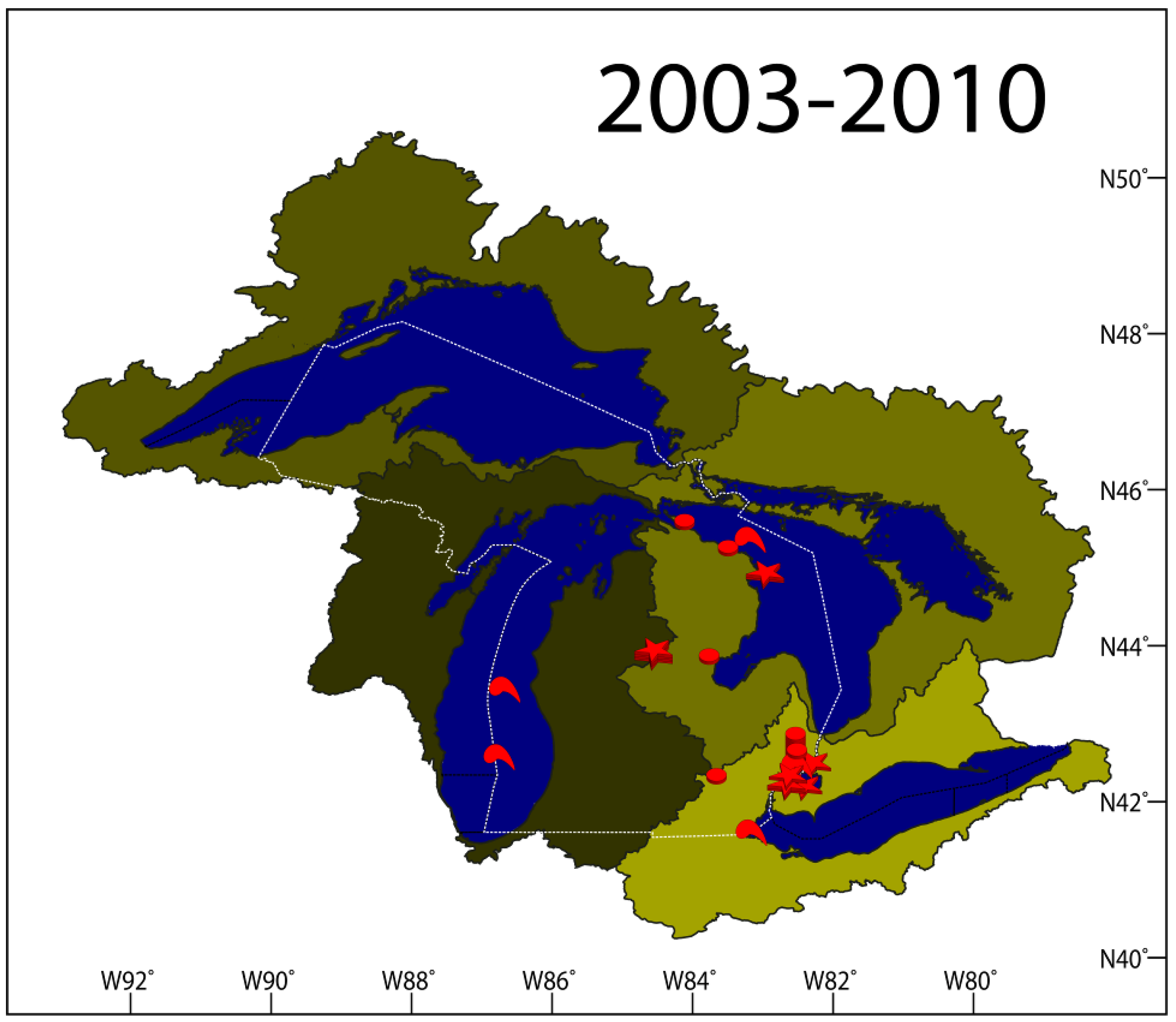
2.1.7. Results Summary 2003–2010
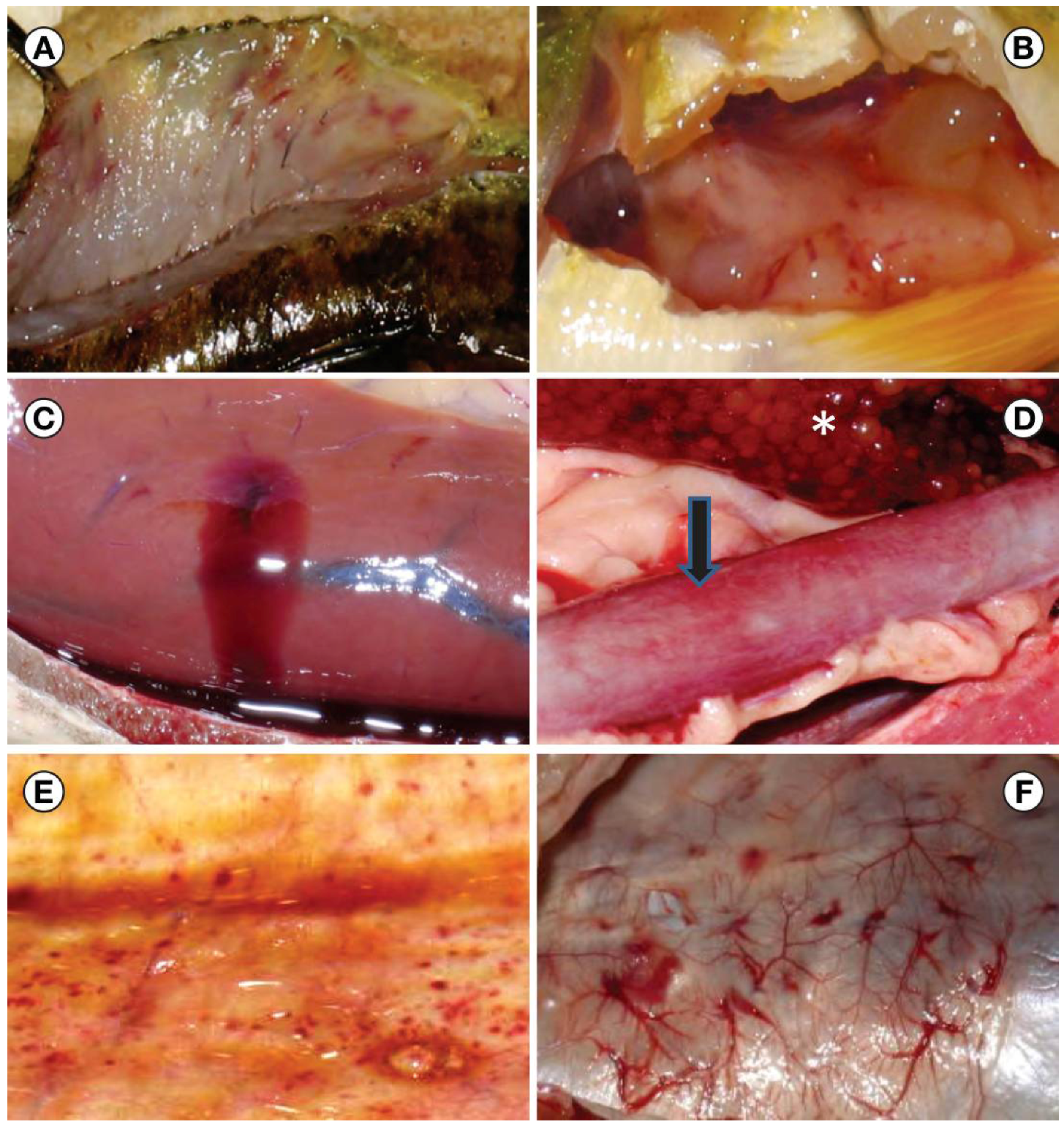
2.2. State and Federal Managerial Response to VHSV Emergence
2.3. Research Efforts Made to Better Understand VHSV-IVb Biology and Ecology
3. Materials and Methods
3.1. Fish and Sample Processing
3.2. Cell Lines
3.3. Virus Isolation
3.4. Virus Identification and PCR Confirmation
4. Conclusions
Acknowledgements
Supplementary Files
References and Notes
- Office International des Epizooties, Manual of Diagnostic Tests for Aquatic Animals, 6th ed; World Animal Health Organization: Paris, France, 2009.
- Schäperclaus, W. Die Immunisierung von Karpfen gegen Bauchwasserssucht auf natürilchen und künstlichen Wege. Fischereig Neudamm 1938, 5, 193–196. [Google Scholar]
- Hopper, K. The isolation of VHSV from Chinook salmon at Glenwood Springs, Orcas Islands, Washington. Fish Health Sect. Am. Fish. Soc. Newsl. 1989, 17, 1–2. [Google Scholar]
- Eaton, W.D.; Hulett, J. The first isolation in North America of Infectious Hematopoietic Necrosis Virus (IHNV) and Viral Hemorrhagic Septicemia Virus (VHSV) in Coho salmon from the same watershed. J. Aquat. Anim. Health 1991, 3, 114–117. [Google Scholar]
- Meyers, T.R.; Sullivan, J.; Emmenegger, E.; Follett, J.; Short, S.; Batts, W.N. Identification of viral hemorrhagic septicemia virus isolated from Pacific cod Gadus macrocephalus in Prince William Sound, Alaska, USA. Dis. Aquat. Org. 1992, 12, 167–175. [Google Scholar] [CrossRef]
- Meyers, T.R.; Short, S.; Lipson, K. Isolation of the North American strain of Viral Hemorrhagic Septicemia Virus (VHSV) associated with epizootic mortality in two new host species of Alaskan marine fish. Dis. Aquat. Org. 1999, 38, 81–86. [Google Scholar]
- Hedrick, R.P.; Batts, W.N.; Yun, S.; Traxler, G.S.; Kaufman, J.; Winton, J.R. Host and geographic range extensions of the North American strain of viral hemorrhagic septicemia virus. Dis. Aquat. Org. 2003, 55, 211–220. [Google Scholar] [CrossRef]
- Schlotfeldt, H.J.; Ahne, W.; Jørgensen, P.E.V.; Glende, W. Occurrence of viral hemorrhagic septicaemia in turbot (Scophthalmus maximus)—A natural outbreak. Bull. Eur. Assoc. Fish Pathol. 1991, 11, 105–107. [Google Scholar]
- Ross, K.; McCarthy, U.; Huntly, P.J.; Wood, B.P.; Stuart, D.; Rough, E.I. An outbreak of Viral Hemorrhagic Septicaemia (VHS) in turbot (Scophthalmus maximus) in Scotland. Bull.Eur. Assoc. Fish Pathol. 1994, 14, 213–214. [Google Scholar]
- Dixon, P.; Feist, S.; Kehoe, E.; Parry, L.; Stone, D.; Way, K. Isolation of viral haemorrhagic septicaemia virus from Atlantic herring Clupea harengus from the English Channel. Dis. Aquat. Org. 1997, 30, 81–89. [Google Scholar] [CrossRef]
- Mortensen, H.F.; Heuer, O.E.; Lorenzen, N.; Otte, L.; Olesen, N.J. Isolation of viral haemorrhagic septicaemia virus (VHSV) from wild marine fish species in the Baltic Sea, Kattegat, Skagerrak and the North Sea. Virus Res. 1999, 63, 95–106. [Google Scholar] [CrossRef]
- Takano, R.; Nishizawa, T.; Arimoto, M.; Muroga, K. Isolation of Viral Haemorrhagic Septicaemia Virus (VHSV) from wild Japanese flounder, Paralichthys olivaceus. Bull. Eur. Assoc. Fish Pathol. 2000, 20, 186–192. [Google Scholar]
- Kim, S.M.; Lee, J.I.; Hong, M.J.; Park, H.S.; Park, S.I. Genetic relationship of the VHSV (Viral Hemorrhagic Septicemia Virus) isolated from cultured olive flounder, Paralichthys olivaceus in Korea. J. Fish Pathol. 2003, 16, 1–12. [Google Scholar]
- Wolf, K. Hemorrhagic septicemia virus. In Fish Viruses and Fish Viral Diseases; Wolf, K., Ed.; Cornell University Press: Ithaca, NY, USA, 1988; pp. 217–248. [Google Scholar]
- De Kinkelin, P.; Chilmonczyk, S.; Dorson, M.; Le Berre, M.; Baudaouy, A. Some pathogenic facets of rhabdoviral infection of salmonid fish. In Proceedings of the 4th Munich Symposium on Microbiology: Mechanisms of Viral Pathogenesis and Virulence; Bachmann, P., Ed.; World Health Organization (WHO): Munich, Germany, 1979; pp. 357–375. [Google Scholar]
- Evensen, O.; Meier, W.; Wahli, T.; Olesen, N.; Jørgensen, P.; Hastein, T. Comparison of immunohistochemistry and virus cultivation for detection of viral haemorrhagic septicaemia virus in experimentally infected rainbow trout Oncorhynchus mykiss. Dis. Aquat. Org. 1994, 20, 101–109. [Google Scholar] [CrossRef]
- VHSV Expert Panel and Working Group. Viral hemorrhagic septicemia virus (VHSV IVb) risk factors and association measures derived by expert panel. Prev. Vet. Med. 2010, 94, 128–139. [CrossRef]
- Yasutake, W.T.; Rasmussen, C.J. Histopathogenesis of experimentally induced viral hemorrhagic septicemia in fingerling rainbow trout (Salmo gairdneri). Bull. Off. Int. Epizoot. 1968, 69, 977–984. [Google Scholar]
- Brudeseth, B.E.; Castric, J.; Evensen, O. Studies on pathogenesis following single and double infection with viral hemorrhagic septicemia virus and infectious hematopoietic necrosis virus in rainbow trout (Oncorhynchus mykiss). Vet. Pathol. 2002, 39, 180–189. [Google Scholar] [CrossRef]
- Kim, R.; Faisal, M. Experimental studies confirm the wide host range of the Great Lakes viral haemorrhagic septicaemia virus genotype IVb. J. Fish Dis. 2010, 33, 83–88. [Google Scholar]
- Kim, R.; Faisal, M. Comparative susceptibility of representative Great Lakes fish species to the North American viral hemorrhagic septicemia virus sublineage IVb. Dis. Aquat. Org. 2010, 91, 23–34. [Google Scholar]
- Kim, R.K.; Faisal, M. The Laurentian Great Lakes strain (MI03) of the viral hemorrhagic septicemia virus is highly pathogenic for juvenile muskellunge, Esox masquinongy (Mitchill). J. Fish Dis. 2010, 33, 513–527. [Google Scholar] [CrossRef]
- Kim, R.; Faisal, M. Emergence and resurgence of the viral hemorrhagic septicemia virus (Novirhabdovirus, Rhabdoviridae, Mononegavirales). J. Adv. Res. 2011, 2, 9–23. [Google Scholar] [CrossRef]
- Kim, R.; Faisal, M. Viral hemorrhagic septicemia virus in the Laurentian Great Lakes. In Bridging America and Russia with Shared Perspectives on Aquatic Animal Health. In Proceedings of the Third Bilateral Conference between Russia and the United States, July 2009; Cipriano, R.C., Bruckner, A.W., Shchelkunov, I.S., Eds.; The Khaled bin Sultan Living Oceans Foundation, Landover, MD, USA, Michigan State University Printing: East Lansing, MI, USA, 2011; pp. 113–122. [Google Scholar]
- Al-Hussinee, L.; Lumsden, J.S.; Lord, S.; Stevenson, R.M.W.; Casey, R.N.; Groocock, G.H.; Britt, K.L.; Kohler, K.H.; Wooster, G.A.; Getchell, R.G.; Bowser, P.R. Immunohistochemistry and pathology of viral hemorrhagic septicemia virus, type IVb associated with mortality in multiple Great Lakes fish. Dis. Aquat. Org. 2011, 93, 117–127. [Google Scholar] [CrossRef]
- Schütze, H.; Mundt, E.; Mettenleiter, T. Complete genomic sequence of viral hemorrhagic septicemia virus, a fish rhabdovirus. Virus Genes 1999, 19, 59–65. [Google Scholar]
- Ammayappan, A.; Vakharia, V.N. Molecular characterization of the Great Lakes viral hemorrhagic septicemia virus (VHSV) isolate from USA. Virol. J. 2009, 6, 171. [Google Scholar]
- Wagner, R.R.; Rose, J.K. Rhabdoviridae: The Viruses and Their Replication; Fields, B.N., Knipe, D.M., Howley, P.M., Eds.; Lippincott-Raven: Philadelphia/New York, PA/NY, USA, 1996. [Google Scholar]
- Emerson, S.U.; Yu, Y. Both NS and L proteins are required for in vitro RNA synthesis by vesicular stomatitis virus. J. Virol. 1975, 15, 1348–1356. [Google Scholar]
- Ivanov, I.; Yabukarski, F.; Ruigrok, R.W.; Jamin, M. Structural insights into the rhabdovirus transcription/replication complex. Virus Res. 2011, 162, 126–137. [Google Scholar]
- Finke, S.; Conzelmann, K.-K. Dissociation of rabies virus matrix protein functions in regulation of viral RNA synthesis and virus assembly. J. Virol. 2003, 77, 12074–12082. [Google Scholar]
- Jørgensen, P.E.V.; Einer-Jensen, K.; Higman, K.H.; Winton, J.R. Sequence comparison of the central region of the glycoprotein gene of neutralizable, non-neutralizable, and serially passed isolates of viral haemorrhagic septicaemia virus. Dis. Aquat. Org. 1995, 23, 77–82. [Google Scholar]
- Ammayappan, A.; Kurath, G.; Thompson, T.M.; Vakharia, V.N. A reverse genetics system for the Great Lakes strain of viral hemorrhagic septicemia virus: The NV gene is required for pathogenicity. Mar. Biotechnol. 2011, 13, 672–683. [Google Scholar]
- Einer-Jensen, K.; Ahrens, P.; Forsberg, R.; Lorenzen, N. Evolution of the fish rhabdovirus viral haemorrhagic septicaemia virus. J. Gen. Virol. 2004, 85, 1167–1179. [Google Scholar]
- Snow, M.; Bain, N.; Black, J.; Taupin, V.; Cunningham, C.; King, J.; Skall, H.; Raynard, R. Genetic population structure of marine viral haemorrhagic septicaemia virus (VHSV). Dis. Aquat. Org. 2004, 61, 11–21. [Google Scholar]
- Stone, D.M.; Way, K.; Dixon, P.F. Nucleotide sequence of the glycoprotein gene of viral haemorrhagic septicaemia (VHS) viruses from different geographical areas: a link between VHS in farmed fish species and viruses isolated from North Sea cod (Gadus morhua L.). J. Gen. Virol. 1997, 78, 1319–1326. [Google Scholar]
- Nishizawa, T.; Iida, H.; Takano, R.; Isshiki, T.; Nakajima, K.; Muroga, K. Genetic relatedness among Japanese, American and European isolates of viral hemorrhagic septicemia virus (VHSV) based on partial G and P genes. Dis. Aquat. Org. 2002, 48, 143–148. [Google Scholar]
- Elsayed, E.; Faisal, M.; Thomas, M.; Whelan, G.; Batts, W.; Winton, J. Isolation of viral haemorrhagic septicaemia virus from muskellunge, Esox masquinongy (Mitchill), in Lake St. Clair, Michigan, USA reveals a new sublineage of the North American genotype. J. Fish Dis. 2006, 29, 611–619. [Google Scholar] [CrossRef]
- Lumsden, J.S.; Morrison, B.; Yason, C.; Russell, S.; Young, K.; Yazdanpanah, A.; Huber, P.; Al-Hussinee, L.; Stone, D.; Way, K. Mortality event in freshwater drum Aplodinotus grunniens from Lake Ontario, Canada, associated with viral hemorrhagic septicemia virus, type IV. Dis. Aquat. Org. 2007, 76, 99–111. [Google Scholar] [CrossRef]
- Groocock, G.H.; Getchell, R.G.; Wooster, G.A.; Britt, K.L.; Batts, W.N.; Winton, J.R.; Casey, R.N.; Casey, J.W.; Bowser, P.R. Detection of viral hemorrhagic septicemia in round gobies in New York State (USA) waters of Lake Ontario and the St. Lawrence River. Dis. Aquat. Org. 2007, 76, 187–92. [Google Scholar]
- Gagné, N.; MacKinnon, A-M.; Boston, L.; Souter, B.; Cook-Versloot, M.; Griffiths, S.; Olivier, G. Isolation of viral haemorrhagic septicaemia virus from mummichog, stickleback, striped bass and brown trout in eastern Canada. J. Fish Dis. 2007, 30, 213–223. [Google Scholar] [CrossRef]
- Thompson, T.M.; Batts, W.N.; Faisal, M.; Bowser, P.; Casey, J.W.; Phillips, K.; Garver, K.A.; Winton, J.; Kurath, G. Emergence of Viral hemorrhagic septicemia virus in the North American Great Lakes region is associated with low viral genetic diversity. Dis. Aquat. Org. 2011, 96, 29–43. [Google Scholar]
- Frattini, S.A.; Groocock, G.H.; Getchell, R.G.; Wooster, G.A.; Casey, R.N.; Casey, J.W.; Bowser, P.R. A 2006 survey of Viral Hemorrhagic Septicemia (VHSV) virus type IVb in New York State waters. J. Great Lakes Res. 2011, 37, 194–198. [Google Scholar]
- Kane-Sutton, M.; Kinter, B.; Dennis, P.M.; Koonce, J.F. Viral hemorrhagic septicemia virus infection in yellow perch, Perca flavescens, in Lake Erie. J. Great Lakes Res. 2010, 36, 37–43. [Google Scholar] [CrossRef]
- Faisal, M.; Winters, A.D. Detection of Viral Hemorrhagic Septicemia Virus (VHSV) from Diporeia spp. (Pontoporeiidae, Amphipoda) in the Laurentian Great Lakes, USA. Parasit. Vectors 2011, 4, 2. [Google Scholar] [CrossRef]
- Faisal, M.; Schulz, C.A. Detection of Viral Hemorrhagic Septicemia virus (VHSV) from the leech Myzobdella lugubris Leidy, 1851. Parasit. Vectors 2009, 2, 45. [Google Scholar] [CrossRef]
- Tuttle-Lau, M.T.; Phillips, K.A.; Gaikowski, M.P. Evaluation of the efficacy of iodophor disinfection of walleye and northern pike eggs to eliminate viral hemorrhagic septicemia virus. U.S. Geological Survey Fact Sheet 2009-3107, 4p.
- USDA-APHIS. VHS Federal Order. United States Department of Agriculture (USDA), Animal and Plant Health Inspection Service (APHIS): Ames, IA, USA, 2008. Available online: http://www.aphis.usda.gov/animal_health/animal_dis_spec/aquaculture (available online 16 April 2012).
- Kim, R.; Faisal, M. Shedding of viral hemorrhagic septicemia virus (Genotype IVb) by experimentally infected muskellunge (Esox masquinongy). J. Microbiol. 2012, in press. [Google Scholar]
- Weeks, C.; Kim, R.; Wolgamood, M.; Whelan, G.; Faisal, M. Experimental infection studies demonstrate the high susceptibility of the salmonid, lake herring, Coregonus artedi (Le Sueur), to the Great Lakes strain of viral haemorrhagic septicaemia virus (genotype IVb). J. Fish Dis. 2011, 34, 887–891. [Google Scholar] [CrossRef]
- Faisal, M.; Schulz, C.; Eissa, A.; Whelan, G. High prevalence of buccal ulcerations in largemouth bass, Micropterus salmoides (Centrarchidae) from Michigan inland lakes associated with Myzobdella lugubris Leidy 1851 (Annelida: Hirudinea). Parasite 2011, 18, 79–84. [Google Scholar]
- Hope, K.M.; Casey, R.N.; Groocock, G.H.; Getchell, R.G.; Bowser, P.R.; Casey, J.W. Comparison of quantitative RT-PCR with cell culture to detect viral hemorrhagic septicemia virus (VHSV) IVb infections in the Great Lakes. J. Aquat. Anim. Health 2010, 22, 50–61. [Google Scholar]
- Bain, M.B.; Cornwell, E.R.; Hope, K.M.; Eckerlin, G.E.; Casey, R.N.; Groocock, G.H.; Getchell, R.G.; Bowser, P.R.; Winton, J.R.; Batts, W.N.; Cangelosi, A.; Casey, J.W. Distribution of an invasive aquatic pathogen (Viral Hemorrhagic Septicemia Virus) in the Great Lakes and its relationship to shipping. PLoS One 2010, 5, e10156. [Google Scholar]
- Garver, K.; Hawley, L.; McClure, C.; Schroeder, T.; Aldous, S.; Doig, F.; Snow, M.; Edes, S.; Baynes, C.; Richard, J. Development and validation of a reverse transcription quantitative PCR for universal detection of viral hemorrhagic septicemia virus. Dis. Aquat. Org. 2011, 95, 97–112. [Google Scholar]
- Millard, E.; Faisal, M. Development of neutralizing antibody responses in muskellunge, Esox masquinongy (Mitchill), experimentally exposed to viral haemorrhagic septicaemia virus (genotype IVb). J. Fish Dis. 2012, in press. [Google Scholar]
- Millard, E.; Faisal, M. Heterogeneity in levels of serum neutralizing antibodies against viral hemorrhagic septicemia virus (Genotype IVb) among fish species in Lake St. Clair, Michigan, USA. J. Wildl. Dis. 2012, in press. [Google Scholar]
- Eckerlin, G.E.; Farrell, J.M.; Casey, R.N.; Hope, K.M.; Groocock, G.H.; Bowser, P.R.; Casey, J. Temporal variation in prevalence of Viral Hemorrhagic Septicemia Virus Type IVb among Upper St. Lawrence River Smallmouth Bass. Trans. Am. Fish. Soc. 2011, 140, 529–536. [Google Scholar] [CrossRef]
- Smail, D.A. Viral haemorrhagic septicaemia. In Fish Diseases and Disorders. Volume 3: Viral, Bacterial and Fungal Infections; Woo, P.T.K., Bruno, D.W., Eds.; FRS Marine Laboratory: Aberdeen, UK, 1999; pp. 123–147. [Google Scholar]
- De Kinkelin, P.; Scherrer, R. Le Virus d'Egtved I. Stabilité, développement et structure du virus de la souche danoise F1. Ann. Rech. Vet. 1970, 1, 17–30. [Google Scholar]
- Meyers, T.; Winton, J. Viral hemorrhagic septicemia virus in North America. Annu. Rev. Fish Dis. 1995, 5, 3–24. [Google Scholar]
- Skall, H.F.; Olesen, N.J.; Mellergaard, S. Viral haemorrhagic septicaemia virus in marine fish and its implications for fish farming—A review. J. Fish Dis. 2005, 28, 509–529. [Google Scholar]
- Kocan, R.; Bradley, M.; Elder, N.; Meyers, T.; Batts, W.; Winton, J. North American strain of viral haemorrhagic septicemia virus is highly pathogenic for laboratory-reared Pacific herring. J. Aquat. Anim. Hlth. 1997, 9, 279–290. [Google Scholar]
- Ghittino, P. Viral hemorrhagic septicemia (VHS) in rainbow trout in Italy. Ann. N. Y. Acad. Sci. 1965, 126, 468–478. [Google Scholar] [CrossRef]
- USDA-APHIS. Species regulated by title 9 CFR Parts 83.1 through 83.7, 93.900 and 93.910 through 93.916. United States Department of Agriculture (USDA),Animal and Plant Health Inspection Service (APHIS): Ames, IA, USA, 2008. Available online: http://www.aphis.usda.gov/ animal_health/animal_dis_spec/aquaculture/downloads/vhs_regulated_spp.pdf (available online 16 April 2012).
- Fijan, N.; Sulimanovic, D.; Bearzotti, M.; Muzinic, D.; Zwillenberg, L.; Chilmonczyk, S.; Vautherot, J.; De kinkelin, P. Some properties of the Epithelioma papulosum cyprini (EPC) cell line from carp cyprinus carpio. Annal. Instit. Pasteur Virol. 1983, 134, 207–220. [Google Scholar]
- Gravell, M.; Malsberger, R.G. A permanent cell line from the fathead minnow (Pimephales promelas). Ann. N. Y. Acad. Sci. 1965, 126, 555–565. [Google Scholar]
- Fryer, J.L.; Yusha, A.; Pilcher, K.S. The in vitro cultivation of tissue and cells of Pacific salmon and steelhead trout. Ann. N. Y. Acad. Sci. 1965, 126, 566–586. [Google Scholar]
- Batts, W.N.; Winton, J.R. Viral hemorrhagic septicemia. In American Fisheries Society-Fish Health Section (AFS-FHS) Blue Book: Suggested Procedures for the Detection and Identification of Certain Finfish and Shellfish Pathogens; American Fisheries Society-Fish Health Section (AFS-FHS): Bethesda, MD, USA, 2007. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Faisal, M.; Shavalier, M.; Kim, R.K.; Millard, E.V.; Gunn, M.R.; Winters, A.D.; Schulz, C.A.; Eissa, A.; Thomas, M.V.; Wolgamood, M.; et al. Spread of the Emerging Viral Hemorrhagic Septicemia Virus Strain, Genotype IVb, in Michigan, USA. Viruses 2012, 4, 734-760. https://doi.org/10.3390/v4050734
Faisal M, Shavalier M, Kim RK, Millard EV, Gunn MR, Winters AD, Schulz CA, Eissa A, Thomas MV, Wolgamood M, et al. Spread of the Emerging Viral Hemorrhagic Septicemia Virus Strain, Genotype IVb, in Michigan, USA. Viruses. 2012; 4(5):734-760. https://doi.org/10.3390/v4050734
Chicago/Turabian StyleFaisal, Mohamed, Megan Shavalier, Robert K. Kim, Elena V. Millard, Michelle R. Gunn, Andrew D. Winters, Carolyn A. Schulz, Alaa Eissa, Michael V. Thomas, Martha Wolgamood, and et al. 2012. "Spread of the Emerging Viral Hemorrhagic Septicemia Virus Strain, Genotype IVb, in Michigan, USA" Viruses 4, no. 5: 734-760. https://doi.org/10.3390/v4050734