Viruses of Fish: An Overview of Significant Pathogens

Abstract
:1. Introduction
2. Aquabirnavirus
3. Betanodavirus
4. Infectious Salmon Anemia Virus
5. Salmon Alphavirus
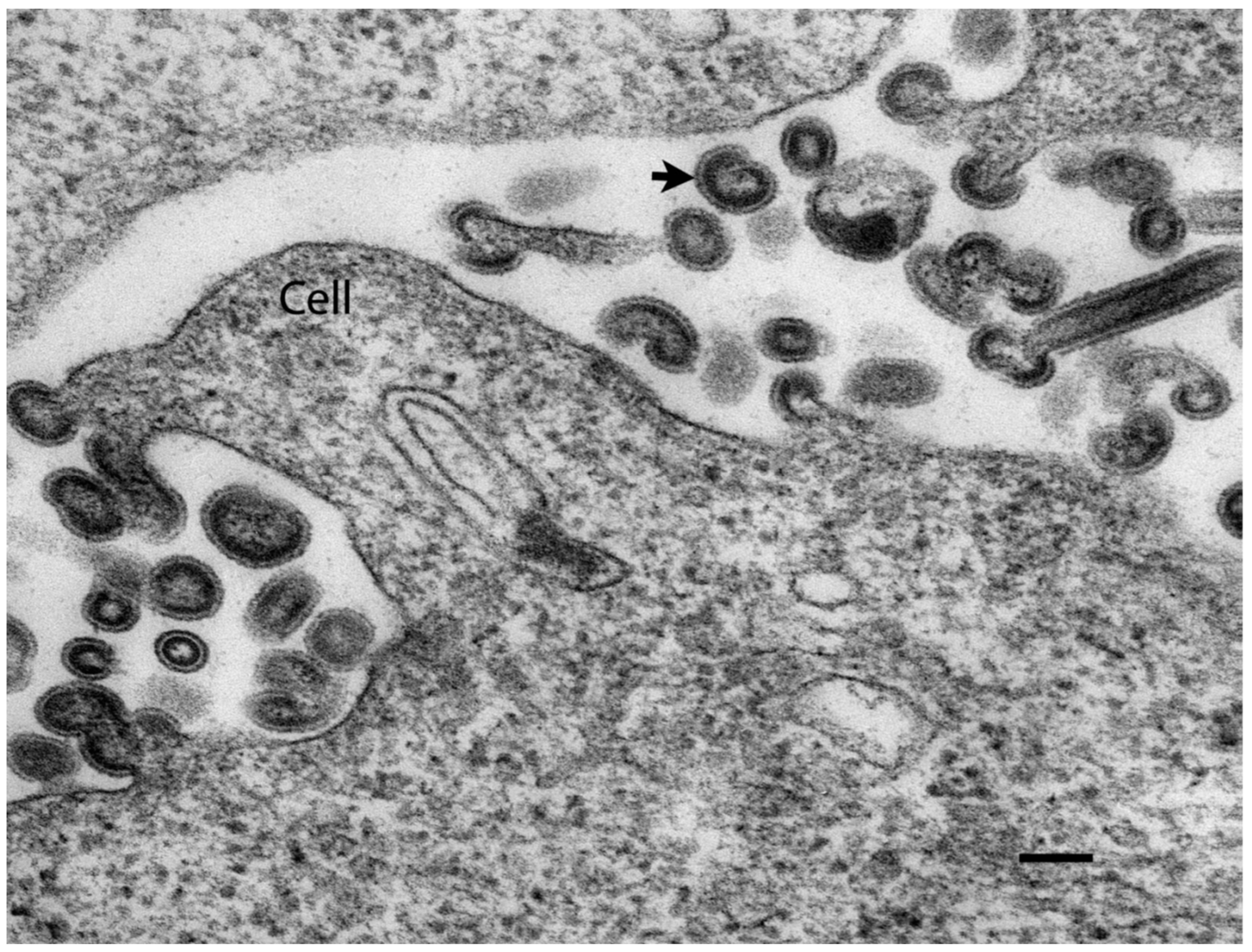
6. Infectious Hematopoietic Necrosis Virus
7. Epizootic Hematopoietic Necrosis Virus
8. Viral Hemorrhagic Septicemia Virus
Conflicts of Interest
References and Notes
- Wolf, K. Fish Viruses and Fish Viral Diseases; Cornell University Press: Ithaca, NY, USA, 1988; p. 476. [Google Scholar]
- Wolf, K.; Quimby, M.C. Established eurythermic line of fish cells in vitro. Science 1962, 135, 1065–1066. [Google Scholar] [CrossRef]
- Gravell, M.; Malsberger, R.G. A permanent cell line from Fathead Minnow (Pimephales promelas). Ann. NY Acad. Sci. 1965, 126, 555–565. [Google Scholar] [CrossRef] [PubMed]
- OIE. Manual of Diagnostic Tests for Aquatic Animals, 6th ed.; World Organisation for Animal Health: Paris, France, 2009; p. 383. [Google Scholar]
- Wolf, K.; Sniesko, S.F.; Dunbar, C.E.; Pyle, E. Virus nature of infectious pancreatic necrosis in trout. Proc. Soc. Exp. Med. Biol. 1960, 104, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Wood, E.M.; Snieszko, S.F.; Yasutake, W.T. Infectious pancreatic necrosis in brook trout. Am. Med. Assoc. Arch. Pathol. 1955, 60, 26–28. [Google Scholar]
- Hill, B.J.; Way, K. Serological classification of infectious pancreatic necrosis (IPN) virus and other aquatic birnaviruses. Ann. Rev. Fish Dis. 1995, 5, 55–77. [Google Scholar] [CrossRef]
- Reno, P.W. Infectious pancreatic necrosis and associated aquatic birnaviruses. In Fish Diseases and Disorders. Volume 3: Viral, Bacterial and Fungal Infections; Woo, P.T.K., Bruno, D.W., Eds.; CABI Publishing: Wallingford, UK, 1999; pp. 1–56. [Google Scholar]
- Sorimachi, M.; Hara, T. Characteristics and pathogenicity of a virus isolated from yellowtail fingerlings showing ascites. Fish Pathol. 1985, 19, 231–238. [Google Scholar] [CrossRef]
- Isshiki, T.; Nagano, T.; Kanehira, K.; Suzuki, S. Distribution of marine birnavirus in cultured marine fish species from Kagawa prefecture, Japan. J. Fish Dis. 2004, 27, 89–98. [Google Scholar] [CrossRef]
- Christie, K.E.; Håvarstein, L.S.; Djuvik, H.O.; Ness, S.; Endresen, C. Characterisation of a new serotype of infectious pancreatic necrosis virus isolated from Atlantic salmon. Arch. Virol. 1988, 103, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.J.; Pearson, M.D. Infectious pancreatic necrosis in Atlantic salmon, Salmo salar L. J. Fish Dis. 2005, 28, 383–390. [Google Scholar] [CrossRef]
- John, K.R.; Richards, R.H. Characteristics of a new birnavirus associated with a warm-water fish cell line. J. Gen. Virol. 1999, 80, 2061–2065. [Google Scholar] [CrossRef]
- Dixon, P.F.; Ngoh, G.H.; Stone, D.M.; Chang, S.F.; Way, K.; Kueh, S.L.F. Proposal for a fourth aquabirnavirus serogroup. Arch. Virol. 2008, 153, 1937–1941. [Google Scholar] [CrossRef]
- Zhang, C.X.; Suzuki, S. Aquabirnaviruses isolated from marine organisms form a distinct genogroup from othe aquabirnaviruses. J. Fish Dis. 2004, 27, 633–43. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, T.; Kinoshita, S.; Yoshimizu, M. An approach for genogrouping of Japanese isolates of aquabirnaviruses in a new genogroup, VII, based on the VP2/NS junction region. J. Gen. Virol. 2005, 86, 1973–1978. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.R.; McColl, K.A.; Wang, L.-F.; Yu, M.; Williams, L.M.; Crane, M.St.J. Molecular characterisation of Australasian isolates of aquabirnavirus. Dis. Aquat. Org. 2010, 93, 1–15. [Google Scholar] [CrossRef]
- Dobos, P. The molecular biology of infectious pancreatic necrosis virus (IPNV). Ann. Rev. Fish Dis. 1995, 5, 25–54. [Google Scholar] [CrossRef]
- Dobos, P.; Hill, B.J.; Hallet, R.; Kells, D.T.; Becht, H.; Teninges, D. Biophysical and biochemical characterisation of five animal viruses with bisegmented double-stranded RNA genomes. J. Virol. 1979, 32, 593–605. [Google Scholar] [CrossRef]
- Snieszko, S.F. Your fishes’ health: Diseases of fishes. Trop. Fish Hobbyist. 1981, 3, 78. [Google Scholar]
- Sano, M.; Okamoto, N.; Fukuda, H.; Saneyoshi, M.; Sano, T. 1992. Virulence of infectious pancreatic necrosis virus is associated with the larger RNA segment (RNA segment A). J. Fish Dis. 1998, 15, 283–293. [Google Scholar] [CrossRef]
- Santi, N.; Vakharia, V.N.; Evensen, Ø. Identification of putative motifs involved in the virulence of infectious pancreatic necrosis virus. Virology 2004, 322, 31–40. [Google Scholar] [CrossRef]
- Santi, N.; Sandtrø, A.; Sindre, H.; Song, H.; Hong, J.-R.; Thu, B.; Wu, J.-L.; Vakharia, V.N.; Evensen, Ø. Infectious pancreatic necrosis virus induces apoptosis in vitro and in vivo independent of VP5 expression. Virology 2005, 342, 13–25. [Google Scholar] [CrossRef]
- Bowden, T.J.; Smail, D.A.; Ellis, A.E. Development of a reproducible infectious pancreatic necrosis virus challenge model for Atlantic salmon, Salmo salar L. J. Fish Dis. 2002, 25, 555–563. [Google Scholar] [CrossRef]
- Ramstad, A.; Romstad, A.B.; Knappskog, D.H.; Midtlyng, P.J. Field validation of experimental challenge models for IPN vaccines. J. Fish Dis. 2007, 30, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Ramstad, A.; Midtlyng, P.J. Strong genetic influence on IPN vaccination-and-challenge trials in Atlantic salmon Salmo salar L. J. Fish Dis. 2008, 31, 567–578. [Google Scholar] [CrossRef]
- Evensen, ø.; Santi, N. Infectious pancreatic necrosis virus. In Encyclopaedia of Virology; Mahy, B.W.J., Van Regenmortel, M.H.V., Eds.; Elsevier: Oxford, UK, 2008; pp. 83–89. [Google Scholar]
- Ball, L.A.; Hendry, D.A.; Johnson, J.E.; Ruekert, R.R.; Scotti, P.D. Family Nodaviridae. In Virus Taxonomy Seventh Report of the International Committee on Taxonomy of Viruses; Regenmortel, M.H.V., Fauquet, C.M., Bishop, D.H.L., Carstens, E.B., Estes, M.K., Lemon, S.D.M., Maniloff, J., Mayo, M.A., McGeoch, D.J., Pringle, C.R., Wickner, R.B., Eds.; Academic Press: San Diego, CA, USA, 2000; pp. 747–755. [Google Scholar]
- Glazebrook, J.S.; Heasman, M.P.; de Beer, S.W. Picorna-like viral particles associated with mass mortalities in larval barramundi, Lates calcarifer (Bloch). J. Fish Dis. 1990, 13, 245–249. [Google Scholar] [CrossRef]
- Munday, B.L.; Kwang, J.; Moody, N. Betanodavirus infections of teleost fish: A review. J. Fish Dis. 2002, 25, 127–142. [Google Scholar] [CrossRef]
- Yoshikoshi, K.; Inoue, K. Viral nervous necrosis in hatchery-rearded larvae and juveniles of Japanese parrotfish, Oplegnathus fasciatus (Temminck & Schlegel). J. Fish Dis. 1990, 13, 69–77. [Google Scholar]
- Bloch, B.; Gravningen, K.; Larsen, J.L. Encephalomyelitis among turbot associated with a picornavirus-like agent. Dis. Aquat. Org. 1991, 10, 65–70. [Google Scholar] [CrossRef]
- Breuil, G.; Bonami, J.R.; Pepin, J.F.; Pichot, Y. Viral infection (picorna-like virus) associated with mass mortalities in hatchery-reared sea bass (Dicentrarchus labrax) larvae and juveniles. Aquaculture 1991, 97, 109–116. [Google Scholar] [CrossRef]
- Mori, K.; Nakai, T.; Nagahara, M.; Muroga, K.; Mekuchi, T.; Kanno, T. A viral disease in hatchery-reared larvae and juveniles of redspotted grouper. Fish Pathol. 1991, 26, 209–210. [Google Scholar] [CrossRef]
- Mori, K.; Nakai, T.; Muroga, K.; Arimoto, M.; Mushiake, K.; Furusawa, I. Properties of a new virus belonging to Nodaviridae found in larval striped jack (Pseudocaranx dentex) with nervous necrosis. Virology 1992, 187, 368–371. [Google Scholar] [CrossRef]
- Tanaka, S.; Takagi, M.; Miyazaki, T. Histopathological studies on viral nervous necrosis of sevenband grouper, Epinephelus septemfasciatus Thunberg, at the grow-out stage. J. Fish Dis. 2004, 27, 385–399. [Google Scholar] [CrossRef] [PubMed]
- Frerichs, G.N.; Roger, H.D.; Peric, Z. Cell culture isolation of piscine neuropathy nodavirus from juvenile sea bass, Dicentrarchus labrax. J. Gen. Virol. 1996, 77, 2067–2071. [Google Scholar] [CrossRef]
- Furusawa, R.; Okinaka, Y.; Uematsu, K.; Nakai, T. Screening of freshwater fish species for their susceptibility to a betanodavirus. Dis. Aquat. Org. 2007, 77, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Moody, N.J.G.; Horwood, P.F.; Reynolds, A.; Mahony, T.J.; Anderson, I.G.; Oakey, H.J. Phylogenetic analysis of betanodavirus isolates from Australian finfish. Dis. Aquat. Org. 2009, 87, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.K.; Mori, K.-I.; Okinaka, Y.; Nakai, T.; Park, S.C. Trash fish can be a source of betanodavirus for cultured marine fish. Aquaculture 2010, 302, 158–163. [Google Scholar] [CrossRef]
- Tan, C.; Huang, B.; Chang, S.F.; Ngoh, G.H.; Munday, B.; Chen, S.C.; Kwang, J. Determination of the complete nucleotide sequences of RNA1 and RNA2 from greasy grouper (Epinephelus tauvina) nervous necrosis virus, Singapore strain. J. Gen. Virol. 2001, 82, 647–653. [Google Scholar] [CrossRef]
- Nagai, T.; Nishizawa, T. Sequence of the non-structural protein gene encoded by RNA1 of striped jack nervous necrosis virus. J. Gen. Virol. 1999, 80, 3019–3022. [Google Scholar] [CrossRef] [PubMed]
- Skliris, G.P.; Krondiris, J.V.; Sieris, D.C.; Shinn, A.P.; Starkey, W.G.; Richards, R.H. Phylogenetic and antigenic characterisation of new fish nodavirus isolates from Europe and Asia. Virus Res. 2001, 75, 59–67. [Google Scholar] [CrossRef]
- Nishizawa, T.; Furuhashi, M.; Nagai, T.; Nakai, T.; Muroga, K. Genomic classification of fish nodaviruses by phylogenetic analysis of the coat protein gene. Appl. Environ. Microbiol. 1997, 63, 1633–1636. [Google Scholar] [CrossRef]
- Gagné, N.; Johnson, S.C.; Cook-Versloot, M.; MacKinnon, A.M.; Olivier, G. Molecular detection and characterization of nodavirus in several marine fish species from the northeastern Atlantic. Dis. Aquat. Org. 2004, 62, 181–189. [Google Scholar] [CrossRef]
- Johansen, R.; Sommerset, I.; Tørud, B.; Korsnes, K.; Hjortaas, M.J.; Nilsen, F.; Nerland, A.H.; Dannevig, B.H. Characterization of nodavirus and viral encephalopathy and retinopathy in farmed turbot, Scophthalmus maximus (L.). J. Fish Dis. 2004, 27, 591–601. [Google Scholar] [CrossRef]
- Nakai, T.; Mori, K.; Sugaya, T.; Nishioka, T.; Mushiake, K.; Yamashita, H. Current knowledge on viral nervous necrosis (VNN) and its causative betanodaviruses. Isr. J. Aquaculture 2009, 61, 198–207. [Google Scholar] [CrossRef]
- Sommerset, I.; Skern, R.; Biering, E.; Bleie, H.; Fiksdal, I.U.; Grove, S.; Nerland, A.H. Protection against Atlantic halibut nodavirus in turbot is induced by recombinant capsid protein vaccination but not following DNA vaccination. Fish Shellfish Immunol. 2005, 18, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Thiéry, R.; Cozien, J.; Cabon, J.; Lamour, F.; Baud, M.; Scheemann, A. Induction of protective immune response against viral nervous necrosis in the European sea bass Dicentrarchus labrax by using betanodavirus virus-like particles. J. Virol. 2006, 80, 10201–10207. [Google Scholar] [CrossRef] [PubMed]
- Kai, Y.-H.; Chi, S.-C. Efficacies of inactivated vaccines against betanodavirus in grouper larvae (Epinephelus coioides) by bath immunization. Vaccine 2008, 26, 1450–1457. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Mori, K,; Kuroda, A. ; Nakai, T. Neutralizing antibody levels for protection against betanodavirus infection in sevenband grouper, Epinephelus septemfasciatus (Thunberg), immunized with an inactivated virus vaccine. J. Fish Dis. 2009, 32, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Pakingking, R.; Bautista, N.B.; de Jesus-Ayson, E.G.; Reyes, O. Protective immunity against viral nervous necrosis (VNN) in brown-marbled grouper (Epinephelus fuscogutattus) following vaccination with inactivated betanodavirus. Fish Shellfish Immunol. 2010, 28, 525–533. [Google Scholar] [CrossRef]
- Pakingking, R.; Seron, R.; dela Pena, L.; Mori, K.; Yamashita, H.; Nakai, T. Immune responses of Asian sea bass, Lates calcarifer Bloch, against inactivated betanodavirus vaccine. J. Fish Dis. 2009, 32, 457–463. [Google Scholar] [CrossRef]
- Thorud, K.E.; Djupvik, H.O. Infectious salmon anaemia in Atlantic salmon (Salmo salar L.). Bull. Eur. Assoc. Fish Pathol. 1988, 8, 109–111. [Google Scholar]
- Dannevig, B.H.; Falk, K.; Namork, E. Isolation of the causal virus of infectious salmon anaemia (ISA) in a long-term cell line from Atlantic salmon head kidney. J Gen. Virol. 1995, 76, 1353–1359. [Google Scholar] [CrossRef]
- Dannevig, B.H.; Mjaaland, S.; Rimstad, E. Infectious salmon anemia virus. In Encyclopaedia of Virology; Mahy, B.W.J., Van Regenmortel, M.H.V., Eds.; Elsevier: Oxford, UK, 2008; pp. 89–95. [Google Scholar]
- Mullins, J.E.; Groman, D.; Wadowska, D. Infectious salmon anaemia in salt water Atlantic salmon (Salmo salar L.) in New Brunswick, Canada. Bull. Eur. Assoc. Fish Pathol. 1998, 18, 110–114. [Google Scholar]
- Bouchard, D.; Keleher, W.; Opitz, H.M.; Blake, S.; Edwards, K.C.; Nicholson, B.L. Isolation of infectious salmon anemia virus (ISAV) from Atlantic salmon in New Brunswick, Canada. Dis. Aquat. Org. 1999, 35, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Lovely, J.E.; Dannevig, B.H.; Falk, K.; Hutchin, L.; Mackinnon, A.M.; Melville, K.J.; Rimstad, E.; Griffiths, S.G. First identification of infectious salmon anaemia virus in North America with haemorrhagic kidney syndrome. Dis. Aquat. Org. 1999, 35, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Rodger, H.D.; Turnbull, T.; Muir, F.; Millar, S.; Richards, R.H. Infectious salmon anaemia (ISA) in the United Kingdom. Bull. Eur. Assoc. Fish Pathol. 1998, 18, 115–116. [Google Scholar]
- Rowley, H.M.; Campbell, S.J.; Curran, W.L.; Turnbull, T.; Bryson, D.G. Isolation of infectious salmon anaemia virus (ISAV) from Scottish farmed Atlantic salmon, Salmo salar L. J. Fish Dis. 1999, 22, 483–487. [Google Scholar] [CrossRef]
- Christiansen, D.H.; Østergaard, P.S.; Snow, M.; Dale, O.B.; Falk, K. A low-pathogenic variant of infectious salmon anemia virus (ISAV-HPR0) is highly prevalent and causes a non-clinical transient infection in farmed Atlantic salmon (Salmo salar L.) in the Faroe Islands. J. Gen. Virol. 2011, 92, 909–18. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, D.A.; Brockway, K.; Giray, C.; Keleher, W.; Merrill, P.L. First report of infectious salmon anaemia (ISA) in the United States. Bull. Eur. Assoc. Fish Pathol. 2001, 21, 86–88. [Google Scholar]
- Godoy, M.G.; Aedo, A.; Kibenge, M.J.T.; Groman, D.B.; Yason, C.V.; Grothusen, H.; Lisperguer, A.; Calbucura, M.; Avendaño, F.; Imilán, M.; Jarpa, M.; Kibenge, F.S.B. First detection, isolation and molecular characterization of infectious salmon anaemia virus associated with clinical disease in farmed Atlantic salmon (Salmo salar) in Chile. BMC Vet. Res. 2008, 4, 28. [Google Scholar] [CrossRef]
- Geoghegan, F. First isolation and identification of ISAV in Ireland. In Proceedings of the 6th Annual Meeting of EU National Reference Laboratories for Fish Diseases, Brussels, Belgium, 23–24 September 2002. [Google Scholar]
- Kibenge, F.S.B.; Gárate, O.N.; Johnson, G.; Arriagada, R.; Kibenge, M.J.T.; Wadowska, D. Isolation and identification of infectious salmon anaemia virus (ISAV) from Coho salmon in Chile. Dis. Aquat. Org. 2001, 45, 9–18. [Google Scholar] [CrossRef]
- Krossøy, B.; Hordvik, I.; Nilsen, F.; Nylund, A.; Endresen, C. The putative polymerase sequence of infectious salmon anemia virus suggests a new genus within the Orthomyxoviridae. J. Virol. 1999, 73, 2136–2142. [Google Scholar] [CrossRef]
- Kawaoka, Y.; Cox, N.J.; Haller, O.; Hongo, S.; Kaverin, N.; Klenk, H.D.; Lamb, R.A.; McCauley, J.; Palese, P.; Rimstad, E.; Webster, R.G. Infectious Salmon Anaemia Virus. In Virus Taxonomy—Eight Report of the International Committee on Taxonomy Viruses; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Academic Press: New York, NY, USA, 2005; pp. 681–693. [Google Scholar]
- Clouthier, S.C.; Rector, T.; Brown, N.E.C.; Anderson, E.D. Genomic organization of infectious salmon anaemia. J. Gen. Virol. 2002, 83, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.O.; Snow. M. Genetic analysis of infectious salmon anaemia virus (ISAV) from Scotland. Dis. Aquat. Org. 2000, 41, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Inglis, J.A.; Bruce, J.; Cunningham, C.O. Nucleotide sequence variation in isolates of infectious salmon anaemia virus (ISAV) from Atlantic salmon Salmo salar in Scotland and Norway. Dis. Aquat. Org. 2001, 43, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, R.J.; Cook, M.; Melville, K.; Simard, N.; Cusack, R.; Griffiths, S. Identification of infectious salmon anaemia virus in Atlantic salmon from Nova Scotia (Canada): Evidence for functional strain differences. Dis. Aquat. Org. 2001, 44, 171–178. [Google Scholar] [CrossRef]
- Devold, M.; Falk, K.; Dale, O.B.; Krossøy, B.; Biering, E.; Aspehaug, V.; Nilsen, F.; Nylund, A. Strain variation, based on the hemagglutinin gene, in Norwegian ISA virus isolates collected from 1987 to 2001: Indications of recombination. Dis. Aquat. Org. 2001, 47, 119–128. [Google Scholar] [CrossRef]
- Kibenge, F.S.B.; Kibenge, M.J.T.; McKenna, P.K.; Stothard, P.; Marshall, R.; Cusack, R.R.; McGeachy, S. Antigenic variation among isolates of infectious salmon anaemia virus correlates with genetic variation of the viral haemagglutinin gene. J. Gen. Virol. 2001, 82, 2869–2879. [Google Scholar] [CrossRef]
- Nylund, A.; Devold, M.; Plarre, H.; Isdal, E.; Aarseth, M. Emergence and maintenance of infectious salmon anaemia virus (ISAV) in Europe: a new hypothesis. Dis. Aquat. Org. 2003, 56, 11–24. [Google Scholar] [CrossRef]
- Cook-Versloot, M.; Griffiths, S.; Cusack, R.; McGeachy, S.; Ritchie, R. Identification and characterisation of infectious salmon anaemia virus (ISAV) haemagglutinin gene highly polymorphic region (HPR) type 0 in North America. Bull. Eur. Assoc. Fish Pathol. 2004, 24, 203–208. [Google Scholar]
- Cunningham, C.O.; Gregory, A.; Black, J.; Simpson, I.; Raynard, R.S. A novel variant of the infectious salmon anaemia virus (ISAV) haemagglutinin gene suggests mechanisms for virus diversity. Bull. Eur. Assoc. Fish Pathol. 2002, 22, 366–373. [Google Scholar]
- Devold, M.; Karlsen, M.; Nylund, A. Sequence analysis of the fusion protein gene from infectious salmon anemia virus isolates: evidence of recombination and reassortment. J. Gen. Virol. 2006, 87, 2031–2040. [Google Scholar] [CrossRef]
- Markussen, T.; Jonassen, C.M.; Numanovic, S.; Braaen, S.; Hjortaas, M.; Nilsen, H.; Mjaaland, S. Evolutionary mechanisms involved in the virulence of infectious salmon anaemia virus (ISAV), a piscine orthomyxovirus. Virology 2008, 374, 515–527. [Google Scholar] [CrossRef] [PubMed]
- McBeath, A.J.; Bain, N.; Snow, M. Surveillance for infectious salmon anaemia virus HPR0 in marine Atlantic salmon farms across Scotland. Dis. Aquat. Org. 2009, 87, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Nylund, A.; Devold, M.; Mullins, J.; Plarre, H. Herring (Clupea harengus): A host for infectious salmon anaemia virus (ISAV). Bull. Eur. Assoc. Fish Pathol. 2002, 22, 311–318. [Google Scholar]
- Snow, M.; Raynard, R.S.; Inglis, J.; Bruno, D.W. Investigation into the potential for seawater rainbow trout (Oncorhynchus mykiss) to act as vectors of infectious salmon anaemia virus (ISAV). Bull. Eur. Assoc. Fish Pathol. 2001, 21, 252–262. [Google Scholar]
- Snow, M.; Raynard, R.S.; Bruno, D.W. Comparative susceptibility of Arctic char (Salvelinus alpinus), rainbow trout (Oncorhynchus mykiss) and brown trout (Salmo trutta) to the Scottish isolate of infectious salmon anaemia virus. Aquaculture 2001, 196, 47–54. [Google Scholar] [CrossRef]
- Nylund, A.; Jakobsen, P. Sea trout as a carrier of infectious salmon anaemia virus. J. Fish Biol. 1995, 47, 174–176. [Google Scholar] [CrossRef]
- Rolland, J.B.; Winton, J.R. Relative resistance of Pacific salmon to infectious salmon anaemia virus. J. Fish Dis. 2003, 26, 511–520. [Google Scholar] [CrossRef]
- Dannevig, B.H.; Thorud, K.E. Other viral diseases and agents of cold-water fish: Infectious salmon anaemia, pancreas disease and viral erythrocytic necrosis. In Fish Diseases and Disorders. Volume 3: Viral, Bacterial and Fungal Infections; Woo, P.T.K., Bruno, D.W., Eds.; CABI Publishing: Wallingford, UK, 1999; pp. 149–175. [Google Scholar]
- Jarp, J.; Karlsen, E. Infectious salmon anaemia (ISA) risk factors in sea-cultured Atlantic salmon Salmo salar. Dis. Aquat. Org. 1997, 28, 79–86. [Google Scholar] [CrossRef]
- Gustafson, L.L.; Ellis, S.K.; Bartlett, C.A. Using expert opinion to identify risk factors important to infectious salmon-anemia (ISA) outbreaks on salmon farms in Maine, USA and New Brunswick, Canada. Prev. Vet. Med. 2005, 70, 17–28. [Google Scholar] [CrossRef]
- Mjaaland, S.; Markussen, T.; Sindre, H.; Kjøglum, S.; Dannevig, B.H.; Larsen, S.; Grimholt, U. Susceptibility and immune responses following experimental infection of MHC compatible Atlantic salmon (Salmo salar L.) with different infectious salmon anaemia virus isolates. Arch. Virol. 2005, 150, 2195–2216. [Google Scholar] [CrossRef]
- Falk, K.; Dannevig, B.H. Demonstration of a protective immune response in infectious salmon anaemia (ISA)-infected Atlantic salmon Salmo salar. Dis. Aquat. Org. 1995, 21, 1–5. [Google Scholar] [CrossRef]
- Jones, S.R.M.; MacKinnon, A.M.; Salonius, K. Vaccination of freshwater-reared Atlantic salmon reduces mortality associated with infectious salmon anaemia virus. Bull. Eur. Assoc. Fish Pathol. 1999, 19, 98–101. [Google Scholar]
- Kibenge, F.S.B.; Kibenge, M.J.T.; Joseph, T.; McDougall, J. The development of infectious salmon anaemia vaccines in Canada. In Technical Bulletin 1902, International Response to Infectious Salmon Anemia: Prevention, Control and Eradication; Miller, O., Cipriano, R.C., Eds.; USDA, APHIS; US Dept Interior, US Geological Survey: US Dept Commerce, National Marine Fisheries Service, Washington DC, USA, 2003; pp. 39–49. [Google Scholar]
- Mikalsen, A.B.; Sindre, H.; Torgersen, J.; Rimstad, E. Protective effects of a DNA vaccine expressing the infectious salmon anemia virus hemagglutinin-esterase in Atlantic salmon. Vaccine 2005, 23, 4895–4905. [Google Scholar] [CrossRef]
- Nelson, R.T.; McLoughlin, M.F.; Rowley, H.M.; Platten, M.A.; McCormick, J.I. Isolation of a toga-like virus from farmed Atlantic salmon Salmon salar with pancreas disease. Dis. Aquat. Org. 1995, 22, 25–32. [Google Scholar] [CrossRef]
- Christie, K.E.; Fyrand, K.; Holtet, L.; Rowley, H.M. Isolation of pancreas disease virus from farmed Atlantic salmon, Salmo salar L., in Norway. J. Fish Dis. 1998, 21, 391–394. [Google Scholar] [CrossRef]
- Rowley, H.M.; Doherty, C.E.; McLoughlin, M.F.; Welsh, M.D. Isolation of salmon pancreas disease virus (SPDV) from farmed Atlantic salmon, Salmo salar L., in Scotland. J. Fish Dis. 1998, 21, 469–471. [Google Scholar] [CrossRef]
- Weston, J.; Welsh, M.D.; McLoughlin, M.F.; Todd, D. Salmon pancreas disease virus, an alphavirus infecting farmed Atlantic salmon, Salmo salar L. Virology 1999, 256, 188–195. [Google Scholar] [CrossRef]
- Fringuelli, E.; Rowley, H.M.; Wilson, J.C.; Hunter, R.; Rodger, H.; Graham, D.A. Phylogenetic analyses and molecular epidemiology of European salmonid alphaviruses (SAV) based on partial E2 and nsP3 gene nucleotide sequences. J. Fish Dis. 2008, 31, 811–823. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, M.F.; Graham, D.A. Alphavirus infections in salmonids—A review. J. Fish Dis. 2007, 30, 511–531. [Google Scholar] [CrossRef]
- Hodneland, K.; Bratland, A.; Christie, K.E.; Endresen, C.; Nylund, A. New subtype of salmonid alphavirus (SAV), Togaviridae, from Atlantic salmon Salmo salar and rainbow trout Oncorhynchus mykiss in Norway. Dis. Aquat. Org. 2005, 66, 113–120. [Google Scholar] [CrossRef]
- Chamberlain, R.W. Epidemiology of arthropod-borne togaviruses: The role of arthropods as hosts and vectors and of vertebrate hosts in natural transmission cycles. In The Togaviruses; Schlesinger, R.W., Ed.; Academic Press: New York, NY, USA, 1980; pp. 175–227. [Google Scholar]
- McLoughlin, M.F.; Nelson, R.T.; Rowley, H.M.; Cox, D.I.; Grant, A.N. Experimental pancreas disease virus in Atlantic salmon Salmo salar post-smolts induced by salmon pancreas disease virus (SPDV). Dis. Aquat. Org. 1996, 26, 117–124. [Google Scholar] [CrossRef]
- Karlsen, M.; Villoing, S.; Rimstad, E.; Nylund, A. Characterisation of untranslated regions of the salmonid alphavirus 3 (SAV3) genome and construction of a SAV3 based replicon. Virol. J. 2009, 6, 173. [Google Scholar] [CrossRef] [PubMed]
- Weston, J.; Villoing, S.; Bremont, M.; Castric, J.; Pfeffer, M.; Jewhurst, V.; McLoughlin, M.; Rødseth, O.; Christie, K.E.; Koumans, J.; Todd, D. Comparison of two aquatic alphaviruses, salmon pancreas disease virus and sleeping disease virus, by using genome sequence analysis, monoclonal reactivity, and cross-infection. J. Virol. 2002, 76, 6155–6163. [Google Scholar] [CrossRef]
- Boucher, P.; Baudin Laurencin, F. Sleeping disease and pancreas disease; comparative histopathology and acquired cross-protection. J. Fish Dis. 1996, 19, 303–310. [Google Scholar] [CrossRef]
- Houghton, G. Acquired protection in Atlantic salmon Salmo salar parr and post-smolts against pancreas disease. Dis. Aquat. Org. 1994, 18, 109–118. [Google Scholar] [CrossRef]
- Lopez-Doriga, M.V.; Smail, D.A.; Smith, R.J.; Domenech, A.; Castric, J.; Smith, P.D.; Ellis, A.E. Isolation of salmon pancreas disease virus (SPDV) in cell culture and its ability to protect against infection by the ‘wild type’agent. Fish Shellfish Immunol. 2001, 11, 505–522. [Google Scholar] [CrossRef]
- Moriette, C.; LeBerre, M.; Lamoureux, A.; Thanh-Lan, L.; Bremont, M. Recovery of a recombinant salmonid alphavirus fully attenuated and protective for rainbow trout. J. Virol. 2005, 80, 4088–4098. [Google Scholar] [CrossRef] [PubMed]
- Bootland, L.M.; Leong, J.C. Infectious haematopoietic necrosis virus. In Fish Diseases and Disorders. Volume 3: Viral, Bacterial and Fungal Infections; Woo, P.T.K., Bruno, D.W., Eds.; CABI Publishing: Wallingford, UK, 1999; pp. 57–122. [Google Scholar]
- Rohovec, J.S.; Fryer, J.L. Dispersal of microbial pathogens through introductions and transfer of finfish. J. Shellfish Res. 1988, 7, 555. [Google Scholar]
- Enzmann, P.-J.; Castric, J.; Bovo, G.; Thiery, R.; Fichtner, D.; Schütze, H.; Wahli, T. Evolution of infectious hematopoietic necrosis virus (IHNV), a fish rhabdovirus, in Europe over 20 years: Implications for control. Dis. Aquat. Org. 2010, 89, 9–15. [Google Scholar] [CrossRef]
- Nishizawa, T.; Kinoshita, S.; Kim, W.-S.; Higashi, S.; Yoshimizu, M. Nucleotide diversity of Japanese isolates of infectious hematopoietic necrosis virus (IHNV) based on the glycoprotein gene. Dis. Aquat. Org. 2006, 71, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Schutze, H.; Enzmann, P.J.; Kuchling. R.; Mundt, E.; Niemann, H.; Mettenleiter, T.C. Complete genomic sequence of the fish rhabdovirus infectious haematopoietic necrosis virus. J. Gen. Virol. 1995, 76, 2519–2527. [Google Scholar] [CrossRef]
- Morzunov, S.P.; Winton, J.R.; Nichol, S.T. The complete genome structure and phylogenetic relationship of infectious hematopoietic necrosis virus. Virus Res. 1995, 38, 175–192. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, D.; Wood, J. A natural epizootic of infectious haematopoietic necrosis in imported Atlantic salmon, Salmo salar L., reared in the enzootic region. J. Fish Dis. 1986, 9, 173–175. [Google Scholar] [CrossRef]
- LaPatra, S.E.; Jones, G.R.; Lauda, K.A.; McDowell, T.S.; Schneider, R.; Hedrick, R.P. White sturgeon as a potential vector of infectious hematopoietic necrosis virus. J. Aquat. Anim. Health 1995, 7, 225–230. [Google Scholar] [CrossRef]
- Kimura, T.; Yoshimizu, M.; Gorie, S. A new rhabdovirus isolated in Japan from cultured hirame (Japanese flounder) Paralichthys olivaceus and ayu Plecoglossus altivelis. Dis. Aquat. Org. 1986, 1, 209–217. [Google Scholar] [CrossRef]
- Kurath, G.; Higman, K.H.; Björklund, H.V. Distribution and variation of NV genes in fish rhabdoviruses. J. Gen. Virol. 1997, 78, 113–117. [Google Scholar] [CrossRef]
- LaPatra, SE. Factors affecting pathogenicity of infectious hematopoietic necrosis virus (IHNV) for salmonid fish. J. Aquat. Anim. Health 1998, 10, 121–131. [Google Scholar] [CrossRef]
- Kurath, G.; Garver, K.A.; Troyer, R.M.; Emmenegger, E.J.; Einer-Jensen, K.; Anderson, E.D. Phylogeography of infectious hematopoietic necrosis virus. J. Gen. Virol. 2003, 84, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Garver, K.A.; Troyer, R.M.; Kurath, G. Two distinct phylogenetic clades of infectious hematopoietic necrosis virus overlap within the Columbia River basin. Dis. Aquat. Org. 2003, 55, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Garver, K.A.; Batts, W.N.; Kurath, G. Virulence comparisons of infectious hematopoietic necrosis virus U and M genogroups in sockeye salmon and rainbow trout. J. Aquat. Anim. Health 2006, 18, 232–243. [Google Scholar] [CrossRef]
- Peñaranda, M.M.D.; Purcell, M.K.; Kurath, G. Differential virulence mechanisms of infectious hematopoietic necrosis virus in rainbow trout (Oncorhynchus mykiss) include host entry and virus replication kinetics. J. Gen. Virol. 2009, 90, 2172–2182. [Google Scholar] [CrossRef]
- Engelking, H.M.; Leong, J.C. Glycoprotein from infectious hematopoietic necrosis virus (IHNV) induces protective immunity against five IHNV types. J. Aquat. Anim. Health 1989, 1, 291–300. [Google Scholar] [CrossRef]
- Hedrick, R.P.; LaPatra, S.E.; Yun, S.; Lauda, K.A.; Jones, G.R.; Congleton, J.L.; De Kinkelin, P. Induction of protection from infectious hematopoietic necrosis virus in rainbow trout Oncorhynchus mykiss by pre-exposure to the avirulent cutthroat trout virus (CTV). Dis. Aquat. Org. 1994, 20, 111–118. [Google Scholar] [CrossRef]
- LaPatra, S.E.; Lauda, K.A.; Jones, G.R. Antigenic variants of infectious hematopoietic necrosis virus and implications for vaccine development. Dis. Aquat. Org. 1994, 20, 119–126. [Google Scholar] [CrossRef]
- Anderson, E.; Clouthier, S.; Shewmaker, W.; Weighall, A.; LaPatra, S. Inactivated infectious haematopoietic necrosis virus (IHNV) vaccines. J. Fish Dis. 2008, 31, 729–745. [Google Scholar] [CrossRef]
- Noonan, B.; Enzmann, P.J.; Trust, T.J. Recombinant infectious hematopoietic necrosis virus and viral hemorrhagic septicemia virus glycoprotein epitopes expressed in Aeromonas salmonicida induce protective immunity in rainbow trout (Oncorhynchus mykiss). Appl. Environ. Microbiol. 1995, 61, 3586–3591. [Google Scholar] [CrossRef]
- Corbeil, S.; LaPatra, S.E.; Anderson, E.D.; Kurath, G. Nanogram quantities of a DNA vaccine protect rainbow trout fry against heterologous strains of infectious hematopoietic necrosis virus. Vaccine 2000, 18, 2817–2824. [Google Scholar] [CrossRef] [PubMed]
- Kurath, G.; Garver, K.A.; Corbeil, S.; Elliott, D.G.; Anderson, E.D.; LaPatra, S.E. Protective immunity and lack of histopathological damage two years after DNA vaccination against infectious hematopoietic necrosis virus in trout. Vaccine 2006, 24, 345–354. [Google Scholar] [CrossRef]
- Peñaranda, M.M.D.; LaPatra, S.; Kurath, G. Specificity of DNA vaccines against the U and M genogroups of infectious hematopoietic necrosis virus (IHNV) in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2011, 31, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.; Chiou, P.P.; Leong, J.-A. Development of a suicidal DNA vaccine for infectious hematopoietic necrosis virus. Fish Shellfish Immunol. 2011, 30, 815–823. [Google Scholar] [CrossRef]
- Yan, X.; Yu, Z.; Zhang, P.; Battisti, A.J.; Holdaway, H.A.; Chipman, P.R.; Bajaj, C.; Bergoin, M.; Rossmann, M.G.; Baker, T.S. The capsid proteins of a large, icosahedral dsDNA virus. J. Mol. Biol. 2009, 385, 1287–1299. [Google Scholar] [CrossRef]
- Langdon, J.S.; Humphrey, J.D.; Williams, L.M.; Hyatt, A.D.; Westbury, H.A. First virus isolation from Australian fish: an iridovirus-like pathogen from redfin perch, Perca fluviatilis L. J. Fish Dis. 1986, 9, 263–268. [Google Scholar] [CrossRef]
- Langdon, J.S.; Humphrey, J.D. Epizootic haematopoietic necrosis a new viral disease in redfin perch, Perca fluviatilis L., in Australia. J. Fish Dis. 1987, 10, 289–298. [Google Scholar] [CrossRef]
- Langdon, J.S.; Humphrey, J.D.; Williams, L.M. Outbreaks of an EHNV-like iridovirus in cultured rainbow trout, Salmo gairdneri Richardson, in Australia. J. Fish Dis. 1988, 11, 93–96. [Google Scholar] [CrossRef]
- Whittington, R.J.; Reddacliff, G.L. Influence of environmental temperature on experimental infection of redfin perch (Perca fluviatilis) and rainbow trout (Oncorhynchus mykiss) with epizootic haematopoietic necrosis virus, an Australian iridovirus. Aus. Vet. J. 1995, 72, 421–424. [Google Scholar] [CrossRef]
- Langdon, J.S. Experimental transmission and pathogenicity of epizootic haematopoietic necrosis virus (EHNV) in redfin perch, Perca fluviatilis L., and 11 other teleosts. J. Fish Dis. 1989, 12, 295–310. [Google Scholar] [CrossRef]
- Whittington, R.J.; Philbey, A.; Reddacliff, G.L.; MacGown, A.R. Epidemiology of epizootic haematopoietic necrosis virus (EHNV) infection in farmed rainbow trout, Oncorhynchus mykiss (Walbaum): Findings based on virus isolation, antigen capture ELISA and serology. J. Fish Dis. 1994, 17, 205–218. [Google Scholar] [CrossRef]
- Reddacliff, L.A.; Whittington, R.J. Pathology of epizootic haematopoietic necrosis virus (EHNV) infection in rainbow trout (Oncorhynchus mykiss Walbaum) and redfin perch (Perca fluviatilis L). J. Comp. Pathol. 1996, 115, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Whittington, R.J.; Becker, J.A.; Dennis, M.M. Iridovirus infections in finfish—A critical review with emphasis on ranaviruses. J. Fish Dis. 2010, 33, 95–122. [Google Scholar] [CrossRef] [PubMed]
- Whittington, R.J.; Kearns, C.; Hyatt, A.D.; Hengstberger, S.; Rutzou, T. Spread of epizootic haematopoietic necrosis virus (EHNV) in redfin perch (Perca fluviatilis) in southern Australia. Aus. Vet. J. 1996, 73, 112–114. [Google Scholar] [CrossRef]
- Meyers, T.R.; Winton, J.R. Viral hemorrhagic septicaemia virus in North America. Ann. Rev. Fish Dis. 1995, 5, 3–24. [Google Scholar] [CrossRef]
- Oshima, K.H.; Higman, K.H.; Arakawa, C.K.; de Kinkelin, P.; Jørgensen, P.E.V.; Meyers, T.R.; Winton, J.R. Genetic comparison of viral hemorrhagic septicaemia virus isolates from North America and Europe. Dis. Aquat. Org. 1993, 17, 73–80. [Google Scholar] [CrossRef]
- Batts, W.N.; Arakawa, C.K.; Bernard, J.; Winton, J.R. Isolates of viral hemorrhagic septicemia virus from North America and Europe can be detected and distinguished by DNA probes. Dis. Aquat. Org. 1993, 17, 67–71. [Google Scholar] [CrossRef]
- Faisal, M. Michigan State University, MI, USA. Viruses to be submitted for publication. 2011. [Google Scholar]
- Smail, D.A. Viral haemorrhagic septicaemia. In Fish Diseases and Disorders. Volume 3: Viral, Bacterial and Fungal Infections; Woo, P.T.K., Bruno, D.W., Eds.; CABI Publishing: Wallingford, UK, 1999; pp. 123–148. [Google Scholar]
- Skall, H.F.; Olesen, N.J.; Mellergaard, S. Viral hemorrhagic septicaemia virus in marine fish and its implications for fish farming—A review. J. Fish Dis. 2005, 28, 509–529. [Google Scholar] [CrossRef] [PubMed]
- Stone, D.M.; Ferguson, H.W.; Tyson, P.A.; Savage, J.; Wood, G.; Dodge, M.J; Woolford, G.; Dixon, P.F.; Feist, S.W.; Way, K. The first report of viral haemorrhagic septicaemia in farmed rainbow trout, Oncorhynchus mykiss (Walbaum), in the United Kingdom. J. Fish Dis. 2008, 31, 775–784. [Google Scholar] [CrossRef]
- Dale, O.B.; Ørpetveit, I.; Lyngstad, T.M.; Kahns, S.; Skall, H.F.; Olesen, N.J.; Dannevig, B.H. Outbreak of viral haemorrhagic septicaemia (VHS) in seawater-farmed rainbow trout in Norway caused by VHS virus Genotype III. Dis. Aquat. Org. 2009, 85, 93–103. [Google Scholar] [CrossRef]
- Mellergaard, S. Surveillance and eradication of VHS in Denmark. In Report on the 15th Annual Meeting of EU National Reference Laboratories for Fish Diseases, Copenhagen, Denmark, 26–27 May; 2011; p. 18. [Google Scholar]
- Vestergard Jorgensen, P.E. Egtved virus: Temperature-dependent immune response of trout to infection with low-virulence virus. J. Fish Dis. 1982, 5, 47–55. [Google Scholar] [CrossRef]
- de Kinkelin, P.; Bearzotti, M.; Castric, J.; Nougayrede, P.; Lecocqxhonneux, F.; Thiry, M. Eighteen years of vaccination against viral haemorrhagic septicaemia in France. Vet. Res. 1995, 26, 379–387. [Google Scholar] [PubMed]
- Lorenzen, E.; Lorenzen, N.; Einer-Jensen, K.; Brudeseth, B.; Evensen, Ø. Time course study of in situ expression of antigens following DNA-vaccination against VHS in rainbow trout Oncorhynchus mykiss (Walbaum) fry. Fish Shellfish Immunol. 2005, 19, 27–41. [Google Scholar] [CrossRef] [PubMed]
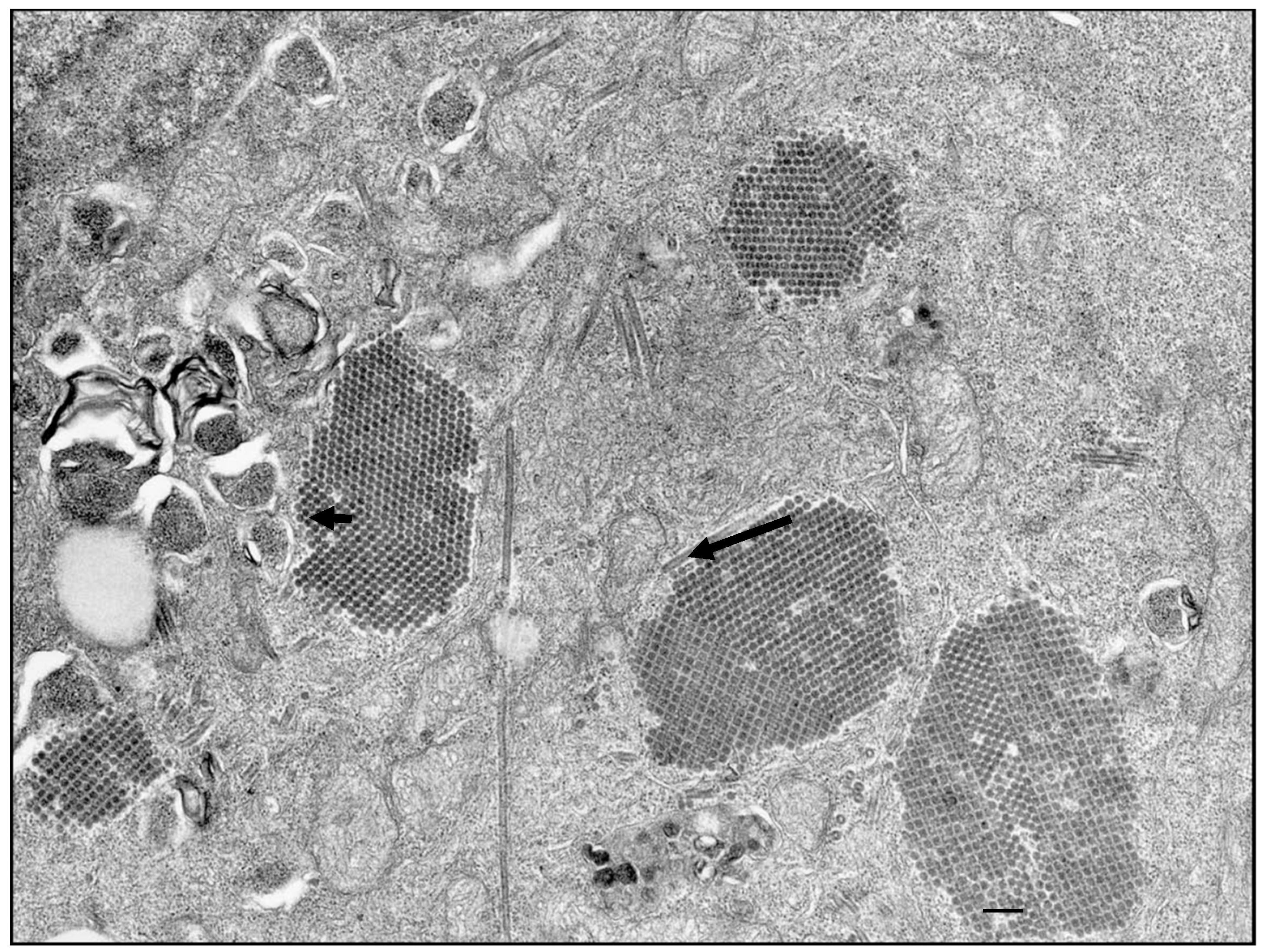
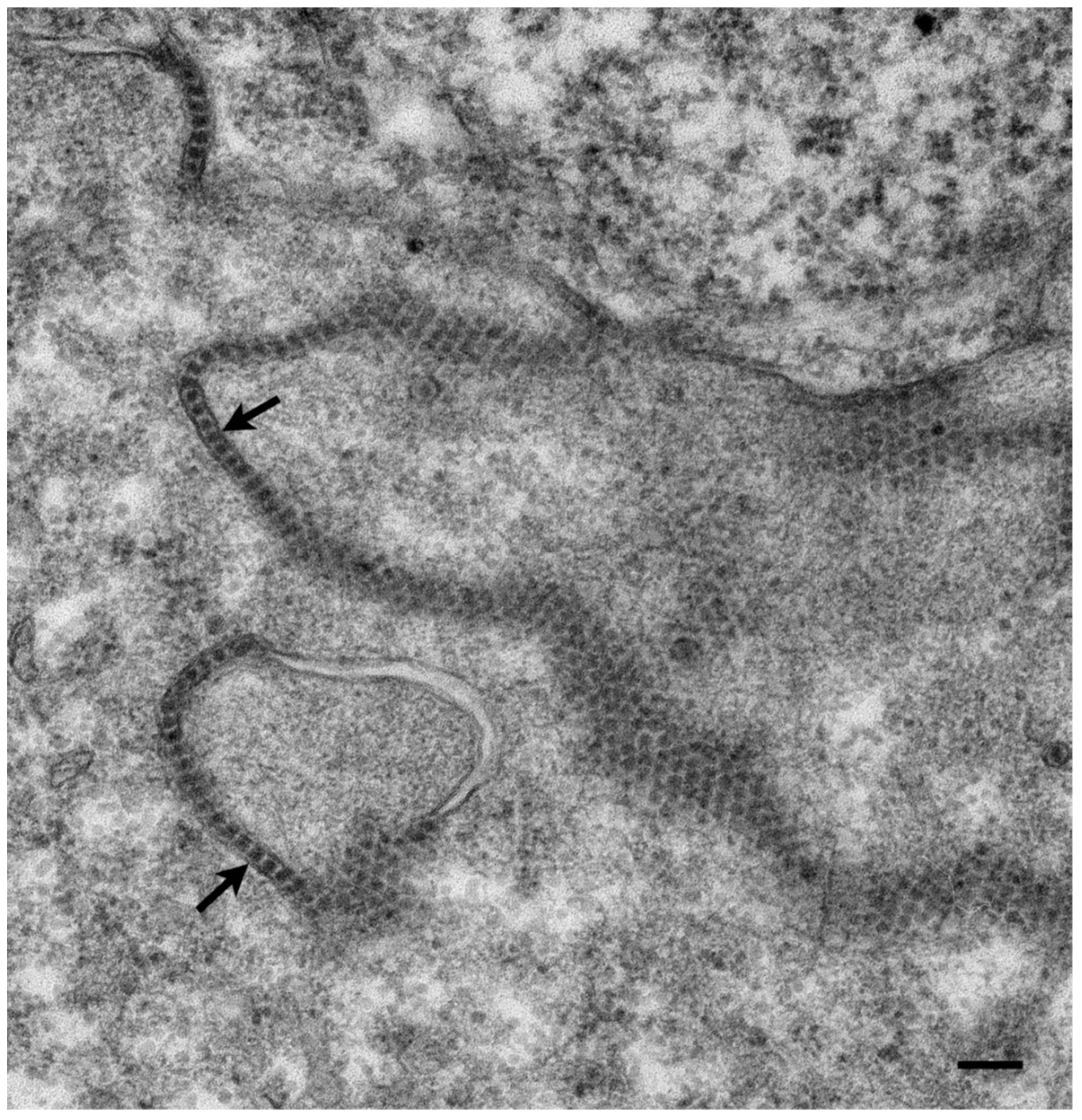



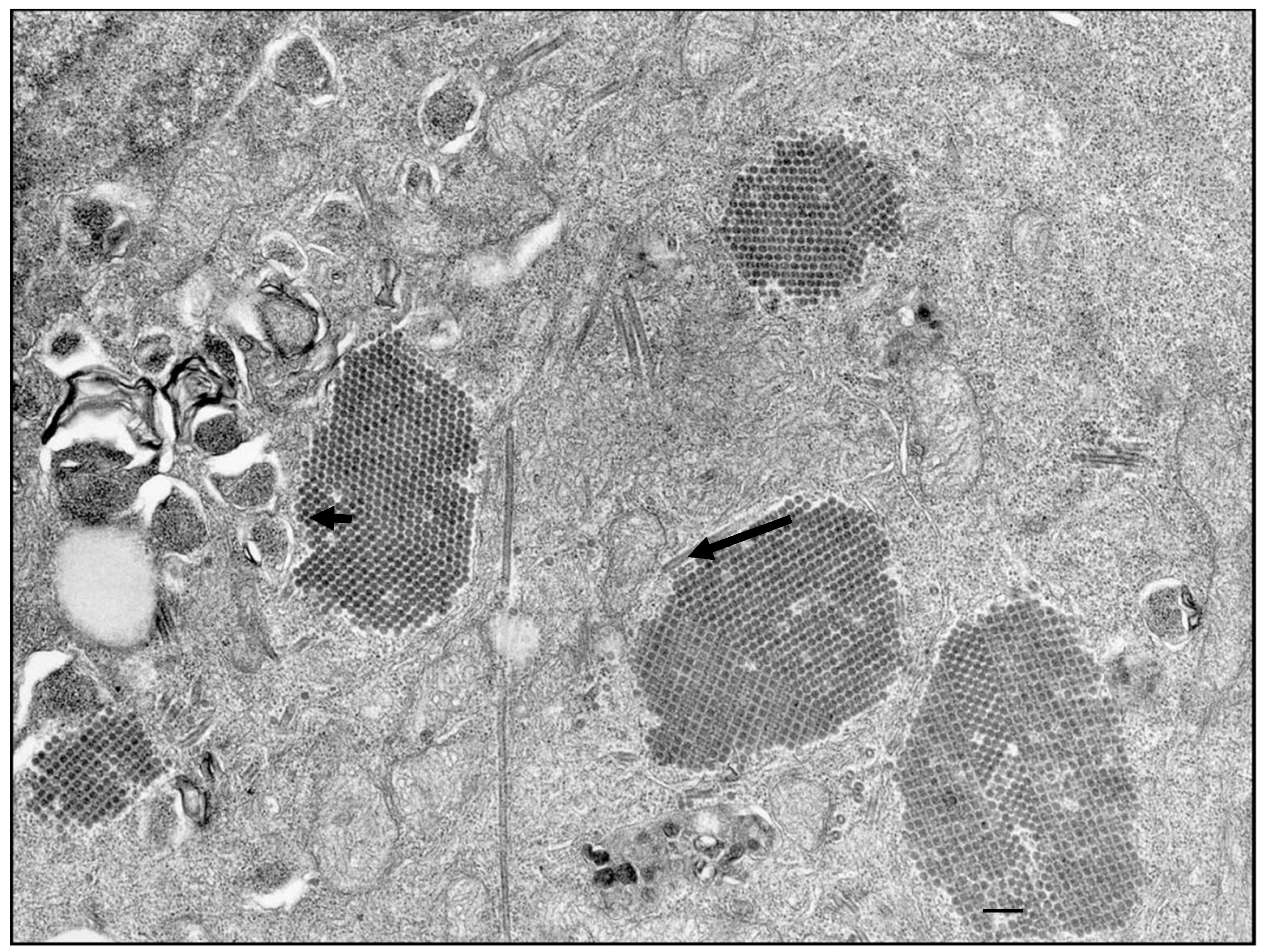{kind=link}
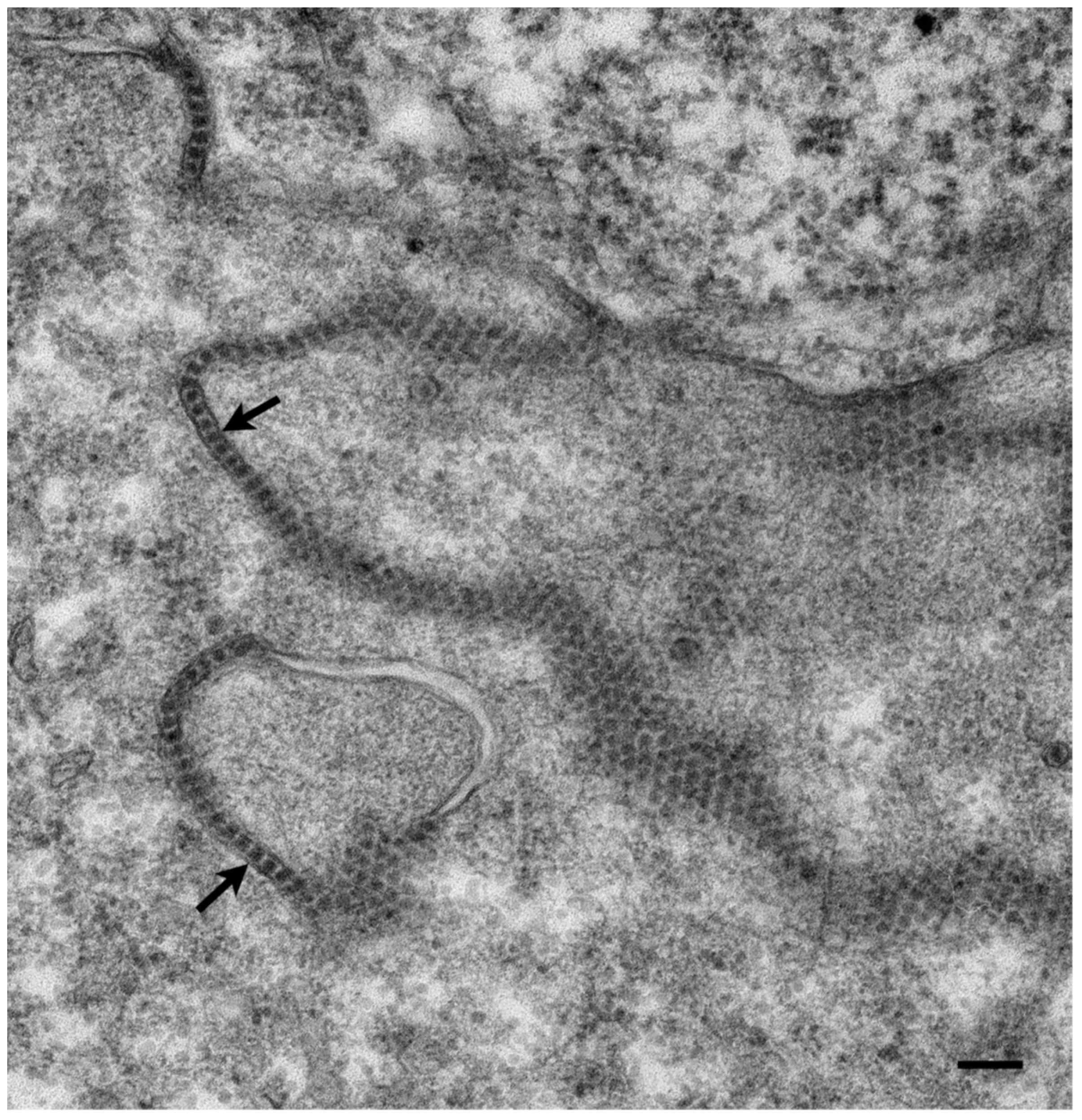{kind=link}
{kind=link}
{kind=link}
| Virus Subtype | Host Range | Disease | Geographical Range |
|---|---|---|---|
| SAV1 | Marine Atlantic salmon | Pancreas disease | Ireland |
| SAV2 | Freshwater rainbow trout | Sleeping disease | England, France, Germany, Italy, Scotland, Spain |
| SAV3 | Sea-reared Atlantic salmon, rainbow trout | Pancreas disease | Norway |
| SAV4 | Marine Atlantic salmon | Pancreas disease | Ireland, Scotland |
| SAV5 | Marine Atlantic salmon | Pancreas disease | Scotland |
| SAV6 | Marine Atlantic salmon | Pancreas disease | Ireland |
© 2011 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Crane, M.; Hyatt, A. Viruses of Fish: An Overview of Significant Pathogens. Viruses 2011, 3, 2025-2046. https://doi.org/10.3390/v3112025
Crane M, Hyatt A. Viruses of Fish: An Overview of Significant Pathogens. Viruses. 2011; 3(11):2025-2046. https://doi.org/10.3390/v3112025
Chicago/Turabian StyleCrane, Mark, and Alex Hyatt. 2011. "Viruses of Fish: An Overview of Significant Pathogens" Viruses 3, no. 11: 2025-2046. https://doi.org/10.3390/v3112025
APA StyleCrane, M., & Hyatt, A. (2011). Viruses of Fish: An Overview of Significant Pathogens. Viruses, 3(11), 2025-2046. https://doi.org/10.3390/v3112025