Abstract
Chronic hepatitis C virus (HCV) infection is a major worldwide cause of liver disease, including cirrhosis and hepatocellular carcinoma. It is estimated that more than 170 million individuals are infected with HCV, with three to four million new cases each year. The current standard of care, combination treatment with interferon and ribavirin, eradicates the virus in only about 50% of chronically infected patients. Notably, neither of these drugs directly target HCV. Many new antiviral therapies that specifically target hepatitis C (e.g. NS3 protease or NS5B polymerase inhibitors) are therefore in development, with a significant number having advanced into clinical trials. The nonstructural 4B (NS4B) protein, is among the least characterized of the HCV structural and nonstructural proteins and has been subjected to few pharmacological studies. NS4B is an integral membrane protein with at least four predicted transmembrane (TM) domains. A variety of functions have been postulated for NS4B, such as the ability to induce the membranous web replication platform, RNA binding and NTPase activity. This review summarizes potential targets within the nonstructural protein NS4B, with a focus on novel classes of NS4B inhibitors.
Keywords:
HCV; hepatitis C virus; antiviral agents; NS4B; clinical trials; RNA binding; amphipathic helix; NTPase 1. Introduction
Hepatitis C virus (HCV) infects an estimated 170 million people around the world []. Infection often leads to cirrhosis and sometimes to hepatocellular carcinoma [,]. No vaccine is currently available. The only FDA approved regimen relies on a combination of subcutaneously administered pegylated interferon-α (IFN) and oral ribavirin (RBV). These non-HCV specific antivirals, however, have limited efficacy. A sustained virologic response (SVR), defined as the absence of HCV RNA in the serum six months after the cessation of HCV therapy, can be achieved in only about 42% to 53% of patients with HCV genotypes 1 and 4, respectively, and up to 78% to 82% of patients infected with HCV genotypes 2 or 3 [,]. These genotype specific differences are hypothesized to be due in part to the evolution of anti-immune factors [], which limit the immune response generated by exogenous interferon. Furthermore, because the majority of patients infected with HCV in the United States are infected with genotype 1, the most treatment-refractory genotype, nearly half of all patients treated for HCV with IFN/RBV therapy will fail treatment. IFN/RBV combination therapy is also expensive, and results in serious side effects such as fever, fatigue, anemia, leucopenia, thrombocytopenia, and depression [,,].
For these many reasons, there is an urgent need for HCV-specific antivirals that are both more efficacious and tolerable. Furthermore, given the ability of HCV to rapidly acquire resistance to antivirals, as has been already shown in multiple clinical trials [,,,], anti-HCV therapeutics that target multiple aspects of the HCV-life cycle are expected to be needed in order to control this infection—as is currently the case for HIV. The rapidity with which HCV acquires resistance is due in part to the fact that the HCV NS5B polymerase lacks a proofreading function, which results in an estimated erroneous base substitution rate of ~1 in 10,000 replicated bases []. Practically, this means that every copy of HCV that is made contains at least one mutation. Given the massive number of HCV virions produced in patients chronically infected with HCV (1.3 × 1012 virions per day) [], every patient is therefore best considered as infected with a population of HCV quasi-species [,]. Such quasi-species, when under selective antiviral pressure, lead to the emergence of resistant strains.
Therefore, a collection of well-tolerated HCV antivirals that target multiple steps of the HCV life cycle, and which thereby can overwhelm the ability of such quasi-species to adapt, represents the idealized goal for future HCV drug development. These agents are termed “specifically targeted antiviral therapy for HCV” or STAT-C drugs [,]. Among the most promising new anti-HCV agents in clinical development are those that target the NS3 protease, the NS5A protein, the RNA-polymerase NS5B, and compounds that directly inhibit HCV replication through interaction with host cell proteins (see [20-23] for more in depth reviews of these agents). Another new promising strategy consists of targeting the NS4B protein. This review focuses on the recent developments of potential antivirals against targets within NS4B.
2. Important features of NS4B
NS4B is a relatively poorly characterized 27 kDa protein with at least four predicted transmembrane (TM) domains [24-26] (Figure 1). As a consequence of polyprotein processing by the NS3-4A protease, the N- and C-terminal parts of NS4B are believed to be oriented towards the cytosolic side of the endoplasmic reticulum (ER) membrane. An amphipathic helix (AH), a helix in which one side is hydrophobic and the other hydrophilic, is located within the first 27 amino acids of NS4B and designated AH1 []. When N-terminally fused to a construct containing GFP, AH1 was shown to mediate membrane association, and point mutations disrupting its hydrophobic face abrogated membrane association []. Of note, fusion of amino acids 1 to 29 or 1 to 40 of NS4B did not confer membrane association [], possibly reflecting differences in the respective expression constructs. Insertion of artificial glycosylation acceptor sites at various positions in NS4B yielded evidence in support of the predicted ER luminal loops around amino acid positions 112 and 161 [,]. In addition, a fraction of NS4B may acquire a fifth TM domain, postulated to result from post-translational translocation of the N-terminal domain into the ER lumen (see Figure 1) [].
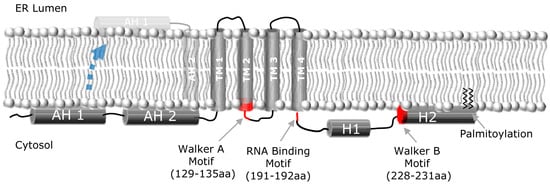
Figure 1.
NS4B membrane topology. Schematic representation of the HCV NS4B protein, and its proposed topology with respect to the ER membrane, depicting the N-terminal amphipathic helices (AH1, AH2), the four transmembrane domains (TM1-4), and the two C-terminal helices (H1, H2). A fraction of NS4B appears to be able to undergo a post-translational translocation event wherein the N-terminus of NS4B adopts a luminal orientation (indicated by dashed blue arrow), thereby creating a fifth transmembrane domain. Walker A and B motifs and RNA binding motif are also indicated.
Recently, we and others have identified a second AH (AH2) immediately following AH1 (Figure 1) that is sufficient to confer tight membrane association and is required for viral replication [,,]. It is suggested that this amphipathic α-helix has the potential to traverse the phospholipid bilayer as a transmembrane domain []. We have shown that AH2 can both oligomerize with itself and specifically mediate lipid vesicle aggregation []—a function that might be relevant for the role of NS4B in forming the membranous web replication platform [] (discussed below). A variety of biophysical measurements have confirmed the predicted helical nature of AH1 [] and AH2 [,].
Secondary structure analyses have also predicted two helices in the C-terminal portion of NS4B. The first of these C-terminal helices (H1) is composed of amino acids ~200–213 and is highly conserved among HCV genotypes. The second helix (H2), also confirmed by CD [], is composed of amino acids ~229–253 and is more variable. H2 also has an amphipathic structure, mediates membrane association, and is involved in the formation of functional HCV replication complexes [26,28,31,32,34-38]. NS4B has also been shown to interact with itself, suggesting an ability to form homo-multimers [,]. Finally, NS4B has been shown to be palmitoylated at two C-terminal cysteine residues []. It is hypothesized that this palmitoylation plays an important role in NS4B oligomerization. However, the role of C-terminal palmitoylation of NS4B in the HCV life cycle remains to be further explored.
One of the more striking NS4B functions identified to date is its reported effect on intracellular membranes. Expression of the HCV NS4B protein alone is sufficient to cause formation of the so-called “membranous web” structure [,] which is thought to represent the platform upon which HCV replication takes places. Like all positive strand RNA viruses, HCV replicates its genome in intimate association with intracellular membranes. The membranous web is believed to be derived in part from the ER. A number of studies have also suggested that the early endosome proteins Rab5 and Rab7 [,], phosphatidylinositol 4-kinase III alpha [] and autophagy protein ATG5 [] may play a role in HCV genome replication or membranous web formation. The membranous web consists of a collection of vesicular-like structures detectable by electron microscopy. Under light microscopy, the membranous web is believed to be correlated with so-called membrane associated foci (MAF) []. Both N- and C-terminal amphipathic helices have been implicated in the formation of such foci [,]. The mechanism(s) by which NS4B induces the membranous web, and which host cell components are hijacked for this purpose are still largely unknown.
Site-directed mutagenesis in the replicon and HCVcc systems has demonstrated the essential role of NS4B in HCV RNA replication [,,,,,,]. Jones et al. [] revealed an additional role of NS4B in viral assembly when a single amino acid substitution, N216A, in NS4B was shown to be sufficient to increase the titer of JFH1 virus by five-fold without affecting HCV RNA replication. This highly conserved position, located between helices H1 and H2, might be critical for interaction between NS4B and other components of the viral assembly machinery.
Physical interactions between NS4B and other nonstructural proteins have been demonstrated by glutathione S-transferase pull-down experiments []. In addition, intragenotypic interactions between NS3 and NS4B were demonstrated in the replicon system [], which may underlie recent findings of synergy between pharmacologic inhibitors of NS3 and NS4B (see Section 3.2. below).
Another functionality of NS4B was identified by Einav and colleagues, who discovered a functional nucleotide-binding motif (NBM) within NS4B. This motif was found to mediate GTP hydrolysis and is essential for HCV replication []. Thompson et al. confirmed this finding recently and also demonstrated ATPase and adenylate kinase activity for NS4B [].
Finally, NS4B has been shown to bind the viral RNA, an interaction that is critical for HCV replication []. These aspects of NS4B functionality are discussed further, below.
4. Conclusions
The functions of NS4B identified to date have enabled the development of two new classes of anti-HCV drugs: one class that interferes with the ability of NS4B to bind HCV RNA; and another that inhibits NS4B’s interactions with membranes. Notably, preliminary data suggests that the drugs that target these individual functions of NS4B are synergistic. It is expected that these inhibitors will be used as a basis for future structure-function studies designed to yield more potent derivatives and eventual new clinical candidates. Furthermore, as other functions for NS4B have already been identified, (e.g. its ability to hydrolyze NTPs), it is hoped that these too can be targeted by separate classes of antivirals.
Knowledge of the molecular virology of HCV, and the functions of its encoded proteins, has allowed for the design of new drugs that directly target HCV. Other than safety and efficacy, the major challenges of HCV drug development are the ability to design therapeutics that inhibit all HCV genotypes and yet are insensitive to the emergence of drug mutants. One solution to this latter challenge is to administer a cocktail of antiviral agents that target different functions in the viral cycle. Accordingly, most current clinical trials test a combination of antiviral agents that combine STAT-C agents with IFN or RBV. As more and more STAT-C agents emerge, and more efficacy data is accumulated, future studies that are able to omit or decrease IFN or RBV may be considered—such an approach would hopefully provide even more tolerable and efficacious therapies to fight HCV and its complications.
Acknowledgements
This work was supported by RO1AI087917 and R41AI088793 and a Burroughs Wellcome Fund Clinical Scientist Award in Translational Research (to J.S.G.). Hadas Dvory-Sobol is the recipient of an ALF Postdoctoral Research Fellow Award. P.S. Pang is supported by an NRSA 1F32AI082930 Fellowship.
Disclosures
J.S.G. is a consultant to, or has an equity interest in Eiger BioPharmaceuticals, Inc., Genentech, Merck, Roche, and Romark Laboratories. H.D.-S. and P.S.P. have an equity interest in Eiger BioPharmaceuticals, Inc.
References and Notes
- Global surveillance and control of hepatitis, C. Report of a WHO Consultation organized in collaboration with the Viral Hepatitis Prevention Board, Antwerp, Belgium. J. Viral. Hepat. 1999, 6, 35–47. [Google Scholar]
- Alter, M.J.; Kruszon-Moran, D.; Nainan, O.V.; McQuillan, G.M.; Gao, F.; Moyer, L.A.; Kaslow, R.A.; Margolis, H.S. The prevalence of hepatitis C virus infection in the United States, 1988 through 1994. N. Engl. J. Med. 1999, 341, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Alter, M.J.; Margolis, H.S.; Krawczynski, K.; Judson, F.N.; Mares, A.; Alexander, W.J.; Hu, P.Y.; Miller, J.K.; Gerber, M.A.; Sampliner, R.E.; et al. The natural history of community-acquired hepatitis C in the United States. The Sentinel Counties Chronic non-A, non-B Hepatitis Study Team. N. Engl. J. Med. 1992, 327, 1899–1905. [Google Scholar] [CrossRef]
- Fried, M.W.; Shiffman, M.L.; Reddy, K.R.; Smith, C.; Marinos, G.; Goncales, F.L., Jr.; Haussinger, D.; Diago, M.; Carosi, G.; Dhumeaux, D.; Craxi, A.; Lin, A.; Hoffman, J.; Yu, J. Peginterferon alfa-2a plus ribavirin for chronic hepatitis C virus infection. N. Engl. J. Med. 2002, 347, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.P.; McHutchison, J.G.; Gordon, S.C.; Rustgi, V.K.; Shiffman, M.; Reindollar, R.; Goodman, Z.D.; Koury, K.; Ling, M.; Albrecht, J.K. Peginterferon alfa-2b plus ribavirin compared with interferon alfa-2b plus ribavirin for initial treatment of chronic hepatitis C: a randomised trial. Lancet 2001, 358, 958–965. [Google Scholar] [CrossRef]
- Pang, P.S.; Planet, P.J.; Glenn, J.S. The evolution of the major hepatitis C genotypes correlates with clinical response to interferon therapy. PLoS One 2009, 4, e6579. [Google Scholar] [CrossRef] [PubMed]
- Di Bisceglie, A.M.; McHutchison, J.; Rice, C.M. New therapeutic strategies for hepatitis C. Hepatology 2002, 35, 224–231. [Google Scholar] [CrossRef]
- Tan, S.L.; Pause, A.; Shi, Y.; Sonenberg, N. Hepatitis C therapeutics: current status and emerging strategies. Nat. Rev. Drug Discov. 2002, 1, 867–881. [Google Scholar] [CrossRef]
- Manns, M.P.; Wedemeyer, H.; Cornberg, M. Treating viral hepatitis C: efficacy, side effects, and complications. Gut 2006, 55, 1350–1359. [Google Scholar] [CrossRef]
- Sarrazin, C.; Kieffer, T.L.; Bartels, D.; Hanzelka, B.; Muh, U.; Welker, M.; Wincheringer, D.; Zhou, Y.; Chu, H.M.; Lin, C.; Weegink, C.; Reesink, H.; Zeuzem, S.; Kwong, A.D. Dynamic hepatitis C virus genotypic and phenotypic changes in patients treated with the protease inhibitor telaprevir. Gastroenterology 2007, 132, 1767–1777. [Google Scholar] [CrossRef]
- Kieffer, T.L.; Sarrazin, C.; Miller, J.S.; Welker, M.W.; Forestier, N.; Reesink, H.W.; Kwong, A.D.; Zeuzem, S. Telaprevir and pegylated interferon-alpha-2a inhibit wild-type and resistant genotype 1 hepatitis C virus replication in patients. Hepatology 2007, 46, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Susser, S.; Welsch, C.; Wang, Y.; Zettler, M.; Domingues, F.S.; Karey, U.; Hughes, E.; Ralston, R.; Tong, X.; Herrmann, E.; Zeuzem, S.; Sarrazin, C. Characterization of resistance to the protease inhibitor boceprevir in hepatitis C virus-infected patients. Hepatology 2009, 50, 1709–1718. [Google Scholar] [CrossRef]
- Sarrazin, C.; Zeuzem, S. Resistance to direct antiviral agents in patients with hepatitis C virus infection. Gastroenterology 2010, 138, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Bartenschlager, R.; Lohmann, V. Replication of hepatitis C virus. J. Gen. Virol. 2000, 81, 1631–1648. [Google Scholar]
- Neumann, A.U.; Lam, N.P.; Dahari, H.; Gretch, D.R.; Wiley, T.E.; Layden, T.J.; Perelson, A.S. Hepatitis C viral dynamics in vivo and the antiviral efficacy of interferon-alpha therapy. Science 1998, 282, 103–107. [Google Scholar] [CrossRef]
- Domingo, E.; Martinez-Salas, E.; Sobrino, F.; de la Torre, J.C.; Portela, A.; Ortin, J.; Lopez-Galindez, C.; Perez-Brena, P.; Villanueva, N.; Najera, R.; et al. The quasispecies (extremely heterogeneous) nature of viral RNA genome populations: biological relevance--a review. Gene 1985, 40, 1–8. [Google Scholar] [CrossRef]
- Martell, M.; Esteban, J.I.; Quer, J.; Genesca, J.; Weiner, A.; Esteban, R.; Guardia, J.; Gomez, J. Hepatitis C virus (HCV) circulates as a population of different but closely related genomes: quasispecies nature of HCV genome distribution. J. Virol. 1992, 66, 3225–3229. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.P.; Foster, G.R.; Rockstroh, J.K.; Zeuzem, S.; Zoulim, F.; Houghton, M. The way forward in HCV treatment--finding the right path. Nat. Rev. Drug Discov. 2007, 6, 991–1000. [Google Scholar] [CrossRef]
- Goto, K.; Watashi, K.; Murata, T.; Hishiki, T.; Hijikata, M.; Shimotohno, K. Evaluation of the anti-hepatitis C virus effects of cyclophilin inhibitors, cyclosporin A, and NIM811. Biochem. Biophys. Res. Commun. 2006, 343, 879–884. [Google Scholar] [CrossRef]
- Chary, A.; Holodniy, M. Recent advances in hepatitis C virus treatment: review of HCV protease inhibitor clinical trials. Rev. Recent. Clin. Trials 2010, 5, 158–173. [Google Scholar] [CrossRef]
- Chen, K.X.; Njoroge, F.G. A review of HCV protease inhibitors. Curr. Opin. Investig. Drugs 2009, 10, 821–837. [Google Scholar] [PubMed]
- Burton, J.R., Jr.; Everson, G.T. HCV NS5B polymerase inhibitors. Clin. Liver Dis. 2009, 13, 453–465. [Google Scholar] [CrossRef]
- Gelman, M.A. , Glenn, J.S. Mixing the right hepatitis C inhibitor cocktail. Trends Mol. Med. 2010, in press. [Google Scholar]
- Lundin, M.; Monne, M.; Widell, A.; Von Heijne, G.; Persson, M.A. Topology of the membrane-associated hepatitis C virus protein NS4B. J. Virol. 2003, 77, 5428–5438. [Google Scholar] [CrossRef]
- Hugle, T.; Fehrmann, F.; Bieck, E.; Kohara, M.; Krausslich, H.G.; Rice, C.M.; Blum, H.E.; Moradpour, D. The hepatitis C virus nonstructural protein 4B is an integral endoplasmic reticulum membrane protein. Virology 2001, 284, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Welsch, C.; Albrecht, M.; Maydt, J.; Herrmann, E.; Welker, M.W.; Sarrazin, C.; Scheidig, A.; Lengauer, T.; Zeuzem, S. Structural and functional comparison of the non-structural protein 4B in flaviviridae. J. Mol. Graph. Model 2007, 26, 546–557. [Google Scholar] [CrossRef]
- Elazar, M.; Liu, P.; Rice, C.M.; Glenn, J.S. An N-terminal amphipathic helix in hepatitis C virus (HCV) NS4B mediates membrane association, correct localization of replication complex proteins, and HCV RNA replication. J. Virol. 2004, 78, 11393–11400. [Google Scholar] [CrossRef]
- Gouttenoire, J.; Castet, V.; Montserret, R.; Arora, N.; Raussens, V.; Ruysschaert, J.M.; Diesis, E.; Blum, H.E.; Penin, F.; Moradpour, D. Identification of a novel determinant for membrane association in hepatitis C virus nonstructural protein 4B. J. Virol. 2009, 83, 6257–6268. [Google Scholar] [CrossRef]
- Lundin, M.; Lindstrom, H.; Gronwall, C.; Persson, M.A. Dual topology of the processed hepatitis C virus protein NS4B is influenced by the NS5A protein. J. Gen. Virol. 2006, 87, 3263–3272. [Google Scholar] [CrossRef]
- Cho, N.J.; Dvory-Sobol, H.; Lee, C.; Cho, S.J.; Bryson, P.; Masek, M.; Elazar, M.; Frank, C.W.; Glenn, J.S. Identification of a class of HCV inhibitors directed against the nonstructural protein NS4B. Sci. Transl. Med. 2010, 2, 15ra16. [Google Scholar] [CrossRef]
- Gouttenoire, J.; Montserret, R.; Kennel, A.; Penin, F.; Moradpour, D. An amphipathic alpha-helix at the C terminus of hepatitis C virus nonstructural protein 4B mediates membrane association. J. Virol. 2009, 83, 11378–11384. [Google Scholar] [CrossRef] [PubMed]
- Egger, D.; Wolk, B.; Gosert, R.; Bianchi, L.; Blum, H.E.; Moradpour, D.; Bienz, K. Expression of hepatitis C virus proteins induces distinct membrane alterations including a candidate viral replication complex. J. Virol. 2002, 76, 5974–5984. [Google Scholar] [CrossRef] [PubMed]
- Cho, N.J. Stanford University, Stanford, CA, USA. Unpublished work, 2010.
- Aligo, J.; Jia, S.; Manna, D.; Konan, K.V. Formation and function of hepatitis C virus replication complexes require residues in the carboxy-terminal domain of NS4B protein. Virology 2009, 393, 68–83. [Google Scholar] [CrossRef]
- Guillen, J.; Gonzalez-Alvarez, A.; Villalain, J. A membranotropic region in the C-terminal domain of hepatitis C virus protein NS4B interaction with membranes. Biochim. Biophys. Acta. 2010, 1798, 327–337. [Google Scholar] [CrossRef]
- Liefhebber, J.M.; Brandt, B.W.; Broer, R.; Spaan, W.J.; van Leeuwen, H.C. Hepatitis C virus NS4B carboxy terminal domain is a membrane binding domain. Virol. J. 2009, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Gosert, R.; Moradpour, D. Reading between the lines: evidence for a new hepatitis C virus protein. Hepatology 2002, 36, 757–760. [Google Scholar] [CrossRef]
- Jones, D.M.; Patel, A.H.; Targett-Adams, P.; McLauchlan, J. The hepatitis C virus NS4B protein can trans-complement viral RNA replication and modulates production of infectious virus. J. Virol. 2009, 83, 2163–2177. [Google Scholar] [CrossRef]
- Dimitrova, M.; Imbert, I.; Kieny, M.P.; Schuster, C. Protein-protein interactions between hepatitis C virus nonstructural proteins. J. Virol. 2003, 77, 5401–5414. [Google Scholar] [CrossRef]
- Yu, G.Y.; Lee, K.J.; Gao, L.; Lai, M.M. Palmitoylation and polymerization of hepatitis C virus NS4B protein. J. Virol. 2006, 80, 6013–6023. [Google Scholar] [CrossRef]
- Stone, M.; Jia, S.; Heo, W.D.; Meyer, T.; Konan, K.V. Participation of rab5, an early endosome protein, in hepatitis C virus RNA replication machinery. J. Virol. 2007, 81, 4551–4563. [Google Scholar] [CrossRef]
- Manna, D.; Aligo, J.; Xu, C.; Park, W.S.; Koc, H.; Do Heo, W.; Konan, K.V. Endocytic Rab proteins are required for hepatitis C virus replication complex formation. Virology 2010, 398, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Berger, K.L.; Cooper, J.D.; Heaton, N.S.; Yoon, R.; Oakland, T.E.; Jordan, T.X.; Mateu, G.; Grakoui, A.; Randall, G. Roles for endocytic trafficking and phosphatidylinositol 4-kinase III alpha in hepatitis C virus replication. Proc. Natl. Acad. Sci. U S A 2009, 106, 7577–7582. [Google Scholar] [CrossRef] [PubMed]
- Guevin, C.; Manna, D.; Belanger, C.; Konan, K.V.; Mak, P.; Labonte, P. Autophagy protein ATG5 interacts transiently with the hepatitis C virus RNA polymerase (NS5B) early during infection. Virology 2010, 405, 1–7. [Google Scholar] [CrossRef]
- Gretton, S.N.; Taylor, A.I.; McLauchlan, J. Mobility of the hepatitis C virus NS4B protein on the endoplasmic reticulum membrane and membrane-associated foci. J. Gen. Virol. 2005, 86, 1415–1421. [Google Scholar] [CrossRef]
- Lindstrom, H.; Lundin, M.; Haggstrom, S.; Persson, M.A. Mutations of the Hepatitis C virus protein NS4B on either side of the ER membrane affect the efficiency of subgenomic replicons. Virus Res. 2006, 121, 169–178. [Google Scholar] [CrossRef]
- Blight, K.J. Allelic variation in the hepatitis C virus NS4B protein dramatically influences RNA replication. J. Virol. 2007, 81, 5724–5736. [Google Scholar] [CrossRef]
- Paredes, A.M.; Blight, K.J. A genetic interaction between hepatitis C virus NS4B and NS3 is important for RNA replication. J. Virol. 2008, 82, 10671–10683. [Google Scholar] [CrossRef]
- Einav, S.; Elazar, M.; Danieli, T.; Glenn, J.S. A nucleotide binding motif in hepatitis C virus (HCV) NS4B mediates HCV RNA replication. J. Virol. 2004, 78, 11288–11295. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.A.; Zou, A.; Yan, J.; Duggal, R.; Hao, W.; Molina, D.; Cronin, C.N.; Wells, P.A. Biochemical characterization of recombinant hepatitis C virus nonstructural protein 4B: evidence for ATP/GTP hydrolysis and adenylate kinase activity. Biochemistry 2009, 48, 906–916. [Google Scholar] [CrossRef]
- Einav, S.; Gerber, D.; Bryson, P.D.; Sklan, E.H.; Elazar, M.; Maerkl, S.J.; Glenn, J.S.; Quake, S.R. Discovery of a hepatitis C target and its pharmacological inhibitors by microfluidic affinity analysis. Nat. Biotechnol. 2008, 26, 1019–1027. [Google Scholar] [CrossRef]
- Einav, S.; Sklan, E.H.; Moon, H.M.; Gehrig, E.; Liu, P.; Hao, Y.; Lowe, A.W.; Glenn, J.S. The nucleotide binding motif of hepatitis C virus NS4B can mediate cellular transformation and tumor formation without Ha-ras co-transfection. Hepatology 2008, 47, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, V.; Hoffmann, S.; Herian, U.; Penin, F.; Bartenschlager, R. Viral and cellular determinants of hepatitis C virus RNA replication in cell culture. J. Virol. 2003, 77, 3007–3019. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.M. Stanford University, Stanford, CA, USA. Unpublished work, 2010.
- Maerkl, S.J.; Quake, S.R. A systems approach to measuring the binding energy landscapes of transcription factors. Science 2007, 315, 233–237. [Google Scholar] [CrossRef]
- Einav, S.; Sobol, H.D.; Gehrig, E.; Glenn, J.S. The hepatitis C virus (HCV) NS4B RNA binding inhibitor clemizole is highly synergistic with HCV protease inhibitors. J. Infect. Dis. 202, 65–67.
- Jacques, A.A.; Fuchs, V.H. Clinical evaluation of clemizole in allergic rhinitis. Int. Rec. Med. 1960, 173, 88–91. [Google Scholar] [PubMed]
- Dvory-Sobol, H. Stanford University, Stanford, CA, USA. Unpublished work, 2010.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).