
Fibrin Hydrogel Based Bone Substitute Tethered with BMP-2 and BMP-2/7 Heterodimers



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
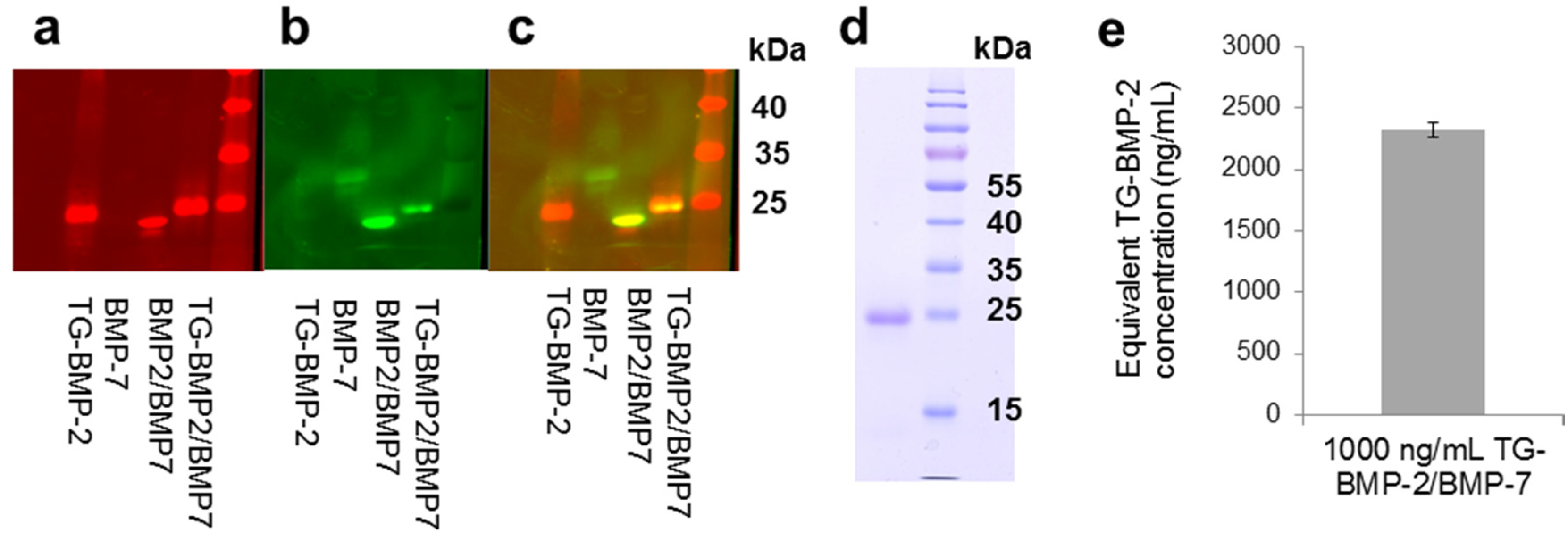
2.1. Heterodimer
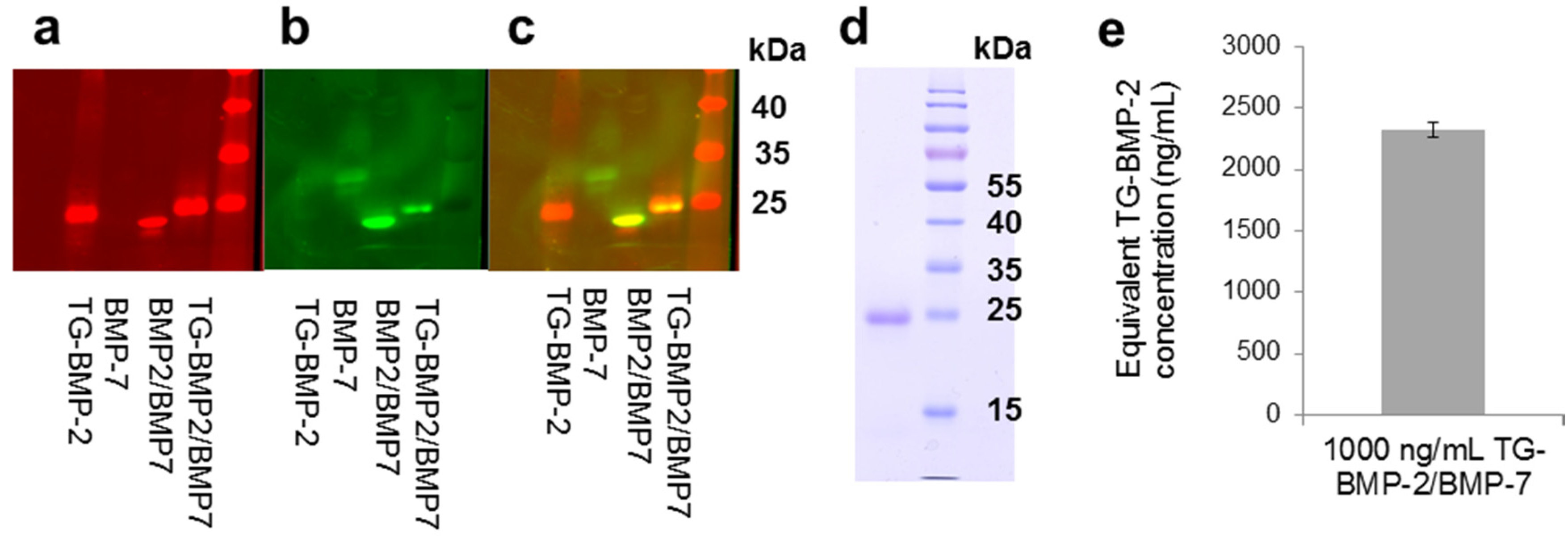
2.2. Functionality of Engineered Growth Factors


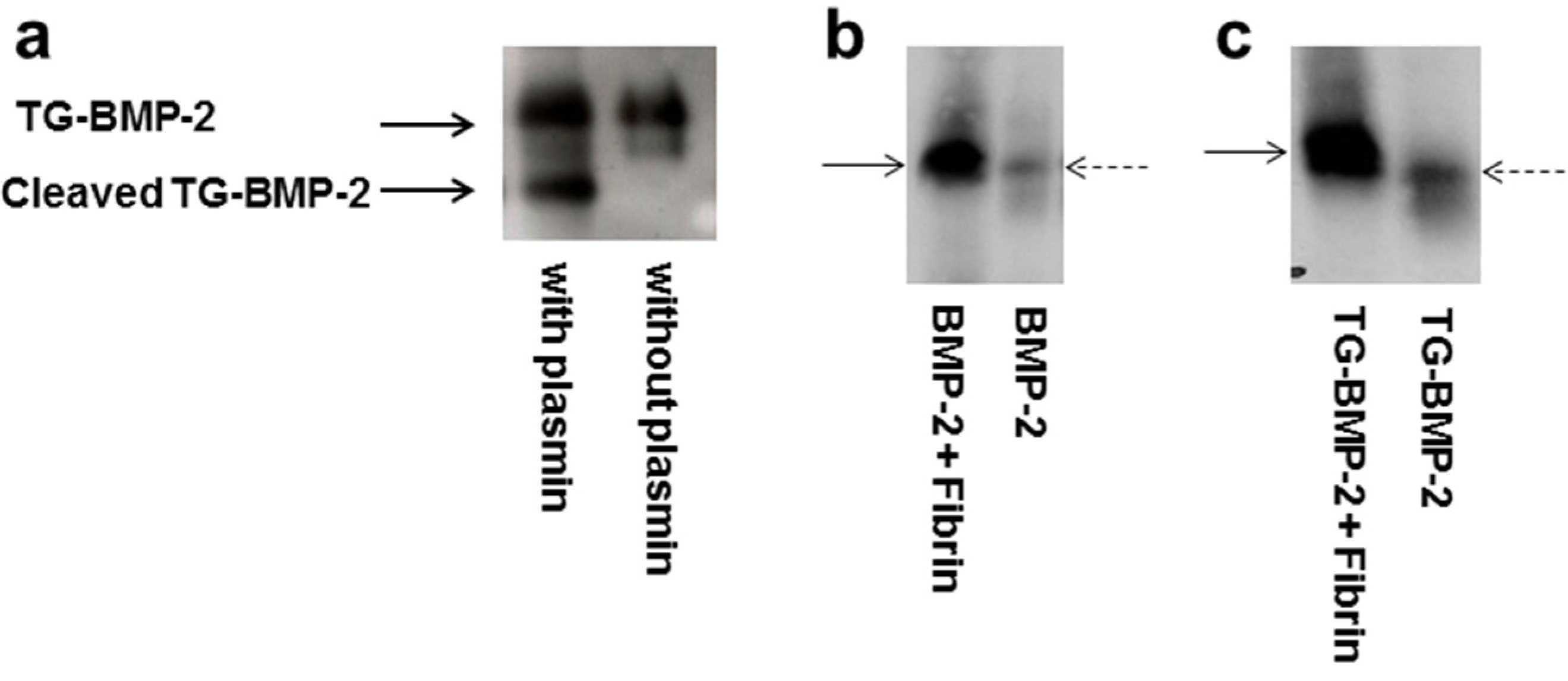
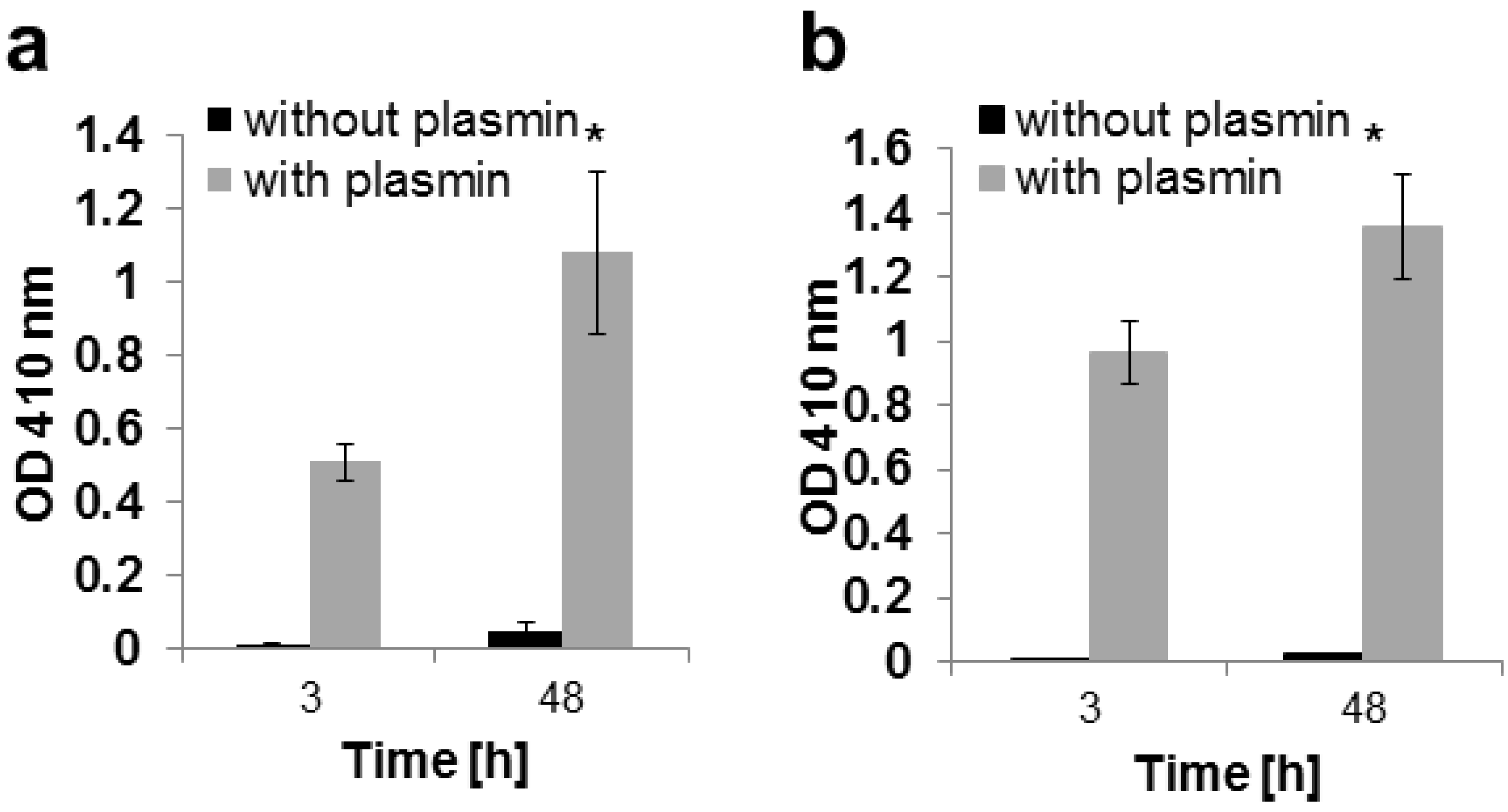
2.3. In Vitro Release

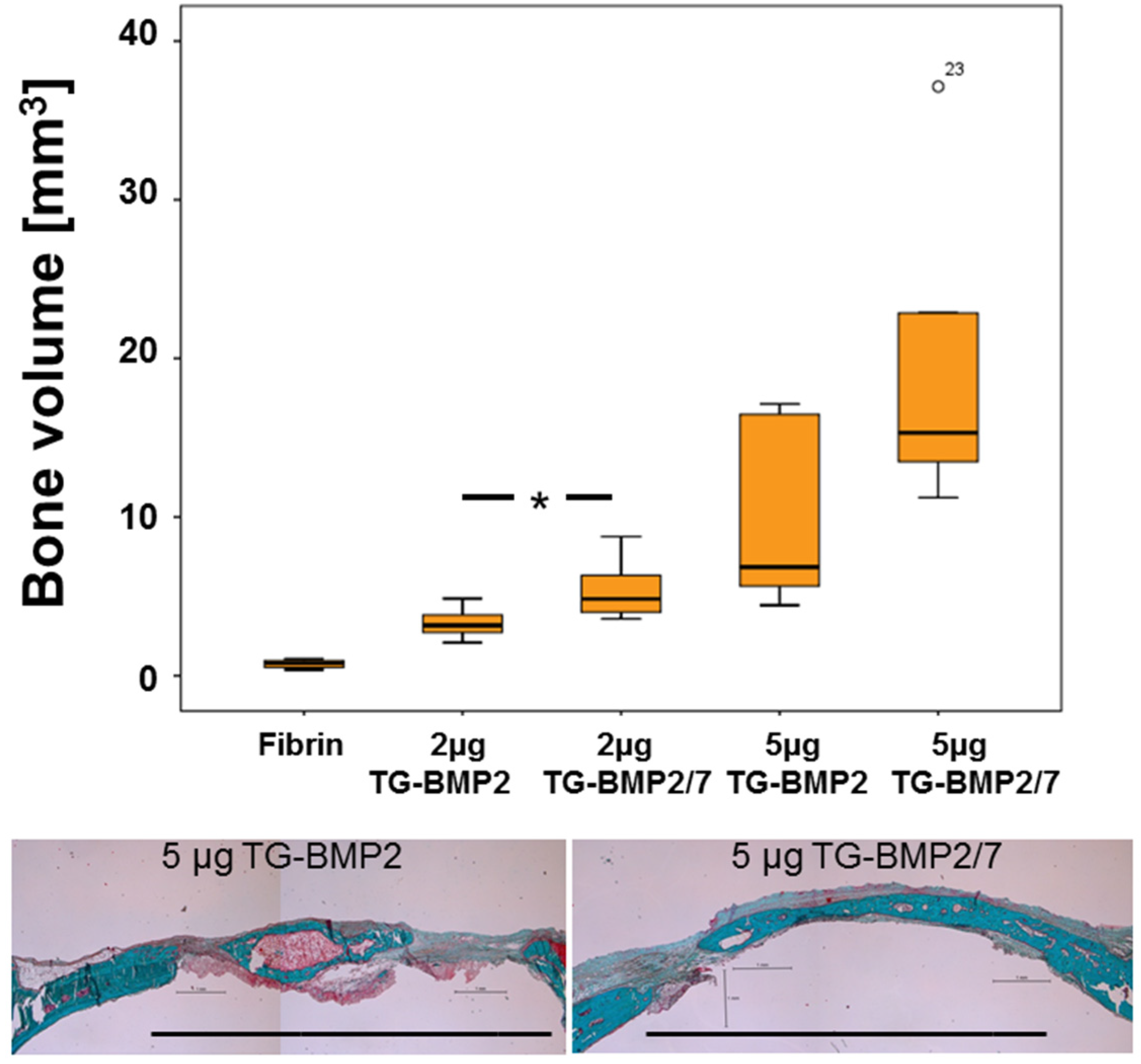
2.4. In Vivo Testing
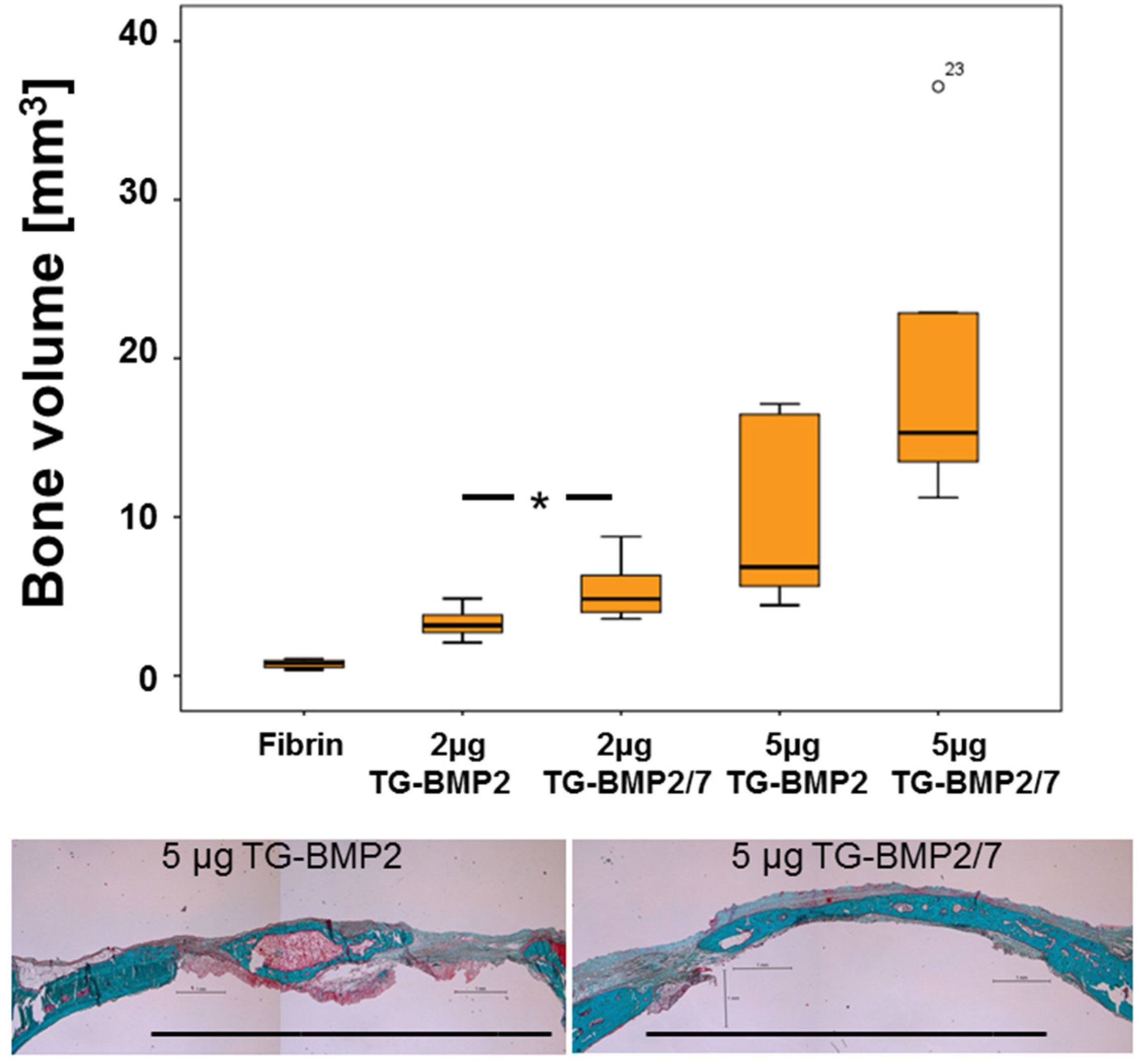

3. Discussion
4. Experimental Section
4.1. BMP-2, TG-BMP-2, and TG-BMP-2/BMP-7 Production
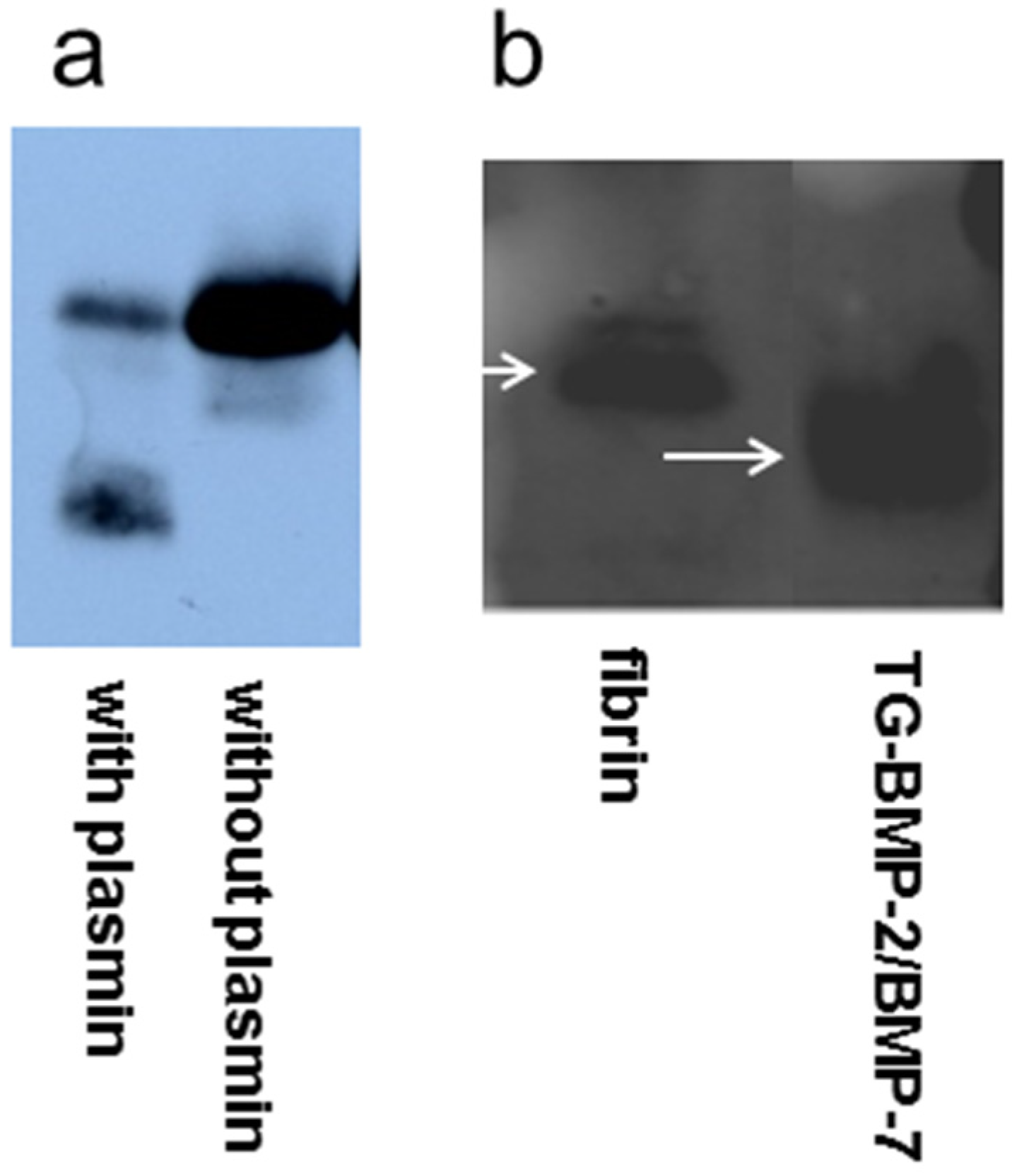
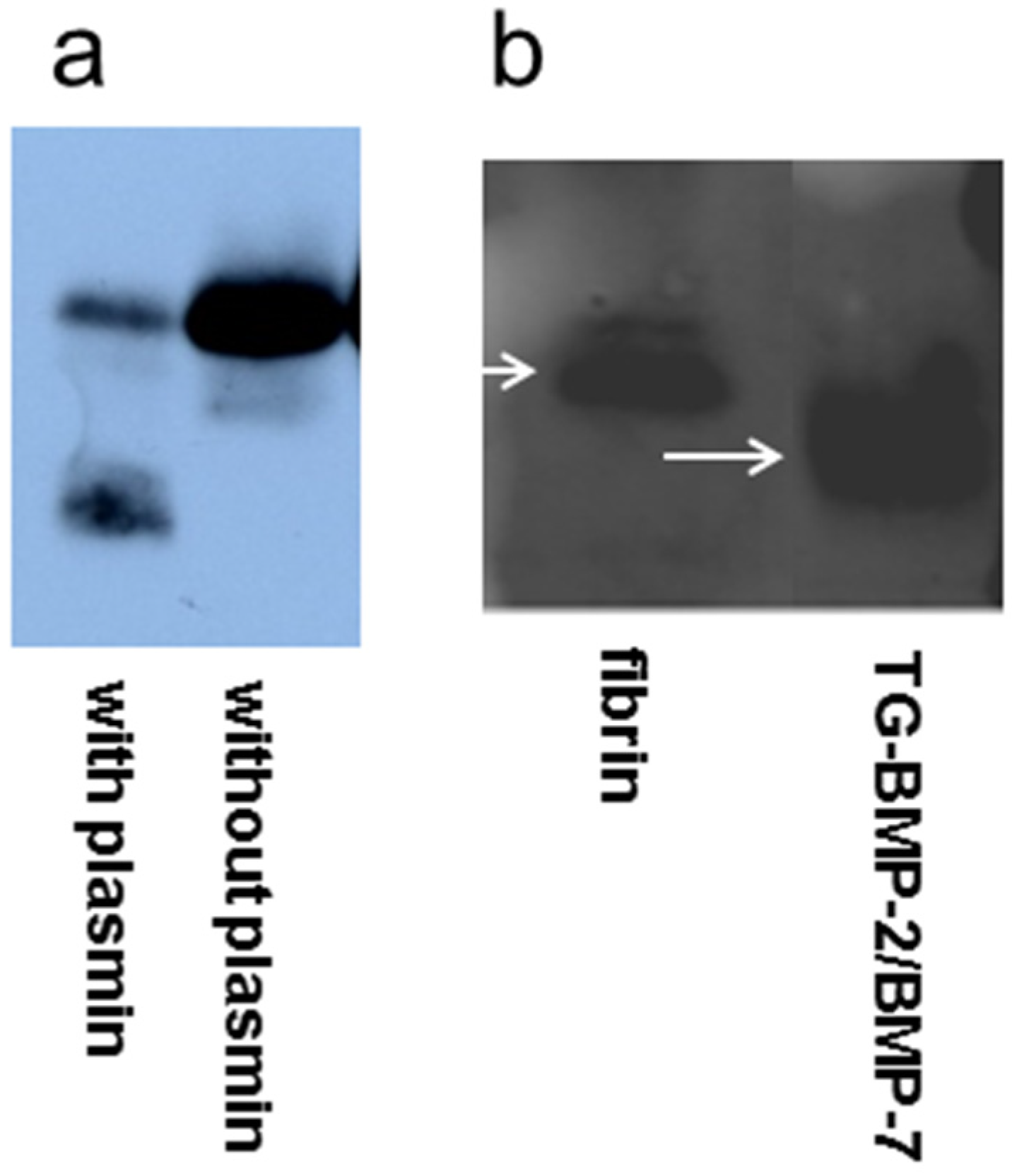
4.2. Plasmin Degradation
4.3. BMP-2 Western Blot
4.4. TG-BMP-2/BMP-7 Western Blot
4.5. Fibrin Gel Formation
4.6. Trypsin Degradation
4.7. Cell Culture
4.8. Alkaline Phosphatase Assay
4.9. In vivo Rat Calvaria Defect
4.10. Microcomputed Tomography (µCT)
4.11. Histology
4.12. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Urist, M.R. Bon: Formation by autoinduction. Science 1965, 150, 893–899. [Google Scholar]
- Ripamonti, U.; Reddi, A.H. Tissue engineering, morphogenesis, and regeneration of the periodontal tissues by bone morphogenetic proteins. Crit. Rev. Oral Biol. Med. 1997, 8, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Elangovan, S.; Srinivasan, S.; Ayilavarapu, S. Novel regenerative strategies to enhance periodontal therapy outcome. Expert Opin. Biol. Ther. 2009, 9, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Seeherman, H.; Wozney, J.M. Delivery of bone morphogenetic proteins for orthopedic tissue regeneration. Cytokine Growth Factor Rev. 2005, 16, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Bessa, P.C.; Casal, M.; Reis, R.L. Bone morphogenetic proteins in tissue engineering: The road from laboratory to clinic, part II (BMP delivery). J. Tissue Eng. Regen. Med. 2008, 2, 81–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Kim, J.; Cheng, C.; Rawlins, B.A.; Boachie-Adjei, O.; Crystal, R.G.; Hidaka, C. Noggin regulation of bone morphogenetic protein (BMP) 2/7 heterodimer activity in vitro. Bone 2006, 39, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Carragee, E.J.; Hurwitz, E.L.; Weiner, B.K. A critical review of recombinant human bone morphogenetic protein-2 trials in spinal surgery: emerging safety concerns and lessons learned. Spine J. 2011, 11, 471–491. [Google Scholar] [CrossRef] [PubMed]
- Haidar, Z.S.; Hamdy, R.C.; Tabrizian, M. Delivery of recombinant bone morphogenetic proteins for bone regeneration and repair. Part A: Current challenges in BMP delivery. Biotechnol. Lett. 2009, 31, 1817–1824. [Google Scholar] [CrossRef] [PubMed]
- Haidar, Z.S.; Hamdy, R.C.; Tabrizian, M. Delivery of recombinant bone morphogenetic proteins for bone regeneration and repair. Part B: Delivery systems for BMPs in orthopaedic and craniofacial tissue engineering. Biotechnol. Lett. 2009, 31, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Luginbuehl, V.; Meinel, L.; Merkle, H.P.; Gander, B. Localized delivery of growth factors for bone repair. Eur. J. Pharm. Biopharm. 2004, 58, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Masters, K.S. Covalent growth factor immobilization strategies for tissue repair and regeneration. Macromol. Biosci. 2011, 11, 1149–1163. [Google Scholar] [CrossRef] [PubMed]
- Martino, M.M.; Tortelli, F.; Mochizuki, M.; Traub, S.; Ben-David, D.; Kuhn, G.A.; Mueller, R.; Livne, E.; Eming, S.A.; Hubbell, J.A. Engineering the growth factor microenvironment with fibronectin domains to promote wound and bone tissue healing. Sci. Transl. Med. 2011, 3, 100ra89. [Google Scholar] [CrossRef] [PubMed]
- Lutolf, M.P.; Hubbell, J.A. Synthetic biomaterials as instructive extracellular microenvironments for morphogenesis in tissue engineering. Nat. Biotechnol. 2005, 23, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Schmoekel, H.G.; Weber, F.E.; Hurter, K.; Schense, J.C.; Seiler, G.; Rytz, U.; Spreng, D.; Schawalder, P.; Hubbell, J. Enhancement of bone healing using non-glycosylated rhBMP-2 released from a fibrin matrix in dogs and cats. J. Small Anim. Pract. 2005, 46, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Macri, L.; Silverstein, D.; Clark, R.A. Growth factor binding to the pericellular matrix and its importance in tissue engineering. Adv. Drug Deliv. Rev. 2007, 59, 1366–1381. [Google Scholar] [CrossRef] [PubMed]
- Schultz, G.S.; Wysocki, A. Interactions between extracellular matrix and growth factors in wound healing. Wound Repair Regen. 2009, 17, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Martino, M.M.; Briquez, P.S.; Ranga, A.; Lutolf, M.P.; Hubbell, J.A. Heparin-binding domain of fibrin(ogen) binds growth factors and promotes tissue repair when incorporated within a synthetic matrix. Proc. Natl. Acad. Sci. USA 2013, 110, 4563–4568. [Google Scholar] [CrossRef] [PubMed]
- Mosesson, M.W. Fibrinogen and fibrin structure and functions. J. Thromb. Haemost. 2005, 3, 1894–1904. [Google Scholar] [CrossRef] [PubMed]
- Jung, R.E.; Schmoekel, H.G.; Zwahlen, R.; Kokovic, V.; Hammerle, C.H.F.; Weber, F.E. Platelet-rich plasma and fibrin as delivery systems for recombinant human bone morphogenetic protein-2. Clin. Oral Implant. Res. 2005, 16, 676–682. [Google Scholar] [CrossRef]
- Schmoekel, H.; Schense, J.C.; Weber, F.E.; Gratz, K.W.; Gnagi, D.; Muller, R.; Hubbell, J.A. Bone healing in the rat and dog with nonglycosylated BMP-2 demonstrating low solubility in fibrin matrices. J. Orthop. Res. 2004, 22, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Schmokel, H.G.; Weber, F.E.; Seiler, G.; von Rechenberg, B.; Schense, J.C.; Schawalder, P.; Hubbell, J. Treatment of nonunions with nonglycosylated recombinant human bone morphogenetic protein-2 delivered from a fibrin matrix. Vet. Surg. 2004, 33, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Schmoekel, H.G.; Weber, F.E.; Schense, J.C.; Gratz, K.W.; Schawalder, P.; Hubbell, J.A. Bone repair with a form of BMP-2 engineered for incorporation into fibrin cell ingrowth matrices. Biotechnol. Bioeng. 2005, 89, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Ehrbar, M.; Rizzi, S.C.; Hlushchuk, R.; Djonov, V.; Zisch, A.H.; Hubbell, J.A.; Weber, F.E.; Lutolf, M.P. Enzymatic formation of modular cell-instructive fibrin analogs for tissue engineering. Biomaterials 2007, 28, 3856–3866. [Google Scholar] [CrossRef] [PubMed]
- Ehrbar, M.; Rizzi, S.C.; Schoenmakers, R.G.; San Miguel, B.; Hubbell, J.A.; Weber, F.E.; Lutolf, M.P. Biomolecular hydrogels formed and degraded via site-specific enzymatic reactions. Biomacromolecules 2007, 8, 3000–3007. [Google Scholar] [CrossRef] [PubMed]
- Sakiyama-Elbert, S.E.; Panitch, A.; Hubbell, J.A. Development of growth factor fusion proteins for cell-triggered drug delivery. FASEB J. 2001, 15, 1300–1302. [Google Scholar] [PubMed]
- Zisch, A.H.; Schenk, U.; Schense, J.C.; Sakiyama-Elbert, S.E.; Hubbell, J.A. Covalently conjugated VEGF-fibrin matrices for endothelialization. J. Control. Release 2001, 72, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Romanic, A.M.; Arleth, A.J.; Willette, R.N.; Ohlstein, E.H. Factor XIIIa cross-links lipoprotein(a) with fibrinogen and is present in human atherosclerotic lesions. Circ. Res. 1998, 83, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Schense, J.C.; Hubbell, J.A. Cross-linking exogenous bifunctional peptides into fibrin gels with factor XIIIa. Bioconjugate Chem. 1999, 10, 75–81. [Google Scholar] [CrossRef]
- Sakiyama-Elbert, S.E.; Hubbell, J.A. Controlled release of nerve growth factor from a heparin-containing fibrin-based cell ingrowth matrix. J. Control. Release 2000, 69, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Sakiyama-Elbert, S.E.; Hubbell, J.A. Development of fibrin derivatives for controlled release of heparin-binding growth factors. J. Control. Release 2000, 65, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Darby, I. Periodontal materials. Aust. Dent. J. 2011, 56, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Wikesjo, U.M.; Qahash, M.; Huang, Y.H.; Xiropaidis, A.; Polimeni, G.; Susin, C. Bone morphogenetic proteins for periodontal and alveolar indications; biological observations-clinical implications. Orthod. Craniofac. Res. 2009, 12, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Zhao, Z.; Koh, J.T.; Jin, T.; Franceschi, R.T. Combinatorial gene therapy for bone regeneration: Cooperative interactions between adenovirus vectors expressing bone morphogenetic proteins 2, 4, and 7. J. Cell. Biochem. 2005, 95, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Wang, J.; Zheng, Y.; Fan, Y.; Gu, Z. BMP2/7 heterodimer is a stronger inducer of bone regeneration in peri-implant bone defects model than BMP2 or BMP7 homodimer. Dental Mater. J. 2012, 31, 239–248. [Google Scholar] [CrossRef]
- Israel, D.I.; Nove, J.; Kerns, K.M.; Kaufman, R.J.; Rosen, V.; Cox, K.A.; Wozney, J.M. Heterodimeric bone morphogenetic proteins show enhanced activity in vitro and in vivo. Growth Factors 1996, 13, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wu, G.; Zhao, J.; Wang, L.; Sun, P.; Gu, Z. Rhbmp2/7 heterodimer: An osteoblastogenesis inducer of not higher potency but lower effective concentration compared with rhBMP2 and rhBMP7 Homodimers. Tissue Eng. Part A 2010, 16, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Valera, E.; Isaacs, M.J.; Kawakami, Y.; Izpisua Belmonte, J.C.; Choe, S. BMP-2/6 heterodimer is more effective than BMP-2 or BMP-6 homodimers as inductor of differentiation of human embryonic stem cells. PLoS One 2010, 5, e11167. [Google Scholar] [CrossRef] [PubMed]
- Aono, A.; Hazama, M.; Notoya, K.; Taketomi, S.; Yamasaki, H.; Tsukuda, R.; Sasaki, S.; Fujisawa, Y. Potent ectopic bone-inducing activity of bone morphogenetic protein-4/7 heterodimer. Biochem. Biophys. Res. Commun. 1995, 210, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Hazama, M.; Aono, A.; Ueno, N.; Fujisawa, Y. Efficient expression of a heterodimer of bone morphogenetic protein subunits using a baculovirus expression system. Biochem. Biophys. Res. Commun. 1995, 209, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Israel, D.; Wolfman, N.M. Recombinant bone morphogenetic protein heterodimers, compositions and methods of use. U.S. Patent 6,190,880, 23 March 2001. [Google Scholar]
- Weber, F.E.; Eyrich, G.; Gratz, K.W.; Maly, F.E.; Sailer, H.F. Slow and continuous application of human recombinant bone morphogenetic protein via biodegradable poly(lactide-co-glycolide) foamspheres. Int. J. Oral Maxillofac. Surg. 2002, 31, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Hanseler, P.; Jung, U.W.; Jung, R.E.; Choi, K.H.; Cho, K.S.; Hammerle, C.H.; Weber, F.E. Analysis of hydrolyzable polyethylene glycol hydrogels and deproteinized bone mineral as delivery systems for glycosylated and non-glycosylated bone morphogenetic protein-2. Acta Biomater. 2012, 8, 116–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, F.E.; Eyrich, G.; Gratz, K.W.; Thomas, R.M.; Maly, F.E.; Sailer, H.F. Disulfide bridge conformers of mature BMP are inhibitors for heterotopic ossification. Biochem. Biophys. Res. Commun. 2001, 286, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Karfeld-Sulzer, L.S.; Ghayor, C.; Siegenthaler, B.; Gjoksi, B.; Pohjonen, T.H.; Weber, F.E. Comparative study of NMP-preloaded and dip-loaded membranes for guided bone regeneration of rabbit cranial defects. J. Tissue Eng. Regen. Med. 2014. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karfeld-Sulzer, L.S.; Siegenthaler, B.; Ghayor, C.; Weber, F.E. Fibrin Hydrogel Based Bone Substitute Tethered with BMP-2 and BMP-2/7 Heterodimers. Materials 2015, 8, 977-991. https://doi.org/10.3390/ma8030977
Karfeld-Sulzer LS, Siegenthaler B, Ghayor C, Weber FE. Fibrin Hydrogel Based Bone Substitute Tethered with BMP-2 and BMP-2/7 Heterodimers. Materials. 2015; 8(3):977-991. https://doi.org/10.3390/ma8030977
Chicago/Turabian StyleKarfeld-Sulzer, Lindsay S., Barbara Siegenthaler, Chafik Ghayor, and Franz E. Weber. 2015. "Fibrin Hydrogel Based Bone Substitute Tethered with BMP-2 and BMP-2/7 Heterodimers" Materials 8, no. 3: 977-991. https://doi.org/10.3390/ma8030977