
Noise in the Sea and Its Impacts on Marine Organisms

Abstract
:1. Introduction
2. Sounds in the Sea and Their Biological Significances
2.1. Propagation of Sounds in the Sea
2.2. Sources of Sounds in the Sea

{kind=link}
| Types of the Anthropogenic Sound | Frequency | Intensity Level | References |
|---|---|---|---|
| Bottom-founded oil drilling and mining | 4–38 Hz | 119–127 dB re 1 μPa | Richardson et al., 1995 [26] |
| Pile driving | 30–40 Hz | 131–135 dB re 1 μPa | Richardson et al., 1995 [26] |
| Drillship | 20–1000 Hz | 174–185 dB re 1 μPa | Richardson et al., 1995 [26] |
| Semisubmersible drilling vessel | 10–4000 Hz | ~154 dB re 1 μPa | Richardson et al., 1995 [26] |
| Seismic airguns | 100–250 Hz | 240–250 dB re 1 μPa | Richardson et al., 1995 [26] |
| The Acoustic Thermometry of Ocean Climate Project (ATOC) | ~75 Hz | ~195 dB re 1 μPa | Buck, 1995 [24] |
| Navy Sonar | 100–500 Hz | ~215 dB re 1 μPa | Conservation and development problem solving team, University of Maryland, 2000 [12] |
| High Frequency Marine Mammal Monitoring Sonar (HF/M3) | ~3000 Hz | ~220 dB re 1 μPa | Conservation and development problem solving team, University of Maryland, 2000 [12] |
| Supertanker & container ship | 6.8–70 Hz | 180–205 dB re 1 μPa | Richardson et al., 1995 [26]; Gisiner et al., 1998 [25] |
| Medium size ship (ferries) | ~50 Hz | 150–170 dB re 1 μPa | Richardson et al., 1995 [26] |
| Boats (<30 m in length) | <300 Hz | ~175 dB re 1 μPa | Richardson et al., 1995 [26] |
| Small ship (support & supply ship) | 20–1000 Hz | 170–180 dB re 1 μPa | Richardson et al., 1995 [26] |
2.3. Biological Significances of Sound in the Sea
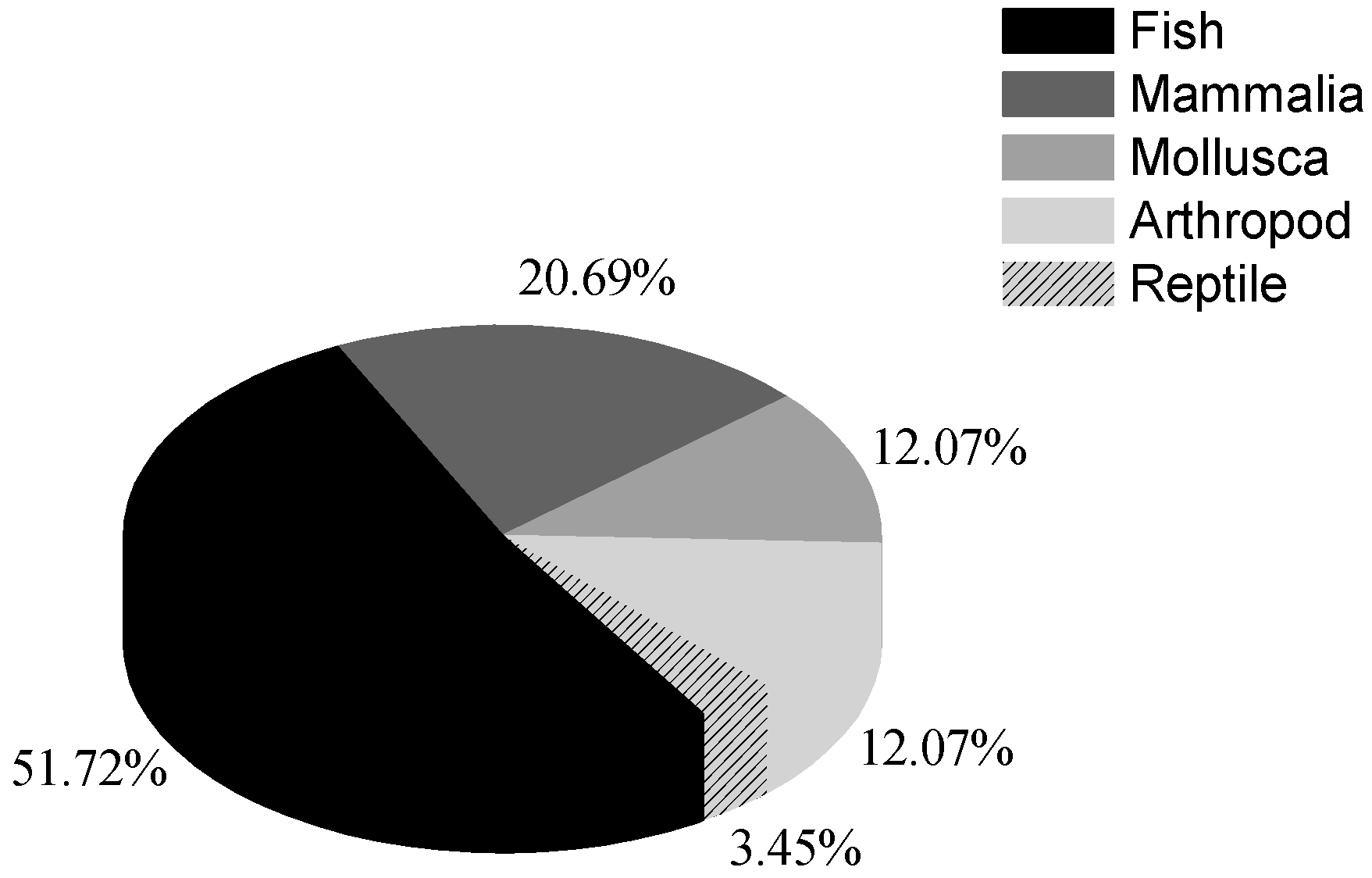
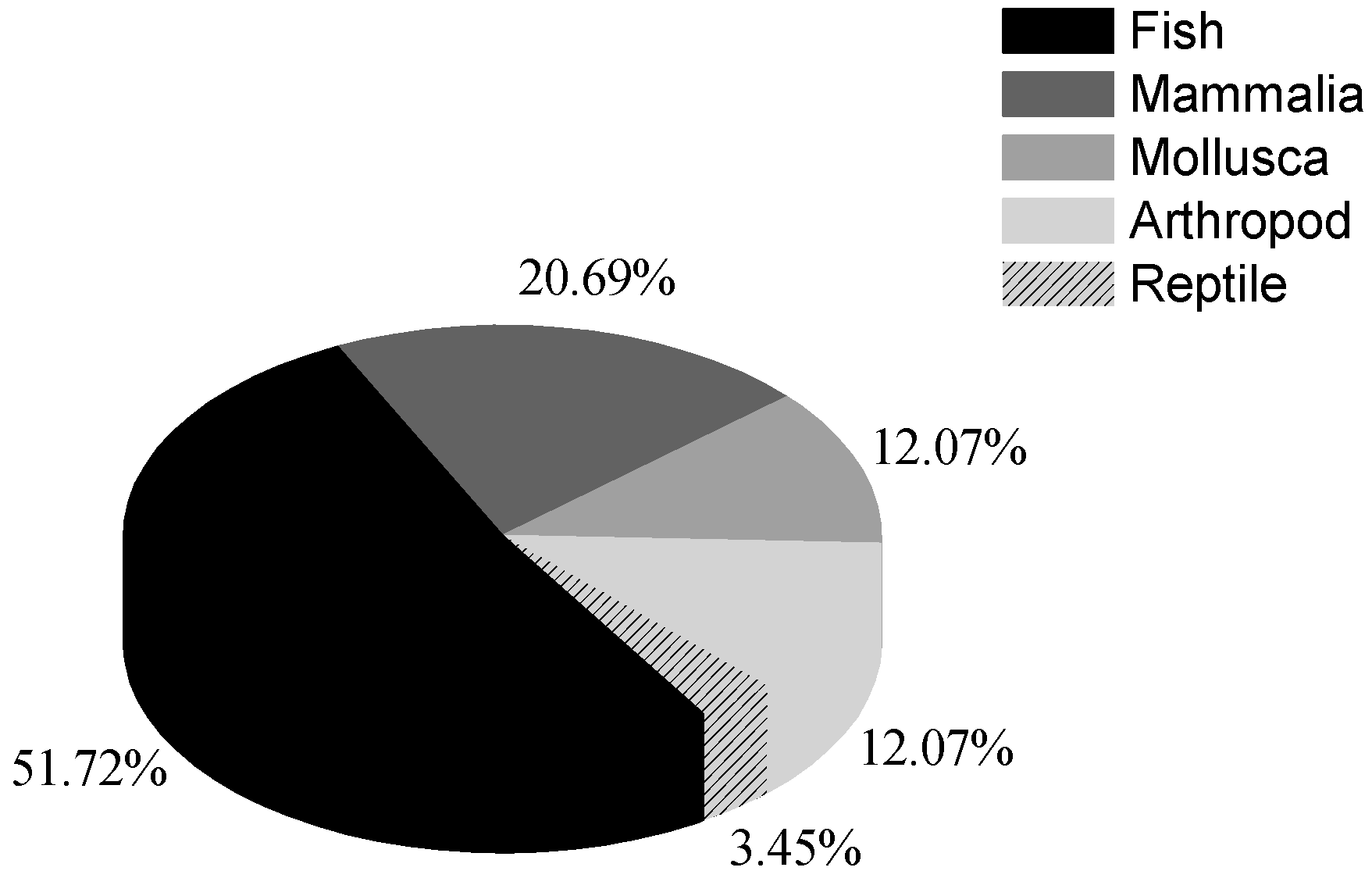
3. The Effects of Anthropogenic Noise on Marine Organisms
3.1. Acoustic Masking and Physiological Damage to Hearing System
| Species | Types of Anthropogenic Noise | Effects | References |
|---|---|---|---|
| M. angustirostris | increased ambient noise | constrains acoustic communication | Southall et al., 2003 [45] |
| C. chromis | boating and shipping noise | reduces auditory sensitivity and shifts the hearing threshold | Codarin et al., 2009 [7] |
| S. umbra | |||
| G. cruentatus | |||
| H. didactylus | boating and shipping noise | constrains acoustic communication and shifts the hearing threshold | Vasconcelos et al., 2007 [46] |
| P. phocoena | seismic air-gun shooting | shifts the hearing threshold | Lucke et al., 2009 [48] |
| T. truncatus | experimental noise emanating device | shifts the hearing threshold | Nachtigall et al., 2004 [49] |
| P. auratus | seismic air-gun shooting | damages the hearing sensory epithelia | McCauley et al., 2003 [37] |
| L. vulgaris | experimental noise emanating device | damages the hearing sensory epithelia | André et al., 2011 [52] |
| S. officinalis | |||
| O. vulgaris | |||
| I. coindetii | |||
| A. dux | seismic air-gun shooting | damage to internal fibers, statocysts, stomachs, and digestive tracts | Guerra et al., 2011 [53] |
3.2. Behavior Alteration
| Species | Types of Anthropogenic Noise | Effects | References |
|---|---|---|---|
| D. labrax | experimental noise emanating device | induces startle response | Kastelein et al., 2008 [6] |
| C. labrosus | |||
| T. luscus | |||
| G. morhua | |||
| P. pollachius | |||
| T. trachurus | |||
| A. Anguilla | |||
| C. harengus | |||
| P. dentex | seismic air-gun shooting | induces startle response | Fewtrell and McCauley, 2012 [54] |
| P. auratus | |||
| S. australis | |||
| C. pallasii | boating and shipping noise | induces avoidance responses | Schwarz and Greer, 1984 [38] |
| N. pulcher | boating and shipping noise | reduces digging and defense capabilities, increases aggression | Bruintjes and Radford, 2013 [58] |
| G. aculeatus | experimental noise emanating device | increases in food-handling error | Purser and Radford, 2011 [59] |
| C. clypeatus | boating and shipping noise | reduces defense capabilities | Chan et al., 2010 [60] |
| C. maenas | boating and shipping noise | reduces defense capabilities | Wale et al., 2013 [61] |
| M. novaeangliae | ATOC (Acoustic Thermometry of Ocean Climate) sound | increases distance and time intervals between successive surfacing | Frankel and Clark, 2000 [65] |
| M. novaeangliae | Sonar | modifies courtship calls | Miller, 2000 [63] |
| T. truncatus | pile driving noise | modifies sound producing | David, 2006 [62] |
| E. glacialis | vessels noise | modifies calling behavior | Parks et al., 2007 [64] |
| E. australis | |||
| G. cruentatus | boating and shipping noise | decreases time in nest caring and increases time in the shelters | Picciulin et al., 2010 [57] |
| C. chromis | |||
| C. caretta | seismic air-gun shooting | induces startle response | DeRuiter et al., 2012 [56] |
| M. densirostris | mid-frequency sonar | disrupts foraging and induces avoidance behavior | Tyack et al., 2011 [55] |
3.3. Changes in Population Distribution and Abundance
3.4. The Other Physiological Impacts
| Species | Types of Anthropogenic Noise | Effects | References |
|---|---|---|---|
| Z. cavirostris | Sonar | causes mass strandings | Frantzis, 1998 [68] |
| A. dux | seismic air-gun shooting | causes mass strandings | Guerra et al., 2011 [53] |
| O. orca | high-amplitude acoustic harassment devices | induces emigration | Morton, 2002 [73] |
| P. phocoena | pile driving noise | induces emigration | Thompson et al., 2010 [75] |
| T. truncatus | |||
| C. harengus, | seismic air-gun shooting | induces emigration | Slotte et al., 2004 [4] |
| M. poutassou | |||
| P. phocoena | wind farm noise | induces emigration and alters vertical distribution | Carstensen et al., 2006 [74] |
| G. flavescens | wind farm noise | no detectable effects on community structure and biodiversity | Wilhelmsson et al., 2006 [78] |
| P. minutus | |||
| P. microps | |||
| T. bubalis | |||
| M. scorpius | |||
| S. goodie | seismic air-gun shooting | decreases catch rate | Skalski et al., 1992 [43]; Løkkeborg et al., 1993 [36]; Engås et al., 1996 [41] |
| S. paucispinis | |||
| S. chlorostictus | |||
| G. morhua | |||
| M. aeglefinus | |||
| P. virens | boating and shipping noise | decreases catch rate | Engås, 1994 [40] |
| M. aeglefinus | experimental noise emanating device | decreases catch rate | Nicholson et al., 1992 [42] |
| P. cygnus | seismic air-gun shooting | no detectable effect on catch rate | Parry and Gason, 2006 [77] |
| P. novaezelandiae | experimental noise emanating device | decreases population recruitment | Aguilar de Soto et al., 2013 [39] |
| A. crassa | tidal turbine and wind turbine noise | decreases population recruitment | Pine et al., 2012 [80] |
| H. crenulatus | |||
| C. crangon | experimental noise emanating device | decreases reproduction rates | Lagardère, 1982 [35] |
| M. magister | seismic air-gun shooting | no detectable effect on larval survival | Pearson et al., 1994 [79] |
| Z. cavirostris | naval sonar | mass strandings | Cox, et al., 2006 [70] |
| M. densirostris | |||
| M. europaeus | |||
| Z. cavirostris | naval sonar | mass strandings | Fernández, et al., 2005 [71] |
| M. densirostris | |||
| M. europaeus | |||
| Z. cavirostris | naval sonar | mass strandings | Jepson, et al., 2003 [72] |
| M. densirostris | |||
| M. europaeus | |||
| L. kempii | Underwater explosives | mass strandings | Klima et al., 1988 [69] |
| T. truncates | |||
| C. caretta |
| Species | Types of Anthropogenic Noise | Effects | References |
|---|---|---|---|
| C. crangon | experimental noise emanating device | increases metabolism and decreases growth | Lagardère, 1982 [35] |
| D. leucas | seismic air-gun shooting | increases metabolism and decreases immunity | Romano et al., 2004 [81] |
| T. truncates | |||
| H. erectus | increased ambient noise | increases metabolism and decreases growth and immunity | Anderson et al., 2011 [83] |
| C. maenas | ship noise | increases metabolism | Wale et al., 2013 [82] |
| D. labrax | seismic air-gun shooting | increases metabolism | Santulli et al., 1999 [84] |
| D. labrax | boating and ship noise | increases metabolism and induces motility | Buscaino et al., 2010 [2] |
| S. aurata | |||
| D. leucas | experimental noise emanating device | increases heart rate | Lyamin et al., 2011 [86] |
| P. elephas | ship noise | decreases immunity | Celi et al., 2014 [87] |
| M. chrysops | pile driving | induces barotraumas injuries | Casper et al., 2013 [85] |
| M. saxatilis |
4. Discussion and Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Andrew, R.K.; Howe, B.M.; Mercer, J.A.; Dzieciuch, M.A. Ocean ambient sound: Comparing the 1960s with the 1990s for a receiver off the California coast. Acoust. Res. Lett. 2002, 3, 65–70. [Google Scholar] [CrossRef]
- Buscaino, G.; Filiciotto, F.; Buffa, G.; Bellante, A.; di Stefano, V.; Assenza, A.; Fazio, F.; Caola, G.; Mazzola, S. Impact of an acoustic stimulus on the motility and blood parameters of European sea bass (Dicentrarchus labrax L.) and gilthead sea bream (Sparus aurata L.). Mar. Environ. Res. 2010, 69, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Johansson, K. Impact of Anthropogenic Noise on Fish Behaviour and Ecology. 2011. Available online: http://pub.epsilon.slu.se/8366/1/Johansson_K_111013. pdf (accessed on 11 July 2015).
- Slotte, A.; Hansen, K.; Dalen, J.; Ona, E. Acoustic mapping of pelagic fish distribution and abundance in relation to a seismic shooting area off the Norwegian west coast. Fish. Res. 2004, 67, 143–150. [Google Scholar] [CrossRef]
- Tyack, P.L. Implications for marine mammals of large-scale changes in the marine acoustic environment. J. Mammal. 2008, 89, 549–558. [Google Scholar] [CrossRef]
- Kastelein, R.A.; Heul, S.; Verboom, W.C.; Jennings, N.; Veen, J.; de Haan, D. Startle response of captive North Sea fish species to underwater tones between 0.1 and 64 kHz. Mar. Environ. Res. 2008, 65, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Codarin, A.; Wysocki, L.E.; Ladich, F.; Picciulin, M. Effects of ambient and boat noise on hearing and communication in three fish species living in a marine protected area (Miramare, Italy). Mar. Pollut. Bull. 2009, 58, 1880–1887. [Google Scholar] [CrossRef] [PubMed]
- André, M. The sperm whale sonar: Monitoring and use in mitigation of anthropogenic noise effects in the marine environment. Nucl. Instrum. Methods Phys. Res. Sect. A 2009, 602, 262–267. [Google Scholar] [CrossRef]
- Kight, C.R.; Swaddle, J.P. How and why environmental noise impacts animals: An integrative, mechanistic review. Ecol. Lett. 2011, 14, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Popper, A.N.; Hastings, M.C. The effects of human-generated sound on fish. Integr. Zool. 2009, 4, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Slabbekoorn, H.; Bouton, N.; van Opzeeland, I.; Coers, A.; Cate, C.; Popper, A.N. A noisy spring: The impact of globally rising underwater sound levels on fish. Trends Ecol. Evolut. 2010, 25, 419–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conservation and Development Problem Solving Team. Anthropogenic Noise in the Marine Environment: Potential Impacts on the Marine Resources and Stellwagen Bank and Channel Islands National Marine Sanctuaries; University of Maryland: Maryland, MD, USA, 2000; pp. 18–19. [Google Scholar]
- Rogers, P.H.; Cox, H. Underwater sound as a biological stimulus. In Sensory Biology of Aquatic Animals; Atema, J., Fay, R.R., Popper, A.N., Eds.; Springer-Verla: New York, NY, USA, 1988; pp. 131–149. [Google Scholar]
- Pitcher, T.J. Behaviour of Teleost Fishes, 2nd ed.; Chapman & Hall: London, UK, 1992. [Google Scholar]
- Simmonds, J.; MacLennan, D.N. Fisheries Acoustics: Theory and Practice, 2nd ed.; Blackwell Publishing: Oxford, UK, 2005. [Google Scholar]
- Helfman, G.; Collette, B.B.; Facey, D.E.; Bowen, B.W. The Diversity of Fishes: Biology, Evolution and Ecology, 2nd ed.; Wiley-Blackwell: Chichester, UK, 2009. [Google Scholar]
- Hawkins, A.; Rasmussen, K.J. The calls of gadoid fish. J. Mar. Biol. Assoc. UK 1978, 58, 891–911. [Google Scholar] [CrossRef]
- Everest, F.A.; Young, R.W.; Johnson, M.W. Acoustical characteristics of noise produced by snapping shrimp. J. Acoust. Soc. Am. 2005, 20, 137–142. [Google Scholar] [CrossRef]
- Ladich, F.; Yan, H. Correlation between auditory sensitivity and vocalization in anabantoid fishes. J. Comp. Physiol. A 1998, 182, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Popper, A.N.; Fay, R.R.; Platt, C.; Sand, O. Sound detection mechanisms and capabilities of teleost fishes. In Sensory Processing in Aquatic Environments; Collins, S.P., Marshall, N.J., Eds.; Springer-Verlag: New York, NY, USA, 2003; pp. 3–38. [Google Scholar]
- Urick, R.J. Principles of Underwater Sound, 3rd ed.; McGraw-Hill: New York, NY, USA, 1983. [Google Scholar]
- McDonald, M.A.; Hildebrand, J.A.; Wiggins, S.M. Increases in deep ocean ambient noise in the Northeast Pacific west of San Nicolas Island, California. J. Acoust. Soc. Am. 2006, 120, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Ross, D. Ship sources of ambient noise. IEEE J. Ocean. Eng. 2005, 30, 257–261. [Google Scholar] [CrossRef]
- Buck, E.H. Acoustic Thermometry of Ocean Climate: Marine Mammal Issues; Congressional Research Service: Washington, DC, USA, 1995.
- Gisiner, R.; Cudahy, E.; Frisk, G.; Gentry, R.; Hofman, R.; Popper, A.; Richardson, W.J. Proceedings: Workshop on the Effects of Anthropogenic Noise in the Marine Environment; Office of Naval Research: Arlington, VA, USA, 1998.
- Richardson, W.J.; Finley, K.J.; Miller, G.W.; Davis, R.A.; Koski, W.R. Feeding, social and migration behavior of bowhead whales, Balaena mysticetus, in Baffin Bay vs. the Beaufort Sea—Regions with different amount of human activity. Mar. Mammal Sci. 1995, 11, 1–45. [Google Scholar] [CrossRef]
- Simard, Y.; Lepage, R.; Gervaise, C. Anthropogenic sound exposure of marine mammals from seaways: Estimates for Lower St. Lawrence Seaway, eastern Canada. Appl. Acoust. 2010, 71, 1093–1098. [Google Scholar] [CrossRef]
- Amoser, S.; Wysocki, L.E.; Ladich, F. Noise emission during the first powerboat race in an Alpine lake and potential impact on fish communities. J. Acoust. Soc. Am. 2004, 116, 3789–3797. [Google Scholar] [CrossRef] [PubMed]
- Bregman, A.S. Auditory Scene Analysis: The Perceptual Organization of Sound; MIT Press: Cambridge, MA, USA, 1994. [Google Scholar]
- Popper, A.N.; Hastings, M.C. The effects of anthropogenic sources of sound on fishes. J. Fish Biol. 2009, 75, 455–489. [Google Scholar] [CrossRef] [PubMed]
- Southall, B.L.; Bowles, A.E.; Ellison, W.T.; Finneran, J.J.; Gentry, R.L.; Greene, C.R.; Kastak, D.; Ketten, D.R.; Miller, J.H.; Nachtigall, P.E.; et al. Marine mammal noise-exposure criteria: Initial scientific recommendations. Bioacoustics 2008, 17, 273–275. [Google Scholar] [CrossRef]
- Brawn, V.M. Sound production by the cod (Gadus callarias L.). Behaviour 1961, 18, 239–255. [Google Scholar] [CrossRef]
- Schwarz, A.L. The behavior of fishes in their acoustic environment. Environ. Biol. Fishes 1985, 13, 3–15. [Google Scholar] [CrossRef]
- Scholik, A.R.; Yan, H.Y. The effects of noise on the auditory sensitivity of the bluegill sunfish, Lepomis macrochirus. Comp. Biochem. Physiol. Part A 2002, 133, 43–52. [Google Scholar] [CrossRef]
- Lagardère, J. Effects of noise on growth and reproduction of Crangon crangon in rearing tanks. Mar. Biol. 1982, 71, 177–185. [Google Scholar] [CrossRef]
- Løkkeborg, S.; Soldal, A.V. The influence of seismic exploration with airguns on cod (Gadus morhua) behaviour and catch rates. ICES J. Mar. Sci. Symp. 1993, 196, 62–67. [Google Scholar]
- McCauley, R.D.; Fewtrell, J.; Popper, A.N. High intensity anthropogenic sound damages fish ears. J. Acoust. Soc. Am. 2003, 113, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, A.L.; Greer, G.L. Responses of Pacific herring, Clupea harengus pallasi, to some underwater sounds. Can. J. Fish. Aquat. Sci. 1984, 41, 1183–1192. [Google Scholar] [CrossRef]
- Aguilar de Soto, N.; Delorme, N.; Atkins, J.; Howard, S.; Williams, J.; Johnson, M. Anthropogenic noise causes body malformations and delays development in marine larvae. Sci. Rep. 2013, 3, 2831–2835. [Google Scholar]
- Engås, A. The effects of trawl performance and fish behaviour on the catching efficiency of demersal sampling trawls. In Marine Fish Behaviour in Capture and Abundance Estimation; Ferno, A., Olsen, S., Eds.; Fishing News Books: Oxford, UK, 1994; pp. 45–68. [Google Scholar]
- Engås, A.; Løkkeborg, S.; Ona, E.; Soldal, A.V. Effects of seismic shooting on local abundance and catch rates of cod (Gadus morhua) and haddock (Melanogrammus aeglefinus). Can. J. Fish. Aquat. Sci. 1996, 53, 2238–2249. [Google Scholar] [CrossRef]
- Nicholson, M.; Rackham, B.; Mitson, R. Measuring the effect of underwater radiated noise on trawl catches. In Proceedings of the ICES FTFB and FAST Joint Working Group Meeting, Bergen, Norway, 16 June 1992.
- Skalski, J.R.; Pearson, W.H.; Malme, C.I. Effects of sounds from a geophysical survey device on catch-per-unit-effort in a hook-and-line fishery for rockfish (Sebastes spp.). Can. J. Fish. Aquat. Sci. 1992, 49, 1357–1365. [Google Scholar] [CrossRef]
- Clark, C.W.; Ellison, W.T.; Southall, B.L.; Hatch, L.; Parijs, S.M.; Frankel, A.; Ponirakis, D. Acoustic masking in marine ecosystems: Intuitions, analysis, and implication. Mar. Ecol. Prog. Ser. 2009, 395, 201–222. [Google Scholar] [CrossRef]
- Southall, B.L.; Schusterman, R.J.; Kastak, D. Acoustic communication ranges for northern elephant seals (Mirounga angustirostris). Aquat. Mammal. 2003, 29, 202–213. [Google Scholar] [CrossRef]
- Vasconcelos, R.O.; Amorim, M.C.; Ladich, F. Effects of ship noise on the detectability of communication signals in the Lusitanian toadfish. J. Exp. Biol. 2007, 210, 2104–2112. [Google Scholar] [CrossRef] [PubMed]
- Aguilar de Soto, N.; Johnson, M.; Madsen, P.T.M.; Tyack, P.L.; Bocconcelli, A.; Borsani, F. Dose intense ship noise disrupt foraging in deep-diving Cuvier’s beaked whales (Ziphius cavirostris)? Mar. Mammal Sci. 2006, 22, 690–699. [Google Scholar] [CrossRef]
- Lucke, K.; Siebert, U.; Lepper, P.A.; Blanchet, M.A. Temporary shift in masked hearing thresholds in a harbor porpoise (Phocoena phocoena) after exposure to seismic airgun stimuli. J. Acoust. Soc. Am. 2009, 125, 4060–4070. [Google Scholar] [CrossRef] [PubMed]
- Nachtigall, P.E.; Supin, A.Y.; Pawloski, J.; Au, W.W. Temporary threshold shifts after noise exposure in the bottlenose dolphin (Tursiops truncatus) measured using evoked auditory potentials. Mar. Mammal Sci. 2004, 20, 673–687. [Google Scholar] [CrossRef]
- Popper, A.N.; Smith, M.E.; Cott, P.A.; Hanna, B.W.; MacGillivray, A.O.; Austin, M.E.; Mann, D.A. Effects of exposure to seismic airgun use on hearing of three fish species. J. Acoust. Soc. Am. 2005, 117, 3958–3971. [Google Scholar] [CrossRef] [PubMed]
- Scholik, A.R.; Yan, H.Y. Effects of underwater noise on auditory sensitivity of a cyprinid fish. Hear. Res. 2001, 152, 17–24. [Google Scholar] [CrossRef]
- André, M.; Solé, M.; Lenoir, M.; Durfort, M.; Quero, C.; Mas, A.; Lombarte, A.; van der Schaar, M.; López-Bejar, M.; Morell, M.; et al. Low-frequency sounds induce acoustic trauma in cephalopods. Front. Ecol. Environ. 2011, 9, 489–493. [Google Scholar] [CrossRef]
- Guerra, Á.; González, Á.F.; Pascual, S.; Dawe, E.G. The giant squid Architeuthis: An emblematic invertebrate that can represent concern for the conservation of marine biodiversity. Biol. Conserv. 2011, 144, 1989–1997. [Google Scholar]
- Fewtrell, J.L.; McCauley, R.D. Impact of air gun noise on the behaviour of marine fish and squid. Mar. Pollut. Bull. 2012, 64, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Tyack, P.L.; Zimmer, W.M.; Moretti, D.; Southall, B.L.; Claridge, D.E.; Durban, J.W.; Clark, C.W.; D’Amico, A.; DiMarzio, N.; Jarvis, S.; et al. Beaked whales respond to simulated and actual navy sonar. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- DeRuiter, S.; Larbi Doukare, K. Loggerhead turtles dive in response to airgun sound exposure. Endanger. Species Res. 2012, 16, 55–63. [Google Scholar] [CrossRef]
- Picciulin, M.; Sebastianutto, L.; Codarin, A.; Farina, A.; Ferrero, E.A. In situ behavioural responses to boat noise exposure of Gobius cruentatus (Gmelin, 1789; fam. Gobiidae) and Chromis chromis (Linnaeus, 1758; fam. Pomacentridae) living in a Marine Protected Area. J. Exp. Mar. Biol. Ecol. 2010, 386, 125–132. [Google Scholar] [CrossRef]
- Bruintjes, R.; Radford, A.N. Context-dependent impacts of anthropogenic noise on individual and social behaviour in a cooperatively breeding fish. Anim. Behav. 2013, 85, 1343–1349. [Google Scholar] [CrossRef]
- Purser, J.; Radford, A.N. Acoustic noise induces attention shifts and reduces foraging performance in three-spined sticklebacks (Gasterosteus aculeatus). PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.A.; Giraldo-Perez, P.; Smith, S.; Blumstein, D.T. Anthropogenic noise affects risk assessment and attention: The distracted prey hypothesis. Biol. Lett. 2010, 6, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Wale, M.A.; Simpson, S.D.; Radford, A.N. Noise negatively affects foraging and antipredator behaviour in shore crabs. Anim. Behav. 2013, 86, 111–118. [Google Scholar] [CrossRef]
- David, J.A. Likely sensitivity of bottlenose dolphins to pile-driving noise. Water Environ. J. 2006, 20, 48–54. [Google Scholar] [CrossRef]
- Miller, P.J.; Biassoni, N.; Samuels, A.; Tyack, P.L. Whale songs lengthen in response to sonar. Nature 2000, 405, 903–903. [Google Scholar] [CrossRef] [PubMed]
- Parks, S.E.; Clark, C.W.; Tyack, P.L. Short- and long-term changes in right whale calling behavior: The potential effects of noise on acoustic communication. J. Acoust. Soc. Am. 2007, 122, 3725–3731. [Google Scholar] [CrossRef] [PubMed]
- Frankel, A.; Clark, C. Behavioral responses of humpback whales (Megaptera novaeangliae) to full-scale ATOC signals. J. Acoust. Soc. Am. 2000, 108, 1930–1937. [Google Scholar] [CrossRef] [PubMed]
- Bailey, H.; Senior, B.; Simmons, D.; Rusin, J.; Picken, G.; Thompson, P.M. Assessing underwater noise levels during pile-driving at an offshore windfarm and its potential effects on marine mammals. Mar. Pollut. Bull. 2010, 60, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Wardle, C.; Carter, T.; Urquhart, G.; Johnstone, A.; Ziolkowski, A.; Hampson, G.; Mackie, D. Effects of seismic air guns on marine fish. Cont. Shelf Res. 2001, 21, 1005–1027. [Google Scholar] [CrossRef]
- Frantzis, A. Does acoustic testing strand whales? Nature 1998, 392. [Google Scholar] [CrossRef] [PubMed]
- Klima, E.F.; Gitschlag, G.R.; Renaud, M.L. Impacts of the explosive removal of offshore petroleum on sea turtles and dolphins. Mar. Fish. Rev. 1988, 50, 33–42. [Google Scholar]
- Cox, T.M.; Ragen, T.J.; Read, A.J.; Vos, E.; Baird, R.W.; Balcomb, K.; Barlow, J.; Caldwell, J.; Cranford, T.; Crum, L.; et al. Understanding the impacts of anthropogenic sound on beaked whales. J. Cetacean Res. Manag. 2006, 7, 177–187. [Google Scholar]
- Fernández, A.; Edwards, J.F.; Rodríguez, F.; Espinosa de los Monteros, A.; Herráez, P.; Castro, P.; Jaber, J.R.; Martín, V.; Arbelo, M. “Gas and fat embolic syndrome” involving a mass stranding of beaked whales (family Ziphiidae) exposed to anthropogenic sonar signals. Vet. Pathol. 2005, 42, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Jepson, P.D.; Arbelo, M.; Deaville, R.; Patterson, I.A.P.; Castro, P.; Baker, J.R.; Degollada, E.; Ross, H.M.; Herráez, P.; Pocknell, A.M.; et al. Gas-bubble lesions in stranded cetaceans: Was sonar responsible for a spate of whale deaths after an Atlantic military exercise? Nature 2003, 425, 575–576. [Google Scholar] [CrossRef] [PubMed]
- Morton, A. Displacement of Orcinus orca (L.) by high amplitude sound in British Columbia, Canada. ICES J. Mar. Sci. 2002, 59, 71–80. [Google Scholar] [CrossRef]
- Carstensen, J.; Henriksen, O.D.; Teilmann, J. Impacts of offshore wind farm construction on harbour porpoises: Acoustic monitoring of echolocation activity using porpoise detectors (T-PODs). Mar. Ecol. Prog. Ser. 2006, 321, 295–308. [Google Scholar] [CrossRef]
- Thompson, P.M.; Lusseau, D.; Barton, T.; Simmons, D.; Rusin, J.; Bailey, H. Assessing the responses of coastal cetaceans to the construction of offshore wind turbines. Mar. Pollut. Bull. 2010, 60, 1200–1208. [Google Scholar] [CrossRef] [PubMed]
- Engås, A.; Løkkeborg, S. Effects of seismic shooting and vessel-generated noise on fish behaviour and catch rates. Bioacoustics 2002, 12, 313–316. [Google Scholar] [CrossRef]
- Parry, G.D.; Gason, A. The effect of seismic surveys on catch rates of rock lobsters in western Victoria, Australia. Fish. Res. 2006, 79, 272–284. [Google Scholar] [CrossRef]
- Wilhelmsson, D.; Malm, T.; Ohman, M. The influence of offshore windpower on demersal fish. ICES J. Mar. Sci. 2006, 63, 775–784. [Google Scholar] [CrossRef]
- Pearson, W.H.; Skalski, J.R.; Sulkin, S.D.; Malme, C.I. Effects of seismic energy releases on the survival and development of zoeal larvae of dungeness crab (Cancer magister). Mar. Environ. Res. 1994, 38, 93–113. [Google Scholar] [CrossRef]
- Pine, M.K.; Jeffs, A.G.; Radford, C.A. Turbine sound may influence the metamorphosis behaviour of estuarine crab megalopae. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Romano, T.A.; Keogh, M.J.; Kelly, C.; Feng, P.; Berk, L.; Schlundt, C.E.; Carder, D.A.; Finneran, J.J. Anthropogenic sound and marine mammal health: Measures of the nervous and immune systems before and after intense sound exposure. Can. J. Fish. Aquat. Sci. 2004, 61, 1124–1134. [Google Scholar] [CrossRef]
- Wale, M.A.; Simpson, S.D.; Radford, A.N. Size-dependent physiological responses of shore crabs to single and repeated playback of ship noise. Biol. Lett. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.A.; Berzins, I.K.; Fogarty, F.; Hamlin, H.J.; Guillette, L.J. Sound, stress, and seahorses: The consequences of a noisy environment to animal health. Aquaculture 2011, 311, 129–138. [Google Scholar] [CrossRef]
- Santulli, A.; Modica, A.; Messina, C.; Ceffa, L.; Curatolo, A.; Rivas, G.; Fabi, G.; D’amelio, V. Biochemical responses of European sea bass (Dicentrarchus labrax L.) to the stress induced by off shore experimental seismic prospecting. Mar. Pollut. Bull. 1999, 38, 1105–1114. [Google Scholar] [CrossRef]
- Casper, B.M.; Halvorsen, M.B.; Matthews, F.; Carlson, T.J.; Pooper, A.N. Recovery of barotraumas injuries resulting from exposure to pile driving sound in two sizes of hybrid striped bass. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Lyamin, O.I.; Korneva, S.M.; Rozhnov, V.V.; Mukhametov, L.M. Cardiorespiratory changes in beluga in response to acoustic noise. Dokl. Biol. Sci. 2011, 440, 257–258. [Google Scholar] [CrossRef] [PubMed]
- Celi, M.; Filiciotto, F.; Vazzana, M.; Arizza, V.; Maccarrone, V.; Ceraulo, M.; Mazzola, S.; Buscaino, G. Shipping noise affecting immune responses of European spiny lobster (Palinurus elephas). Can. J. Zool. 2014, 93, 113–121. [Google Scholar] [CrossRef]
- Filiciotto, F.; Giacalone, V.M.; Fazio, F.; Buffa, G.; Piccione, G.; Maccarrone, V.; di Stefano, V.; Mazzola, S.; Buscaino, G. Effect of acoustic environment on gilthead sea bream (Sparus aurata): Sea and onshore aquaculture background noise. Aquaculture 2013, 414, 36–45. [Google Scholar] [CrossRef]
- Papoutsoglou, S.E.; Karakatsouli, N.; Batzina, A.; Papoutsoglou, E.S.; Tsopelakos, A. Effect of music stimulus on gilthead seabream Sparus aurata physiology under different light intensity in a re-circulating water system. J. Fish Biol. 2008, 73, 980–1004. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, C.; Zhao, X.; Liu, G. Noise in the Sea and Its Impacts on Marine Organisms. Int. J. Environ. Res. Public Health 2015, 12, 12304-12323. https://doi.org/10.3390/ijerph121012304
Peng C, Zhao X, Liu G. Noise in the Sea and Its Impacts on Marine Organisms. International Journal of Environmental Research and Public Health. 2015; 12(10):12304-12323. https://doi.org/10.3390/ijerph121012304
Chicago/Turabian StylePeng, Chao, Xinguo Zhao, and Guangxu Liu. 2015. "Noise in the Sea and Its Impacts on Marine Organisms" International Journal of Environmental Research and Public Health 12, no. 10: 12304-12323. https://doi.org/10.3390/ijerph121012304