Perinatal BPA Exposure Induces Hyperglycemia, Oxidative Stress and Decreased Adiponectin Production in Later Life of Male Rat Offspring

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Treatment
2.2. Histologic Examination of Subcutaneous Tissue Oil Red O
2.3. Blood and Tissue Collection
2.4. Biochemical Assays
2.5. Western Blotting
2.6. Real-Time PCR
2.7. Statistical Analysis
3. Results
3.1. Maternal Physiology during Gestation
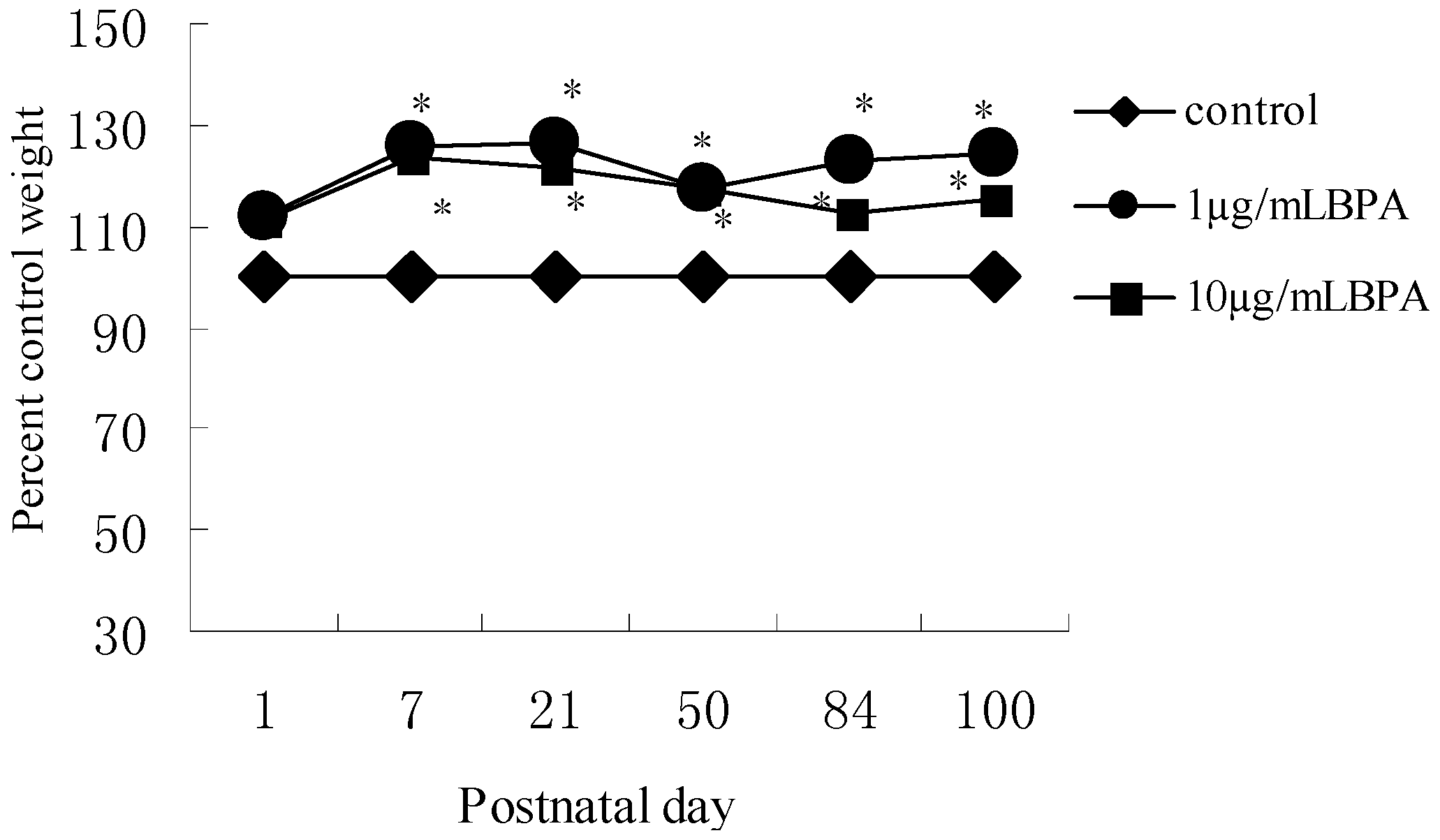
3.2. Effect of BPA Exposure on Body Weight in Male Offspring from PND1 to PND100

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F0-Control | F0-BPA1 | F0-BPA10 | |
|---|---|---|---|
| Body weight on GD 0 (g) | 262.7 ± 9.58 | 258.8 ± 6.49 | 251.3 ± 6.81 |
| Body weight gain (g) | |||
| GD 0–7 | 25.3 ± 3.1 | 23.6 ± 1.3 | 25.2 ± 1.9 |
| GD 8–14 | 27.6 ± 4.7 | 29.9 ± 4.6 | 28.4 ± 2.9 |
| GD 15–21 | 55.9 ± 8.2 | 60.0 ± 4.8 | 59.6 ± 6.4 |
| Food consumption (g per day) | |||
| GD 0–7 | 24.9 ± 3.4 | 27.6 ± 1.5 | 24.7 ± 1.2 |
| GD 8–14 | 22.5 ± 4.3 | 28.3 ± 3.4 | 25.9 ± 4.9 |
| GD 15–21 | 25.4 ± 4.1 | 26.0 ± 3.7 | 23.6 ± 1.6 |
| Water consumption (mL per day) | |||
| GD 0–7 | 33.8 ± 2.3 | 33.6 ± 2.6 | 27.7 ± 1.8 |
| GD 8–14 | 33.2 ± 4.9 | 35.8 ± 4.8 | 29.4 ± 2.4 |
| GD 15–21 | 20.2 ± 2.1 | 25.0 ± 2.3 | 25.6 ± 2.9 |

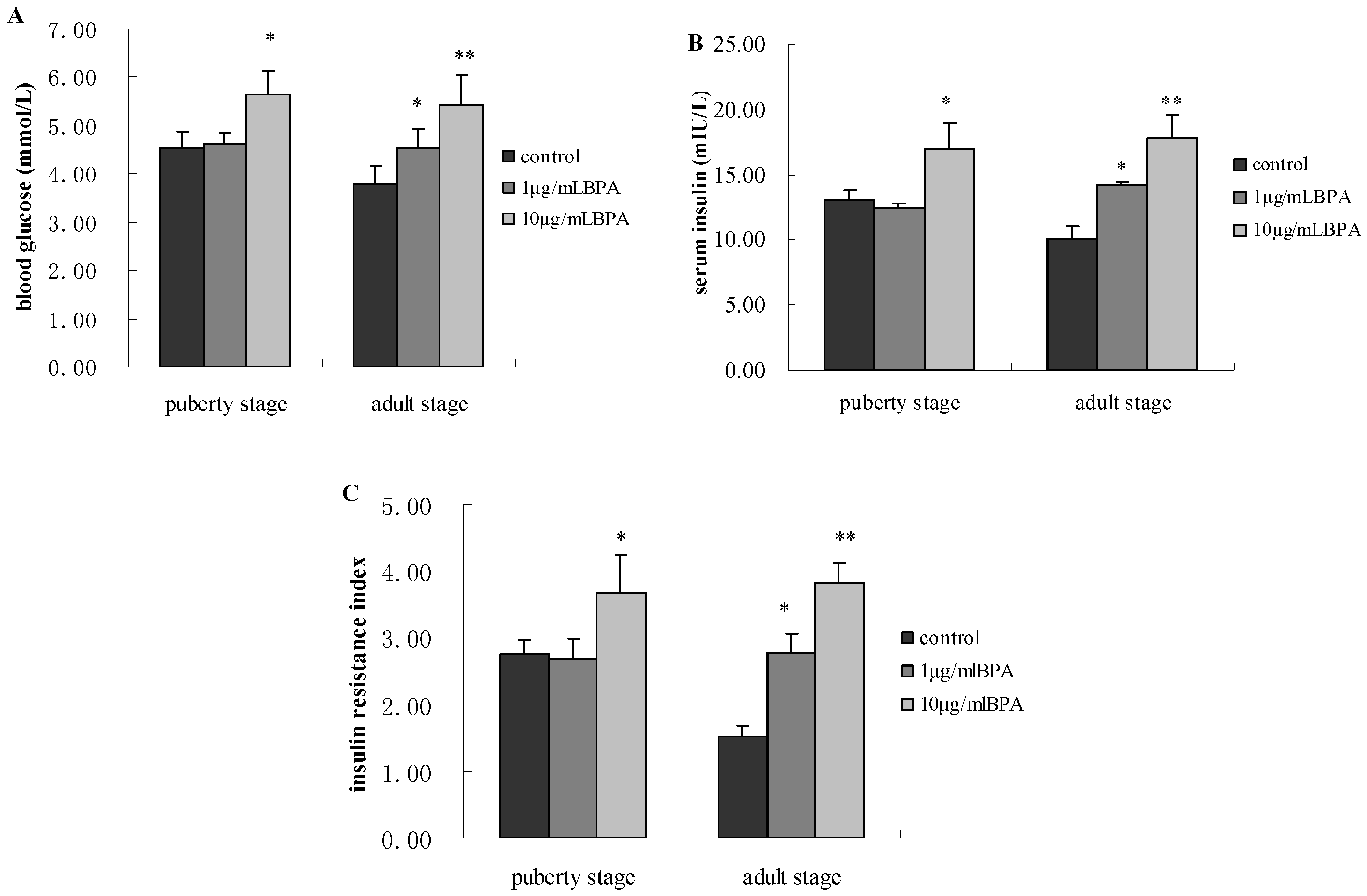
3.3. Effect of BPA Exposure on Glucose Metabolism of Male Offspring at Puberty Stage and Adult Stage
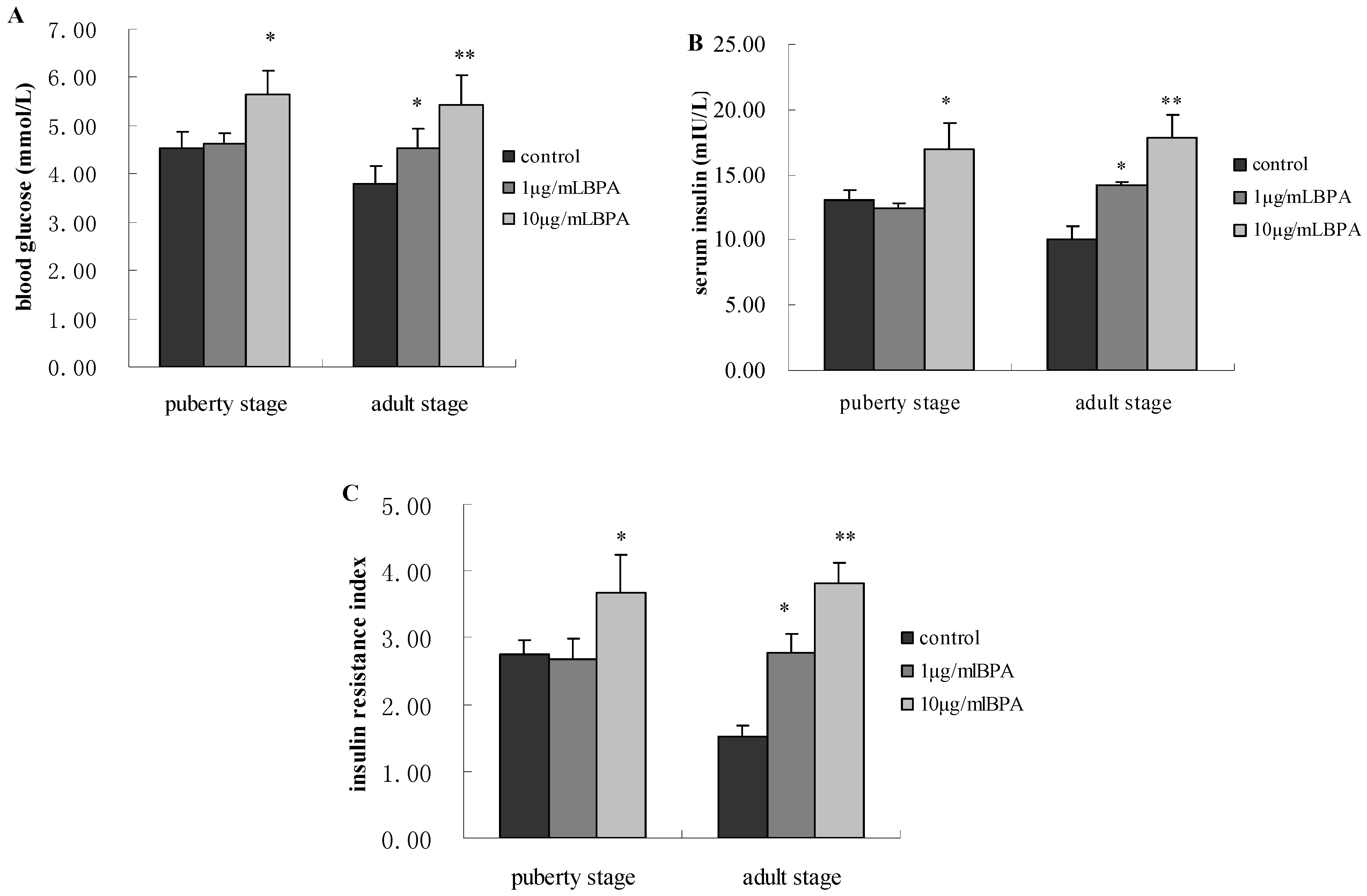
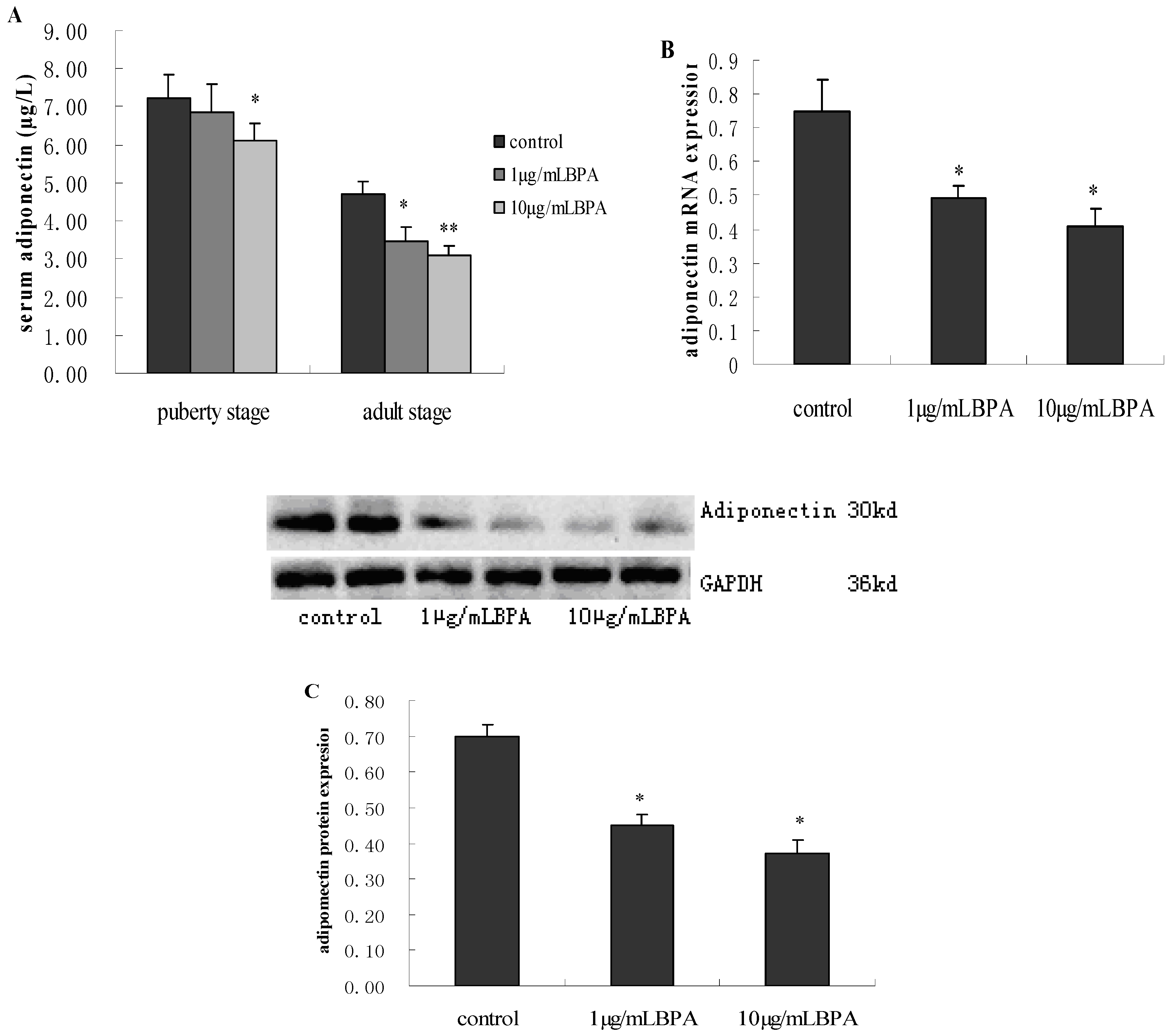
3.4. Effect of BPA Exposure on ADP Production and Release in Male Offspring at Puberty Stage or Adult Stage

3.5. Effect of BPA Exposure on the Size of Subcutaneous Adipocytes of Male Offspring at Adult Stage

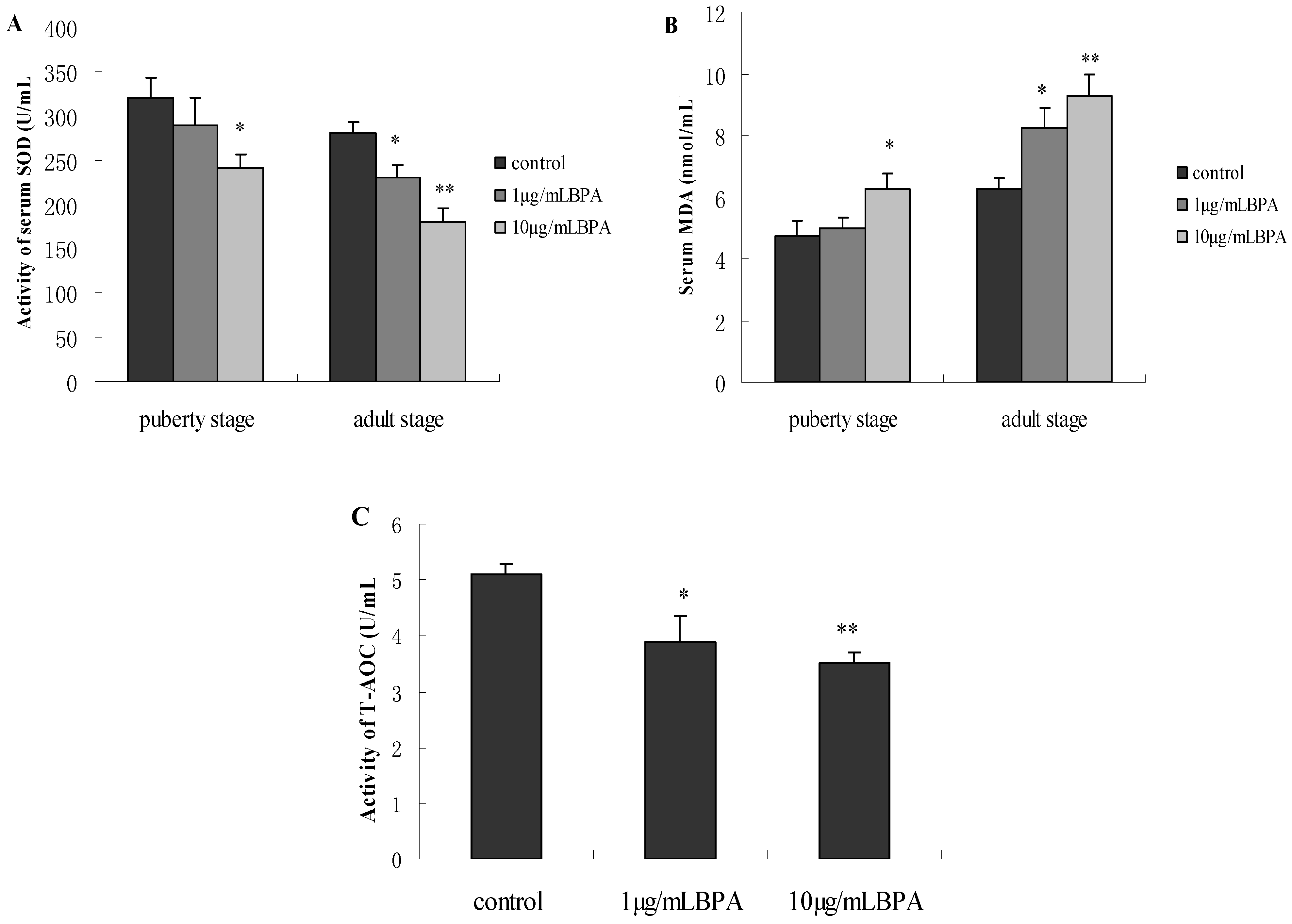
3.6. Effect of BPA Exposure on Serum SOD, MDA and T-AOC in Male Offspring at Puberty Stage or Adult Stage
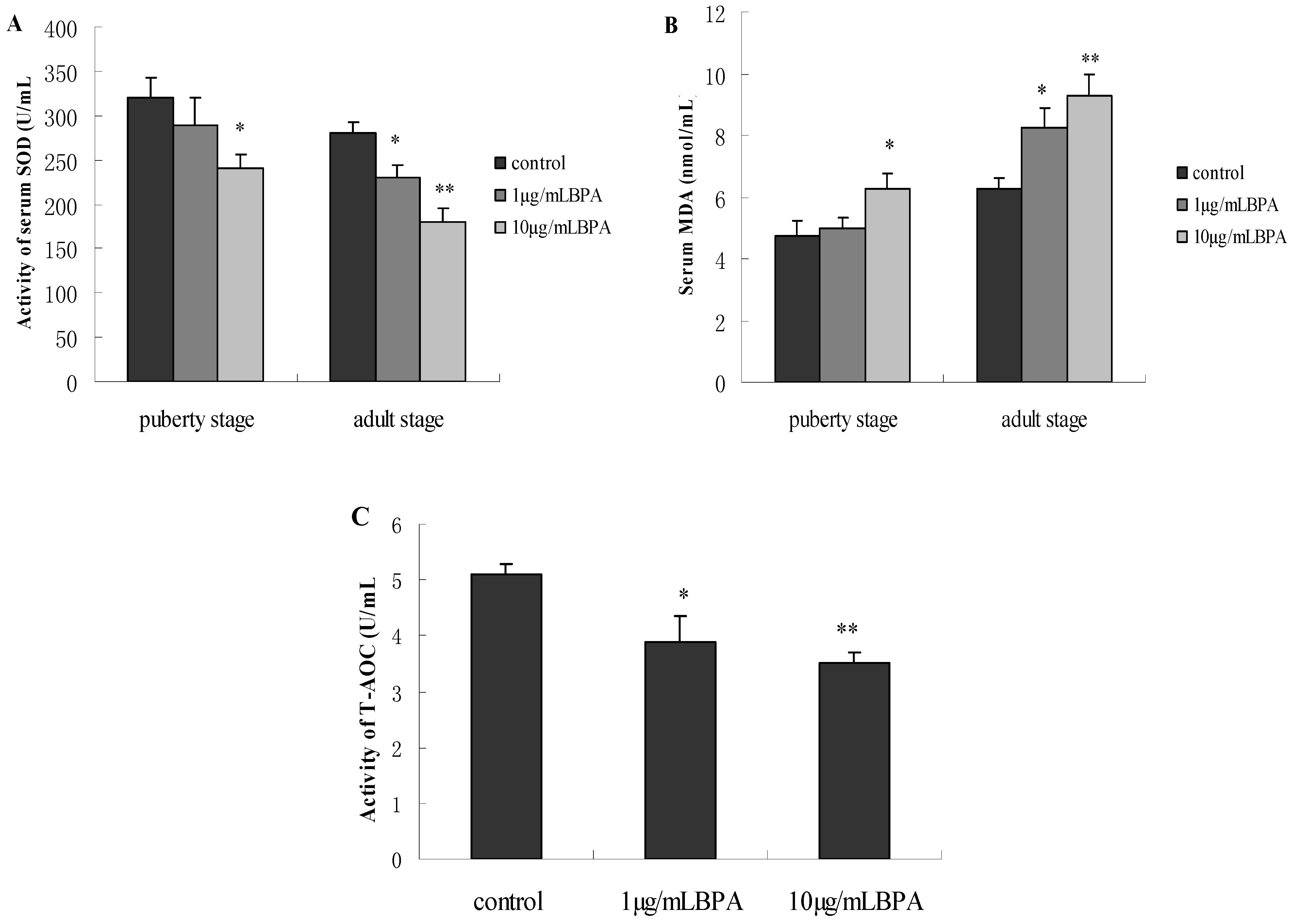
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schoenfelder, G. Urinary, circulating and tissue biomonitoring studies indicate widespread exposure to bisphenol A. Environ. Health Perspect. 2010, 118, 1055–1070. [Google Scholar] [CrossRef]
- Carwile, J.L.; Michels, K.B. Urinary bisphenol A and obesity: NHANES 2003–2006. Environ. Res. 2011, 111, 825–830. [Google Scholar] [CrossRef]
- Groff, T. Bisphenol A: Invisible pollution. Curr. Opin. Pediatr. 2010, 22, 524–529. [Google Scholar] [CrossRef]
- Fall, C.H. Fetal programming and the risk of noncommunication disease. Indian J. Pediatr. 2013, 1, 13–20. [Google Scholar] [CrossRef]
- Alonso-Magdalena, P.; Vieira, E.; Soriano, S.; Menes, L.; Burks, D.; Quesada, I.; Nadal, A. Bisphenol A exposure during pregnancy disrupts glucose homeostasis in mothers and adult male offspring. Environ. Health Perspect. 2010, 118, 1243–1250. [Google Scholar] [CrossRef]
- Batista, T.M.; Alonso-Magdalena, P.; Viera, E.; Amaral, M.E.; Cederroth, C.R.; Nef, S.; Quesada, I.; Carneiro, E.M.; Nadal, A. Short-term treatment with bisphenol-A leads to metabolic abnormalities in adult male rats. PLoS One 2012, 7, e33814. [Google Scholar] [CrossRef]
- Alonso-Magdalena, P.; Ropero, A.B.; Soriano, S.; Quesada, I.; Nadal, A. Bisphenol-A: A new diabetogenic factor? Hormones 2010, 9, 118–126. [Google Scholar]
- Kim, K.; Park, H. Association between urinary concentrations of bisphenol A and type 2 diabetes in Korean adults: A population-based cross-sectional study. Int. J. Hyg. Environ. Health 2013, 216, 467–471. [Google Scholar] [CrossRef]
- Nadal, A.; Alonso-Magdalena, P.; Soriano, S.; Quesada, I.; Ropero, A.B. The pancreatic beta-cell as a target of estrogens and xenoestrogens: Implications for blood glucose homeostasis and diabetes. Mol. Cell. Endocrinol. 2009, 304, 63–68. [Google Scholar] [CrossRef]
- Alonso-Magdalena, P.; Ropero, A.B.; Carrera, M.P.; Cederroth, C.R.; Baquié, M.; Gauthier, B.R.; Nef, S.; Stefani, E.; Nadal, A. Pancreatic insulin content regulation by the estrogen receptor ER alpha. PLoS One 2008, 3, e2069. [Google Scholar] [CrossRef]
- Indumathi, D.; Jayashree, S.; Selvaraj, J.; Sathish, S.; Mayilvanan, C.; Akilavalli, N.; Balasubramanian, K. Effect of bisphenol A on insulin signal transduction and glucose oxidation in skeletal muscle of adult male albino rat. Hum. Exp. Toxicol. 2013, 32, 960–971. [Google Scholar] [CrossRef]
- Lin, Y.; Sun, X.; Qiu, L.; Wei, J.; Huang, Q.; Fang, C.; Ye, T.; Kang, M.; Shen, H.; Dong, S. Exposure to bisphenol A induces dysfunction of insulin secretion and apoptosis through the damage of mitochondria in rat insulinoma (INS-1) cells. Cell Death Dis. 2013, 4, e460. [Google Scholar] [CrossRef]
- Kidani, T.; Kamei, S.; Miyawaki, J.; Aizawa, J.; Sakayama, K.; Masuno, H. Bisphenol A downregulates Akt signaling and inhibits adiponectin production and secretion in 3T3-L1 adipocytes. J. Atheroscler. Thromb. 2010, 17, 834–843. [Google Scholar] [CrossRef]
- Hassan, Z.K.; Elobeid, M.A.; Virk, P.; Omer, S.A.; ElAmin, M.; Daghestani, M.H.; Alolayan, E.M. Bisphenol A induces hepatotoxicity through oxidative stress in rat model. Oxid. Med. Cell. Longev. 2012, 2012, 194829. [Google Scholar]
- Rolo, A.P.; Palmeira, C.M. Diabetes and mitochondrial function: Role of hyperglycemia and oxidative stress. Toxicol. Appl. Pharmacol. 2006, 212, 167–178. [Google Scholar] [CrossRef]
- Gerritsen, W.B.; van Boven, W.J.; Boss, D.S.; Haas, F.J.; van Dongen, E.P.; Aarts, L.P. Malondialdehyde in plasma, a biomarker of global oxidative stress during mini-CABG compared to on- and off-pump CABG surgery: A pilot study. Interact. Cardiovasc. Thorac. Surg. 2006, 5, 27–31. [Google Scholar]
- Wang, T.; Mao, X.; Li, H.; Qiao, S.; Xu, A.; Wang, J.; Lei, S.; Liu, Z.; Ng, K.F.; Wong, G.T.; et al. N-Acetylcysteine and allopurinol up-regulated the Jak/STAT3 and PI3K/Akt pathways via adiponectin and attenuated myocardial postischemic injury in diabetes. Free Radic. Biol. Med. 2013, 63, 291–303. [Google Scholar] [CrossRef]
- Koleva, D.I.; Orbetzova, M.M.; Atanassova, P.K. Adipose tissue hormones and appetite and body weight regulators in insulin resistance. Folia Med. 2013, 55, 25–32. [Google Scholar]
- Ben-Jonathan, N.; Hugo, E.R.; Brandebourg, T.D. Effects of bisphenol A on adipokine release from human adipose tissue: Implications for the metabolic syndrome. Mol. Cell. Endocrinol. 2009, 304, 49–54. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Rubin, B.S.; Soto, A.M. Bisphenol A: Perinatal exposure and body weight. Mol. Cell. Endocrinol. 2009, 304, 55–62. [Google Scholar] [CrossRef]
- Hugo, E.R.; Brandebourg, T.D.; Woo, J.G.; Loftus, J.; Alexander, J.W.; Ben-Jonathan, N. Bisphenol A at environmentally relevant doses inhibits adiponectin release from human adipose tissue explants and adipocytes. Environ. Health Perspect. 2008, 116, 1642–1647. [Google Scholar] [CrossRef]
- Korkmaz, G.G.; Konukoglu, D.; Kurtulus, E.M.; Irmak, H.; Bolayirli, M.; Uzun, H. Total antioxidant status and markers of oxidative stress in subjects with normal or impaired glucose regulation (IFG, IGT) in diabetic patients. Scand. J. Clin. Lab. Investig. 2013, 73, 641–649. [Google Scholar] [CrossRef]
- Iwakami, S.; Misu, H.; Takeda, T.; Sugimori, M.; Matsugo, S.; Kaneko, S.; Takamura, T. Concentration dependent dual effect effects of hydrogen peroxide on insulin signal transduction in H4IIEC hepatocyte. PLoS One 2011, 6, e27401. [Google Scholar]
- Harley, K.G.; Aguilar Schall, R.; Chevrier, J.; Tyler, K.; Aguirre, H.; Bradman, A.; Holland, N.T.; Lustig, R.H.; Calafat, A.M.; Eskenazi, B. Prenatal and postnatal bisphenol A exposure and body mass index in childhood in the CHAMACOS cohort. Environ. Health Perspect. 2013, 121, 514–520. [Google Scholar] [CrossRef]
- Somm, E.; Schwitzgebel, V.M.; Toulotte, A.; Cederroth, C.R.; Combescure, C.; Nef, S.; Aubert, M.L. Perinatal exposure to bisphenol a alters early adipogenesis in the rat. Environ. Health Perspect. 2009, 117, 1549–1555. [Google Scholar] [CrossRef]
- Sakurai, K.; Kawazuma, M.; Adachi, T.; Harigaya, T.; Saito, Y.; Hashimoto, N.; Mori, C. Bisphenol A affects glucose transport in mouse 3T3-F442A adipocytes. Br. J. Pharmacol. 2004, 141, 209–214. [Google Scholar] [CrossRef]
- Masuno, H.; Kidani, T.; Sekiya, K.; Sakayama, K.; Shiosaka, T.; Yamamoto, H.; Honda, K. Bisphenol A in combination with insulin can accelerate the conversion of 3T3-L1 fibroblasts to adipocytes. J. Lipid Res. 2002, 43, 676–684. [Google Scholar]
- Valentino, R.; D’Esposite, V.; Passaretti, F.; Liotti, A.; Cabaro, S.; Longo, M.; Perruolo, G.; Oriente, F.; Beguinot, F.; Formisano, P. Bisphenol-A impairs insulin action and up-regulates inflammatory pathways in human subcutaneous adipocytes and 3T3-L1 cells. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Kent, A.L. Developmental origins of health and adult disease: What should neonatologists/paediatricians be considering about the long-term health of their patients? J. Paediatr. Child. Health 2012, 48, 730–734. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Song, S.; Zhang, L.; Zhang, H.; Wei, W.; Jia, L. Perinatal BPA Exposure Induces Hyperglycemia, Oxidative Stress and Decreased Adiponectin Production in Later Life of Male Rat Offspring. Int. J. Environ. Res. Public Health 2014, 11, 3728-3742. https://doi.org/10.3390/ijerph110403728
Song S, Zhang L, Zhang H, Wei W, Jia L. Perinatal BPA Exposure Induces Hyperglycemia, Oxidative Stress and Decreased Adiponectin Production in Later Life of Male Rat Offspring. International Journal of Environmental Research and Public Health. 2014; 11(4):3728-3742. https://doi.org/10.3390/ijerph110403728
Chicago/Turabian StyleSong, Shunzhe, Ling Zhang, Hongyuan Zhang, Wei Wei, and Lihong Jia. 2014. "Perinatal BPA Exposure Induces Hyperglycemia, Oxidative Stress and Decreased Adiponectin Production in Later Life of Male Rat Offspring" International Journal of Environmental Research and Public Health 11, no. 4: 3728-3742. https://doi.org/10.3390/ijerph110403728
APA StyleSong, S., Zhang, L., Zhang, H., Wei, W., & Jia, L. (2014). Perinatal BPA Exposure Induces Hyperglycemia, Oxidative Stress and Decreased Adiponectin Production in Later Life of Male Rat Offspring. International Journal of Environmental Research and Public Health, 11(4), 3728-3742. https://doi.org/10.3390/ijerph110403728