Predictive Modeling of West Nile Virus Transmission Risk in the Mediterranean Basin: How Far from Landing?

Abstract
:1. Introduction
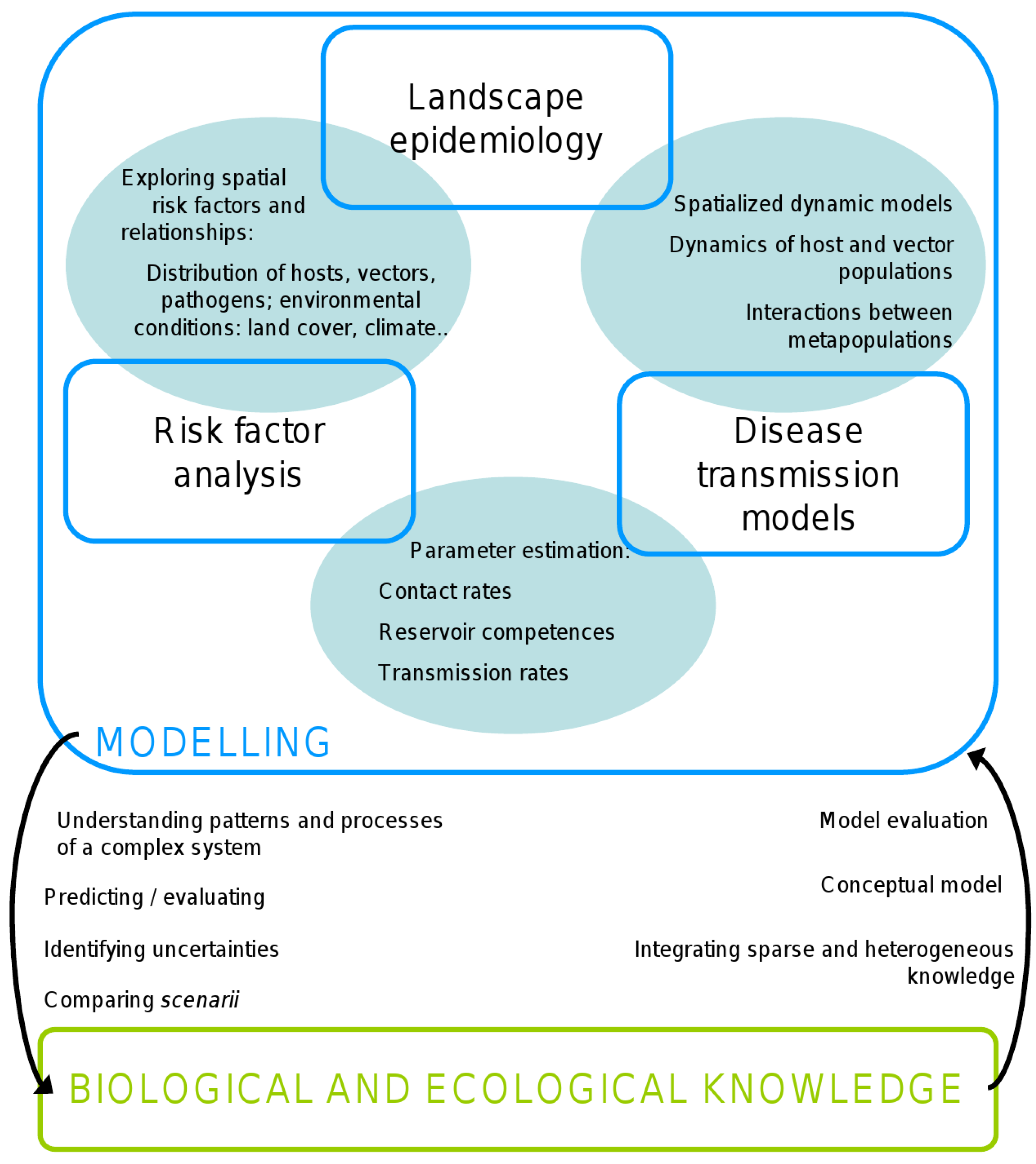
2. Method
3. Results
3.1. Risk Factor Analyses of WNF Occurrence and/or Transmission

{kind=link}
| Infection Marker | Scale | Explicative Variables | Validation | Prediction | Ref. | |||
|---|---|---|---|---|---|---|---|---|
| Abiotic | Landcover | Landscape | Other | |||||
| Horse seroprevalence | Local (France) | Age, breed, group size | Yes, internal | No | [36] | |||
| Horse cases and seroprevalence | Local (France) | Wet sansouire, open water, rice fields, dry bushes | No | No | [37] | |||
| Horse seroprevalence | Local (France) | Density of hetero-geneous agricultural areas | Insterspersion and juxtaposition index | Yes, internal | Yes | [38] | ||
| Horse seroprevalence | Local (Iran) | Elevation | Age | No | No | [39] | ||
| Horse seroprevalence | Local (Spain) | Number of horses within the holding, transport within the last 6 months, presence of mosquitoes | No | No | [40] | |||
| Horse seroprevalence | Country (Tunisia) | Night-time land surface temperature, biannual phase of NDVI | Distance to the nearest RAMSAR site | Yes, external | Yes | [41] | ||
| Horse cases | Local (Morocco) | NDVI, rainfall | No | No | [42] | |||
| Human and horse cases | Continental (Russia, Greece, Israel, Romania, Turkey, Hungary, Italy, Spain) | Temperature, Relative Humidity | No | No | [43] | |||
| Scale | Wild Birds | Mosquitoes | Risk Indices/Model | Ref. | ||
|---|---|---|---|---|---|---|
| Species | Abundance Model | Species | Abundance Model | |||
| Local (France) | 60 species | Qualitative probability of presence according to land cover (6 classes) | Cx. pipiens Cx. modestus | Qualitative density level (5 classes), data: bird-baited trapping | Vector and host occurrence probability indexes, host richness and abundance indexes | [44] |
| Country (Israel) | Cx. pipiens | Spearman and Pearson correlation with temperature and precipitation | [45] | |||
| Local (Italy) | Oc Caspius Cx. pipiens Cx. modestus | Bayesian Generalized Linear Mixed Model (GLMM) of CO2-baited trapping data according to elevation, rainfall, temperature, NDVI, season | [46] | |||
| Local (Italy) | Cx. pipiens | GLMM | [47] | |||
| Local (Spain) | 32 migratory species, present in large numbers, associated with aquatic habitat | Presence/absence: only abundant species (>2,000 pairs) are addressed | Cx. pipiens | Weighted Linear Combination (WLC) of temperature, rainfall rate, distance to the nearest humid area | WLC of wild bird presence, Cx. pipiens abundance and equid density | [48] |
| Study Type | Method | Ref. | ||
|---|---|---|---|---|
| Species/Genus | Explanatory/Calibrated Variables | Method | ||
| Respective roles of wild bird species | 25 Bird species | Migrating status | t-test | [49] |
| 72 Bird species | Migrating status Body weight | GLMM | [50] | |
| Respective roles of mosquito species | Duck, Horse Cx. pipiens, Cx. modestus, Ae Caspius, Ae. vexans | Host abundance and biomass | Multi-host model of host choice by vectors | [51] |
| WNV transmission dynamics | Passerines Cx. pipiens/Cx univittatus Horses Chicken | Vector-host ratio in each population Nestling-adult bite relative risk Passerine-incidental host bite relative risk | Meta-population model (3 vector populations, 5 host populations) | [52] |
| Use of a meta-population model | [53] | |||
3.2. Landscape Epidemiology Studies
3.3. Integrative Studies
3.3.1. The Respective Roles of Mosquito Species in WNV Transmission
3.3.2. The Respective Roles of Wild Bird Species in WNV Transmission
3.3.3. Mathematical Models of Transmission Dynamics
4. Discussion/Perspectives
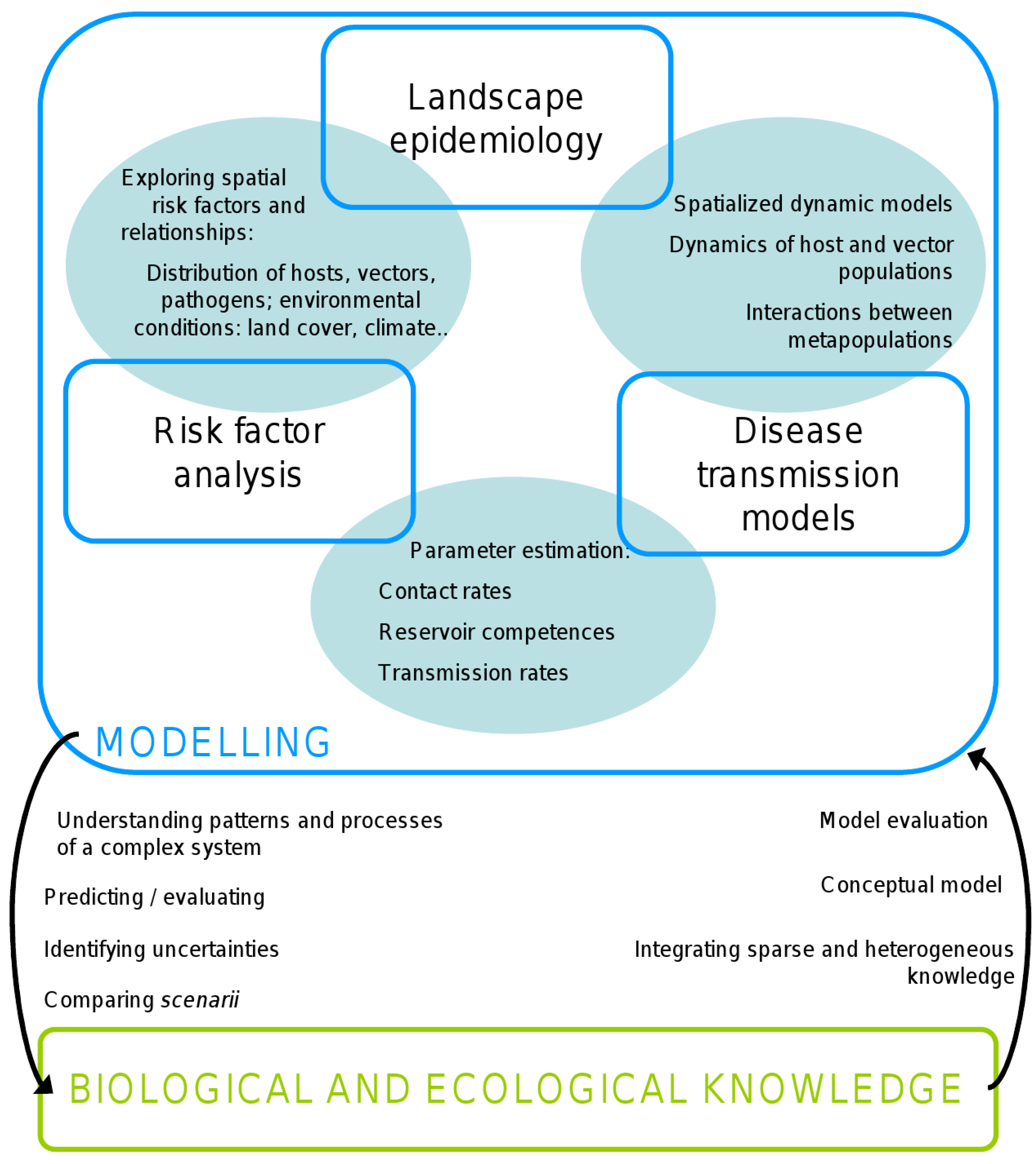
5. Conclusions
Conflicts of Interest
References
- Bunning, M.L.; Bowen, R.A.; Cropp, B.; Sullivan, K.; Davis, B.; Komar, N.; Godsey, M.; Baker, D.; Jetter, D.; Holmes, D.; et al. Experimental infection of horses with West Nile virus and their potential to infect mosquitoes and serve as amplifying hosts. Ann. N. Y. Acad. Sci. 2001, 951, 338–339. [Google Scholar]
- Rossi, S.; Ross, T.; Evans, J. West Nile virus. Clin. Lab. Med. 2010, 30, 47–65. [Google Scholar] [CrossRef]
- Sambri, V.; Capobianchi, M.; Charrel, R.; Fyodorova, M.; Gaibani, P.; Gould, E.; Niedrig, M.; Papa, A.; Pierro, A.; Rossini, G.; et al. West Nile virus in Europe: Emergence, epidemiology, diagnosis, treatment, and prevention. Clin. Microbiol. Infect. 2013, 19, 699–704. [Google Scholar] [CrossRef]
- Castillo-Olivares, J.; Wood, J. West Nile virus infection of horses. Vet. Res. 2004, 35, 467–483. [Google Scholar] [CrossRef]
- Gardner, I.; Wong, S.; Ferraro, G.; Balasuriya, U.; Hullinger, P.; Wilson, W.; Shi, P.; MacLachlan, N. Incidence and effects of West Nile virus infection in vaccinated and unvaccinated horses in California. Vet. Res. 2007, 38, 109–116. [Google Scholar] [CrossRef]
- Pybus, O.; Rambaut, A. Evolutionary analysis of the dynamics of viral infectious disease. Nat. Rev. Genet. 2009, 10, 540–550. [Google Scholar] [CrossRef]
- May, F.; Davis, C.; Tesh, R.; Barrett, A. Phylogeography of West Nile virus: From the cradle of evolution in Africa to Eurasia, Australia, and the Americas. J. Virol. 2011, 85, 2964–2974. [Google Scholar] [CrossRef]
- Ciccozzi, M.; Peletto, S.; Cella, E.; Giovanetti, M.; Lai, A.; Gabanelli, E.; Acutis, P.; Modesto, P.; Rezza, G.; Platonov, A.; et al. Epidemiological history and phylogeography of West Nile virus lineage 2. Infect. Genet. Evol. 2013, 17, 46–50. [Google Scholar] [CrossRef]
- Zehender, G.; Ebranati, E.; Bernini, F.; lo Presti, A.; Rezza, G.; Delogu, M.; Galli, M.; Ciccozzi, M. Phylogeography and epidemiological history of West Nile virus genotype 1a in Europe and the Mediterranean basin. Infect. Genet. Evol. 2011, 11, 646–653. [Google Scholar] [CrossRef]
- Kilpatrick, A. Globalization, land use, and the invasion of West Nile virus. Science 2011, 21, 323–327. [Google Scholar] [CrossRef]
- Nemeth, N.; Beckett, S.; Edwards, E.; Klenk, K.; Komar, N. Avian mortality surveillance for West Nile virus in Colorado. Am. J. Trop. Med. Hyg. 2007, 76, 431–437. [Google Scholar]
- Calistri, P.; Giovannini, A.; Hubalek, Z.; Ionescu, A.; Monaco, F.; Savini, G.; Lelli, R. Epidemiology of West Nile in Europe and in the Mediterranean basin. Open Vir. J. 2010, 4, 29–37. [Google Scholar]
- Bakoni, T.; Hubalek, Z.; Rudolf, I.; Nowotny, N. Novel flavivirus or new lineage of West Nile virus, central Europe. Emerg. Inf. Dis. 2005, 11, 225–231. [Google Scholar] [CrossRef]
- Bakoni, T.; Ivanics, E.; Erdélyi, K.; Ursu, K.; Ferenczi, E.; Weissenböck, H.; Nowotny, N. Lineage 1 and 2 strains of encephalitic West Nile virus, central Europe. Emerg. Infect. Dis. 2005, 12, 618–623. [Google Scholar]
- Papa, A.; Politis, C.; Tsoukala, A.; Eglezou, A.; Bakaloudi, V.; Hatzitaki, M.; Tsergouli, K. West Nile virus lineage 2 from blood donor, Greece. Emerg. Infect. Dis. 2012, 18, 688–689. [Google Scholar] [CrossRef]
- Savini, G.; Capelli, G.; Monaco, F.; Monaco, F.; Polci, A.; Russo, F.; di Gennaro, A.; Marini, V.; Teodori, L.; Montarsi, F.; Pinoni, C.; et al. Evidence of West Nile virus lineage 2 circulation in northern Italy. Vet. Microbiol. 2012, 158, 267–273. [Google Scholar] [CrossRef]
- Papa, A.; Bakonyi, T.; Xanthopoulou, K.; Vázquez, A.; Tenorio, A.; Nowotny, N. Genetic characterization of West Nile virus lineage 2, Greece, 2010. Emerg. Infect. Dis. 2011, 17, 920–922. [Google Scholar] [CrossRef]
- Chaintoutis, S.; Chaskopoulou, A.; Chassalevris, T.; Koehler, P.; Papanastassopoulou, M.; Dovas, C. West Nile virus lineage 2 strain in Greece, 2012. Emerg. Infect. Dis. 2013, 19, 827–829. [Google Scholar]
- Papa, A.; Danis, K.; Baka, A.; Bakas, A.; Dougas, G.; Lytras, T.; Theocharopoulos, G.; Chrysagis, D.; Vassiliadou, E.; Kamaria, F.; et al. Ongoing outbreak of West Nile virus infections in human in Greece, July–August 2010. Eur. Surveill. 2010, 15, 1–5. [Google Scholar]
- Sirbu, A.; Ceianu, C.; Panculescu-Gatej, R.; Vazquez, A.; Tenorio, A.; Rebreanu, R.; Niedrig, M.; Nicolescu, G.; Pistol, A. Outbreak of West Nile virus infection in humans, Romania, July to October 2010. Eur. Surveill. 2011, 2, 1–5. [Google Scholar]
- García-Bocanegra, I.; Jaén-Téllez, J.; Napp, S.; Arenas-Montes, A.; Fernández-Morente, M.; Fernández-Molera, V.; Arenas, A. Monitoring of the West Nile virus epidemic in Spain between 2010 and 2011. Transbound. Emerg. Dis. 2012, 59, 448–455. [Google Scholar] [CrossRef]
- Pfeiffer, D. Veterinary Epidemiology: An Introduction; Wiley-Balckwell: Chicester, UK, 2010. [Google Scholar]
- Pavlovsky, E. The Natural Nidality of Transmissible Disease; Levine, N.D., Ed.; University of Illinois Press: Urbana, IL, USA, 1966. [Google Scholar]
- Lambin, E.; Tran, A.; Vanwambeke, S.; Linard, C.; Soti, V. Pathogenic landscapes: Interactions between land, people, disease vectors, and their animal hosts. Int. J. Health Geogr. 2010, 9. [Google Scholar] [CrossRef]
- Ostfeld, R.; Glass, G.; Keesing, F. Spatial epidemiology: An emerging (or re-emerging) discipline. Trend. Ecol. Evolut. 2005, 20, 328–336. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Infectious Diseases of Humans. Dynamics and Control; Oxford University Press: Oxford, UK, 1991; p. 768. [Google Scholar]
- Reiner, R.J.; Perkins, T.; Barker, C.; Niu, T.; Chaves, L.; Ellis, A.; George, D.; le Menach, A.; Pulliam, J.; Bisanzio, D.; et al. A systematic review of mathematical models of mosquito-borne pathogen transmission: 1970–2010. J. R. Soc. Interface 2013, 10. [Google Scholar] [CrossRef]
- Gu, W.; Lampman, R.; Krasavin, N.; Berry, R.; Novak, R.J. Spatio-temporal analyses of West Nile virus transmission in Culex mosquitoes in northern Illinois, USA, 2004. Vector Borne Zoonotic Dis. 2006, 6, 91–98. [Google Scholar] [CrossRef]
- Beroll, H.; Berke, O.; Wilson, J.; Barker, I. Investigating the spatial risk distribution of West Nile virus disease in birds and humans in southern Ontario from 2002 to 2005. Popul. Health Metr. 2007, 5. [Google Scholar] [CrossRef]
- Roberts, R.S.; Foppa, I.M. Prediction of equine based risk of West Nile virus infection based on dead bird surveillance. Vector Borne Zoonotic Dis. 2006, 6, 1–6. [Google Scholar] [CrossRef]
- Esteves, A.; Almeida, A.; Galão, R.; Parreira, R.; Piedade, J.; Rodrigues, J.; Sousa, C.; Novo, M. West Nile virus in Southern Portugal, 2004. Vector Borne Zoonotic Dis. 2004, 5, 410–413. [Google Scholar]
- Savage, H.M.; Ceianu, C.; Nicolescu, G.; Karabatsos, N.; Lanciotti, R.; Vladimirescu, A.; Laiv, L.; Ungureanu, A.; Romanca, C.; Tsai, T.F.; et al. Entomologic and avian investigations of an epidemic of West Nile fever in Romania in 1996, with serologic and molecular characterization of a virus isolate from mosquitoes. Amer. J. Trop. Med. Hyg. 1999, 61, 600–611. [Google Scholar]
- Fyodorova, M.; Savage, H.; Lopatina, J.; Bulgakova, T.; Ivanitsky, A.; Platonova, O.; Platonov, A. Evaluation of potential West Nile virus vectors in Volgograd region, Russia, 2003 (Diptera: Culicidae): Species composition, blood meal host utilization, and virus infection rates of mosquitoes. J. Med. Entomol. 2003, 43, 552–563. [Google Scholar]
- Jourdain, E.; Gauthier-Clerc, M.; Bicout, D.J.; Sabatier, P. Bird migration routes and risk for pathogen dispersion into western Mediterranean wetlands. Emerg. Inf. Dis. 2007, 13, 365–372. [Google Scholar] [CrossRef]
- Bakonyi, T.; Ferenczi, E.; Erdélyi, K.; Kutasi, O.; Csörgő, T.; Seidel, B.; Weissenböck, H.; Brugger, K.; Bán, E.; Nowotny, N. Explosive spread of a neuroinvasive lineage 2 West Nile virus in central Europe, 2008/2009. Vet. Microbiol. 2013, 165, 61–70. [Google Scholar] [CrossRef]
- Durand, B.; Chevalier, V.; Pouillot, R.; Labie, J.; Marendat, I.; Murgue, B.; Zeller, H.; Zientara, S. West Nile virus outbreak in horses, southern France, 2000: Results of a serosurvey. Emerg. Infect. Dis. 2002, 8, 777–782. [Google Scholar] [CrossRef]
- Leblond, A.; Sandoz, A.; Lefebvre, G.; Zeller, H.; Bicout, D. Remote sensing based identification of environmental risk factors associated with West Nile disease in horses in Camargue, France. Prev. Vet. Med. 2007, 79, 20–31. [Google Scholar] [CrossRef]
- Pradier, S.; Leblond, A.; Durand, B. Landcover, landscape structure, and West Nile virus circulation in France. Vector Borne Zoonotic Dis. 2008, 8, 253–263. [Google Scholar] [CrossRef]
- Ahmadnejad, F.; Otarod, V.; Fallah, M.; Lowenski, S.; Sedighi-Moghaddam, R.; Zavareh, A.; Durand, B.; Lecollinet, S.; Sabatier, P. Spread of West Nile virus in Iran: A cross-sectional serosurvey in equines, 2008–2009. Epid. Inf. 2011, 139, 1587–1593. [Google Scholar] [CrossRef]
- García-Bocanegra, I.; Arenas-Montes, A.; Napp, S.; Jaén-Téllez, J.; Fernández-Morente, M.; Fernández-Molera, V.; Arenas, A. Seroprevalence and risk factors associated to West Nile virus in horses from Andalusia, southern Spain. Vet. Microbiol. 2012, 160, 341–346. [Google Scholar] [CrossRef]
- Bargaoui, R.; Lecollinet, S.; Lancelot, R. Mapping the serological prevalence rate of West Nile fever in Equids, Tunisia. Trans. Emerg. Dis. 2013. [Google Scholar] [CrossRef]
- Calistri, P.; Ippoliti, C.; Candeloro, L.; Benjelloun, A.; el Harrak, M.; Bouchra, B.; Danzetta, M.; di Sabatino, D.; Conte, A. Analysis of climatic and environmental variables associated with the occurrence of West Nile virus in Morocco. Prev. Vet. Med. 2013, 110, 549–553. [Google Scholar] [CrossRef]
- Paz, S.; Malkinson, D.; Green, M.S.; Tsioni, G.; Papa, A.; Danis, K.; Sirbu, A.; Ceianu, C.; Katalinn, K.; Ferenczi, E.; Zeller, H.; Semenza, J.C. Permissive summer temperatures of the 2010 European West Nile fever upsurge. Plos One 2013, 8. [Google Scholar] [CrossRef]
- Tran, A.; Gaidet, N.; L’Ambert, G.; Balenghien, T.; Balança, G.; Chevalier, V.; Soti, V.; Ivanes, C.; Etter, E.; Schaffner, F.; et al. The use of remote sensing for the ecological description of multi-host disease systems: A case study on West Nile virus in southern France. Vet. Ital. 2007, 43, 687–697. [Google Scholar]
- Paz, S.; Albersheim, I. Influence of warming tendency on Culex pipiens population abundance and on the probability of West Nile fever outbreaks. EcoHealth 2008, 5, 40–48. [Google Scholar] [CrossRef]
- Bisanzio, D.; Giacobini, M.; Bertolotti, L.; Mosca, A.; Balbo, L.; Kitron, U.; Vazquez-Prokopec, G. Spatio-temporal patterns of distribution of West Nile virus vectors in eastern Piedmont Region, Italy. Parasit. Vectors 2011, 4. [Google Scholar] [CrossRef]
- Roiz, D.; Neteler, M.; Castellani, C.; Arnoldi, D.; Rizzoli, A. Climatic factors driving invasion of the tiger mosquito (Aedes albopictus) into new areas of Trentino, northern Italy. PloS One 2011, 6. [Google Scholar] [CrossRef]
- Rodriguez-Prieto, V.; Martinez-Lopez, B.; Martinez, M.; Munoz, M.J.; Sanchez-Vizcaino, J.M. Identification of suitable areas for West Nile virus outbreaks in equid populations for application in surveillance plans: The example of the Castile and Leon region of Spain. Epidemiol. Infect. 2012, 140, 1617–1631. [Google Scholar] [CrossRef]
- Lopez, G.; Jimenez-Clavero, M.A.; Tejedor, C.G.; Soriguer, R.; Figuerola, J. Prevalence of West Nile virus neutralizing antibodies in Spain is related to the behavior of migratory birds. Vector Borne Zoonotic Dis. 2008, 8, 615–621. [Google Scholar] [CrossRef]
- Figuerola, J.; Jiménez-Clavero, M.; López, G.; Rubio, C.; Soriguer, R.; Gómez-Tejedor, C.; Tenorio, A. Size matters: West Nile Virus neutralizing antibodies in resident and migratory birds in Spain. Vet. Microbiol. 2008, 132, 39–46. [Google Scholar] [CrossRef]
- Balenghien, T.; Fouque, F.; Sabatier, P.; Bicout, D. Theoretical formulation for mosquito host-feeding patterns: Application to a West Nile virus focus of southern France. J. Med. Entomol. 2011, 48, 1076–1090. [Google Scholar] [CrossRef]
- Durand, B.; Balança, G.; Baldet, T.; Chevalier, V. A metapopulation model to simulate West Nile virus circulation in the old world. Vet. Res. 2010, 41. [Google Scholar] [CrossRef]
- Chevalier, V.; Lecollinet, S.; Durand, B. West Nile virus in Europe: A comparison of surveillance system designs in a changing epidemiological context. Vector Born Zoonotic Dis. 2011, 11, 1085–1091. [Google Scholar] [CrossRef]
- Cornel, A.J.; Jupp, P.G.; Blackburn, N.K. Environmental temperature on the vector competence of Culex univittatus (Diptera: Culicidae) for West Nile virus. J. Med. Entomol. 1993, 30, 449–456. [Google Scholar]
- Andrade, C.; Maharaj, P.; Reisen, W.; Brault, A. North American West Nile virus genotype isolates demonstrate differential replicative capacities in response to temperature. J. Gen. Virol. 2011, 92, 2523–2533. [Google Scholar] [CrossRef]
- Epstein, P. Climate change and human health. N. Engl. J. Med. 2005, 353, 1433–1436. [Google Scholar] [CrossRef]
- Epstein, P. West Nile virus and the climate. J. Urban Health 2001, 78, 367–371. [Google Scholar] [CrossRef]
- Dohm, D.; O’Guinn, M.; Turell, M.J. Effect of environmental temperature on the ability of Culex pipiens (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2002, 39, 221–225. [Google Scholar] [CrossRef]
- Kilpatrick, A.; Meola, M.; Moudy, R.; Kramer, L. Temperature, viral genetics, and the transmission of West Nile virus by Culex pipiens mosquitoes. PLoS Pathog. 2008, 4. [Google Scholar] [CrossRef]
- Ruiz, M.; Chaves, L.; Hamer, G.; Sun, T.; Brown, W.; Walker, E.; Haramis, L.; Goldberg, T.; Kitron, U. Local impact of temperature and precipitation on West Nile virus infection in Culex species mosquitoes in northeast Illinois, USA. Parasit. Vectors 2010, 3. [Google Scholar] [CrossRef]
- Dieng, H.; Rahman, G.; Hassan, A.A.; Salmah, M.C.; Satho, T.; Miake, F.; Boots, M.; Sazaly, A. The effects of simulated rainfall on immature population dynamics of Aedes albopictus and female oviposition. Int. J. Biometeorol. 2010, 56, 113–120. [Google Scholar]
- Shaman, J.; Day, J.F.; Stieglitz, M. Drought induced amplification and epidemic transmission of West Nile virus in southern Florida. J. Med. Entomol. 2005, 42, 134–141. [Google Scholar] [CrossRef]
- Chase, J.; Knight, T. Drought-induced mosquito outbreaks in wetlands. Ecol. Lett. 2003, 6, 1017–1024. [Google Scholar] [CrossRef]
- Brownstein, J.S.; Rosen, D.; Purdy, J.; Miller, M.; Merlino, F.; Fish, D. Spatial analysis of West Nile virus: Rapid risk assessment of an introduced vector-borne zoonosis. Vector Borne Zoonotic Dis. 2002, 2, 157–164. [Google Scholar] [CrossRef]
- McGarigal, K.; Marks, B. FRAGSTATS: Spatial Pattern Analysis Program for Quantifying Landscape Structure. Available online: http://www.umass.edu/landeco/pubs/mcgarigal.marks.1995.pdf (accessed on 10 December 2013).
- Bailey, D.; Billeter, R.; Aviron, S.; Schweiger, O. The influence of thematic resolution on metric selection for biodiversity monitoring in agricultural landscapes. Landscape Ecol. 2007, 22, 461–473. [Google Scholar] [CrossRef]
- Guis, H. Géomatique et épidémiologie: Caractérisation des paysages favorables à Culicoides imicola, vecteur de la fièvre catarrhale ovine en Corse, in Sciences de la vie et de la santé; (in French). Université de Franche-Comté: Besançon, France, 2007; p. 418. [Google Scholar]
- Durand, B.; Zanella, G.; Biteau-Coroller, F.; Locatelli, C.; Baurier, F.; Simon, C.; le Dréan, E.; Delaval, J.; Prengère, E.; Beauté, V.; et al. Anatomy of bluetongue virus serotype 8 epizootic wave, France, 2007–2008. Emerg. Inf. Dis. 2010, 16, 1861–1868. [Google Scholar] [CrossRef]
- Guis, H.; Tran, A.; de la Rocque, S.; Baldet, T.; Gerbier, G.; Barragué, B.; Biteau-Coroller, F.; Roger, F.; Viel, J.; Mauny, F. Use of high spatial resolution satellite imagery to characterize landscapes at risk for bluetongue. Vet. Res. 2007, 38, 669–683. [Google Scholar] [CrossRef]
- Kramer, L.; Styer, L.; Ebel, G.D. A global perspective on the epidemiology of West Nile virus. Annu. Rev. Entomol. 2008, 53, 61–84. [Google Scholar] [CrossRef]
- Durand, B.; Dauphin, G.; Zeller, H.; Labie, J.; Schuffenecker, I.; Murri, S.; Moutou, F.; Zientara, S. Serosurvey for West Nile viruses in horses in southern France. Vet. Rec. 2005, 157, 711–713. [Google Scholar] [CrossRef]
- Roiz, D.; Vazquez, A.; Rosà, R.; Muñoz, J.; Arnoldi, D.; Rosso, F.; Figuerola, J.; Tenorio, A.; Rizzoli, A. Blood meal analysis, flavivirus screening, and influence of meteorological variables on the dynamics of potential mosquito vectors of West Nile virus in northern Italy. J. Vector Ecol. 2012, 37, 20–28. [Google Scholar] [CrossRef]
- Chaves, L.; Harrington, L.; Keogh, C.; Nguyen, A.; Kitron, U. Blood feeding patterns of mosquitoes: Random or structured? Front. Zool. 2010, 7. [Google Scholar] [CrossRef]
- Lyimo, I.; Ferguson, H. Ecological and evolutionary determinants of host species choice in mosquito vectors. Trends Parasitol. 2009, 25, 189–226. [Google Scholar] [CrossRef]
- Huang, Z.; de Boer, W.; van Langevelde, F.; Olson, V.; Blackburn, T.; Prins, H. Species’ life-history traits explain interspecific variation in reservoir competence: A possible mechanism underlying the dilution effect. Plos One 2013, 8. [Google Scholar] [CrossRef]
- Chevalier, V.; Reynaud, P.; Lefrançois, T.; Durand, B.; Baillon, F.; Balança, G.; Gaidet, N.; Mondet, B.; Lancelot, R. Predicting West Nile virus seroprevalence in wild birds in Senegal. Vector Borne Zoonotic Dis. 2009, 9, 589–596. [Google Scholar] [CrossRef]
- Hamer, G.; Walker, E.; Brawn, J.; Loss, S.; Ruiz, M.; Goldberg, T.; Schotthoefer, A.; Brown, W.; Wheeler, E.; Kitron, U. Rapid amplification of West Nile virus: The role of hatch-year birds. Vector Borne Zoonotic Dis. 2008, 8, 57–67. [Google Scholar] [CrossRef]
- Burkett-Cadena, N.; Ligon, R.; Liu, M.; Hassan, H.; Hill, G.; Eubanks, M.; Unnasch, T. Vector-host interactions in avian nests: Do mosquitoes prefer nestlings over adults? Am. J. Trop. Med. Hyg. 2010, 83, 395–399. [Google Scholar] [CrossRef]
- Loss, S.; Hamer, G.; Goldberg, T.; Ruiz, M.; Kitron, U.; Walker, E.; Brawn, J. Nestling passerines are not important hosts for amplification of West Nile Virus in Chicago, Illinois. Vector Borne Zoonotic Dis. 2009, 9, 13–18. [Google Scholar] [CrossRef]
- Caillouët, K.; Riggan, A.; Rider, M.; Bulluck, L. Nest mosquito trap quantifies contact rates between nesting birds and mosquitoes. J. Vector Ecol. 2012, 37, 210–215. [Google Scholar] [CrossRef]
- Caillouët, K.; Riggan, A.; Bulluck, L.; Carlson, J.; Sabo, R. Nesting bird “host funnel” increases mosquito-bird contact rate. J. Med. Entomol. 2013, 50, 462–466. [Google Scholar] [CrossRef]
- Chen, C.; Epp, T.; Jenkins, E.; Waldner, C.; Curry, P.; Soos, C. Modeling monthly variation of Culex tarsalis (Diptera: Culicidae) abundance and West Nile Virus infection rate in the Canadian Prairies. Int. J. Environ. Res. Public Health 2013, 10, 3033–3051. [Google Scholar] [CrossRef]
- Baleghien, T. De l’identification des vecteurs de West Nile à la modélisation du risque d'infection dans le Sud de la France. (in French). In ED Ingenierie pour la Santé, la Cognition et l’Environnement; Université Pierre Mendes-France: Grenoble, France, 2006; p. 235. [Google Scholar]
- Ward, M.P. Epidemic West Nile virus encephalomyelitis: A temperature-dependent, spatial model of disease dynamics. Prev. Vet. Med. 2005, 71, 253–264. [Google Scholar] [CrossRef]
- Zou, L.; Miller, S.N.; Schmidtmann, E.T. A GIS tool to estimate West Nile virus risk based on a degree-day model. Environ. Monit. Assess. 2007, 129, 413–420. [Google Scholar] [CrossRef]
- Bowman, C.; Gumel, A.B.; van den Driessche, P.; Wu, J.; Zhu, H. A mathematical model for assessing control strategies against West Nile virus. Bull. Math. Biol. 2005, 67, 1107–1133. [Google Scholar] [CrossRef]
- Cruz-Pacheco, G.; Esteva, L.; Montano-Hirose, J.; Vargas, C. Modelling the dynamics of West Nile virus. Bull. Math. Biol. 2005, 67, 1157–1172. [Google Scholar] [CrossRef]
- Lewis, M.; Renclawowicz, J.; van den Driessche, P. Traveling waves and spread rates for a West Nile virus model. Bull. Math. Biol. 2006, 68, 3–23. [Google Scholar] [CrossRef]
- Rappole, J.; Compton, B.; Leimgruber, P.; Robertson, J.; King, D.; Renner, S. Modeling movement of West Nile virus in the western hemisphere. Vector Borne Zoonotic Dis. 2006, 6, 128–139. [Google Scholar] [CrossRef]
- Wonham, M.J.; de Camino-Beck, T.; Lewis, M.A. An epidemiological model for West Nile virus: Invasion analysis and control applications. Proc. R. Soc. B 2004, 271, 501–507. [Google Scholar] [CrossRef]
- Komar, N.; Panella, N.; Boyce, E. Exposure of domestic mammals to West Nile virus during an outbreak of human encephalitis, New York City, 1999. Emerg. Inf. Dis. 2001, 7, 736–738. [Google Scholar]
- Boroushaki, S.; Malczewski, J. Using the fuzzy majority approach for GIS-based multicriteria group decision-making. Comput. Geosci. 2010, 36, 302–312. [Google Scholar] [CrossRef]
- Hongoh, V.; Hoen, A.; Aenishaenslin, C.; Waaub, J.; Belanger, D.; Michel, P. Spatially explicit multi-criteria decision analysis for managing vector-borne diseases. Int. J. Health Geogr. 2011, 10. [Google Scholar] [CrossRef]
- Stevens, K.; Gilbert, M.; Pfeiffer, D. Modeling habitat suitability for occurence of highky pathogenic avian influenza virsu H5N1 in domestic poultry in Asia: A spatial multicriteria decision analysis approach. Spat. Spatiotemporal Epidemiol. 2012, 4, 1–14. [Google Scholar] [CrossRef]
- Tran, A.; Ippoliti, C.; Balenghien, T.; Conte, A.; Gely, M.; Calistri, P.; Goffredo, M.; Baldet, T.; Chevalier, V. A geographic information system-based multi-criteria evaluation to map areas at risk for Rift Valley fever vector-borne transmission in Italy. Transbound. Emerg. Dis. 2013, 60, 14–23. [Google Scholar]
- Hubalek, Z.; Halouzka, J. West Nile fever—A reemerging mosquito-borne viral disease in Europe. Emerg. Infect. Dis. 1999, 5, 643–650. [Google Scholar] [CrossRef]
- Turell, M.J.; Sardelis, D.J.D.; O’Guinn, M.L. Potential north American vectors of West Nile virus. Ann. N. Y. Acad. Sci. 2001, 951, 317–324. [Google Scholar]
- Kilpatrick, A.; Peters, R.; Dupuis, A.; Jones, M.; Marra, P.; Kramer, L. Predicted and observed mortality from vector-borne disease in small songbirds. Biol. Conserv. 2013, 165, 79–85. [Google Scholar] [CrossRef]
- Thiemann, T.; Wheeler, S.; Barker, C.; Reisen, W. Mosquito host selection varies seasonally with host availability and mosquito density. PLoS Negl. Trop. Dis. 2011, 5. [Google Scholar] [CrossRef]
- Komar, N.; Langevin, S.A.; Hinten, S.R.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.A.; Bunning, M. Experimental infection of north American birds with the New York 1999 strain of West Nile virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef]
- Busquets, N.; Bertran, K.; Costa, T.; Rivas, R.; de la Fuente, J.; Villalba, R.; Solanes, D.; Bensaid, A.; Majó, N.; Pagès, N. Experimental West Nile virus infection in Gyr-Saker hybrid falcons. Vector Borne Zoonotic Dis. 2012, 12, 482–489. [Google Scholar] [CrossRef]
- Dridi, M.; Vangeluwe, D.; Lecollinet, S.; van den Berg, T.; Lambrecht, B. Experimental infection of Carrion crows (Corvus corone) with two European West Nile virus (WNV) strains. Vet. Microbiol. 2013, 165, 160–166. [Google Scholar] [CrossRef]
- Sotelo, E.; Gutierrez-Guzmán, A.; del Amo, J.; Llorente, F.; el Harrak, M.; Pérez-Ramírez, E.; Blanco, J.; Höfle, U.; Jiménez-Clavero, M. Pathogenicity of two recent western Mediterranean West Nile virus isolates in a wild bird species indigenous to Southern Europe: The red-legged partridge. Vet. Res. 2011, 42. [Google Scholar] [CrossRef]
- Ziegler, U.; Angenvoort, J.; Fischer, D.; Fast, C.; Eiden, M.; Rodriguez, A.; Revilla-Fernández, S.; Nowotny, N.; de la Fuente, J.; Lierz, M. Pathogenesis of West Nile virus lineage 1 and 2 in experimentally infected large falcons. Vet. Microbiol. 2013, 161, 263–273. [Google Scholar]
- Roche, B.; Guegan, J.; Bousquet, F. Multi-agent systems in epidemiology: A first step for computational biology in the study of vector-borne disease transmission. BMC Bioinformatics 2008, 9. [Google Scholar] [CrossRef]
- Ezanno, P.; Vergu, E.; Langlais, M.; Gilot-Fromont, E. Modelling the Dynamics of Host-Parasite Interactions: Basic Principles. In New Frontiers of Molecular Epidemiology of Infectious Diseases; Springer: Morand, France, 2012; pp. 79–101. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chevalier, V.; Tran, A.; Durand, B. Predictive Modeling of West Nile Virus Transmission Risk in the Mediterranean Basin: How Far from Landing? Int. J. Environ. Res. Public Health 2014, 11, 67-90. https://doi.org/10.3390/ijerph110100067
Chevalier V, Tran A, Durand B. Predictive Modeling of West Nile Virus Transmission Risk in the Mediterranean Basin: How Far from Landing? International Journal of Environmental Research and Public Health. 2014; 11(1):67-90. https://doi.org/10.3390/ijerph110100067
Chicago/Turabian StyleChevalier, Véronique, Annelise Tran, and Benoit Durand. 2014. "Predictive Modeling of West Nile Virus Transmission Risk in the Mediterranean Basin: How Far from Landing?" International Journal of Environmental Research and Public Health 11, no. 1: 67-90. https://doi.org/10.3390/ijerph110100067
APA StyleChevalier, V., Tran, A., & Durand, B. (2014). Predictive Modeling of West Nile Virus Transmission Risk in the Mediterranean Basin: How Far from Landing? International Journal of Environmental Research and Public Health, 11(1), 67-90. https://doi.org/10.3390/ijerph110100067