Flaviviruses in Europe: Complex Circulation Patterns and Their Consequences for the Diagnosis and Control of West Nile Disease


 ,
, 

Abstract
:1. Introduction
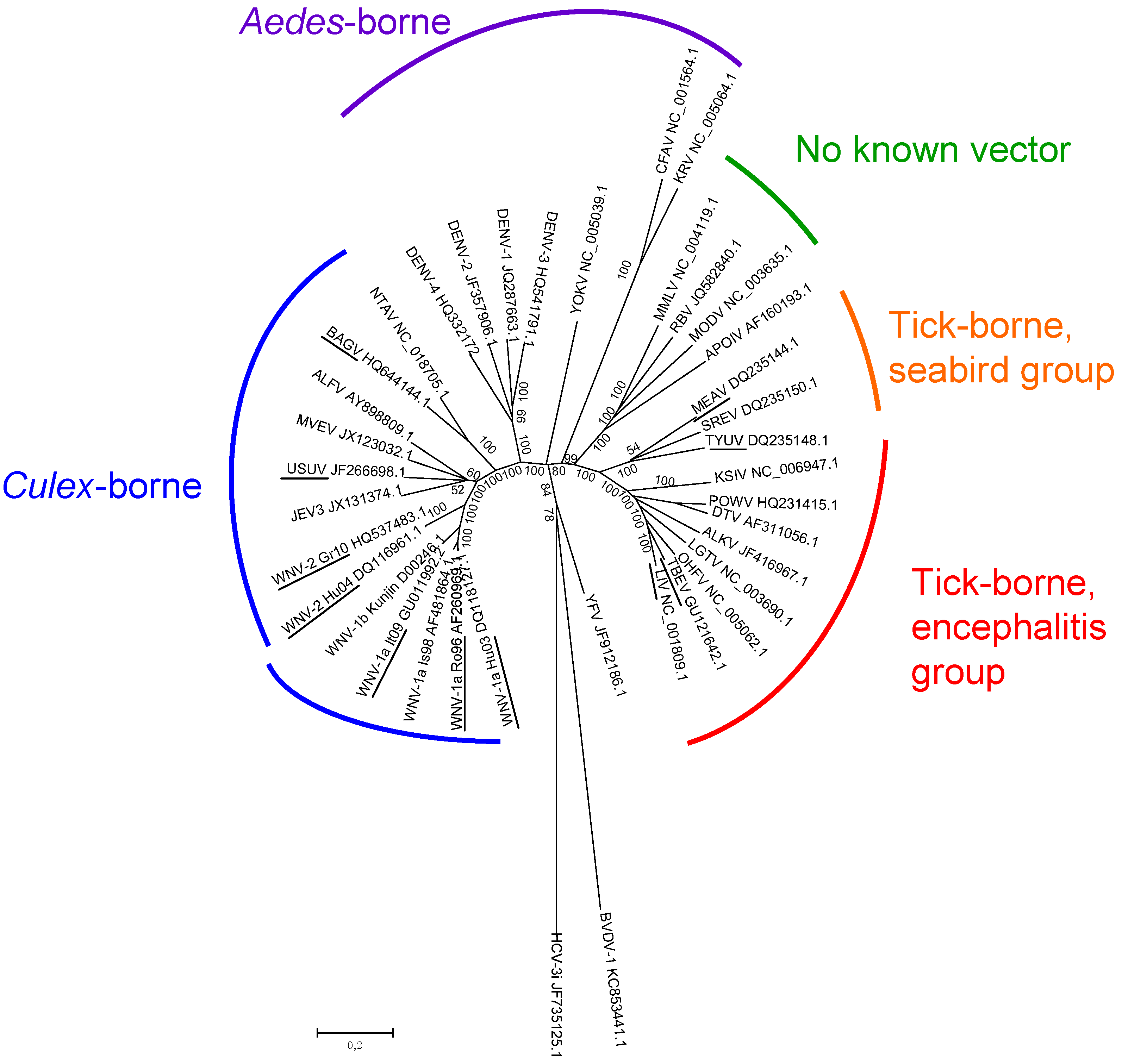
2. Flavivirus Diversity in Europe
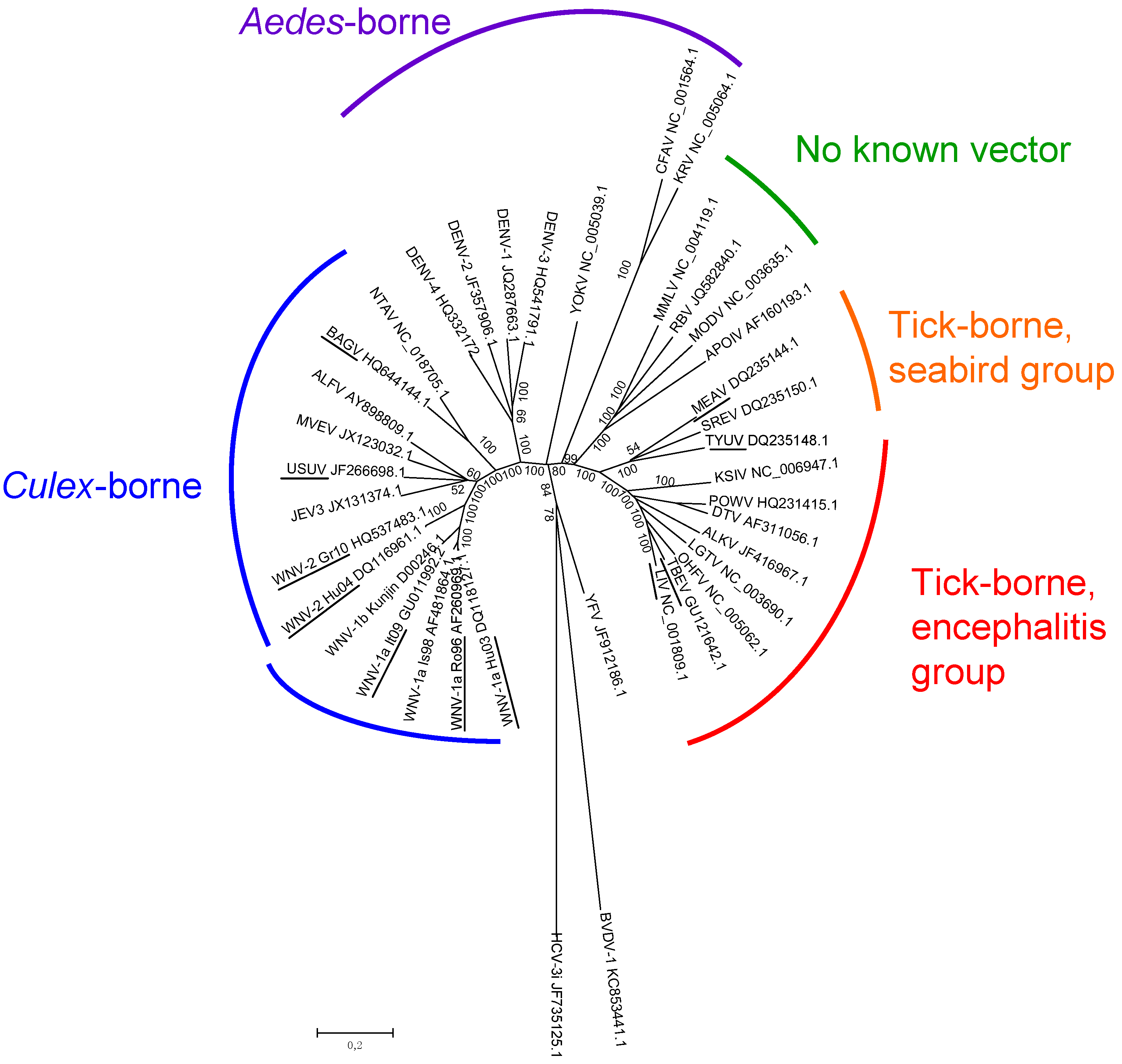
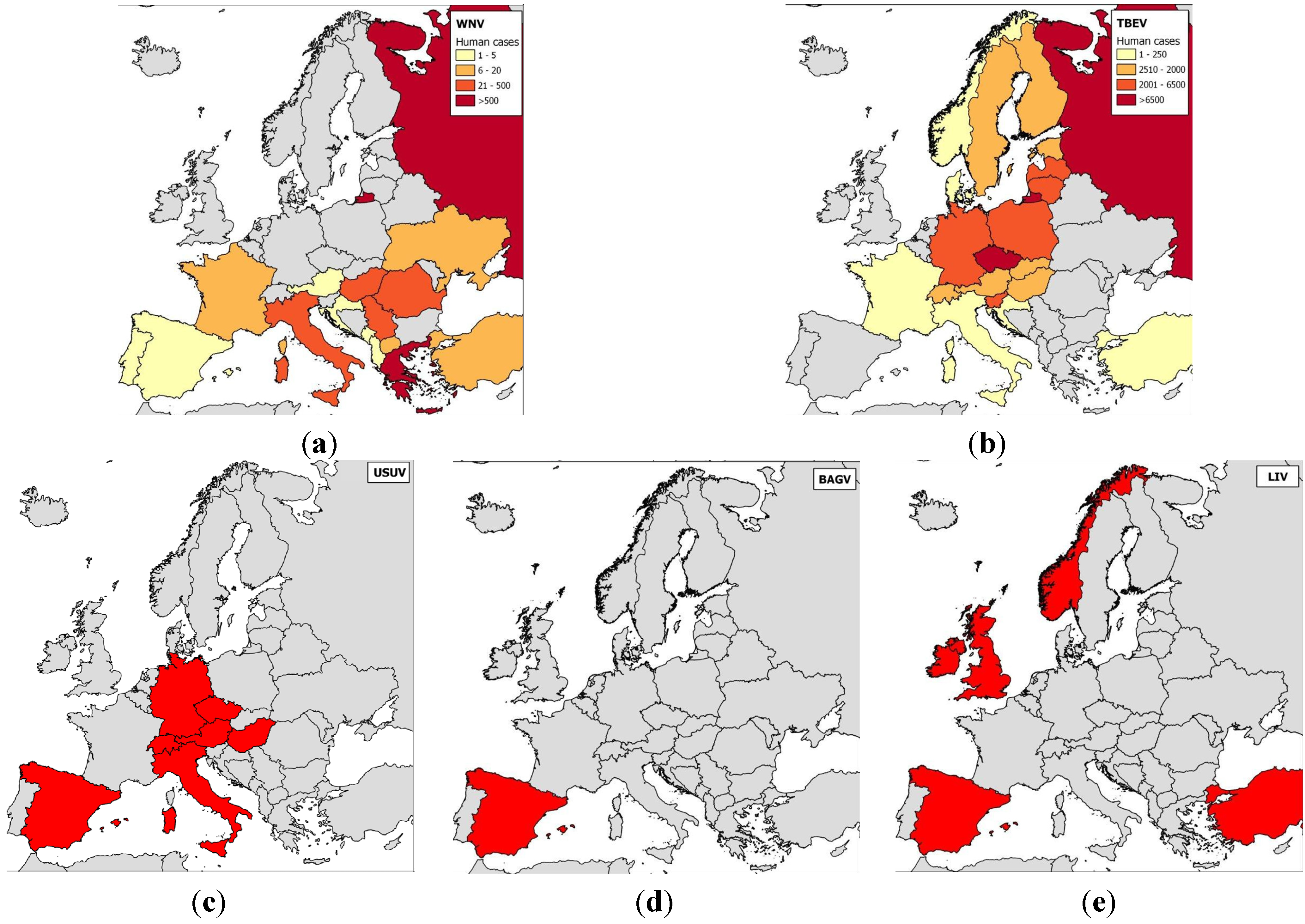
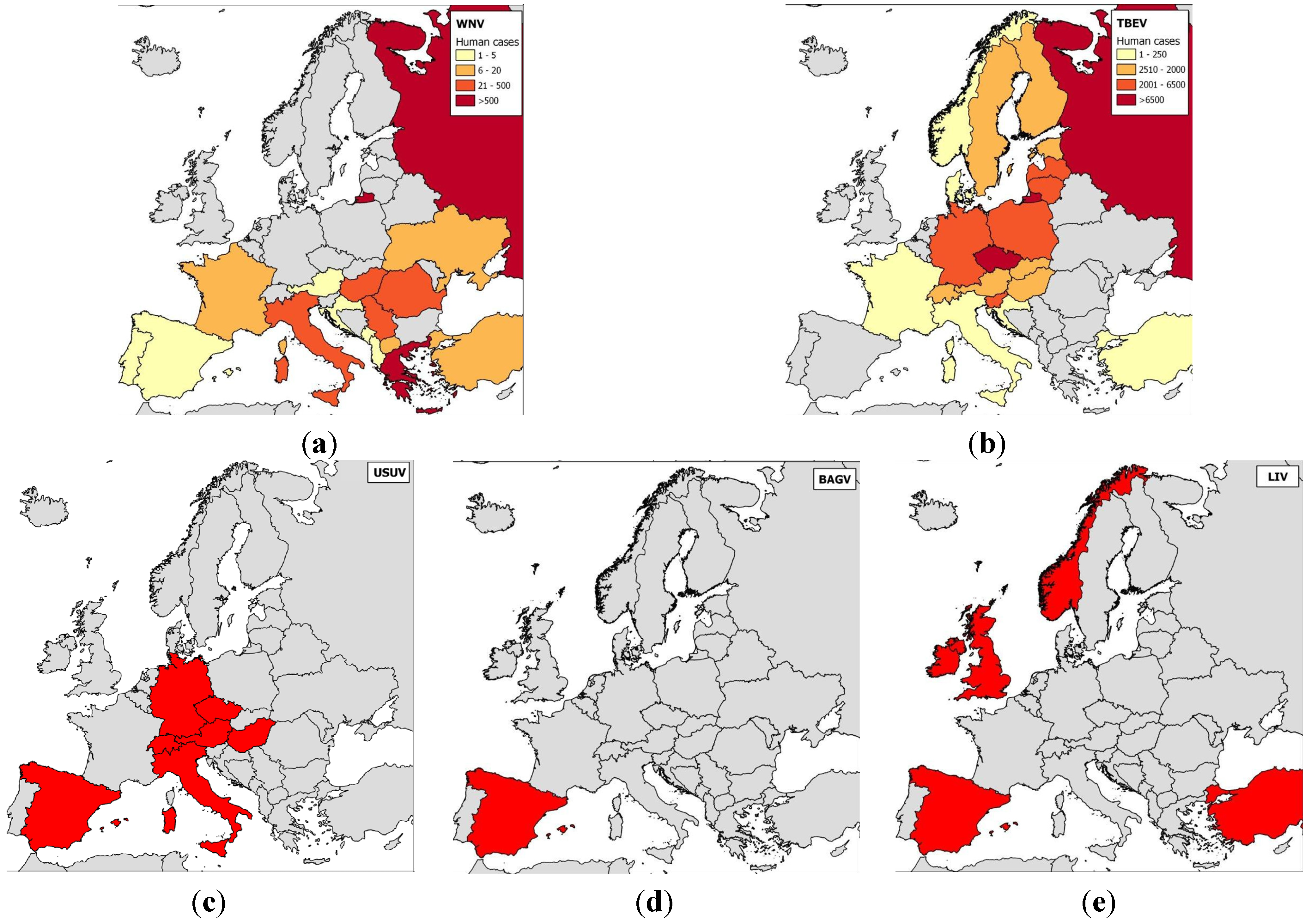
2.1. Epidemiology of Flavivirus Infections in Europe
2.2. Vaccination Programs in Europe

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Vaccine form | Antigen | |
|---|---|---|---|
| Human vaccine | JEV | Inactivated vaccine Ixiaro®—licensed in Europe, JE-VAX®—commercialization stopped | Whole virus |
| Attenuated JEV vaccine (strain SA14-14-2) Available only in China and South Korea | Whole virus | ||
| Chimeric vaccine combining the YFV non structural (NS) proteins and the JEV prM-E Chimeri Vax-JE® Available only in Australia and Thailand | Precursor membrane-Envelope (prM-E) | ||
| TBEV | Inactivated vaccine Ticovac® or Encepur® or FSME-Immune®—licensed in Europe TBE Moscow vaccine® and EnceVir®—licensed in Russia strain from Brazil); attenuated by passage through embryonated eggs Stamaril® or YF-VAX®—licensed in Europe and the rest of the world | Whole virus | |
| YFV | Live-attenuated vaccine (Rockefeller 17D strain or 17DD strain from Brazil); attenuated by passage through embryonated eggs Stamaril® or YF-VAX®—licensed in Europe and the rest of the world | Whole virus | |
| Horse vaccine | JEV | Inactivated vaccine Nisseiken® Available only in Japan | Whole virus |
| WNV | Inactivated vaccine + adjuvant West Nile-Innovator® (U.S.) or Vetera® WNV (U.S.) or EQUIP® WNV (Europe) | Whole virus | |
| Chimeric recombinant canarypox virus Recombitek® Equine WNV (U.S.), Proteq West Nile® (Europe) | prM-E | ||
| Chimeric vaccine combining YFV NS proteins and WNV prM-E PreveNile® Available only in the U.S., recalled in 2010 | prM-E | ||
| DNA vaccine + adjuvant West Nile-Innovator® DNA Available only in the U.S. | prM-E |
3. Serological Cross-Reactions between West Nile and Related Flaviviruses
3.1. Viral Determinants of Serological Reactions
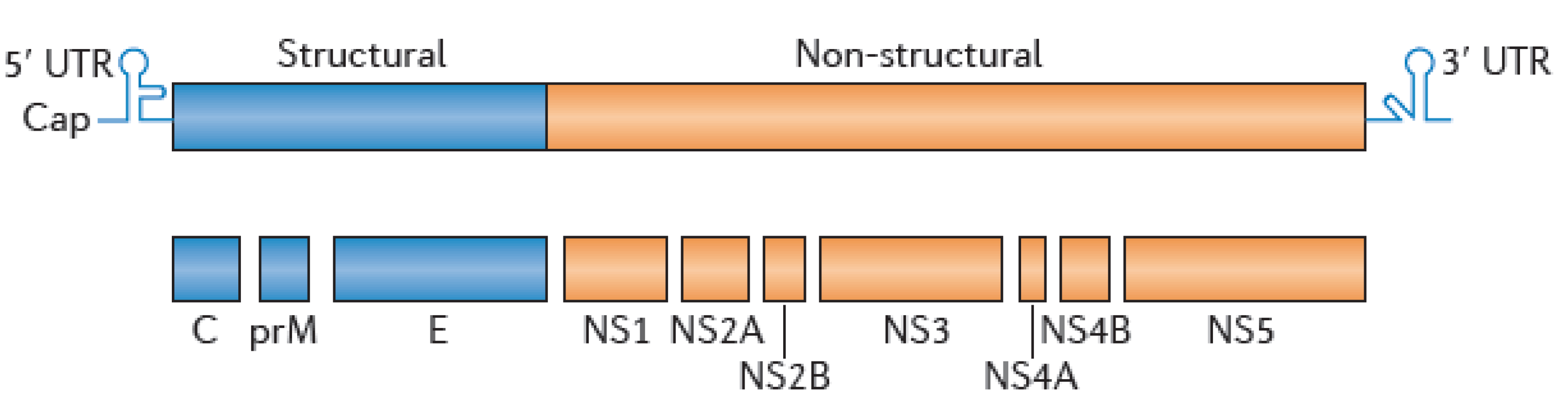
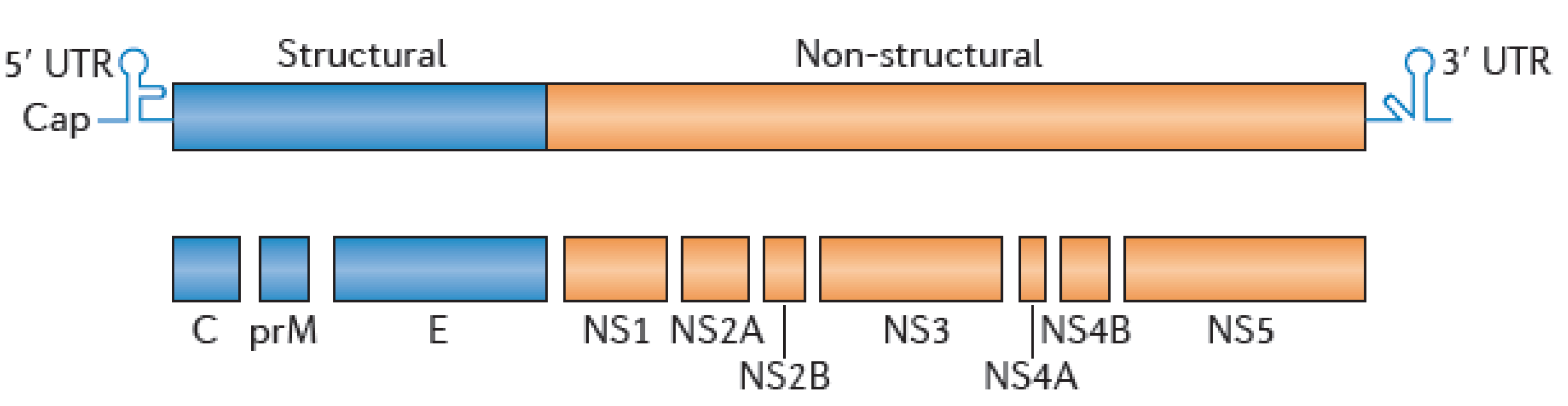
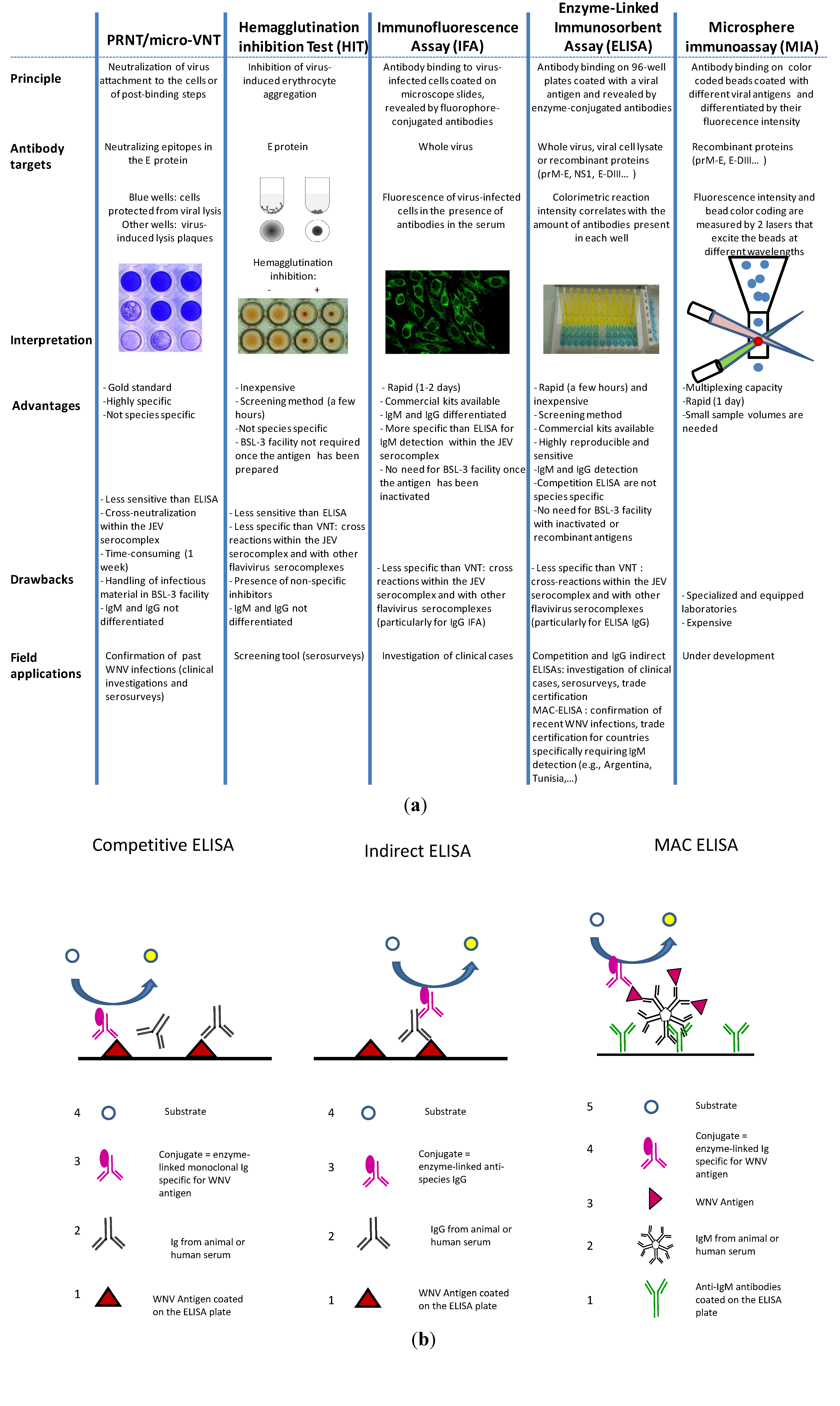
3.2. WNV Serological Diagnostic Tools
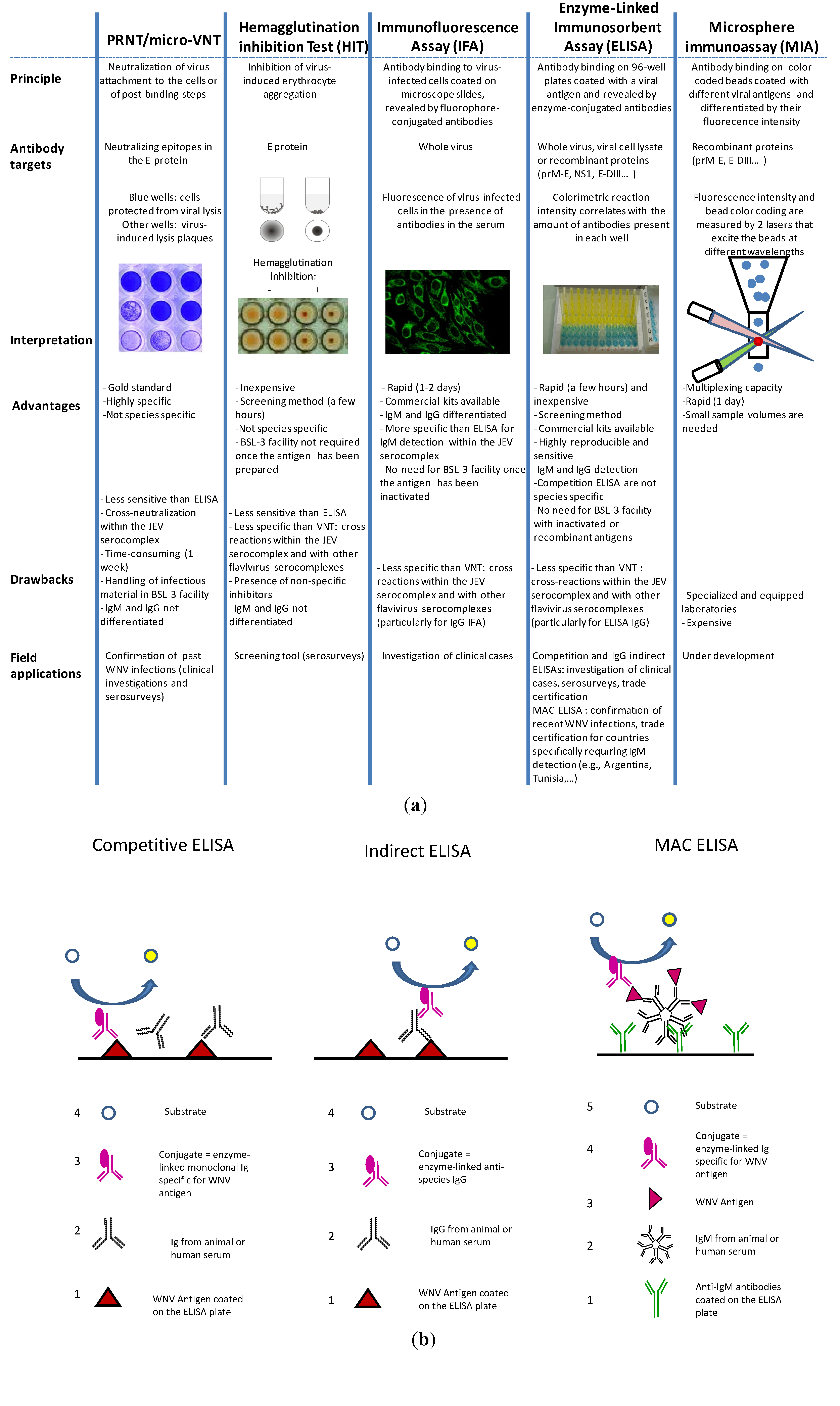
3.2.1. PRNT/Micro-VNT
3.2.2. Hemagglutination Inhibition Tests (HITs) and Complement Fixation Tests (CFTs)
3.2.3. Immunofluorescence Assays
3.2.4. ELISA
3.2.5. Microsphere Immunoassays
3.3. Misinterpretation in the Diagnosis and Surveillance
3.3.1. Difficulties for Accurate Diagnosis
Use of ELISAs
Use of Virus Neutralization Tests
3.3.2. Challenges for WNV Surveillance Programs
4. Antibody-Dependent Cross-Protection or Enhancement among Flaviviruses
4.1. Evaluation of Cross-Protection in Animal Models
| Species (sample size) | Infection or Vaccination | Challenge | Heterologous protection | Publication | |
|---|---|---|---|---|---|
| Birds | House finches (8) | SLEV Kern217 virulent strain | WNV NY99 virulent strain | Complete clinical protection (0/8 death vs. 3/4 deaths in control group), but insufficient virological protection (viremia, mean of 4.6 (2.7–6.4) log10 PFU/mL considered sufficient to infect susceptible mosquitoes) | [170] |
| House finches (8) | WNV NY99 virulent strain | SLEV Kern217 virulent strain | Sterilizing immunity: absence of viremia (0/8 vs. 4/4 in control group) | [170] | |
| Red-winged blackbirds (8) | WNV NY99 virulent strain | JEV virulent Indian strain (genotype III) | Nearly complete virological protection (1/16 viremic birds vs. 16/16 controls) | [171] | |
| Red-winged blackbirds (8) | JEV virulent Vietnamese strain (genotype I) | ||||
| Mammals | Pigs (2) | MVEV virulent OR2 strain | JEV virulent Nakayama strain | Sterilizing immunity: absence of viremia (0/2 vs. 1/1 in control group) | [155] |
| Pigs (2) | WNV mildly virulent KUN HU6774 strain | JEV virulent Nakayama strain | Sterilizing immunity: absence of viremia (0/2 vs. 1/1 in control group) | ||
| Bonnet macaques (3) | Formalin-inactivated JEV strain (733913) | WNV virulent 68856 strain | Complete clinical protection (0/3 death vs. 2/3 deaths in control group) | [172] | |
| Bonnet macaques (5) | Formalin-inactivated WNV strain (68856) | JEV virulent 733913 strain | Partial clinical protection (1/5 death vs. 1/1 death in control group) | ||
| Hamsters (30) | JEV SA14-2-8 vaccine strain | WNV virulent NY99 strain | Complete clinical protection (0/30 death vs. 14/30 deaths in control group). Viremia lowered by about 3 log | [173] | |
| Hamsters (32) | SLEV virulent Be Ar 23379 strain | WNV virulent NY99 strain | Complete clinical protection (0/32 death vs. 14/30 deaths in control group). Viremia lowered by about 4 log | ||
| Hamsters (30) | YF 17D vaccine strain | WNV virulent NY99 strain | Partial clinical protection (4/30 deaths vs. 14/30 deaths in control group). Viremia slightly lowered | ||
| Hamsters (50) | DENV-2 New Guinea C strain | WNV virulent strain | Partial clinical protection (8/50 deaths vs. 50/50 deaths in control group). | [174] | |
| Swiss mice (29) | DENV-2 New Guinea C strain | JEV virulent Peking strain | Complete clinical protection (0/29 deaths vs. 38/60 deaths in control group). Sterilizing immunity: absence of viremia. | [175] | |
| Swiss mice (45) | DENV-2 New Guinea C strain | SLEV virulent Pinellus P 15 strain | Partial clinical protection (17/45 deaths vs. 80/90 deaths in control group). |
4.2. Evaluation of Cross-Protection in the Field
4.3. ADE Risk
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Colpitts, T.M.; Conway, M.J.; Montgomery, R.R.; Fikrig, E. West Nile Virus: Biology, transmission, and human infection. Clin. Microbiol. Rev. 2012, 25, 635–648. [Google Scholar] [CrossRef]
- Dauphin, G.; Zientara, S.; Zeller, H.; Murgue, B. West Nile: Worldwide current situation in animals and humans. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 343–355. [Google Scholar] [CrossRef]
- Porter, R.S.; Leblond, A.; Lecollinet, S.; Tritz, P.; Cantile, C.; Kutasi, O.; Zientara, S.; Pradier, S.; van Galen, G.; Speybroek, N.; Saegerman, C. Clinical diagnosis of West Nile Fever in Equids by classification and regression tree (CART) analysis and comparative study of clinical appearance in three European countries. Transbound. Emerg. Dis. 2011, 58, 197–205. [Google Scholar] [CrossRef]
- Dauphin, G.; Zientara, S. West Nile virus: Recent trends in diagnosis and vaccine development. Vaccine 2007, 25, 5563–5576. [Google Scholar] [CrossRef]
- de Madrid, A.T.; Porterfield, J.S. The flaviviruses (group B arboviruses): A cross-neutralization study. J. Gen. Virol. 1974, 23, 91–96. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Available online: http://www.cdc.gov/westnile/index.html (accessed on 4 Novermber 2013).
- Kilpatrick, A.M. Globalization, land use, and the invasion of West Nile virus. Science 2011, 334, 323–327. [Google Scholar] [CrossRef]
- Murray, K.O.; Mertens, E.; Despres, P. West Nile virus and its emergence in the United States of America. Vet. Res. 2010, 41, 67–81. [Google Scholar] [CrossRef]
- LaDeau, S.L.; Kilpatrick, A.M.; Marra, P.P. West Nile virus emergence and large-scale declines of North American bird populations. Nature 2007, 447, 710–713. [Google Scholar] [CrossRef]
- Blitvich, B.J. Transmission dynamics and changing epidemiology of West Nile virus. Anim. Health Res. Rev. 2008, 9, 71–86. [Google Scholar] [CrossRef]
- Brault, A.C. Changing patterns of West Nile virus transmission: Altered vector competence and host susceptibility. Vet. Res. 2009, 40. [Google Scholar] [CrossRef]
- Dridi, M.; Vangeluwe, D.; Lecollinet, S.; van den Berg, T.; Lambrecht, B. Experimental infection of Carrion crows (Corvus corone) with two European West Nile virus (WNV) strains. Vet. Microbiol. 2013, 165, 160–166. [Google Scholar] [CrossRef]
- Brault, A.C.; Langevin, S.A.; Bowen, R.A.; Panella, N.A.; Biggerstaff, B.J.; Miller, B.R.; Komar, N. Differential virulence of West Nile strains for American crows. Emerg. Infect. Dis. 2004, 10, 2161–2168. [Google Scholar] [CrossRef]
- Reiter, P. West Nile virus in Europe: Understanding the present to gauge the future. Euro. Surveill. 2010, 15, 19508:1–19508:7. [Google Scholar]
- Fonseca, D.M.; Keyghobadi, N.; Malcolm, C.A.; Mehmet, C.; Schaffner, F.; Mogi, M.; Fleischer, R.C.; Wilkerson, R.C. Emerging vectors in the Culex pipiens complex. Science 2004, 303, 1535–1538. [Google Scholar] [CrossRef]
- Gould, E.A. Implications for Northern Europe of the emergence of West Nile virus in the USA. Epidemiol. Infect. 2003, 131, 583–589. [Google Scholar] [CrossRef]
- Swaddle, J.P.; Calos, S.E. Increased avian diversity is associated with lower incidence of human West Nile infection: Observation of the dilution effect. PloS One 2008, 3. [Google Scholar] [CrossRef]
- Ezenwa, V.O.; Godsey, M.S.; King, R.J.; Guptill, S.C. Avian diversity and West Nile virus: Testing associations between biodiversity and infectious disease risk. Proc. Biol. Sci. 2006, 273, 109–117. [Google Scholar] [CrossRef]
- Durand, B.; Dauphin, G.; Zeller, H.; Labie, J.; Schuffenecker, I.; Murri, S.; Moutou, F.; Zientara, S. Serosurvey for West Nile virus in horses in southern France. Vet. Rec. 2005, 157, 711–713. [Google Scholar] [CrossRef]
- West Nile Fever. Available online: http://ecdc.europa.eu/en/healthtopics/west_nile_fever/Pages/index.aspx (accessed on 4 November 2013).
- Pradier, S.; Lecollinet, S.; Leblond, A. West Nile virus epidemiology and factors triggering change in its distribution in Europe. Rev. Sci. Tech. 2012, 31, 829–844. [Google Scholar]
- Lopez, G.; Jimenez-Clavero, M.A.; Vazquez, A.; Soriguer, R.; Gomez-Tejedor, C.; Tenorio, A.; Figuerola, J. Incidence of West Nile virus in birds arriving in wildlife rehabilitation centers in southern Spain. Vector-Borne Zoonotic Dis. 2011, 11, 285–290. [Google Scholar] [CrossRef] [Green Version]
- Eidson, M.; Komar, N.; Sorhage, F.; Nelson, R.; Talbot, T.; Mostashari, F.; McLean, R. Crow deaths as a sentinel surveillance system for West Nile virus in the northeastern United States, 1999. Emerg. Infect. Dis. 2001, 7, 615–620. [Google Scholar]
- Wodak, E.; Richter, S.; Bago, Z.; Revilla-Fernandez, S.; Weissenbock, H.; Nowotny, N.; Winter, P. Detection and molecular analysis of West Nile virus infections in birds of prey in the eastern part of Austria in 2008 and 2009. Vet. Microbiol. 2011, 149, 358–366. [Google Scholar] [CrossRef]
- Chevalier, V.; Lecollinet, S.; Durand, B. West Nile virus in Europe: A comparison of surveillance system designs in a changing epidemiological context. Vector-Borne Zoonotic Dis. 2011, 11, 1085–1091. [Google Scholar] [CrossRef]
- Angelini, P.; Tamba, M.; Finarelli, A.C.; Bellini, R.; Albieri, A.; Bonilauri, P.; Cavrini, F.; Dottori, M.; Gaibani, P.; Martini, E.; et al. West Nile virus circulation in Emilia-Romagna, Italy: The integrated surveillance system 2009. Euro. Surveill. 2010, 15, 19547:1–19547:5. [Google Scholar]
- Calzolari, M.; Monaco, F.; Montarsi, F.; Bonilauri, P.; Ravagnan, S.; Bellini, R.; Cattoli, G.; Cordioli, P.; Cazzin, S.; Pinoni, C.; et al. New incursions of West Nile virus lineage 2 in Italy in 2013: The value of the entomological surveillance as early warning system. Vet. Ital. 2013, 2013. [Google Scholar] [CrossRef]
- Chaskopoulou, A.; Dovas, C.I.; Chaintoutis, S.C.; Kashefi, J.; Koehler, P.; Papanastassopoulou, M. Detection and early warning of West Nile Virus circulation in central Macedonia, Greece, using sentinel chickens and mosquitoes. Vector-Borne Zoonotic Dis. 2013, 13, 723–732. [Google Scholar] [CrossRef]
- Niedrig, M.; Linke, S.; Zeller, H.; Drosten, C. First international proficiency study on West Nile virus molecular detection. Clin. Chem. 2006, 52, 1851–1854. [Google Scholar] [CrossRef]
- Vazquez, A.; Sanchez-Seco, M.P.; Ruiz, S.; Molero, F.; Hernandez, L.; Moreno, J.; Magallanes, A.; Tejedor, C.G.; Tenorio, A. Putative new lineage of west nile virus, Spain. Emerg. Infect. Dis. 2010, 16, 549–552. [Google Scholar] [CrossRef]
- Bakonyi, T.; Ivanics, E.; Erdélyi, K.; Ursu, K.; Ferenczi, E.; Weissenböck, H.; Nowotny, N. Lineage 1 and 2 strains of encephalitic West Nile Virus, central Europe. Emerg. Infect. Dis. 2006, 12, 618–623. [Google Scholar] [CrossRef]
- Bakonyi, T.; Hubalek, Z.; Rudolf, I.; Nowotny, N. Novel flavivirus or new lineage of West Nile virus, central Europe. Emerg. Infect. Dis. 2005, 11, 225–231. [Google Scholar] [CrossRef]
- Lvov, D.K.; Butenko, A.M.; Gromashevsky, V.L.; Larichev, V.P.; Gaidamovich, S.Y.; Vyshemirsky, O.I.; Zhukov, A.N.; Lazorenko, V.V.; Salko, V.N.; Kovtunov, A.I.; et al. Isolation of two strains of West Nile virus during an outbreak in southern Russia, 1999. Emerg. Infect. Dis. 2000, 6, 373–376. [Google Scholar] [CrossRef]
- Sirbu, A.; Celanu, C.S.; Panculescu-Gatej, R.I.; Vazquez, A.; Tenorio, A.; Rebreanu, R.; Niedrig, M.; Nicolescu, G.; Pistol, A. Outbreak of West Nile virus infection in humans, Romania, July to October 2010. Euro. Surveill. 2011, 16, 19762:1–19762:5. [Google Scholar]
- May, F.J.; Davis, C.T.; Tesh, R.B.; Barrett, A.D.T. Phylogeography of West Nile virus: From the cradle of evolution in Africa to Eurasia, Australia, and the Americas. J. Virol. 2011, 85, 2964–2974. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Gubler, D.J.; Petersen, L.R. Emerging flaviviruses: The spread and resurgence of Japanese encephalitis, West Nile and dengue viruses. Nat. Med. 2004, 10, S98–S109. [Google Scholar] [CrossRef]
- Heinz, F.X.; Stiasny, K. Flaviviruses and flavivirus vaccines. Vaccine 2012, 30, 4301–4306. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Horton, D.L.; Johnson, N.; Li, L.; Barrett, A.D.T.; Smith, D.J.; Galbraith, S.E.; Solomon, T.; Fooks, A.R. Flavivirus-induced antibody cross-reactivity. J. Gen. Virol. 2011, 92, 2821–2829. [Google Scholar] [CrossRef]
- Koraka, P.; Zeller, H.; Niedrig, M.; Osterhaus, A.D.M.E.; Groen, J. Reactivity of serum samples from patients with a flavivirus infection measured by immunofluorescence assay and ELISA. Microbes Infect. 2002, 4, 1209–1215. [Google Scholar] [CrossRef]
- Rushton, J.O.; Lecollinet, S.; Hubalek, Z.; Svobodova, P.; Lussy, H.; Nowotny, N. Tick-borne encephalitis virus in horses, Austria, 2011. Emerg. Infect. Dis. 2013, 19, 635–637. [Google Scholar] [CrossRef]
- Ziegler, U.; Angenvoort, J.; Klaus, C.; Nagel-Kohl, U.; Sauerwald, C.; Thalheim, S.; Horner, S.; Braun, B.; Kenklies, S.; Tyczka, J.; et al. Use of competition ELISA for monitoring of West Nile virus infections in horses in Germany. Int. J. Environ. Res. Public Health 2013, 10, 3112–3120. [Google Scholar] [CrossRef]
- ICTV 2012 Master Species List (MSL). Available online: http://www.ictvonline.org/ (accessed on 5 November 2013).
- Gaunt, M.W.; Sall, A.A.; de Lamballerie, X.; Falconar, A.K.I.; Dzhivanian, T.I.; Gould, E.A. Phylogenetic relationships of flaviviruses correlate with their epidemiology, disease association and biogeography. J. Gen. Virol. 2001, 82, 1867–1876. [Google Scholar]
- Kuno, G.; Chang, G.-J.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar]
- Calisher, C.H.; Karabatsos, N.; Dalrymple, J.M.; Shope, R.E.; Porterfield, J.S.; Westaway, E.G.; Brandt, W.E. Antigenic relationships between flaviviruses as determined by cross-neutralization tests with polyclonal antisera. J. Gen. Virol. 1989, 70, 37–43. [Google Scholar] [CrossRef]
- Gould, E.A.; Moss, S.R.; Turner, S.L. Evolution and Dispersal of Encephalitic Flaviviruses. In Emergence and Control of Zoonotic Viral Encephalitis, Les Pensières, France 6–8 April 2003; Springer-Verlag Wien: Berlin, Germany, 2004; pp. 65–84. [Google Scholar]
- Cook, S.; Moureau, G.; Kitchen, A.; Gould, E.A.; de Lamballerie, X.; Holmes, E.C.; Harbach, R.E. Molecular evolution of the insect-specific flaviviruses. J. Gen. Virol. 2012, 93, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Solomon, T.; Dung, N.M.; Vaughn, D.W.; Kneen, R.; Thao, L.T.T.; Raengsakulrach, B.; Loan, H.T.; Day, N.P.J.; Farrar, J.; Myint, K.S.A.; et al. Neurological manifestations of dengue infection. Lancet 2000, 355, 1053–1059. [Google Scholar] [CrossRef]
- Gould, E.A.; Zanotto, P.A.; Holmes, E.C. The genetic evolution of flaviviruses. In Factors in the Emergence of Arbovirus Diseases; Saluzzo, J.F., Dodet, B., Eds.; Elsevier: Amsterdam, The Netherlands, 1997; pp. 51–63. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Ergunay, K.; Saygan, M.B.; Aydogan, S.; Litzba, N.; Sener, B.; Lederer, S.; Niedrig, M.; Hascelik, G.; Us, D. Confirmed exposure to tick-borne encephalitis virus and probable human cases of tick-borne encephalitis in Central/Northern Anatolia, Turkey. Zoonoses Public Health 2011, 58, 220–227. [Google Scholar] [CrossRef]
- Suss, J. Tick-borne encephalitis 2010: Epidemiology, risk areas, and virus strains in Europe and Asia—An overview. Ticks Tick-Borne Dis. 2011, 2, 2–15. [Google Scholar] [CrossRef]
- Dobler, G. Zoonotic tick-borne flaviviruses. Vet. Microbiol. 2010, 140, 221–228. [Google Scholar] [CrossRef]
- M’Fadzean, J. Louping-ill in sheep. J. Comp. Pathol. Ther. 1894, 7, 207–209. [Google Scholar] [CrossRef]
- Lvov, D.K. Omsk Hemorrhagic Fever. In The Arboviruses: Epidemiology and Ecology, Vol. III; Monath, T.P., Ed.; CRC Press, Inc.: Boca Raton, FL, USA, 1988; pp. 205–216. [Google Scholar]
- Chastel, C.; Main, A.J.; Guiguen, C.; le Lay, G.; Quillien, M.C.; Monnat, J.Y.; Beaucournu, J.C. The isolation of Meaban virus, a new Flavivirus from the seabird tick Ornithodoros (Alectorobius) maritimus in France. Arch. Virol. 1985, 83, 129–140. [Google Scholar] [CrossRef]
- Hubalek, Z.; Halouzka, J. Arthropod-Borne Viruses of Vertebrates in Europe; Institute of Landscape Ecology of the Academy of Sciences of the Czech Republic: Prague, Czech Republic, 1996. [Google Scholar]
- Weissenbock, H.; Hubalek, Z.; Bakonyi, T.; Nowotny, N. Zoonotic mosquito-borne flaviviruses: Worldwide presence of agents with proven pathogenicity and potential candidates of future emerging diseases. Vet. Microbiol. 2010, 140, 271–280. [Google Scholar] [CrossRef]
- Filipe, A.R.; Pinto, M.R. Survey for antibodies to arboviruses in serum of animals from southern Portugal. Am. J. Trop. Med. Hyg. 1969, 18, 423–426. [Google Scholar]
- Joubert, L.; Oudar, J.; Hannoun, C.; Beytout, D.; Corniou, B.; Guillon, J.C.; Panthier, R. Epidemiology of the West Nile virus: Study of a focus in Camargue. IV. Meningo-encephalomyelitis of the horse. Ann. Inst. Pasteur (Paris) 1970, 118, 239–247. [Google Scholar]
- Malkinson, M.; Banet, C.; Weisman, Y.; Pokamunski, S.; King, R.; Drouet, M.T.; Deubel, V. Introduction of West Nile virus in the Middle East by migrating white storks. Emerg. Infect. Dis. 2002, 8, 392–397. [Google Scholar] [CrossRef]
- Papa, A.; Danis, K.; Baka, A.; Bakas, A.; Dougas, G.; Lytras, T.; Theocharopoulos, G.; Chrysagis, D.; Vassiliadou, E.; Kamaria, F.; et al. Ongoing outbreak of West Nile virus infections in humans in Greece, July–August 2010. Euro. Surveill. 2010, 15, 20–29. [Google Scholar]
- Bakonyi, T.; Ferenczi, E.; Erdelyi, K.; Kutasi, O.; Csorgo, T.; Seidel, B.; Weissenbock, H.; Brugger, K.; Ban, E.; Nowotny, N. Explosive spread of a neuroinvasive lineage 2 West Nile virus in Central Europe, 2008/2009. Vet. Microbiol. 2013, 165, 61–70. [Google Scholar] [CrossRef]
- Onishchenko, G.G.; Lipnitskii, A.V.; Alekseev, V.V.; Antonov, V.A.; Kriuchkova, T.P.; Krutogolovova, T.A. Epidemiologic situation of West Nile fever in Russia in 2010. Zh. Mikrobiol. Epidemiol. Immunobiol. 2011, 3, 115–120. [Google Scholar]
- Calistri, P.; Monaco, F.; Savini, G.; Guercio, A.; Purpari, G.; Vicari, D.; Cascio, S.; Lelli, R. Further spread of West Nile virus in Italy. Vet. Ital. 2010, 46, 471–474. [Google Scholar]
- Weissenbock, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu virus, Italy, 1996. Emerg. Infect. Dis. 2013, 19, 274–277. [Google Scholar] [CrossRef]
- Weissenbock, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African mosquito-borne flavivirus of the Japanese encephalitis virus group, central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef]
- Meister, T.; Lussy, H.; Bakonyi, T.; Sikutova, S.; Rudolf, I.; Vogl, W.; Winkler, H.; Frey, H.; Hubalek, Z.; Nowotny, N.; et al. Serological evidence of continuing high Usutu virus (Flaviviridae) activity and establishment of herd immunity in wild birds in Austria. Vet. Microbiol. 2008, 127, 237–248. [Google Scholar] [CrossRef]
- Becker, N.; Jost, H.; Ziegler, U.; Eiden, M.; Hoper, D.; Emmerich, P.; Fichet-Calvet, E.; Ehichioya, D.U.; Czajka, C.; Gabriel, M.; et al. Epizootic emergence of Usutu virus in wild and captive birds in Germany. PloS One 2012, 7, e32604. [Google Scholar] [CrossRef]
- Busquets, N.; Alba, A.; Allepuz, A.; Aranda, C.; Nunez, J.I. Usutu virus sequences in Culex pipiens (Diptera : Culicidae), Spain. Emerg. Infect. Dis. 2008, 14, 861–863. [Google Scholar] [CrossRef]
- Höfle, U.; Gamino, V.; Fernández-de-Mera, I.G.; Mangold, A.J.; Ortíz, J.-A.; de la Fuente, J. Usuto virus in migratory song thrushes, Spain. Emerg. Infect. Dis. 2013, 19, 1173–1175. [Google Scholar] [CrossRef] [Green Version]
- Hubalek, Z.; Rudolf, I.; Capek, M.; Bakonyi, T.; Betasova, L.; Nowotny, N. Usutu virus in Blackbirds (Turdus merula), Czech Republic, 2011–2012. Emerg. Dis. 2012, 2012. [Google Scholar] [CrossRef]
- Bakonyi, T.; Erdelyi, K.; Ursu, K.; Ferenczi, E.; Csorgo, T.; Lussy, H.; Chvala, S.; Bukovsky, C.; Meister, T.; Weissenbock, H.; et al. Emergence of Usutu virus in Hungary. J. Clin. Microbiol. 2007, 45, 3870–3874. [Google Scholar] [CrossRef]
- Manarolla, G.; Bakonyi, T.; Gallazzi, D.; Crosta, L.; Weissenbock, H.; Dorrestein, G.M.; Nowotny, N. Usutu virus in wild birds in northern Italy. Vet. Microbiol. 2010, 141, 159–163. [Google Scholar] [CrossRef]
- Steinmetz, H.W.; Bakonyi, T.; Weissenbock, H.; Hatt, J.-M.; Eulenberger, U.; Robert, N.; Hoop, R.; Nowotny, N. Emergence and establishment of Usutu virus infection in wild and captive avian species in and around Zurich, Switzerland—Genomic and pathologic comparison to other central European outbreaks. Vet. Microbiol. 2011, 148, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Vittecoq, M.; Lecollinet, S.; Jourdain, E.; Thomas, F.; Blanchon, T.; Arnal, A.; Lowenski, S.; Gauthier-Clerc, M. Recent circulation of West Nile virus and potentially other closely related flaviviruses in Southern France. Vector-Borne Zoonotic Dis. 2013, 13, 610–613. [Google Scholar] [CrossRef]
- Barbic, L.; Vilibic-Cavlek, T.; Listes, E.; Stevanovic, V.; Gjenero-Margan, I.; Ljubin-Sternak, S.; Pem-Novosel, I.; Listes, I.; Mlinaric-Galinovic, G.; Di Gennaro, A.; et al. Demonstration of Usutu virus antibodies in Horses, Croatia. Vector-Borne Zoonotic Dis. 2013, 13, 772–774. [Google Scholar] [CrossRef]
- Aguero, M.; Fernandez-Pinero, J.; Buitrago, D.; Sanchez, A.; Elizalde, M.; San Miguel, E.; Villalba, R.; Llorente, F.; Jimenez-Clavero, M.A. Bagaza virus in partridges and pheasants, Spain, 2010. Emerg. Infect. Dis. 2011, 17, 1498–1501. [Google Scholar]
- Bondre, V.P.; Sapkal, G.N.; Yergolkar, P.N.; Fulmali, P.V.; Sankararaman, V.; Ayachit, V.M.; Mishra, A.C.; Gore, M.M. Genetic characterization of Bagaza virus (BAGV) isolated in India and evidence of anti-BAGV antibodies in sera collected from encephalitis patients. J. Gen. Virol. 2009, 90, 2644–2649. [Google Scholar] [CrossRef]
- Cerutti, F.; Giacobini, M.; Mosca, A.; Grasso, I.; Rambozzi, L.; Rossi, L.; Bertolotti, L. Evidence of mosquito-transmitted flavivirus circulation in Piedmont, north-western Italy. Parasites Vectors 2012, 5, 99. [Google Scholar] [CrossRef]
- Sanchez-Seco, M.-P.; Vazquez, A.; Collao, X.; Hernandez, L.; Aranda, C.; Ruiz, S.; Escosa, R.; Marques, E.; Bustillo, M.-A.; Molero, F.; et al. Surveillance of arboviruses in Spanish wetlands: Detection of new flavi- and phleboviruses. Vector-Borne Zoonotic Dis. 2010, 10, 203–206. [Google Scholar] [CrossRef]
- Ravanini, P.; Huhtamo, E.; Ilaria, V.; Crobu, M.G.; Nicosia, A.M.; Servino, L.; Rivasi, F.; Allegrini, S.; Miglio, U.; Magri, A.; et al. Japanese encephalitis virus RNA detected in Culex pipiens mosquitoes in Italy. Euro. Surveill. 2012, 17, 20221:1–20221:4. [Google Scholar]
- Huhtamo, E.; Putkuri, N.; Kurkela, S.; Manni, T.; Vaheri, A.; Vapalahti, O.; Uzcategui, N.Y. Characterization of a novel flavivirus from mosquitoes in northern europe that is related to mosquito-borne flaviviruses of the tropics. J. Virol. 2009, 83, 9532–9540. [Google Scholar] [CrossRef]
- Calzolari, M.; Bonilauri, P.; Bellini, R.; Albieri, A.; Defilippo, F.; Maioli, G.; Galletti, G.; Gelati, A.; Barbieri, I.; Tamba, M.; et al. Evidence of simultaneous circulation of West Nile and Usutu viruses in mosquitoes sampled in Emilia-Romagna region (Italy) in 2009. PloS One 2010, 5, e14324. [Google Scholar] [CrossRef]
- Llorente, F.; Pérez-Ramírez, E.; Fernández-Pinero, J.; Soriguer, R.; Figuerola, J.; Jiménez-Clavero, M.A. Flaviviruses in game birds, Southern Spain, 2011–2012. Emerg. Infect. Dis. 2013, 19, 1023–1025. [Google Scholar] [CrossRef]
- Brown, E.B.E.; Adkin, A.; Fooks, A.R.; Stephenson, B.; Medlock, J.M.; Snary, E.L. Assessing the risks of West Nile virus-infected mosquitoes from transatlantic aircraft: Implications for disease emergence in the United Kingdom. Vector-Borne Zoonotic Dis. 2012, 12, 310–320. [Google Scholar] [CrossRef]
- Jourdain, E.; Toussaint, Y.; Leblond, A.; Bicout, D.J.; Sabatier, P.; Gauthier-Clerc, M. Bird species potentially involved in introduction, amplification, and spread of West Nile virus in a Mediterranean wetland, the Camargue (Southern France). Vector-Borne Zoonotic Dis. 2007, 7, 15–33. [Google Scholar] [CrossRef]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Daszak, P.; Goodman, S.J.; Rogg, H.; Kramer, L.D.; Cedeno, V.; Cunningham, A.A. Predicting pathogen introduction: West Nile virus spread to Galaipagos. Conserv. Biol. 2006, 20, 1224–1231. [Google Scholar] [CrossRef]
- Douglas, K.O.; Kilpatrick, A.M.; Levett, P.N.; Lavoie, M.C. A quantitative risk assessment of West Nile virus introduction into Barbados. West Indian Med. J. 2007, 56, 394–397. [Google Scholar]
- Caminade, C.; Medlock, J.M.; Ducheyne, E.; McIntyre, K.M.; Leach, S.; Baylis, M.; Morse, A.P. Suitability of European climate for the Asian tiger mosquito Aedes albopictus: Recent trends and future scenarios. J. R. Soc. Interface/R. Soc. 2012, 9, 2708–2717. [Google Scholar] [CrossRef]
- Schaffner, F.; Medlock, J.M.; van Bortel, W. Public health significance of invasive mosquitoes in Europe. Clin. Microbiol. Infect. 2013, 19, 685–692. [Google Scholar] [CrossRef]
- Gjenero-Margan, I.; Aleraj, B.; Krajcar, D.; Lesnikar, V.; Klobucar, A.; Pem-Novosel, I.; Kurecic-Filipovic, S.; Komparak, S.; Martic, R.; Duricic, S.; et al. Autochthonous dengue fever in Croatia, August-September 2010. Euro. Surveill. 2011, 16, 19805:1–19805:4. [Google Scholar]
- La Ruche, G.; Souares, Y.; Armengaud, A.; Peloux-Petiot, F.; Delaunay, P.; Despres, P.; Lenglet, A.; Jourdain, F.; Leparc-Goffart, I.; Charlet, F.; et al. First two autochthonous dengue virus infections in metropolitan France, September 2010. Euro. Surveill. 2010, 15, 19676:1–19676:5. [Google Scholar]
- Ulbert, S. West Nile virus: The complex biology of an emerging pathogen. Intervirology 2011, 54, 171–184. [Google Scholar] [CrossRef]
- De Filette, M.; Ulbert, S.; Diamond, M.; Sanders, N.N. Recent progress in West Nile virus diagnosis and vaccination. Vet. Res. 2012, 43, 16. [Google Scholar] [CrossRef] [Green Version]
- Epp, T.; Waldner, C.; Townsend, H.G.G. A case-control study of factors associated with development of clinical disease due to West Nile virus, Saskatchewan 2003. Equine Vet. J. 2007, 39, 498–503. [Google Scholar]
- Minke, J.M.; Siger, L.; Cupillard, L.; Powers, B.; Bakonyi, T.; Boyum, S.; Nowotny, N.; Bowen, R. Protection provided by a recombinant ALVAC®-WNV vaccine expressing the prM/E genes of a lineage 1 strain of WNV against a virulent challenge with a lineage 2 strain. Vaccine 2011, 29, 4608–4612. [Google Scholar]
- Venter, M.; van Vuren, P.J.; Mentoor, J.; Paweska, J.; Williams, J. Inactivated West Nile Virus (WNV) vaccine, Duvaxyn WNV, protects against a highly neuroinvasive lineage 2 WNV strain in mice. Vaccine 2013, 31, 3856–3862. [Google Scholar] [CrossRef]
- Samina, I.; Khinich, Y.; Simanov, M.; Malkinson, M. An inactivated West Nile virus vaccine for domestic geese-efficacy study and a summary of 4 years of field application. Vaccine 2005, 23, 4955–4958. [Google Scholar] [CrossRef]
- Chang, G.-J.J.; Davis, B.S.; Stringfield, C.; Lutz, C. Prospective immunization of the endangered California condors (Gymnogyps californianus) protects this species from lethal West Nile virus infection. Vaccine 2007, 25, 2325–2330. [Google Scholar] [CrossRef]
- Boyce, W.M.; Vickers, W.; Morrison, S.A.; Sillett, T.S.; Caldwell, L.; Wheeler, S.S.; Barker, C.M.; Cummings, R.; Reisen, W.K. Surveillance for West Nile virus and vaccination of free-ranging island scrub-jays (Aphelocoma insularis) on Santa Cruz Island, California. Vector-Borne Zoonotic Dis. 2011, 11, 1063–1068. [Google Scholar] [CrossRef]
- Bunning, M.L.; Fox, P.E.; Bowen, R.A.; Komar, N.; Chang, G.-J.J.; Speaker, T.J.; Stephens, M.R.; Nemeth, N.; Panella, N.A.; Langevin, S.A.; et al. DNA vaccination of the American crow (Corvus brachyrhynchos) provides partial protection against lethal challenge with West Nile virus. Avian Dis. 2007, 51, 573–577. [Google Scholar] [CrossRef]
- Turell, M.J.; Bunning, M.; Ludwig, G.V.; Ortman, B.; Chang, J.; Speaker, T.; Spielman, A.; McLean, R.; Komar, N.; Gates, R.; et al. DNA vaccine for West Nile virus infection in fish crows (Corvus ossifragus). Emerg. Infect. Dis. 2003, 9, 1077–1081. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Dupuis, A.P.; Chang, G.-J.J.; Kramer, L.D. DNA vaccination of American robins (Turdus migratorius) against West Nile virus. Vector-Borne Zoonotic Dis. 2010, 10, 377–380. [Google Scholar]
- Barrett, A.D.T.; Teuwen, D.E. Yellow fever vaccine—How does it work and why do rare cases of serious adverse events take place? Curr. Opin. Immunol. 2009, 21, 308–313. [Google Scholar] [CrossRef]
- Tauber, E.; Kollaritsch, H.; von Sonnenburg, F.; Lademann, M.; Jilma, B.; Firbas, C.; Jelinek, T.; Beckett, C.; Knobloch, J.; McBride, W.J.H.; et al. Randomized, double-blind, placebo-controlled phase 3 trial of the safety and tolerability of IC51, an inactivated Japanese encephalitis vaccine. J. Infect. Dis. 2008, 198, 493–499. [Google Scholar] [CrossRef]
- Tauber, E.; Kollaritsch, H.; Korinek, M.; Rendi-Wagner, P.; Jilma, B.; Firbas, C.; Schranz, S.; Jong, E.; Klingler, A.; Dewasthaly, S.; et al. Safety and immunogenicity of a Vero-cell-derived, inactivated Japanese encephalitis vaccine: A non-inferiority, phase III, randomised controlled trial. Lancet 2007, 370, 1847–1853. [Google Scholar] [CrossRef]
- Heinz, F.X.; Holzmann, H.; Essl, A.; Kundi, M. Field effectiveness of vaccination against tick-borne encephalitis. Vaccine 2007, 25, 7559–7567. [Google Scholar] [CrossRef]
- Clarke, D.H.; Casals, J. Techniques for hemagglutination and hemagglutination-inhibition with arthropod-borne viruses. Am. J. Trop. Med. Hyg. 1958, 7, 561–573. [Google Scholar]
- Luca, V. The Structural Basis of Flaviviridae Interaction with Antibodies and Receptors; Washington University: Saint Louis, MO, USA, 2011. [Google Scholar]
- Sanchez, M.D.; Pierson, T.C.; McAllister, D.; Hanna, S.L.; Puffer, B.A.; Valentine, L.E.; Murtadha, M.M.; Hoxie, J.A.; Doms, R.W. Characterization of neutralizing antibodies to West Nile virus. Virology 2005, 336, 70–82. [Google Scholar] [CrossRef]
- Seligman, S.J.; Bucher, D.J. The importance of being outer: Consequences of the distinction between the outer and inner surfaces of flavivirus glycoprotein E. Trends Microbiol. 2003, 11, 108–110. [Google Scholar] [CrossRef]
- Pierson, T.C.; Diamond, M.S. Molecular mechanisms of antibody-mediated neutralisation of flavivirus infection. Expert Rev. Mol. Med. 2008, 10. [Google Scholar] [CrossRef]
- Bressanelli, S.; Stiasny, K.; Allison, S.L.; Stura, E.A.; Duquerroy, S.; Lescar, J.; Heinz, F.X.; Rey, F.A. Structure of a flavivirus envelope glycoprotein in its low-pH-induced membrane fusion conformation. EMBO J. 2004, 23, 728–738. [Google Scholar] [CrossRef]
- Diamond, M.S.; Pierson, T.C.; Fremont, D.H. The structural immunology of antibody protection against West Nile virus. Immunol. Rev. 2008, 225, 212–225. [Google Scholar] [CrossRef]
- Crill, W.D.; Chang, G.-J.J. Localization and characterization of flavivirus envelope glycoprotein cross-reactive epitopes. J. Virol. 2004, 78, 13975–13986. [Google Scholar] [CrossRef]
- Nybakken, G.E.; Oliphant, T.; Johnson, S.; Burke, S.; Diamond, M.S.; Fremont, D.H. Structural basis of West Nile virus neutralization by a therapeutic antibody. Nature 2005, 437, 764–769. [Google Scholar] [CrossRef]
- Beasley, D.W.C.; Barrett, A.D.T. Identification of neutralizing epitopes within structural domain III of the West Nile virus envelope protein. J. Virol. 2002, 76, 13097–13100. [Google Scholar] [CrossRef]
- Oliphant, T.; Nybakken, G.E.; Austin, S.K.; Xu, Q.; Bramson, J.; Loeb, M.; Throsby, M.; Fremont, D.H.; Pierson, T.C.; Diamond, M.S. Induction of epitope-specific neutralizing antibodies against West Nile virus. J. Virol. 2007, 81, 11828–11839. [Google Scholar] [CrossRef]
- Stiasny, K.; Kiermayr, S.; Holzmann, H.; Heinz, F.X. Cryptic properties of a cluster of dominant flavivirus cross-reactive antigenic sites. J. Virol. 2006, 80, 9557–9568. [Google Scholar] [CrossRef]
- Vogt, M.R.; Dowd, K.A.; Engle, M.; Tesh, R.B.; Johnson, S.; Pierson, T.C.; Diamond, M.S. Poorly neutralizing cross-reactive antibodies against the fusion loop of West Nile virus envelope protein protect in vivo via Fcgamma receptor and complement-dependent effector mechanisms. J. Virol. 2011, 85, 11567–11580. [Google Scholar]
- Lobigs, M.; Diamond, M.S. Feasibility of cross-protective vaccination against flaviviruses of the Japanese encephalitis serocomplex. Expert Rev. Vaccines 2012, 11, 177–187. [Google Scholar] [CrossRef]
- Post, P.R.; Carvalho, R.; Galler, R. Glycosylation and secretion of yellow fever virus nonstructural protein NS1. Virus Res. 1991, 18, 291–302. [Google Scholar] [CrossRef]
- Flamand, M.; Megret, F.; Mathieu, M.; Lepault, J.; Rey, F.A.; Deubel, V. Dengue virus type 1 nonstructural glycoprotein NS1 is secreted from mammalian cells as a soluble hexamer in a glycosylation-dependent fashion. J. Virol. 1999, 73, 6104–6110. [Google Scholar]
- Chung, K.M.; Thompson, B.S.; Fremont, D.H.; Diamond, M.S. Antibody recognition of cell surface-associated NS1 triggers Fc-γ receptor-mediated phagocytosis and clearance of West Nile Virus-infected cells. J. Virol. 2007, 81, 9551–9555. [Google Scholar] [CrossRef]
- Hirota, J.; Shimizu, S.; Shibahara, T. Application of West Nile virus diagnostic techniques. Expert Rev. Anti-Infect. Ther. 2013, 11, 793–803. [Google Scholar] [CrossRef]
- Lindsey, H.S.; Calisher, C.H.; Mathews, J.H. Serum dilution neutralization test for California group virus identification and serology. J. Clin. Microbiol. 1976, 4, 503–510. [Google Scholar]
- OIE. West Nile Fever. In OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 6th ed.; Office International des Epizooties: Paris, France, 2008; pp. 377–385. [Google Scholar]
- Weingartl, H.M.; Drebot, M.A.; Hubalek, Z.; Halouzka, J.; Andonova, M.; Dibernardo, A.; Cottam-Birt, C.; Larence, J.; Marszal, P. Comparison of assays for the detection of West Nile virus antibodies in chicken serum. Can. J. Vet. Res.-Rev. Can. Rech. Veterinaire 2003, 67, 128–132. [Google Scholar]
- Buckley, A; Dawson, A; Moss, S.R.; Hlinsley, S.A.; Bellamy, P.E.; Gould, E.A. Serological evidence of West Nile virus, Usutu virus and Sindbis virus infection of birds in the UK. J. Gen. Virol. 2003, 84, 2807–2817. [Google Scholar] [CrossRef]
- Hubalek, Z.; Wegner, E.; Halouzka, J.; Tryjanowski, P.; Jerzak, L.; Sikutova, S.; Rudolf, I.; Kruszewicz, A.G.; Jaworski, Z.; Wlodarczyk, R. Serologic survey of potential vertebrate hosts for West Nile virus in Poland. Viral. Immunol. 2008, 21, 247–254. [Google Scholar]
- Hobson-Peters, J. Approaches for the development of rapid serological assays for surveillance and diagnosis of infections caused by zoonotic flaviviruses of the Japanese encephalitis virus serocomplex. J Biomed. Biotechnol. 2012, 2012. [Google Scholar] [CrossRef]
- Niedrig, M.; Sonnenberg, K.; Steinhagen, K.; Paweska, J.T. Comparison of ELISA and immunoassays for measurement of IgG and IgM antibody to West Nile virus in human sera against virus neutralisation. J. Virol. Methods 2007, 139, 103–105. [Google Scholar] [CrossRef]
- Martin, D.A.; Muth, D.A.; Brown, T.; Johnson, A.J.; Karabatsos, N.; Roehrig, J.T. Standardization of immunoglobulin M capture enzyme-linked immunosorbent assays for routine diagnosis of arboviral infections. J. Clin. Microbiol. 2000, 38, 1823–1826. [Google Scholar]
- Tardei, G.; Ruta, S.; Chitu, V.; Rossi, C.; Tsai, T.F.; Cernescu, C. Evaluation of immunoglobulin M (IgM) and IgG enzyme immunoassays in serologic diagnosis of West Nile virus infection. J. Clin. Microbiol. 2000, 38, 2232–2239. [Google Scholar]
- Niedrig, M.; Donoso-Mantke, O.; Altmann, D.; Zeller, H. First international diagnostic accuracy study for the serological detection of West Nile virus infection. BMC Infect. Dis. 2007, 7. [Google Scholar] [CrossRef]
- Alonso-Padilla, J.; Jimenez de Oya, N.; Blazquez, A.-B.; Loza-Rubio, E.; Escribano, J.M.; Saiz, J.-C.; Escribano-Romero, E. Evaluation of an enzyme-linked immunosorbent assay for detection of West Nile virus infection based on a recombinant envelope protein produced in Trichoplusia ni larvae. J. Virol. Methods 2010, 166, 37–41. [Google Scholar] [CrossRef]
- Sotelo, E.; Llorente, F.; Rebollo, B.; Camunas, A.; Venteo, A.; Gallardo, C.; Lubisi, A.; Rodriguez, M.J.; Sanz, A.J.; Figuerola, J.; et al. Development and evaluation of a new epitope-blocking ELISA for universal detection of antibodies to West Nile virus. J. Virol. Methods 2011, 174, 35–41. [Google Scholar] [CrossRef]
- Holbrook, M.R.; Shope, R.E.; Barrett, A.D.T. Use of recombinant E protein domain III-based enzyme-linked immunosorbent assays for differentiation of tick-borne encephalitis serocomplex flaviviruses from mosquito-borne flaviviruses. J. Clin. Microbiol. 2004, 42, 4101–4110. [Google Scholar] [CrossRef]
- Beasley, D.W.C.; Holbrook, M.R.; Travassos da Rosa, A.P.A.; Coffey, L.; Carrara, A.-S.; Phillippi-Falkenstein, K.; Bohm, R.P., Jr.; Ratterree, M.S.; Lillibridge, K.M.; Ludwig, G.V.; et al. Use of a recombinant envelope protein subunit antigen for specific serological diagnosis of West Nile virus infection. J. Clin. Microbiol. 2004, 42, 2759–2765. [Google Scholar] [CrossRef]
- Kitai, Y.; Shoda, M.; Kondo, T.; Konishi, E. Epitope-blocking enzyme-linked immunosorbent assay to differentiate west nile virus from Japanese encephalitis virus infections in equine sera. Clin. Vaccine Immunol. 2007, 14, 1024–1031. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Marlenee, N.L.; Hall, R.A.; Calisher, C.H.; Bowen, R.A.; Roehrig, J.T.; Komar, N.; Langevin, S.A.; Beaty, B.J. Epitope-blocking enzyme-linked immunosorbent assays for the detection of serum antibodies to west nile virus in multiple avian species. J. Clin. Microbiol. 2003, 41, 1041–1047. [Google Scholar] [CrossRef]
- Houser, B. Bio-Rad’s Bio-Plex® suspension array system, xMAP technology overview. Arch. Physiol. Biochem. 2012, 118, 192–196. [Google Scholar] [CrossRef]
- Lelli, D.; Moreno, A.; Brocchi, E.; Sozzi, E.; Capucci, L.; Canelli, E.; Barbieri, I.; Zeller, H.; Cordioli, P. West Nile virus: Characterization and diagnostic applications of monoclonal antibodies. Virol. J. 2012, 9. [Google Scholar] [CrossRef]
- Nash, D.; Mostashari, F.; Fine, A.; Miller, J.; O’Leary, D.; Murray, K.; Huang, A.; Rosenberg, A.; Greenberg, A.; Sherman, M.; et al. The outbreak of West Nile virus infection in the New York city area in 1999. N. Engl. J. Med. 2001, 344, 1807–1814. [Google Scholar] [CrossRef]
- Shieh, W.J.; Guarner, J.; Layton, M.; Fine, A.; Miller, J.; Nash, D.; Campbell, G.L.; Roehrig, J.T.; Gubler, D.J.; Zaki, S.R. The role of pathology in an investigation of an outbreak of West Nile encephalitis in New York, 1999. Emerg. Infect. Dis. 2000, 6, 370–372. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Roehrig, J.T.; Deubel, V.; Smith, J.; Parker, M.; Steele, K.; Crise, B.; Volpe, K.E.; Crabtree, M.B.; Scherret, J.H.; et al. Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science 1999, 286, 2333–2337. [Google Scholar] [CrossRef]
- Ledermann, J.P.; Lorono-Pino, M.A.; Ellis, C.; Saxton-Shaw, K.D.; Blitvich, B.J.; Beaty, B.J.; Bowen, R.A.; Powers, A.M. Evaluation of widely used diagnostic tests to detect West Nile virus infections in horses previously infected with St. Louis encephalitis virus or dengue virus type 2. Clin. Vaccine Immunol. 2011, 18, 580–587. [Google Scholar] [CrossRef]
- Lorono-Pino, M.A.; Farfan-Ale, J.A.; Blitvich, B.J.; Beebe, J.L.; Jarman, R.G.; Beaty, B.J. Evaluation of an epitope-blocking enzyme-linked immunosorbent assay for the diagnosis of West Nile virus infections in humans. Clin. Vaccine Immunol. 2009, 16, 749–755. [Google Scholar] [CrossRef]
- Papa, A.; Karabaxoglou, D.; Kansouzidou, A. Acute West Nile virus neuroinvasive infections: cross-reactivity with dengue virus and tick-borne encephalitis virus. J. Med. Virol. 2011, 83, 1861–1865. [Google Scholar] [CrossRef]
- Shirafuji, H.; Kanehira, K.; Kamio, T.; Kubo, M.; Shibahara, T.; Konishi, M.; Murakami, K.; Nakamura, Y.; Yamanaka, T.; Kondo, T.; et al. Antibody responses induced by experimental West Nile Virus infection with or without previous immunization with inactivated Japanese encephalitis vaccine in horses. J. Vet. Med. Sci. 2009, 71, 969–974. [Google Scholar] [CrossRef]
- Hirota, J.; Nishi, H.; Matsuda, H.; Tsunemitsu, H.; Shimizu, S. Cross-reactivity of Japanese encephalitis virus-vaccinated horse sera in serodiagnosis of West Nile virus. J. Vet. Med. Sci. 2010, 72, 369–372. [Google Scholar] [CrossRef]
- Yeh, J.-Y.; Lee, J.-H.; Park, J.-Y.; Seo, H.-J.; Moon, J.-S.; Cho, I.-S.; Kim, H.-P.; Yang, Y.-J.; Ahn, K.-M.; Kyung, S.-G.; et al. A diagnostic algorithm to serologically differentiate West Nile Virus from Japanese Encephalitis Virus infections and its validation in field surveillance of poultry and horses. Vector-Borne Zoonotic Dis. 2012, 12, 372–379. [Google Scholar] [CrossRef]
- Williams, D.T.; Daniels, P.W.; Lunt, R.A.; Wang, L.-F.; Newberry, K.M.; MacKenzie, J.S. Experimental infections of pigs with Japanese encephalitis virus and closely related Australian flaviviruses. Am. J. Trop. Med. Hyg. 2001, 65, 379–387. [Google Scholar]
- Buckley, A.; Dawson, A.; Gould, E.A. Detection of seroconversion to West Nile virus, Usutu virus and Sindbis virus in UK sentinel chickens. Virol. J. 2006, 3. [Google Scholar] [CrossRef] [Green Version]
- Rabel, P.O.; Planitzer, C.B.; Farcet, M.R.; Orlinger, K.K.; Ilk, R.; Barrett, P.N.; Kreill, T.R. Increasing West Nile virus antibody titres in central European plasma donors from 2006 to 2010. Euro. Surveill. 2011, 16, 19812:1–19812:5. [Google Scholar]
- Sonnleitner, S.T.; Simeoni, J.; Baumgartner, R.; Zelger, R.; Prader, A.; Piccolini, G.; Nowotny, N.; Walder, G. The spreading of flaviviruses over the continental divide: A challenge for serologic diagnostics. J. Med. Microbiol. Diagn. 2013, 3. [Google Scholar] [CrossRef]
- Jourdain, E.; Schuffenecker, I.; Korimbocus, J.; Reynard, S.; Murri, S.; Kayser, Y.; Gauthier-Clerc, M.; Sabatier, P.; Zeller, H.G. West Nile virus in wild resident birds, Southern France, 2004. Vector-Borne Zoonotic Dis. 2007, 7, 448–452. [Google Scholar] [CrossRef]
- Alba, A.; Allepuz, A.; Napp, S.; Soler, M.; Selga, I.; Aranda, C.; Casal, J.; Pages, N.; Hayes, E.B.; Busquets, N. Ecological Surveillance for West Nile in Catalonia (Spain), Learning from a Five-Year Period of Follow-up. Zoonoses Public Health 2013, 2013. [Google Scholar] [CrossRef]
- Lopez, G.; Jimenez-Clavero, M.A.; Tejedor, C.G.; Soriguer, R.; Figuerola, J. Prevalence of West Nile virus neutralizing antibodies in Spain is related to the behavior of migratory birds. Vector-Borne Zoonotic Dis. 2008, 8, 615–621. [Google Scholar] [CrossRef] [Green Version]
- Figuerola, J.; Soriguer, R.; Rojo, G.; Tejedor, C.G.; Jimenez-Clavero, M.A. Seroconversion in wild birds and local circulation of west nile virus, Spain. Emerg. Infect. Dis. 2007, 13, 1915–1917. [Google Scholar] [CrossRef]
- Jourdain, E.; Olsen, B.; Lundkvist, A.; Hubálek, Z.; Šikutová, S.; Waldenström, J.; Karlsson, M.; Wahlström, M.; Jozan, M.; Falk, K.I. Surveillance for West Nile virus in wild birds from Northern Europe. Vector-Borne Zoonotic Dis. 2011, 11, 77–79. [Google Scholar] [CrossRef]
- Valiakos, G.; Touloudi, A.; Athanasiou, L.V.; Giannakopoulos, A.; Iacovakis, C.; Birtsas, P.; Spyrou, V.; Dalabiras, Z.; Petrovska, L.; Billinis, C. Serological and molecular investigation into the role of wild birds in the epidemiology of West Nile virus in Greece. Virol. J. 2012, 9. [Google Scholar] [CrossRef]
- Boadella, M.; Diez-Delgado, I.; Gutierrez-Guzman, A.V.; Hofle, U.; Gortazar, C. Do wild ungulates allow improved monitoring of flavivirus circulation in Spain? Vector-Borne Zoonotic Dis. 2012, 12, 490–495. [Google Scholar] [CrossRef]
- Turner, J.L.; Waggoner, J.W.; Rose, S.S.; Arns, M.J.; Hankins, K.G.; Tuttle, J. West Nile Virus antibody titers and total immunoglobulin G concentrations in Foals from Mares Vaccinated in Late Gestation. J. Equine Vet. Sci. 2008, 28, 17–21. [Google Scholar] [CrossRef]
- Nemeth, N.M.; Oesterle, P.T.; Bowen, R.A. Passive immunity to West Nile virus provides limited protection in a common passerine species. Am. J. Trop. Med. Hyg. 2008, 79, 283–290. [Google Scholar]
- Nemeth, N.M.; Bowen, R.A. Dynamics of passive immunity to West Nile virus in domestic chickens (Gallus gallus domesticus). Am. J. Trop. Med. Hyg. 2007, 76, 310–317. [Google Scholar]
- Ahmadnejad, F.; Otarod, V.; Fallah, M.H.; Lowenski, S.; Sedighi-Moghaddam, R.; Zavareh, A.; Durand, B.; Lecollinet, S.; Sabatier, P. Spread of West Nile virus in Iran: A cross-sectional serosurvey in equines, 2008–2009. Epidemiol. Infect. 2011, 139, 1587–1593. [Google Scholar] [CrossRef]
- Fang, Y.; Reisen, W.K. Previous infection with West Nile or St. Louis encephalitis viruses provides cross protection during reinfection in house finches. Am. J. Trop. Med. Hyg. 2006, 75, 480–485. [Google Scholar]
- Nemeth, N.M.; Bosco-Lauth, A.M.; Bowen, R.A. Cross-protection between West Nile and Japanese encephalitis viruses in red-winged blackbirds (Agelaius phoeniceus). Avian Dis. 2009, 53, 421–425. [Google Scholar] [CrossRef]
- Goverdhan, M.K.; Kulkarni, A.B.; Gupta, A.K.; Tupe, C.D.; Rodrigues, J.J. Two-way cross-protection between West Nile and Japanese encephalitis viruses in bonnet macaques. Acta. Virol. 1992, 36, 277–283. [Google Scholar]
- Tesh, R.B.; da Rosa, A.; Guzman, H.; Araujo, T.P.; Xiao, S.Y. Immunization with heterologous flaviviruses protective against fatal West Nile encephalitis. Emerg. Infect. Dis. 2002, 8, 245–251. [Google Scholar] [CrossRef]
- Price, W.H.; Thind, I.S. The mechanism of cross-protection afforded by dengue virus against West Nile virus in hamsters. J. Hyg. 1972, 70, 611–617. [Google Scholar] [CrossRef]
- Tarr, G.C.; Hammon, W.M. Cross-protection between group B arboviruses: Resistance in mice to Japanese B encephalitis and St. Louis encephalitis viruses induced by dengue virus immunization. Infect. Immun. 1974, 9, 909–915. [Google Scholar]
- Yamshchikov, G.; Borisevich, V.; Kwok, C.W.; Nistler, R.; Kohlmeier, J.; Seregin, A.; Chaporgina, E.; Benedict, S.; Yamshchikov, V. The suitability of yellow fever and Japanese encephalitis vaccines for immunization against West Nile virus. Vaccine 2005, 23, 4785–4792. [Google Scholar] [CrossRef]
- Petrovsky, N.; Larena, M.; Siddharthan, V.; Prow, N.A.; Hall, R.A.; Lobigs, M.; Morrey, J. An inactivated cell-culture Japanese encephalitis vaccine (JE-ADVAX) formulated with delta inulin adjuvant provides robust heterologous protection against West Nile encephalitis via cross-protective memory B cells and neutralizing antibody. J. Virol. 2013, 87, 10324–10333. [Google Scholar] [CrossRef]
- Lobigs, M.; Larena, M.; Alsharifi, M.; Lee, E.; Pavy, M. Live chimeric and inactivated Japanese encephalitis virus vaccines differ in their cross-protective values against Murray Valley encephalitis virus. J. Virol. 2009, 83, 2436–2445. [Google Scholar] [CrossRef]
- Orlinger, K.K.; Hofmeister, Y.; Fritz, R.; Holzer, G.W.; Falkner, F.G.; Unger, B.; Loew-Baselli, A.; Poellabauer, E.-M.; Ehrlich, H.J.; Barrett, P.N.; Kreil, T.R. A tick-borne encephalitis virus vaccine based on the European prototype strain induces broadly reactive cross-neutralizing antibodies in humans. J. Infect. Dis. 2011, 203, 1556–1564. [Google Scholar] [CrossRef]
- Arroyo, J.; Miller, C.; Catalan, J.; Myers, G.A.; Ratterree, M.S.; Trent, D.W.; Monath, T.P. ChimeriVax-West Nile virus live-attenuated vaccine: Preclinical evaluation of safety, immunogenicity, and efficacy. J. Virol. 2004, 78, 12497–12507. [Google Scholar] [CrossRef]
- Porterfield, J.S. Immunological enhancement and the pathogenesis of dengue haemorrhagic fever. J. Hyg. 1982, 89, 355–364. [Google Scholar] [CrossRef]
- Halstead, S.B. Immune enhancement of viral infection. Prog. Allergy 1982, 31, 301–364. [Google Scholar]
- Guzman, M.G.; Vazquez, S. The complexity of antibody-dependent enhancement of dengue virus infection. Viruses 2010, 2, 2649–2662. [Google Scholar] [CrossRef]
- Phillpotts, R.J.; Stephenson, J.R.; Porterfield, J.S. Antibody-dependent enhancement of tick-borne encephalitis virus infectivity. J. Gen. Virol. 1985, 66, 1831–1837. [Google Scholar] [CrossRef]
- Ferenczi, E.; Ban, E.; Abraham, A.; Kaposi, T.; Petranyi, G.; Berencsi, G.; Vaheri, A. Severe tick-borne encephalitis in a patient previously infected by West Nile virus. Scand. J. Infect. Dis. 2008, 40, 759–761. [Google Scholar] [CrossRef]
- Vittecoq, M.; Thomas, F.; Jourdain, E.; Moutou, F.; Renaud, F.; Gauthier-Clerc, M. Risks of emerging infectious diseases: Evolving threats in a changing area, the mediterranean basin. Transbound. Emerg. Dis. 2012, 2012. [Google Scholar] [CrossRef]
- Cameron, A.R. The consequences of risk-based surveillance: Developing output-based standards for surveillance to demonstrate freedom from disease. Prev. Vet. Med. 2012, 105, 280–286. [Google Scholar] [CrossRef]
- Reist, M.; Jemmi, T.; Stark, K.D.C. Policy-driven development of cost-effective, risk-based surveillance strategies. Prev. Vet. Med. 2012, 105, 176–184. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Beck, C.; Jimenez-Clavero, M.A.; Leblond, A.; Durand, B.; Nowotny, N.; Leparc-Goffart, I.; Zientara, S.; Jourdain, E.; Lecollinet, S. Flaviviruses in Europe: Complex Circulation Patterns and Their Consequences for the Diagnosis and Control of West Nile Disease. Int. J. Environ. Res. Public Health 2013, 10, 6049-6083. https://doi.org/10.3390/ijerph10116049
Beck C, Jimenez-Clavero MA, Leblond A, Durand B, Nowotny N, Leparc-Goffart I, Zientara S, Jourdain E, Lecollinet S. Flaviviruses in Europe: Complex Circulation Patterns and Their Consequences for the Diagnosis and Control of West Nile Disease. International Journal of Environmental Research and Public Health. 2013; 10(11):6049-6083. https://doi.org/10.3390/ijerph10116049
Chicago/Turabian StyleBeck, Cécile, Miguel Angel Jimenez-Clavero, Agnès Leblond, Benoît Durand, Norbert Nowotny, Isabelle Leparc-Goffart, Stéphan Zientara, Elsa Jourdain, and Sylvie Lecollinet. 2013. "Flaviviruses in Europe: Complex Circulation Patterns and Their Consequences for the Diagnosis and Control of West Nile Disease" International Journal of Environmental Research and Public Health 10, no. 11: 6049-6083. https://doi.org/10.3390/ijerph10116049