Marine Drugs from Sponge-Microbe Association—A Review

Abstract
:1. Introduction
2. Sponges and Associated Microbes Involved in Drug Production
2.1. Class: Demospongiae
2.1.1. Order: Astrophorida
2.1.2. Order: Chondrosida
2.1.3. Order: Dendroceratida
2.1.4. Order: Dictyoceratida
2.1.4.1. Family: Dysideidae
2.1.4.2. Family: Irciniidae
2.1.4.3. Family: Spongiidae
2.1.4.4. Family: Thorectidae
2.1.5. Order: Hadromerida
2.1.5.1. Family: Spirastrellidae
2.1.5.2. Family: Suberitidae
2.1.6. Order: Halichondrida
2.1.6.1. Family: Axinellidae
2.1.6.2. Family: Halichondriidae
2.1.7. Order: Haplosclerida
2.1.7.1. Family: Callyspongiidae
2.1.7.2. Family: Chalinidae
2.1.7.3. Family: Niphatidae
2.1.7.4. Family: Petrosiidae
2.1.8. Order: Lithistida
2.1.8.1. Family: Neopeltidae
2.1.8.2. Family: Theonellidae
2.1.9. Order: Poecilosclerida
2.1.9.1. Family: Acarnidae
2.1.9.2. Family: Isodictyidae
2.1.9.3. Family: Raspailiidae
2.1.9.4. Family: Mycalidae
2.1.9.5. Family: Myxillidae
2.1.10. Order: Spirophorida
2.1.10.1. Family: Tetillidae
2.1.11. Order: Verongida
2.1.11.1. Family: Aplysinellidae
2.1.11.2. Family: Aplysinidae
2.1.11.3. Family: Pseudoceratinidae
2.2. Class: Calcarea
2.3. Unidentified sponges
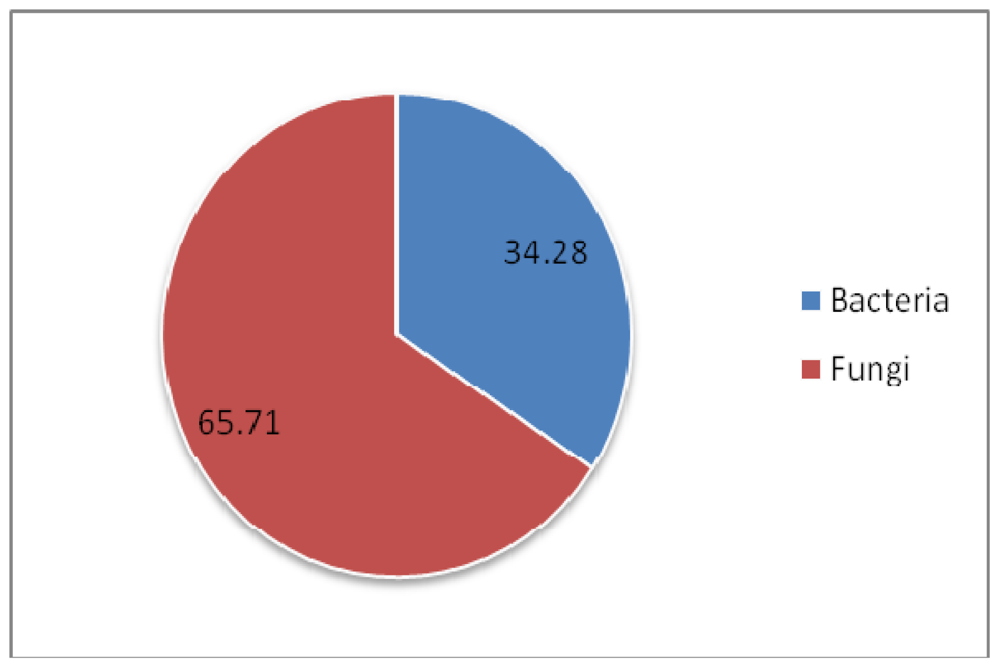
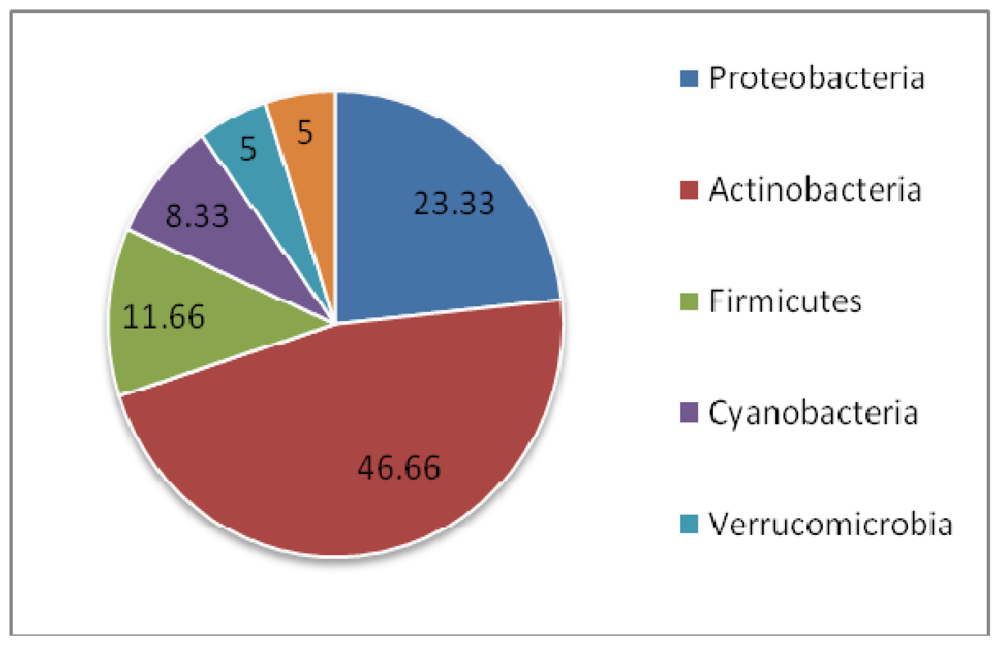
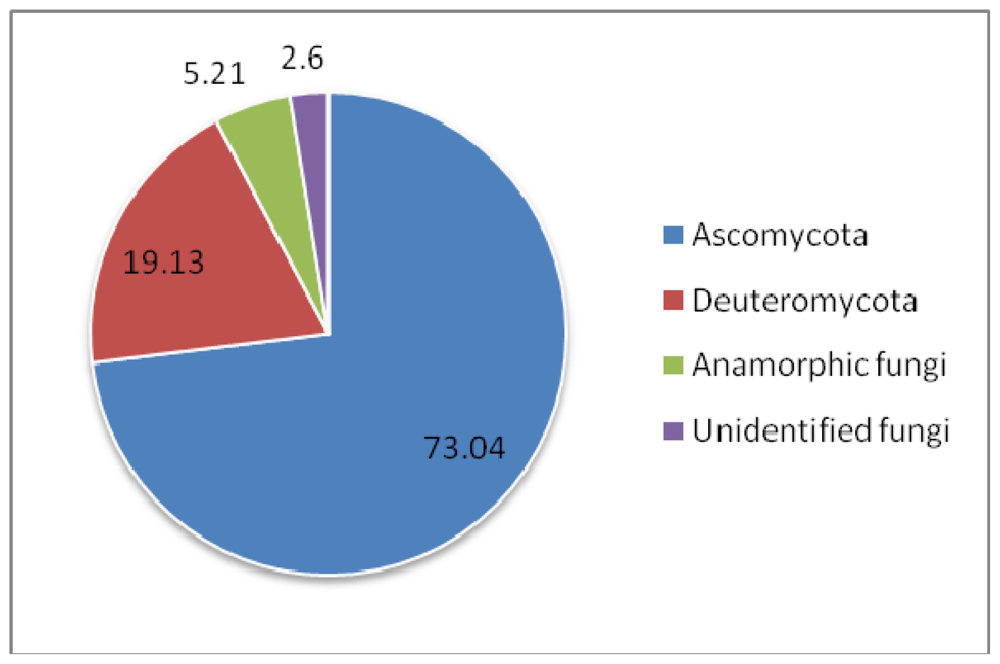
3. Discussion
4. Ecological and Cultural Aspects of Sponge Symbionts
5. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Hentschel, U; Hopke, J; Horn, M; Friedrich, AB; Wagner, M; Hacker, J; Moore, BS. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl Environ Microbiol 2002, 68, 4431–4440. [Google Scholar]
- Radjasa, OK; Sabdono, A; Junaidi; Zocchi, E. Richness of secondary metabolite producing marine bacteria associated with sponge Haliclona sp. Int J Pharm 2007, 3, 275–279. [Google Scholar]
- Fusetani, N; Matsunaga, S. Bioactive sponge peptides. Chem Rev 1993, 93, 1793–1806. [Google Scholar]
- Lee, YK; Lee, JH; Lee, HK. Microbial symbiosis in marine sponges. J Microbiol 2001, 39, 254–264. [Google Scholar]
- Fieseler, L; Horn, M; Wagner, M; Hentschel, U. Discovery of the novel candidate Phylum “Poribacteria” in marine sponges. Appl Environ Microbiol 2004, 70, 3724–3732. [Google Scholar]
- Belarbi, EH; Gomez, AC; Chisti, Y; Camacho, FG; Grima, EM. Producing drugs from marine sponges. Biotechnol Adv 2003, 21, 585–598. [Google Scholar]
- Thakur, NL; Müller, WEG. Biotechnological potential of marine sponges. Curr Sci 2004, 86, 1506–1512. [Google Scholar]
- Jensen, PR; Fenical, W. Strategies for the discovery of secondary metabolites from marine bacteria: ecological perspectives. Annu Rev Microbiol 1994, 48, 559–584. [Google Scholar]
- Bernan, VS; Greenstein, M; Maise, WM. Marine microorganisms as a source of new natural products. Adv Appl Microbiol 1997, 43, 57–89. [Google Scholar]
- Haygood, MG; Schmidt, EW; Davidson, SK; Faulkner, DJ. Microbial symbionts of marine invertebrates: opportunities for microbial biotechnology. J Molec Microbiol Biotechnol 1999, 1, 33–43. [Google Scholar]
- Osinga, R; Armstrong, E; Burgess, JG; Hoffmann, F; Reitner, J; Schumann-Kindel, G. Sponge microbe associations and their importance for sponge bioprocess engineering. Hydrobiologia 2001, 461, 55–62. [Google Scholar]
- Proksch, P; Edrada, RA; Ebel, R. Drugs from the seas: current status and microbiological imblications. Appl Microbiol Biotechnol 2002, 59, 125–134. [Google Scholar]
- Taylor, MW; Radax, R; Steger, D; Wagner, M. Sponge associated microorganisms: Evolution, ecology and biotechnological potential. Microbiol Mol Biol Rev 2007, 71, 295–347. [Google Scholar]
- Wang, G. Diversity and biotechnological potential of the sponge-associated microbial consortia. J Ind Microbiol Biotechnol 2006, 33, 545–551. [Google Scholar]
- Thakur, AN; Thakur, NL; Indap, MM; Pandit, RA; Datar, VV; Müller, WEG. Antiangiogenic, antimicrobial and cytotoxic potential of sponge-associated bacteria. Mar Biotechnol 2005, 7, 245–252. [Google Scholar]
- Gunasekera, AS; Sfanos, KS; Harmody, DK; Pomponi, SA; McCarthy, PJ; Lopez, JV. An enhanced database of the microorganisms associated with deeper water marine invertebrates. Appl Microbiol Biotechnol 2004, 66, 373–376. [Google Scholar]
- Kobayashi, J; Ishibashi, M. Bioactive metabolites of symbiotic marine microorganism. Chem Rev 1993, 93, 1753–1769. [Google Scholar]
- Ridley, CP; Faulkner, DJ; Haygood, MG. Investigation of Oscillatoria spongeliae dominated bacterial communities in four dictyoceratid sponges. Appl Environ Microbiol 2005, 71, 7366–7375. [Google Scholar]
- Alvarez, B; Crisp, MD; Driver, F; Hooper, JNA; Soest, RWMV. Phylogenetic relationships of the family Axinellidae (Porifera: Demospongiae) using morphological and molecular data. Zool Scripta 2000, 29, 169–198. [Google Scholar]
- Bergmann, W; Burke, DC. Contributions to the study of marine products. XXXIX. The nucleosides of sponges. III. Spongothymidine and spongouridine. J Org Chem 1955, 20, 1501–1507. [Google Scholar]
- Wang, G. Diversity and biotechnological potential of the sponge-associated microbial consortia. J Ind Microbiol Biotechnol 2006, 33, 545–551. [Google Scholar]
- Kennedy, J; Baker, P; Piper, C; Cotter, PD; Walsh, M; Mooij, MJ; Bourke, MB; Rea, MC; O’Connor, PM; Ross, RP; Hill, C; O’Gara, F; Marchesi, JR; Dobson, ADW. Isolation and analysis of bacteria with antimicrobial activities from the marine sponge Haliclona simulans collected from Irish waters. Mar Biotechnol 2009, 11, 384–396. [Google Scholar]
- Li, Z; He, L; Miao, X. Cultivable bacterial community from South China Sea sponge as revealed by DGGE fingerprinting and 16S rDNA phylogenetic analysis. Curr Microbiol 2007, 55, 465–472. [Google Scholar]
- Head, IM; Saunders, JR; Pickup, RW. Microbial Evolution, Diversity, and Ecology: A Decade of Ribosomal RNA Analysis of Uncultivated Microorganisms. Microb Ecol 1998, 35, 1–21. [Google Scholar]
- Juretschko, S; Timmermann, G; Schmid, M; Schleifer, K; Pommerening-Röser, A; Koops, H; Wagner, M. Combined Molecular and Conventional Analyses of Nitrifying Bacterium Diversity in Activated Sludge: Nitrosococcus mobilis and Nitrospira-Like Bacteria as Dominant Populations. Appl Environ Microbiol 1998, 64, 3042–3051. [Google Scholar]
- Schmidt, EW; Obraztsova, AY; Davidson, SK; Faulkner, DJ; Haygood, MG. Identification of the antifungal peptide-containing symbiont of the marine sponge Theonella swinhoei as a novel δ-Proteobacterium Candidatus Entotheonella palauensis. Mar Biol 2000, 136, 969–977. [Google Scholar]
- Vacelet, J; Gallissian, M. Virus-like particles in cells of the sponge Virongia cavernicola (demospongiae, dictyoceratida) and accompanying tissue changes. J Invertebr Pathol 1978, 31, 246–254. [Google Scholar]
- Olson, JB; Mccarthy, PJ. Associated bacterial communities of two deep-water sponges. Aquat Microb Ecol 2005, 39, 47–55. [Google Scholar]
- Hill, M; Hill, A; Lopez, N; Harriott, O. Sponge-specific bacterial symbionts in the Caribbean sponge, Chondrilla nucula (Demospongiae, Chondrosida). Mar Biol 2006, 148, 1221–1230. [Google Scholar]
- Kasai, H; Katsuta, A; Sekiguchi, H; Matsuda, S; Adachi, K; Shindo, K; Yoon, J; Yokota, A; Shizuri, Y. Rubritalea squalenifaciens sp. nov., a squalene-producing marine bacterium belonging to subdivision 1 of the phylum ‘Verrucomicrobia’. Int J Syst Evol Microbiol 2007, 57, 1630–1634. [Google Scholar]
- Enticknap, JJ; Kelly, M; Peraud, O; Hill, RT. Characterization of a culturable alphaproteobacterial symbiont common to many marine sponges and evidence for vertical transmission via sponge larvae. Appl Environ Microbiol 2006, 72, 3724–3732. [Google Scholar]
- Thiel, V; Imhoff, JF. Phylogenetic identification of bacteria with antimicrobial activities isolated from Mediterranean sponges. Biomol Eng 2003, 20, 421–423. [Google Scholar]
- Radjasa, OK; Martens, T; Grossart, H; Brinkhoff, T; Sabdono, A; Simmon, M. Antagonistic activity of a marine bacterium Pseudoalteromonas luteoviolacea TAB4.2 associated with coral Acropora sp. J Biol Sci 2007, 7, 239–246. [Google Scholar]
- Zhang, L; An, R; Wang, J; Sun, N; Zhang, S; Hu, J; Kuai, J. Exploring novel bioactive compounds from marine microbes. Curr Opin Microbiol 2005, 8, 276–281. [Google Scholar]
- Newman, DJ; Hill, RT. New drugs from marine microbes: the tide is turning. J Ind Microbiol Biotechnol 2006, 33, 539–544. [Google Scholar]
- Perry, NB; Blunt, JW; Munro, MHG. Mycalamide A, an antiviral compound from a New Zealand sponge of the genus Mycale. J Am Chem Soc 1988, 110, 4850–4851. [Google Scholar]
- Hood, KA; West, LM; Northcote, PT; Berridge, MV; Miller, JH. Induction of apoptosis by the marine sponge (Mycale) metabolites, mycalamide A and pateamine. Apoptosis 2001, 6, 207–219. [Google Scholar]
- Laport, MS; Santos, OCS; Muricy, G. Marine sponges: Potential sources of new antimicrobial drugs. Curr Pharmaceut Biotechnol 2009, 10, 86–105. [Google Scholar]
- Unson, MD; Holland, ND; Faulkner, DJ. A brominated secondary metabolite synthesized by the cyanobacterial symbiont of a marine sponge and accumulation of the crystalline metabolite in the sponge tissue. Mar Biol 1994, 119, 1–11. [Google Scholar]
- Hentschel, U; Schmid, M; Wagner, M; Fieseler, L; Gernert, C; Hacker, J. Isolation and phylogenetic analyses of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicula. FEMS Microbiol Ecol 2001, 35, 305–312. [Google Scholar]
- Zheng, Z; Zeng, W; Huang, Y; Yang, Z; Li, J; Cai, H; Su, W. Detection of antitumor and antimicrobial activities in marine organism associated actinomycetes isolated from the Taiwan Strait, China. FEMS Microbiol Lett 2000, 188, 87–91. [Google Scholar]
- Li, Z. Advances in marine microbial symbionts in the China Sea and related pharmaceutical metabolites. Mar Drugs 2009, 7, 113–129. [Google Scholar]
- Zhang, Y; Mu, J; Feng, Y; Kang, Y; Zhang, J; Gu, P; Wang, Y; Ma, L; Zhu, Y. Broad-spectrum antimicrobial epiphytic and endophytic fungi from marine organisms: Isolation, bioassay and taxonomy. Mar Drugs 2009, 7, 97–112. [Google Scholar]
- Biabani, MAF; Laatsch, H. Advances in chemical studies on low-molecular weight metabolites of marine fungi. J Prakt Chem 1998, 340, 589–607. [Google Scholar]
- Cheng, X; Varoglu, M; Abrell, L; Crews, P; Lobkovsky, E; Clardy, J. Chloriolins A-C, chlorinated sesquiterpenes produced by fungal cultures separated from a Jaspis marine sponge. J Org Chem 1994, 59, 6344–6348. [Google Scholar]
- Sufrin, JR; Finckbeiner, S; Oliver, CM. Marine-Derived Metabolites of S-Adenosylmethionine as Templates for New Anti-Infectives. Mar Drugs 2009, 7, 401–434. [Google Scholar]
- Lang, G; Wiese, J; Schmaljohann, R; Imhoff, JF. New pentaenes from the sponge-derived marine fungus Penicillium rugulosum: structure determination and biosynthetic studies. Tetrahedron 2007, 63, 11844–11849. [Google Scholar]
- Blunt, JW; Copp, BR; Hu, W; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat Prod Rep 2009, 26, 170–244. [Google Scholar]
- Selvin, J; Gandhimathi, R; Seghal Kiran, G; Shanmugha Priya, S; Ravji, TR; Hema, TA. Culturable heterotrophic bacteria from the marine sponge Dendrilla nigra: Isolation and phylogenetic diversity of actinobacteria. Helgol Mar Res 2009, 63, 239–247. [Google Scholar]
- Selvin, J. Exploring the antagonistic producer Streptomyces MSI051: Implications of polyketide synthase gene type II and a ubiquitous defense enzyme phospholipase A2 in the host sponge Dendrilla nigra. Curr Microbiol 2009, 58, 459–463. [Google Scholar]
- Selvin, J; Lipton, AP. Biopotentials of secondary metabolites isolated from marine sponges. Hydrobiologia 2004, 513, 231–238. [Google Scholar]
- Selvin, J; Joseph, S; Asha, KRT; Manjusha, WA; Sangeetha, VS; Jayaseema, DM; Antony, MC; Vinitha, AJD. Antibacterial potential of antagonistic Streptomyces sp. isolated from marine sponge Dendrilla nigra. FEMS Microbiol Ecol 2004, 50, 117–122. [Google Scholar]
- Selvin, J; Shanmughapriya, S; Gandhimathi, R; Kiran, GS; Ravji, TR; Natarajaseenivasan, K; Hema, TA. Optimization and production of novel antimicrobial agents from sponge associated marine actinomycetes Nocardiopsis dassonvillei MAD08. Appl Microbiol Biotechnol 2009, 83, 435–445. [Google Scholar]
- Hinde, R; Pironet, F; Borowitzka, MA. Isolation of Oscillatoria spongeliae, the filamentous cyanobacterial symbiont of the marine sponge Dysidea herbacea. Mar Biol 1994, 119, 99–104. [Google Scholar]
- Arillo, A; Bavestrello, G; Burlando, B; Sara, M. Metabolic integration between symbiotic cyanobacteria and sponges: a possible mechanism. Mar Biol 1993, 117, 159–162. [Google Scholar]
- Unson, MD; Faulkner, DJ. Cyanobacterial symbiont biosynthesis of chlorinated metabolites from Dysidea herbacea (Porifera). Cell Mol Life Sci 1993, 49, 349–353. [Google Scholar]
- Flowers, AE; Garson, MJ; Webb, RI; Dumdei, EJ; Charan, RD. Cellular origin of chlorinated diketopiperazines in the dictyoceratid sponge Dysidea herbacea (Keller). Cell Tissue Res 1998, 292, 597–607. [Google Scholar]
- Besada, P; Mamedova, L; Thomas, CJ; Costanzi, S; Jacobson, KA. Design and synthesis of new bicyclic diketopiperazines as scaffolds for receptor probes of structurally diverse functionality. Org Biomol Chem 2005, 3, 2016–2025. [Google Scholar]
- Elyakov, GB; Kuznetsova, T; Mikhailov, VV; Maltsev, II; Voinov, VG; Fedoreyev, SA. Brominated diphenyl ethers from a marine bacterium associated with the sponge Dysidea sp. Cell Mol Life Sci 47, 632–633.
- Scheuermayer, M; Pimentel-Elardo, S; Fieseler, L; Grozdanov, L; Hentschel, U. Proksch, P, Müller, WEG, Eds.; Microorganisms of sponges: Phylogenetic diversity and biotechnological potential. In Frontiers in Marine Biotechnology; Horizon Bioscience: Norfolk, UK, 2006; pp. 289–312. [Google Scholar]
- Thakur, NL; Müller, WEG. Sponge-bacteria association: A useful model to explore symbiosis in marine invertebrates. Symbiosis 2005, 39, 109–116. [Google Scholar]
- Müller, WEG; Thakur, NL; Ushijima, H; Thakur, AN; Krasko, A; Pennec, G; Indap, MM; Perovic-Ottstadt, S; Schröder, HC; Lang, G; Bringmann, G. Matrix-mediated canal formation in primmorphs from the sponge Suberites domuncula involves the expression of a CD36 receptor-ligand system. J Cell Sci 2004, 117, 2579–2590. [Google Scholar]
- Mohamed, NM; Rao, V; Hamann, MT; Kelly, M; Hill, RT. Monitoring bacterial diversity of the marine sponge Ircinia strobilina upon transfer to aquaculture. Appl Environ Microbiol 2008, 74, 4133–4143. [Google Scholar]
- Bringmann, G; Lang, G; Muhlbacher, J; Schaumann, K; Steffens, S; Rytik, PG; Hentschel, U; Morschhauser, J; Müller, WEG. Sorbicillactone A: A structurally unprecedented bioactive novel-type alkaloid from a sponge-derived fungus. Prog Mol Subcell Biol 2003, 37, 231–253. [Google Scholar]
- Bringmann, G; Gulder, TAM; Lang, G; Schmitt, S; Stöhr, R; Wiese, J; Nagel, K; Imhoff, JF. Large–scale biotechnological production of the antileukemic marine natural product sorbicillactone A. Mar Drugs 2007, 5, 23–30. [Google Scholar]
- Oclarit, JM; Okada, H; Ohta, S; Kaminura, K; Yamaoka, Y; Iizuka, T; Miyashiro, S; Ikegami, S. Anti-bacillus substance in the marine sponge, Hyatella species, produced by an associated Vibrio species bacterium. Microbios 1994, 78, 7–16. [Google Scholar]
- Fredenhagen, A; Tamura, SY; Kenny, PTM; Komura, H; Naya, Y; Nakanishi, K; Nishiyama, K; Sugiura, M; Kita, H. Andrimid, a new peptide antibiotic produced by an intracellular bacterial symbiont isolated from a brown planthopper. J Am Chem Soc 1987, 109, 4409–4411. [Google Scholar]
- Needham, J; Kelly, MT; Ishige, M; Andersen, RJ. Andrimid and moiramides A-C, metabolites produced in culture by a marine isolate of the bacterium Pseudomonas fluorescens: Structure elucidation and biosynthesis. J Org Chem 1994, 59, 2058–2063. [Google Scholar]
- Amagata, T; Rath, C; Rigot, JF; Tarlov, N; Tenney, K; Valeriote, FA; Crews, P. Structures and cytotoxic properties of trichoverroids and their macrolide analogues produced by saltwater culture of Myrothecium verrucaria. J Med Chem 2003, 46, 4342–4350. [Google Scholar]
- Kobayashi, M; Kitagawa, I. Bioactive substances isolated from marine sponge, a miniature conglomerate of various organisms. Pure Appl Chem 1994, 66, 819–826. [Google Scholar]
- Kobayashi, M; Aoki, S; Gato, K; Matsunami, K; Kurosu, M; Kitagawa, I. Marine natural products. XXXIV. Trisindoline, a new antibiotic indole trimer, produced by a bacterium of Vibrio sp. separated from the marine sponge Hyrtios altum. Chem Pharm Bull 1994, 42, 2449–2451. [Google Scholar]
- Braekman, J; Daloze, D. Chemical and biological aspects of sponge secondary metabolites. Phytochem Rev 2004, 3, 275–283. [Google Scholar]
- Varoglu, M; Corbett, TH; Valeriote, FA; Crews, P. Asperazine, a selective cytotoxic alkaloid from a sponge-derived culture of Aspergillus niger. J Org Chem 1997, 62, 7078–7079. [Google Scholar]
- Govek, SP; Overman, LE. Total synthesis of asperazine. J Am Chem Soc 2001, 123, 9468–9469. [Google Scholar]
- Varoglu, M; Crews, P. Biosynthetically diverse compounds from a saltwater culture of sponge derived Aspergillus niger. J Nat Prod 2000, 63, 41–43. [Google Scholar]
- Speitling, M; Smetanina, OF; Kuznetsova, TA; Laatsch, H. Bromoalterochromides A and A′, unprecedented chromopeptides from a marine Pseudoalteromonas maricaloris strain KMM 636T. J Antibiot 2007, 60, 36–42. [Google Scholar]
- Abrell, LM; Borgeson, B; Crews, P. A new polyketide, secocurvularin, from the salt water culture of a sponge derived fungus. Tetrahedon Lett 1996, 37, 8983–8984. [Google Scholar]
- Bugni, TS; Ireland, CM. Marine derived fungi: a chemically and biologically diverse group of microorganisms. Nat Prod Rep 2004, 21, 143–163. [Google Scholar]
- Thakur, NL; Hentschel, U; Krasko, A; Pabel, CT; ANR, AC; Müller, WEG. Antibacterial activity of the sponge Suberites domuncula and its primmorphs: potential basis for epibacterial chemical defense. Aquat Microb Ecol 2003, 31, 77–83. [Google Scholar]
- Webster, NS; Hill, RT. The culturable microbial community of the Great Barrier Reef sponge Rhopaloeides odorabile is dominated by an α-Proteobacterium. Mar Biol 2001, 138, 843–851. [Google Scholar]
- Williams, DE; Burgoyne, DL; Rettig, SJ; Andersen, RJ; Fathi-Afshar, ZR; Allen, TM. The isolation of Majusculamide C from the sponge Ptilocaulis trachys collected in Enewetak and determination of the absolute configuration of the 2-methyl-3-aminopentanoic acid residue. J Nat Prod 1993, 56, 545–551. [Google Scholar]
- Dunlap, WC; Battershill, CN; Liptrot, CH; Cobb, RE; Bourne, DG; Jaspars, M; Long, PF; Newman, DJ. Biomedicinals from the phytosymbionts of marine invertebrates: A molecular approach. Methods 2007, 42, 358–376. [Google Scholar]
- Xie, LW; Jiang, SM; Zhu, HHl; Sun, W; Ouyang, YC; Dai, SK; Li, X. Potential inhibitors against Sclerotinia sclerotiorum, produced by the fungus Myrothecium sp. associated with the marine sponge Axinella sp. Eur J Plant Pathol 2008, 122, 571–578. [Google Scholar]
- Amagata, T; Amagata, A; Tenney, K; Valeriote, FA; Lobkovsky, E; Clardy, J; Crews, P. Unusual C25 steroids produced by a sponge-derived Penicillium citrinum. Org Lett 2003, 5, 4393–4396. [Google Scholar]
- Koizumi, Y; Arai, M; Tomoda, H; Omura, S. Oxaline, a fungal alkaloid, arrests the cell cycle in M phase by inhibition of tubulin polymerization. Biochim Biophys Acta 2004, 1693, 47–55. [Google Scholar]
- Kolachana, P; Smith, MT. Induction of kinetochore-positive micronuclei in human lymphocytes by the anti-fungal drug griseofulvin. Mutat Res 1994, 322, 151–159. [Google Scholar]
- Jadulco, R; Edrada, RA; Ebel, R; Berg, A; Schaumann, K; Wray, V; Steube, K; Proksch, P. New communesin derivatives from the fungus Penicillium sp. derived from the Mediterranean sponge Axinella verrucosa. J Nat Prod 2004, 67, 78–81. [Google Scholar]
- Hiort, J; Maksimenka, K; Reichert, M; Perovic-Ottstadt, S; Lin, WH; Wray, V; Steube, K; Schaumann, K; Weber, H; Proksch, P; Ebel, R; Müller, WEG; Bringmann, G. New natural products from the sponge-derived fungus Aspergillus niger. J Nat Prod 2004, 67, 1532–1543. [Google Scholar]
- Boot, CM; Tenney, K; Valeriote, FA; Crews, P. Highly N-methylated linear peptides produced by an atypical sponge-derived Acremonium sp. J Nat Prod 2006, 69, 83–92. [Google Scholar]
- Boot, CM; Amagata, T; Tenney, K; Compton, JE; Pietraszkiewicz, H; Valeriote, FA; Crews, P. Four classes of structurally unusual peptides from two marine-derived fungi: structures and bioactivities. Tetrahedon 2007, 63, 9903–9914. [Google Scholar]
- Erpenbeck, D; Breeuwer, JAJ; van der Velde, HC; van Soest, RWM. Unravelling host and symbiont phylogenies of halichondrid sponges (Demospongiae, Porifera) using mitochondrial marker. Mar Biol 2002, 141, 377–386. [Google Scholar]
- Miki, W; Otaki, N; Yokoyama, A; Izumida, H; Shimidzu, N. Okadaxanthin, a novel C50-carotenoid from a bacterium Pseudomonas sp. KK10206C associated with a marine sponge Halichondria okadai. Experientia 1994, 50, 684–686. [Google Scholar]
- Kelecom, A. Secondary metabolites from marine microorganisms. An Acad Bras Cienc 2002, 74, 151–170. [Google Scholar]
- Shigemori, H; Bae, MA; Yazawa, K; Sasaki, T; Kobayashi, J. Alteramide A, a new tetracyclic alkaloid from a bacterium Alteromonas sp. associated with the marine sponge Halichondria okadai. J Org Chem 1992, 57, 4317–4320. [Google Scholar]
- Bhalla, TC; Sharma, M; Sharma, NN. Satyanarayana, T, Chand, S, Eds.; Microbial production of flavours and fragrances; fats and oils; dyes; bioplastics (PHAS); polysaccharides; pharmacologically active substances from marine microbes; anticancer agents and microbial transformation. In Applied Microbiology; National Science Digital Library NISCAIR: New Delhi, India, 2008; Volume 7, pp. 1–34. [Google Scholar]
- Amagata, T; Usami, Y; Minoura, K; Ito, T; Numata, A. Cytotoxic substances produced by a fungal strain from a sponge: physico-chemical properties and structures. J Antibiot 1998, 51, 33–40. [Google Scholar]
- Usami, Y; Ikura, T; Amagata, T; Numata, A. First total syntheses and configurational assignments of cytotoxic trichodenones A–C. Tetrahedron-Asymmetry 2000, 11, 3711–3725. [Google Scholar]
- Shindo, K; Asagi, E; Sano, A; Hotta, E; Minemura, N; Mikami, K; Tamesada, E; Misawa, N; Maoka, T. Diapolycopenedioic Acid Xylosyl Esters A, B, and C, Novel Antioxidative Glyco-C30-carotenoic Acids Produced by a New Marine Bacterium Rubritalea squalenifaciens. J Antibiot 2008, 61, 185–191. [Google Scholar]
- Althoff, K; Schutt, C; Steffen, R; Batel, R; Müller, WEG. Evidence for a symbiosis between bacteria of the genus Rhodobacter and the marine sponge Halichondria panicea: Harbor also for putatively toxic bacteria? Mar Biol 1998, 130, 529–536. [Google Scholar]
- Perovic, S; Wichels, A; Schutt, C; Gerdts, G; Pahler, S; Steffen, R; Müller, WEG. Neuroactive compounds produced by bacteria from the marine sponge Halichondria panicea: activation of the neuronal NMDA receptor. Environ Toxicol Pharmacol 1998, 6, 125–133. [Google Scholar]
- Wicke, C; Hners, M; Wray, V; Nimtz, M; Bilitewski, U; Lang, S. Production and structure elucidation of glycoglycerolipids from a marine sponge associated Microbacterium species. J Nat Prod 2000, 63, 621–626. [Google Scholar]
- Numata, A; Amagata, T; Minoura, K; lto, T. Gymnastatins, novel cytotoxic metabolites produced by a fungal strain from a sponge. Tetrahedon Lett 1997, 38, 5675–5678. [Google Scholar]
- Amagata, T; Doi, M; Ohta, T; Minoura, K; Numata, A. Absolute stereostructures of novel cytotoxic metabolites, gymnastatins A–E, from a Gymnascella species separated from a Halichondria sponge. J Chem Soc Perkin Trans 1 1998, 1, 3585–3599. [Google Scholar]
- Amagata, T; Doi, M; Tohgo, M; Minoura, K; Numata, A. Dankasterone, a new class of cytotoxic steroid produced by a Gymnascella species from a marine sponge. Chem Commun 1999, 1321–1322. [Google Scholar]
- Mayer, AMS. Marine Pharmacology in 1998: Antitumor and Cytotoxic Compounds. Pharmacologist 1999, 41, 159–164. [Google Scholar]
- Amagata, T; Minoura, K; Numata, A. 2006. Gymnastatins F-H, Cytostatic Metabolites from the Sponge-Derived Fungus Gymnascella dankaliensis. J Nat Prod 2006, 69, 1384–1388. [Google Scholar]
- Amagata, T; Tanaka, M; Yamada, T; Minoura, K; Numata, A. Gymnastatins and Dankastatins, Growth Inhibitory Metabolites of a Gymnascella Species from a Halichondria Sponge. J Nat Prod 2008, 71, 340–345. [Google Scholar]
- Amagata, T; Tanaka, M; Yamada, T; Doi, M; Minoura, K; Ohishi, H; Yamori, T; Numata, A. Variation in Cytostatic Constituents of a Sponge-Derived Gymnascella dankaliensis by Manipulating the Carbon Source. J Nat Prod 2007, 70, 1731–1740. [Google Scholar]
- Amagata, T; Minoura, K; Numata, A. Gymnasterones, novel cytotoxic metabolites produced by a fungal strain from a sponge. Tetrahedron Lett 1998, 39, 3773–3774. [Google Scholar]
- Nagai, K; Kamigiri, K; Matsumoto, H; Kawano, Y; Yamaoka, M; Shimoi, H; Watanabe, M; Suzuki, K. YM-202204, a new antifungal antibiotic produced by marine fungus Phoma sp. J Antibiot 2002, 55, 1036–1041. [Google Scholar]
- Nagai, K; Kamigiri, K; Arao, N; Suzumura, K; Kawano, Y; Yamaoka, M; Zhang, H; Watanabe, M; Suzuki, K. YM-266183 and YM-266184, novel thiopeptide antibiotics produced by Bacillus cereus isolated from a marine sponge. I. Taxonomy, fermentation, isolation, physico-chemical properties and biological properties. J Antibiot 2003, 56, 123–128. [Google Scholar]
- Suzumura, K; Yokoi, T; Funatsu, M; Nagai, K; Tanaka, K; Zhang, H; Suzuki, K. YM-266183 and YM-266184, novel thiopeptide antibiotics produced by Bacillus cereus isolated from a marine sponge II. Structure elucidation. J Antibiot 2003, 56, 129–134. [Google Scholar]
- Laatsch, H. Proksch, P, Müller, WEG, Eds.; Marine bacterial metabolite. In Frontiers in Marine Biotechnology; Horizon Bioscience: Norfolk, UK, 2006; pp. 225–288. [Google Scholar]
- Hildebrand, M; Waggoner, LE; Lim, GE; Sharp, KH; Ridley, CP; Haygood, MG. Approaches to identify, clone, and express symbiont bioactive metabolite genes. Nat Prod Rep 2003, 21, 122–142. [Google Scholar]
- El Sayed, KA; Dunbar, DC; Perry, TL; Wilkins, SP; Hamann, MT. Marine natural products as prototype insecticidal agents. J Agric Food Chem 1997, 45, 2735–2739. [Google Scholar]
- Ramm, W; Schatton, W; Wagner-Dobler, I; Wray, V; Nimtz, M; Tokuda, H; Enjyo, F; Nishino, H; Beil, W; Heckmann, R; Lurtz, V; Lang, S. Diglucosyl-glycerolipids from the marine sponge-associated Bacillus pumilus strain AAS3: their production, enzymatic modification and properties. Appl Microbiol Biotechnol 2004, 64, 497–504. [Google Scholar]
- Zheng, L; Chen, H; Han, X; Line, W; Yan, X. Antimicrobial screening and active compound isolation from marine bacterium NJ6-3-1 associated with the sponge Hymeniacidon perleve. World J Microbiol Biotechnol 2005, 21, 201–206. [Google Scholar]
- Jadulco, R; Proksch, P; Wray, V; Sudarsono; Berg, A; Grafe, U. New macrolides and furan carboxylic acid derivative from the sponge derived fungus Cladosporium herbarum. J Nat Prod 2001, 64, 527–530. [Google Scholar]
- Höller, U; König, GM; Wright, AD. A new tyrosine kinase inhibitor from a marine isolate of Ulocladium botrytis and new metabolites from the marine fungi Asteromyces cruciatus and Varicosporina ramulosa. Eur J Org Chem 1999, 1999, 2949–2955. [Google Scholar]
- König, GM; Kehraus, S; Seibert, SF; Abdel-Lateff, A; Müller, D. Natural products from marine organisms and their associated microbes. Chem Bio Chem 2005, 7, 229–238. [Google Scholar]
- Yu, S; Deng, Z; Proksch, P; Lin, W. Oculatol, oculatolide and A-nor sterols from the sponge Haliclona oculata. J Nat Prod 2006, 69, 1330–1334. [Google Scholar]
- Bringmann, G; Lang, G; Steffens, S; Gunther, E; Schaumann, K. Evariquinone, isoemericellin, and stromemycin from a sponge derived strain of the fungus Emericella variecolor. Phytochemistry 2003, 63, 437–443. [Google Scholar]
- Wang, G; Abrell, LM; Avelar, A; Borgeson, BM; Crews, P. New hirsutane based sesquiterpenes from salt water cultures of a marine sponge derived fungus and the terrestrial fungus Coriolus consors. Tetrahedron 1998, 54, 7335–7342. [Google Scholar]
- Bhadury, P; Mohammad, BT; Wright, PC. The current status of natural products from marine fungi and their potential as anti-infective agents. J Ind Microbiol Biotechnol 2006, 33, 325–337. [Google Scholar]
- Jadulco, R; Brauers, G; Edrada, RU; Ebel, R; Wray, V; Sudarsono; Proksch, P. New metabolites from sponge derived fungi Curvularia lunata and Cladosporium herbarum. J Nat Prod 2002, 65, 730–733. [Google Scholar]
- Kim, JS; Im, KS; Jung, JH. New bioactive polyacetylenes from the marine sponge Petrosia sp. Tetrahedron 1998, 54, 3151–3158. [Google Scholar]
- Lim, YJ; Park, HS; Im, KS; Lee, CO; Hong, J; Lee, M; Kim, D; Jung, JH. Additional cytotoxic polyacetylenes from the marine sponge Petrosia sp. J Nat Prod 2001, 64, 46–53. [Google Scholar]
- Vacelet, J; Donadey, C. Electron microscope study of the association between some sponges and bacteria. J Exp Mar Bio Ecol 1977, 30, 301–314. [Google Scholar]
- Chelossi, E; Milanese, M; Milano, A; Pronzato, R; Riccardi, G. Characterisation and antimicrobial activity of epibiotic bacteria from Petrosia ficiformis (Porifera, Demospongiae). J Exp Mar Biol Ecol 2004, 309, 21–33. [Google Scholar]
- Bringmann, G; Lang, G; Steffens, S; Schaumann, K. Petrosifungins A and B, novel cyclodepsipeptides from a sponge-derived strain of Penicillium brevicompactum. J Nat Prod 2004, 67, 311–315. [Google Scholar]
- Lemmens-Gruber, R; Kamyar, MR; Dornetshuber, R. Cyclodepsipeptides - Potential drugs and lead compounds in the drug development process. Curr Med Chem 2009, 16, 1122–1137. [Google Scholar]
- López-Gresa, MP; Cabedo, N; González-Mas, MC; Ciavatta, MA; Avila, C; Primo, J. Terretonins E and F, Inhibitors of the Mitochondrial Respiratory Chain from the Marine-Derived Fungus Aspergillus insuetus. J Nat Prod 2009, 72, 1348–1351. [Google Scholar]
- Bultel-Poncé, V; Debitus, C; Berge, J; Cerceau, C; Guyot, M. Metabolites from the sponge-associated bacterium Micrococcus luteus. J Mar Biotechnol 1998, 6, 233–236. [Google Scholar]
- Edrada, RA; Heubes, M; Brauers, G; Wray, V; Berg, A; Grafe, U; Wohlfarth, M; Muhlbacher, J; Schaumann, K; Sudarsono, S; Bringmann, G; Proksch, P. Online analysis of xestodecalactones A-C, novel bioactive metabolites from the fungus Penicillium cf. montanense and their subsequent isolation from the sponge Xestospongia exigua. J Nat Prod 2002, 65, 1598–1604. [Google Scholar]
- Lin, W; Brauers, G; Ebel, R; Wray, V; Berg, A; Sudarsono; Proksch, P. Novel chromone derivatives from the fungus Aspergillus versicolor isolated from the marine sponge Xestospongia exigua. J Nat Prod 2002, 66, 57–61. [Google Scholar]
- El Sayed, KA; Kelly, M; Kara, UAK; Ang, KKH; Katsuyama, T; Dumbar, DC; Khan, AA; Hamann, MT. New manzamine alkaloids with potent activity against infectious diseases. J Am Chem Soc 2001, 123, 1804–1808. [Google Scholar]
- Sakai, R; Higa, T; Jefford, CW; Bernardinelli, G. Manzamine A, a novel antitumor alkaloid from a sponge. J Am Chem Soc 1986, 108, 6404–6405. [Google Scholar]
- Ang, KKH; Holmes, MJ; Higa, T; Hamann, MT; Kara, UAK. In vivo antimalarial activity of the beta-carboline alkaloid manzamine A. Antimicrob Agents Chemother 2000, 44, 1645–1649. [Google Scholar]
- Lee, YM; Mansoor, TA; Hong, J; Lee, C-O; Bae, KS; Jung, JH. Polyketides from a Sponge-Derived Fungus, Aspergillus versicolor. Nat Prod Sci 2007, 13, 90–96. [Google Scholar]
- Lee, YM; Dang, HT; Hong, J; Lee, C-O; Bae, KS; Kim, DK; Jung, JH. A Cytotoxic Lipopeptide from the Sponge-Derived Fungus Aspergillus versicolor. Bull Korean Chem Soc 2010, 31, 205–208. [Google Scholar]
- Fujii, Y; Asahara, M; Ichinoe, M; Nakajima, H. Fungal melanin inhibitor and related compounds from Penicillium decumbens. Phytochemistry 2002, 60, 703–708. [Google Scholar]
- Bultel-Poncé, V; Berge, J; Debitus, C; Nicolas, J; Guyot, M. Metabolites from the sponge associated bacterium Pseudomonas species. Mar Biotechnol 1999, 1, 384–390. [Google Scholar]
- Capon, RJ; Ford, J; Lacey, E; Gill, JH; Heiland, K; Friedel, T. Phoriospongin A and B: Two new nematocidal depsipeptides from the Australian marine sponges Phoriospongia sp. and Callyspongia bilamellata. J Nat Prod 2002, 65, 358–363. [Google Scholar]
- Bewley, CA; Faulkner, DJ. Lithistid sponges: Star performers or hosts to the stars. Angew Chem Int Ed 1998, 37, 2162–2178. [Google Scholar]
- Bewley, CA; Holland, ND; Faulkner, DJ. Two classes of metabolites from Theonella swinhoei are localized in distinct populations of bacterial symbionts. Experientia 1996, 52, 716–722. [Google Scholar]
- Schmidt, EW; Bewley, CA; Faulkner, DJ. Theopalauamide, a bicyclic glycopeptide from filamentous bacterial symbionts of the lithistid sponge Theonella swinhoei from Palau and Mozambique. J Org Chem 1998, 63, 1254–1258. [Google Scholar]
- Bewley, CA; Faulkner, DJ. Theonegramide, an antifungal glycopeptide from the Philippine lithistid sponge Theonella swinhoei. J Org Chem 1994, 59, 4849–4852. [Google Scholar]
- Piel, J. Metabolites from symbiotic bacteria. Nat Prod Rep 2004, 21, 519–538. [Google Scholar]
- Piel, J; Hui, D; Wen, G; Butzke, D; Platzer, M; Fusetani, N; Matsunaga, S. Antitumor polyketide biosynthesis by an uncultivated bacterial symbiont of the marine sponge Theonella swinhoei. Proc Natl Acad Sci 2004, 101, 16222–16227. [Google Scholar]
- Grozdanov, L; Hentschel, U. An environmental genomics perspective on the diversity and function of marine sponge-associated microbiota. Curr Opinion Microbiol 2007, 10, 215–220. [Google Scholar]
- Bugni, TS; Bernan, VS; Greenstein, M; Janso, JE; Maiese, WM; Mayne, CL; Ireland, CM. Brocaenols A–C: Novel polyketides from a marine-derived Penicillium brocae. J Org Chem 2003, 68, 2014–2017. [Google Scholar]
- Ebel, R. Proksch, P, Müller, WEG, Eds.; Secondary metabolites from marine-derived fungi. In Frontiers in Marine Biotechnology; Horizon Scientific Press: Norwich, UK, 2006; pp. 73–143. [Google Scholar]
- Jayatilake, GS; Thornton, MP; Leonard, AC; Grimwade, JE; Baker, BJ. Metabolites from an Antarctic sponge associated bacterium Pseudomonas aeruginosa. J Nat Prod 1996, 59, 293–296. [Google Scholar]
- Höller, U; König, GM; Wright, AD. Three new metabolites from marine derived fungi of the genera Coniothyrium and Microsphaeropsis. J Nat Prod 1999, 62, 114–118. [Google Scholar]
- Mohamed, IE; Gross, H; Pontius, A; Kehraus, S; Krick, A; Kelter, G; Maier, A; Fiebig, H; König, GM. Epoxyphomalin A and B, Prenylated Polyketides with Potent Cytotoxicity from the Marine-Derived Fungus Phoma sp. Org Lett 2009, 11, 5014–5017. [Google Scholar]
- Kralj, A; Gurgui, M; König, GM; van Echten-Deckert, G. Trichothecenes induce accumulation of glucosylceramide in neural cells by interfering with lactosylceramide synthase activity. Toxicol Appl Pharmacol 2007, 225, 113–122. [Google Scholar]
- Chatterjee, S; Kolmakova, A. Lactosylceramide synthase: From molecular biochemistry to biological function. In Lipids (sphingolipid metabolizing enzymes 2004); Research Signpost: Trivandrum, India, 2004; pp. 33–41. [Google Scholar]
- Kralj, A; Kehraus, S; Krick, A; van Echten-Deckert, G; König, GM. Two new depsipeptides from the marine fungus Spicellum roseum. Planta Med 2007, 73, 366–371. [Google Scholar]
- Liu, R; Cui, C; Duan, L; Gu, Q; Zhu, W. Potent in Vitro anticancer activity of metacycloprodigiosin and undecylprodigiosin from a sponge-derived actinomycete Saccharopolyspora sp nov. Arch Pharm Res 2005, 28, 1341–1344. [Google Scholar]
- Xin, ZH; Zhu, WM; Gu, QQ; Fang, LD; Cui, CB. A new cytotoxic compound from Penicillium auratiogriseum, symbiotic or epiphytic fungus of sponge Mycale plumose. Chin Chem Lett 2005, 16, 1227–1229. [Google Scholar]
- Xin, ZH; Fang, Y; Du, L; Zhu, T; Duan, L; Chen, J; Gu, Q; Zhu, W. Aurantiomides A C, quinazoline alkaloids from the sponge-derived fungus Penicillium aurantiogriseum SP0-19. J Nat Prod 2007, 70, 853–855. [Google Scholar]
- Doshida, J; Hasegawa, H; Onuki, H; Shimidzu, N. Exophilin A, a new antibiotic from a marine microorganism Exophiala pisciphila. J Antibiot 1996, 49, 1105–1109. [Google Scholar]
- Han, Y; Yang, B; Zhang, F; Miao, X; Li, Z. Characterization of antifungal chitinase from marine Streptomyces sp. DA11 associated with south China sea sponge Craniella australiensis. Mar Biotechnol 2009, 11, 132–140. [Google Scholar]
- Kim, TK; Garson, MJ; Fuerst, JA. Marine actinomycetes related to the ‘Salinospora’ group from the Great Barrier Reef sponge Pseudoceratina clavata. Environ Microbiol 2005, 7, 509–518. [Google Scholar]
- Mincer, TJ; Jensen, PR; Kauffman, CA; Fenical, W. Widespread and persistent populations of a major new marine Actinomycete taxon in ocean sediments. Appl Environ Microbiol 2002, 68, 5005–5011. [Google Scholar]
- Kim, TK; Hewavitharana, AK; Shaw, PN; Fuerst, JA. Discovery of a new source of rifamycin antibiotics in marine sponge Actinobacteria by phylogenetic prediction. Appl Environ Microbiol 2006, 72, 2118–2125. [Google Scholar]
- Lafi, FF; Garson, MJ; Fuerst, JA. Culturable bacterial symbionts isolated from two distinct sponge species (Pseudoceratina clavata and Rhabdastrella globostellata) from the Great Barrier Reef of Australia. Microb Ecol 2005, 50, 213–220. [Google Scholar]
- Friedrich, AB; Merkert, H; Fendert, T; Hacker, J; Proksch, P; Hentschel, U. Microbial diversity in the marine sponge Aplysina cavernicola (formerly Verongia cavernicola) analyzed by fluorescence in situ hybridization (FISH). Mar Biol 1999, 134, 461–470. [Google Scholar]
- Friedrich, AB; Fischer, I; Proksch, P; Hacker, J; Hentschel, U. Temporal variation of the microbial community associated with the Mediterranean sponge Aplysina aerophoba. FEMS Microbiol Ecol 2001, 38, 105–113. [Google Scholar]
- Pabel, CT; Vater, J; Wilde, C; Franke, P; Hofemeister, J; Adler, B; Bringmann, G; Hacker, J; Hentschel, U. Antimicrobial activities and Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry of Bacillus Isolates from the marine sponge Aplysina aerophoba. Mar Biotechnol 2003, 5, 424–434. [Google Scholar]
- Maget-Dana, R; Peypoux, F. Iturins, a special class of pore-forming lipopeptides: biological and physicochemical properties. Toxicology 1994, 87, 151–174. [Google Scholar]
- Besson, F; Peypoux, F; Michel, G; Delcambe, L. Mode of action of iturin A, an antibiotic isolated from Bacillus subtilis on Micrococcus luteus. Biochem Biophys Res Commun 1978, 81, 297–304. [Google Scholar]
- Klich, MA; Lax, AR; Bland, JM. Inhibition of some mycotoxigenic fungi by iturin A, a peptidolipid produced by Bacillus subtilis. Mycopathologia 2004, 116, 77–80. [Google Scholar]
- Brauers, G; Edrada, RA; Ebel, R; Proksch, P; Wray, V; Berg, A; Grafe, U; Schachtele, C; Totzke, F; Finkenzeller, G; Marme, D; Kraus, J; Munchbach, M; Michel, M; Bringmann, G; Schaumann, K. Anthraquinones and betaenone derivatives from the sponge-associated fungus Microsphaeropsis species: Novel inhibitors of protein kinases. J Nat Prod 2000, 63, 739–745. [Google Scholar]
- Mackay, HJ; Twelves, CJ. Protein kinase C: a target for anticancer drugs. Endocr Relat Cancer 2003, 10, 389–396. [Google Scholar]
- Malmstroem, J; Christophersen, C; Barrero, AF; Oltra, JE; Justicia, J; Rosales, A. Bioactive metabolites from a marine-derived strain of the fungus Emericella Variecolor. J Nat Prod 2002, 65, 364–367. [Google Scholar]
- Imamura, N; Nishijima, M; Adachi, K; Sano, H. Novel antimycin antibiotics, urauchimycins A and B, produced by marine actinomycete. J Antibiot 1993, 46, 241–246. [Google Scholar]
- Yu, C; Curtis, JM; Walter, JA; Wright, JLC; Ayer, SW; Kaleta, J; Querengesser, L; Fathi-Afshar, ZR. Potent inhibitors of cysteine proteases from the marine fungus Microascus longirostris. J Antibiot 1996, 49, 395–397. [Google Scholar]
- Namikoshi, M; Negishi, R; Nagai, H; Dmitrenok, A; Kobayashi, H. Three new chlorine containing antibiotics from a marine-derived fungus Aspergillus ostianus collected in Pohnpei. J Antibiot 2003, 56, 755–761. [Google Scholar]
- Kito, K; Ookura, R; Yoshida, S; Namikoshi, M; Ooi, T; Kusumi, T. Pentaketides Relating to Aspinonene and Dihydroaspyrone from a Marine-Derived Fungus Aspergillus ostianus. J Nat Prod 2007, 70, 2022–2025. [Google Scholar]
- Kito, K; Ookura, R; Yoshida, S; Namikoshi, M; Ooi, T; Kusumi, T. New Cytotoxic 14-Membered Macrolides from Marine-Derived Fungus Aspergillus ostianus. Org Lett 2008, 10, 225–228. [Google Scholar]
- Yoshida, S; Kito, K; Ooi, T; Kanoh, K; Shizuri, Y; Kusumi, T. 2008. Four Pimarane Diterpenes from Marine Fungus: Chloroform Incorporated in Crystal Lattice for Absolute Configuration Analysis by X-Ray. Chem Lett 2007, 36, 1386. [Google Scholar]
- Dettrakul, S; Kittakoop, P; Isaka, M; Nopichai, S; Suyarnsestakorn, C; Tanticharoen, M; Thebtaranonth, Y. Antimycobacterial pimarane diterpenes from the Fungus Diaporthe sp. Bioorg Med Chem Lett 2003, 13, 1253–1255. [Google Scholar]
- Höller, U; Wright, AD; Matthee, GF; König, GM; Draeger, S; Aust, H; Schulz, B. Fungi from marine sponges: Diversity, biological activity and secondary metabolites. Mycol Res 2000, 104, 1354–1365. [Google Scholar]
- Müller, WEG; Grebenjuk, VA; Pennec, G; Schröder, H; Brummer, F; Hentschel, U; Müller, IM; Breter, H. Sustainable production of bioactive compounds by sponges-cell culture and gene cluster approach: A review. Mar Biotechnol 2004, 6, 105–117. [Google Scholar]
- Petit, KE; Mondeguer, F; Roquebert, MF; Biard, JF; Pouches, YF. Detection of griseofulvin in a marine strain of Penicillium waksmanii by ion trap mass spectrometry. J Microbiol Methods 2004, 58, 59–65. [Google Scholar]
- Muth, WL; Nash, CH, III. Biosynthesis of Mycophenolic acid: Purification and characterization of S-Adenosyl-L-Methionine: Demethylmycophenolic Acid O-Methyltransferase. Antimicrob Agents Chemother 1975, 8, 321–327. [Google Scholar]
- Engel, G; Milczewski, KE; Prokopek, D; Teuber, M. Strain-specific synthesis of mycophenolic acid by P. roqueforti in blue-veined cheese. Appl Environ Microbiol 1982, 43, 1034–1040. [Google Scholar]
- Sabdono, A; Radjasa, OK. Microbial symbionts in marine sponges: Marine natural product factory. J Coast Dev 2008, 11, 57–61. [Google Scholar]
- Zhang, W; Zhang, F; Li, Z; Miao, X; Meng, Q; Zhang, X. Investigation of bacteria with polyketide synthase genes and antimicrobial activity isolated from South China Sea sponges. J Appl Microbiol 2009, 107, 567–575. [Google Scholar]
- Müller, WEG; Brummer, F; Batel, R; Müller, IM; Schröder, HC. Molecular biodiversity. Case study: Porifera (Sponges). Naturwissenschaften 2003, 90, 103–120. [Google Scholar]
- Hentschel, U; Usher, KM; Taylor, MW. Marine sponges as microbial fermenters. FEMS Microbiol Ecol 2006, 55, 167–177. [Google Scholar]
- Hoffmann, F; Rapp, HT; Reitner, J. Monitoring microbial community composition by Fluorescence In situ Hybridization during cultivation of the marine cold-water sponge Geodia barretti. Mar Biotechnol 2006, 8, 373–379. [Google Scholar]
- Taylor, MW; Schupp, PJ; Dahllof, I; Kjelleberg, S; Steinberg, PD. Host specificity in marine sponge-associated bacteria, and potential implications for marine microbial diversity. Environ Microbiol 2004, 6, 121–130. [Google Scholar]
- Webster, NS; Negri, AP; Munro, MMHG; Battershill, N. Diverse microbial communities inhabit Antarctic sponges. Environ Microbiol 2004, 6, 288–300. [Google Scholar]
- Wichels, A; Wurtz, S; Dopke, H; Schutt, C; Gerdts, G. Bacterial diversity in the breadcrumb sponge Halichondria panacea (Pallas). FEMS Microbiol Ecol 2006, 56, 102–118. [Google Scholar]
- Thiel, V; Neulinger, SC; Staufenberger, T; Schmaljohann, R; Imhoff, JF. Spatial distribution of sponge-associated bacteria in the Mediterranean sponge Tethya aurantium. FEMS Microbiol Ecol 2006, 59, 47–63. [Google Scholar]
- Taylor, MW; Schupp, PJ; Nys, R; Kjelleberg, S; Steinberg, PD. Biogeography of bacteria associated with the marine sponge Cymbastela concentrica. Environ Microbiol 2005, 7, 419–433. [Google Scholar]
- Giese, AC; Pearse, JS. Giese, AC, Pearse, JS, Eds.; Introduction: general principles. In Reproduction of Marine Invertebrates. Vol I. Acoelomate and Pseudocoelomate Metazoans; Academic Press: New York, USA, 1974; pp. 1–49. [Google Scholar]
- Turon, X; Becerro, MA; Uriz, MJ. Seasonal patterns of toxicity in benthic invertebrates: The encrusting sponge Crambe crambe (Pecilosclerida). Oikos 1996, 75, 33–46. [Google Scholar]
- Thakur, NL; Anil, AC. Antibacterial activity of the sponge Ircinia ramosa: Importance of its surface-associatedbacteria. J Chem Ecol 2000, 26, 57–71. [Google Scholar]
- Imhoff, JF; Stohr, R. Sponge-associated bacteria: general overview and special aspects of bacteria associated with Halichondria panicea. Prog Mol Subcell Biol 2003, 37, 35–57. [Google Scholar]
- Erpenbeck, D; van Soest, RWM. Status and perspective of sponge chemosystematics. Mar Biotechnol 2006, 9, 2–19. [Google Scholar]
- Proksch, P; Edrada-Ebel, R; Ebel, R. Drugs from the Sea-Opportunities and obstacles. Mar Drugs 2003, 1, 5–17. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
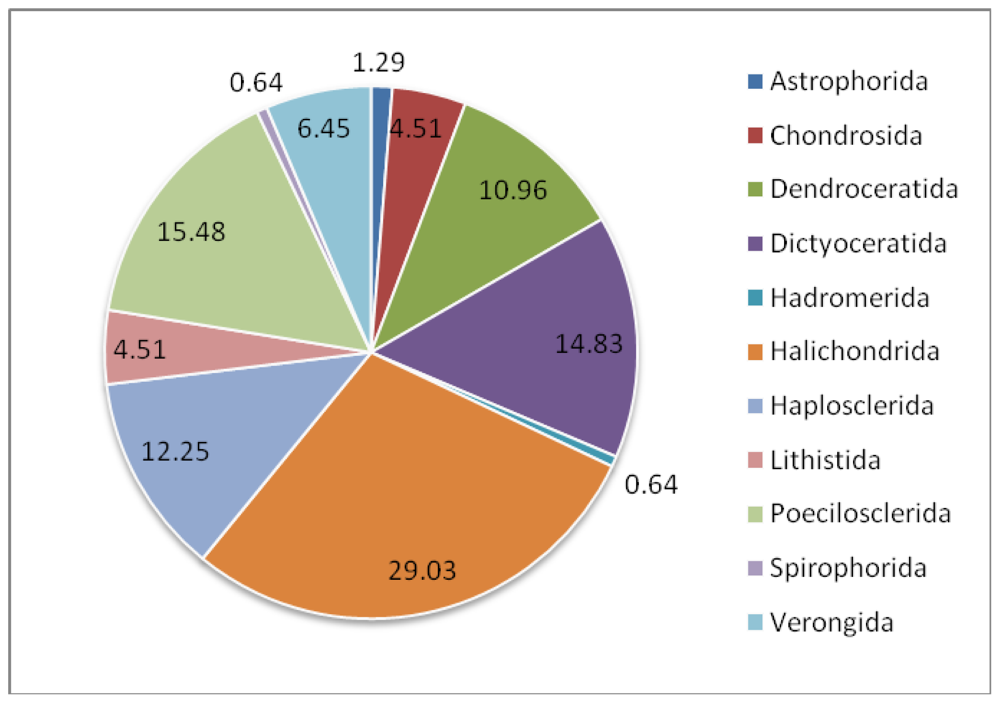
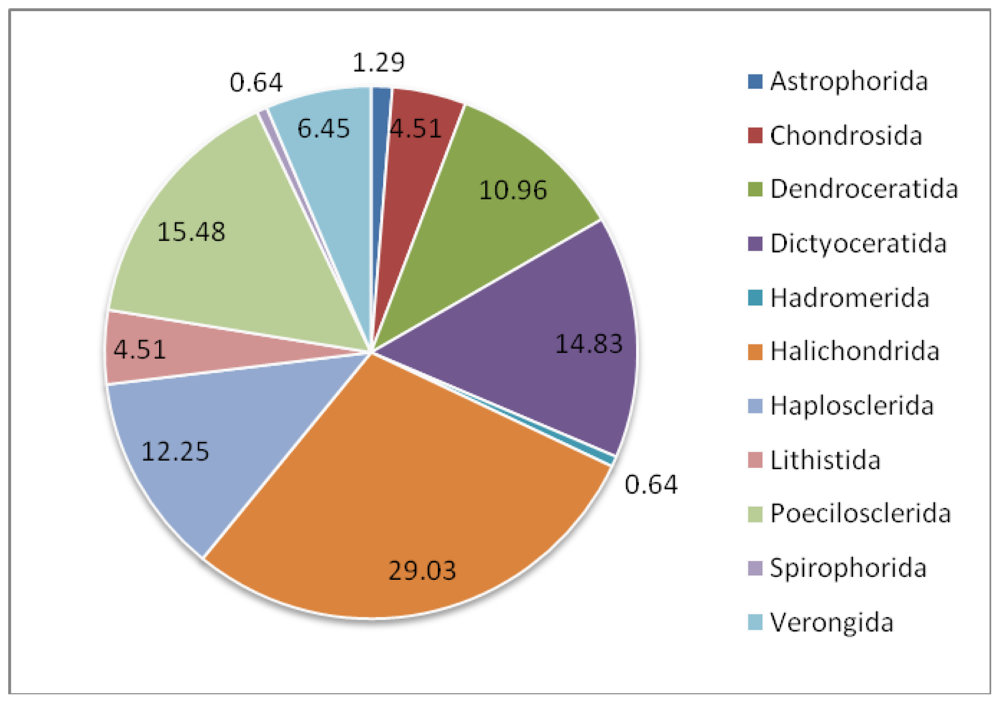
| Class: Demospongiae | |||
|---|---|---|---|
| Order | Family | Species | Reference |
| Astrophorida | Ancorinidae | Stelletta tenuis | [23,42] |
| Jaspis aff. johnstoni | [43–45] | ||
| Chondrosida | Chondrillidae | Chondrosia reniformis | [46–48] |
| Dendroceratida | Darwinellidae | Dendrilla nigra | [50–53] |
| Dictyoceratida | Dysideidae | Lamellodysidea herbacea | [40,57] |
| Dysidea sp. | [59] | ||
| Dysidea avara | [61,62] | ||
| Irciniidae | Ircinia fasciculata | [33,64,65] | |
| Spongiidae | Hyatella sp. | [66] | |
| Spongia sp. | [69] | ||
| Thorectidae | Hyrtios altum | [70–72] | |
| Hyrtios sp. | [45,73] | ||
| Hyrtios proteus | [75] | ||
| Fascaplysinopsis reticulata | [48,76] | ||
| Hadromerida | Spirastrellidae | Spirastrella vagabunda | [77,78] |
| Suberitidae | Suberites domuncula | [15,79] | |
| Halichondrida | Axinellidae | Ptilocaulis trachys | [81,82] |
| Axinella sp. 1 | [82] | ||
| Axinella sp. 2 | [84] | ||
| Axinella verrucosa | [85–87] | ||
| Axinella damicornis | [88] | ||
| Axinella sp. 3 | [89] | ||
| Halichondriidae | Halichondria okadai | [78,93–97] | |
| Halichondria panacea | [100,101] | ||
| Halichondria japonica | [79,102,104,109–113] | ||
| Acanthella acuta | [116] | ||
| Hymeniacidon perlevis | [43,44,117] | ||
| Haplosclerida | Callyspongiidae | Callyspongia aerizusa | [118] |
| Callyspongia vaginalis | [119,120] | ||
| Chalinidae | Haliclona valliculata | [121] | |
| Haliclona simulans | [25] | ||
| Haliclona sp. 1 | [123] | ||
| Haliclona sp. 2 | [2] | ||
| Niphatidae | Niphates olemda | [124,125] | |
| Petrosiidae | Petrosia ficiformis | [129–131] | |
| Xestospongia sp. | [133] | ||
| Xestospongia exigua | [79,124,134,135] | ||
| Acanthostrongylohpora sp. | [82] | ||
| Petrosia sp. | [139–141] | ||
| Lithistida | Neopeltidae | Homophymia sp. | [142] |
| Theonellidae | Theonella swinhoei | [145–148,150] | |
| Poecilosclerida | Acarnidae | Zyzzya sp. | [151,152] |
| Isodictyidae | Isodictya setifera | [153] | |
| Raspailiidae | Ectyoplasia ferox | [154] | |
| Mycalidae | Mycale plumose | [159,160] | |
| Mycale adhaerens | [48,162] | ||
| Myxillidae | Myxilla incrustance | [79,158] | |
| Spirophorida | Tetillidae | Craniella australiensis | [163] |
| Verongida | Aplysinellidae | Suberea clavata | [167] |
| Aplysinidae | Aplysina aerophoba | [170,174] | |
| Aplysina cavernicola | [41] | ||
| Pseudoceratinidae | Pseudoceratina purpurea | [90] | |
| Class: Calcarea | |||
| Leucettidae | Leucetta microraphis | [120] | |
| Class: Demospongiae | |||||
|---|---|---|---|---|---|
| Order Astrophorida Family Ancorinidae | Sponge | Symbiont | Compound | Property | Reference |
| Stelletta tenuis (South China Sea) | Alcaligenes faecalis A72 (β-Proteobacteria) | Cyclo-(L-Pro-L-Phe) | Antimicrobial | [42] | |
| Jaspis aff. johnstoni (Indo-Pacific) | Hyphomycete fungus (Deuteromycota (fungus)) | Chloriolin B | Antitumor | [43–45] | |
| Order Chondrosida Family Chondrillidae | Chondrosia reniformis (Elba, Italy) | Penicillium rugulosum (Ascomycota (fungus)) | Prugosene A1 | Anti-infective | [46–48] |
| Prugosene A2 | Anti-infective | [46–48] | |||
| Prugosene A3 | Anti-infective | [46–48] | |||
| Prugosene B1 | Anti-infective | [46–48] | |||
| Prugosene B2 | Anti-infective | [46–48] | |||
| Prugosene C1 | Anti-infective | [46–48] | |||
| Prugosene C2 | Anti-infective | [46–48] | |||
| Order Dendroceratida Family Darwinellidae | Dendrilla nigra (Vizhinjam, India) | Streptomyces dendra sp. nov. MSI051 (Actinobacteria) | Unidentified compound | Antibacterial | [51] |
| Dendrilla nigra (Kanyakumari, India) | Streptomyces sp. BLT7 (Actinobacteria) | Unidentified compound | Antibacterial | [52,53] | |
| Dendrilla nigra (South east coast, India) | Nocardiopsis dassonvillei MAD08 (Actinobacteria) | Acetic acid,-butyl-ester | Antimicrobial | [53] | |
| Ethanol, 2-(octyloxy)- | Antimicrobial | [53] | |||
| Oxalic acid, allyl-nonyl ester | Antimicrobial | [54] | |||
| 2-Isopropyl-5-methyl-1-heptanol | Antimicrobial | [53] | |||
| Butylated-hydroxytoluene | Antimicrobial | [53] | |||
| Cyclohexane-carboxylic acid, hexyl ester | Antimicrobial | [53] | |||
| Diethyl-phthalate | Antimicrobial | [53] | |||
| Pentadecanal- | Antimicrobial | [53] | |||
| 1-Tridecanol | Antimicrobial | [53] | |||
| 9-Octadecenal | Antimicrobial | [53] | |||
| Hexadecanoic acid, methyl-ester | Antioxidant, hypo-cholesterolemic, nematicide, antiandrogenic, hemolytic | [53] | |||
| n-Hexadecanoic-acid | Antioxidant, hypo-cholesterolemic, nematicide, antiandrogenic, hemolytic | [53] | |||
| Hexadecanoic-acid, ethyl ester | Antioxidant, hypo-cholesterolemic, nematicide, antiandrogenic, hemolytic | [53] | |||
| 9-Octadecenoic-acid-(Z)-, methyl-ester | Anti-inflammatory, antiandrogenic, cancer-preventive, dermatitigenic, hypo-cholesterolemic, anemiagenic | [53] | |||
| Oleic Acid | Anti-inflammatory, antiandrogenic, cancer-preventive, dermatitigenic, hypo-cholesterolemic, anemiagenic | [53] | |||
| (E)-9-Octadecenoic-acid ethyl ester | Anti-inflammatory, antiandrogenic, cancer-preventive, dermatitigenic, hypo-cholesterolemic, anemiagenic | [53] | |||
| 9-Octa-decenamide-(Z)- | Anti-inflammatory, antiandrogenic, cancer-preventive, dermatitigenic, hypo-cholesterolemic, anemiagenic | [53] | |||
| Order Dictyoceratida Family Dysideidae | Lamellodysidea herbacea (Great Barrier Reef, Australia) | Oscillatoria spongeliae (Cyanobacteria) | Dihydrodysamide C | Therapeutic (unknown action) | [57] |
| Didechloro-dihydrodysamide C | Therapeutic (unknown action) | [57] | |||
| Dysidea sp. (Eastern Samoa) | Vibrio sp. (γ-Proteobacteria) | Tetrabromo-diphenyl ethers | Cytotoxic, antibacterial | [59] | |
| Lamellodysidea herbacea (Republic of Palau) | Oscillatoria spongeliae (Cyanobacteria) | 2-(2′,4′-dibromo-phenyl)-4,6-dibromophenol | Anibacterial | [40] | |
| Dysidea avara (Adriatic Sea) | Unidentified bacterium | 2-methylthio-1,4-naphthoquinone | Antiangiogenic, antimicrobial | [61,62] | |
| Irciniidae | Ircinia fasciculata (Mediterranean Sea) | Penicillium chrysogenum (Ascomycota (fungus)) | Sorbicillactone A | Antileukemic, anti HIV | [64,65] |
| Spongiidae | Hyatella sp. | Vibrio sp. M22-1 (γ-Proteobacteria) | Andrimid | Antibiotic | [66] |
| Spongia sp. (Hawaii) | Myrothecium verrucaria 973023 (Deuteromycota (fungus)) | 3-hydroxyroridin E | Antileukemic, antitumor | [69] | |
| 13′-acetyl-trichoverrin B | Antileukemic, antitumor | [69] | |||
| Roridin A | Antileukemic, antitumor | [69] | |||
| Roridin L | Antileukemic, Antitumor | [69] | |||
| Roridin M | Antileukemic, Antitumor | [69] | |||
| Verrucarin M | Antileukemic, antitumor | [69] | |||
| Verrucarin A | Antileukemic, antitumor | [69] | |||
| Isororidin A | Antileukemic, antitumor | [69] | |||
| Epiroridin E | Antileukemic, antitumor | [69] | |||
| Trichoverrin A | Antileukemic, antitumor | [69] | |||
| Trichoverrin B | Antileukemic, antitumor | [69] | |||
| Thorectidae | Hyrtios altum (Okinawa) | Vibrio sp. (γ-Proteobacteria) | Trisindoline | Antibiotic | [70–72] |
| Hyrtios sp. (Caribbean Sea) | Aspergillus niger (Ascomycota (fungus)) | Asperazine | Antileukemic, cytotoxic | [45,74,75] | |
| Hyrtios proteus (Dry Tortugas National Park, Florida ) | Aspergillus niger (Ascomycota (fungus)) | Asperazine | Antileukemic, cytotoxic | [76] | |
| Malformin C | Antitumor | [76] | |||
| Fascaplysinopsis reticulate (Great Barrier Reef, Australia) | Pseudo-alteromonas maricaloris KMM 636T (γ-Proteobacteria) | Bromo-alterochromide A | Cytotoxic | [48,76] | |
| Bromo-alterochromide A | Cytotoxic | [48,76] | |||
| Order Hadromerida Family Spirastrellidae | Spirastrella vagabunda (Indonesia) | Unidentified fungus | 14,15-secocurvularin | Antibiotic | [77,78] |
| Suberitidae | Suberites domuncula (Northern Adriatic Sea) | α-Proteobacterium MBIC3368 (isolate 1) | Unidentified compound | Antiangiogenic, antimicrobial, hemolytic, cytotoxic | [15,79] |
| α-Proteobacterium MBIC3368 (isolate 2) | Unidentified compound | Antimicrobial, hemolytic | [15,80] | ||
| Idiomarina sp. (γ-Proteobacteria) | Unidentified compound | Hemolytic | [15,80] | ||
| Pseudomonas sp. (isolate 1) (γ-Proteobacteria) | Unidentified compound | Hemolytic, cytotoxic | [15,80] | ||
| Pseudomonas sp. (isolate 2) (γ-Proteobacteria) | Unidentified compound | Antiangiogenic, antimicrobial, hemolytic, cytotoxic | [15,80] | ||
| Order Halichondrida Family Axinellidae | Ptilocaulis trachys (Enewetak Atoll, Marshall Island, Pacific Ocean) | Lyngbya majuscula (Cyanobacteria) | Majusculamide C | Antifungal | [81,82] |
| Axinella sp. (South China Sea) | Myrothecium sp. JS9 (Deuteromycota (fungus)) | Roridin A | Antifungal | [83] | |
| Roridin D | Antifungal | [83] | |||
| Axinella sp. (Papaua New Guinea) | Penicillium citrinum (Ascomycota (fungus)) | Isocyclocitrinol A | Antibacterial | [84] | |
| 22-acetyl-isocyclocitrinol A | Antibacterial | [84] | |||
| Axinella verrucosa (Mediterranean Sea) | Penicillium sp. (Ascomycota (fungus)) | Oxaline | Anti-proliferative | [85] | |
| Griseofulvin | Antifungal | [85,86] | |||
| Communesin B | Antileukemic | [85,87] | |||
| Communesin C | Antileukemic | [85,87] | |||
| Communesin D | Antileukemic | [85,87] | |||
| Axinella sp. (Papua New Guinea) | Acremonium sp. (Ascomycota (fungus)) | Efrapeptin E | Cytotoxic, antibacterial | [90] | |
| Efrapeptin F | Cytotoxic, antibacterial | [90] | |||
| Efrapeptin Eα | Cytotoxic, antibacterial | [90] | |||
| Efrapeptin G | Cytotoxic, antibacterial | [89] | |||
| Efrapeptin H | Cytotoxic, antibacterial | [90] | |||
| RHM1 | Antibacterial | [89] | |||
| Axinella damicornis (Mediterranean Sea) | Aspergillus niger (Ascomycota (fungus)) | Bicoumanigrin | Anticancer, cytotoxic | [88] | |
| Aspernigrin B | Neuroprotective | [88] | |||
| Halichondriidae | Halichondria okadai | Alteromonas sp. (γ-Proteobacteria) | Alteramide A | Anticancer, cytotoxic | [93–95] |
| Halichondria okadai (Japan) | Trichoderma harzianum OUPS-N115 (Ascomycota (fungus)) | Trichodenone A | Antileukemic, cytotoxic | [78,96,97] | |
| Trichodenone B | Antileukemic, cytotoxic | [78,96,97] | |||
| Trichodenone C | Antileukemic, cytotoxic | [78,96,97] | |||
| Halichondria okadai | Rubritalea squalenifasciens HOact23T (Verrucomicrobiae) | Dia-polycopenedioic acid xylosyl esters A | Antioxidant | [30,48,98] | |
| Dia-polycopenedioic acid xylosyl esters B | Antioxidant | [30,48,98] | |||
| Dia-polycopenedioic acid xylosyl esters C | Antioxidant | [30,48,98] | |||
| Halichondria panacea | Unidentified bacterium | Unidentified compound | Neuroactive | [100] | |
| Halichondria panacea (Adriatic coast, Croatia) | Microbacterium sp. (Actinobacteria) | 1-O-acyl-3-[R-glucopyranosyl-(1–3)-(6-O-acyl-R-manno-pyranosyl)]-glycerol | Antitumor | [101] | |
| Halichondria japonica (Osaka Bay, Japan) | Gymnascella dankaliensis OUPS-N134 (Ascomycota (fungus)) | Gymnostatin A | Antileukemic, cytotoxic | [78,102,103,105] | |
| Gymnostatin B | Antileukemic, cytotoxic | [78,102,103,105] | |||
| Gymnostatin C | Antileukemic, cytotoxic | [78,102,103,105] | |||
| Gymnostatin F | Antileukemic, cytotoxic | [106] | |||
| Gymnostatin G | Antileukemic, cytotoxic | [106] | |||
| Gymnostatin Q | Antileukemic, anti cancer, cytotoxic | [107] | |||
| Gymnostatin R | Antileukemic, cytotoxic | [107] | |||
| Gymnasterone A | Cytotoxic | [108,109] | |||
| Gymnasterone B | Antileukemic, cytotoxic | [108,109] | |||
| Gymnasterone C | Antileukemic, cytotoxic | [108] | |||
| Gymnasterone D | Antileukemic, cytotoxic | [108] | |||
| Dankastatin A | Antileukemic, cytotoxic | [107] | |||
| Dankastatin B | Antileukemic, cytotoxic | [107] | |||
| Dankasterone A | Antileukemic, anticancer, cytotoxic | [104] | |||
| Halichondria Japonica (Japan) | Phoma sp. Q60596 (Ascomycota (fungus)) | YM-202204 | Antifungal | [110] | |
| Halichondria Japonica | Bacillus cereus QN03323 (Firmicutes) | YM-266183 | Antibacterial | [111–113] | |
| YM-266184 | Antibacterial | [111–113] | |||
| Acanthella acuta (Mediterranean Sea) | Bacillus pumilus AAS3 (Firmicutes) | GG11 | Antitumor | [116] | |
| Hymeniacidon perlevis (Nanji Island, China Sea) | Pseudo-alteromonas piscicida NJ6-3-1 (γ-Proteobacteria) | Norharman | Antimicrobial | [43,117] | |
| Hymeniacidon perlevis (Fujiazhuang coast, China) | Fusarium oxysporum DLFP2008005 (Ascomycota (fungus)) | Unidentified compound | Antibacterial, antifungal | [44] | |
| Order Haplosclerida Family Callyspongiidae | Callyspongia aerizusa (Indonesia) | Cladosporium herbarum (Deuteromycota (fungus)) | Sumiki’s acid | Antibacterial | [118] |
| Acetyl Sumiki’s acid | Antibacterial | [118] | |||
| Callyspongia vaginalis (Caribbean Sea) | Ulocladium botrylis 193A4 (Ascomycota (fungus)) | Ulocladol | Antimicrobial | [119,120] | |
| 1-hydroxy-6-methyl-8-(hydroxylmethyl)-xanthone | Antifungal | [119,120] | |||
| Chalinidae | Haliclona valliculata (Elba, Italy) | Emericella variecolor (Ascomycota (fungus)) | Evariquinone | Anti-proliferative | [122] |
| Haliclona simulans (Ireland) | Pseudo-alteromonas sp. PA2 (γ-Proteobacteria) | Unidentified | Antimicrobial | [123] | |
| Pseudo-alteromonas sp. PA4 (γ-Proteobacteria) | Unidentified | Antimicrobial | [123] | ||
| Pseudo-alteromonas sp. PA5 (γ-Proteobacteria) | Unidentified | Antimicrobial | [123] | ||
| Pseudo-alteromonas sp. PA5 (γ-Proteobacteria) | Unidentified | Antimicrobial | [123] | ||
| Halomonas sp. HM4 ( γ-Proteobacteria) | Unidentified | Antimicrobial | [123] | ||
| Psychrobacter sp. PB1 (γ-Proteobacteria) | Unidentified | Antimicrobial | [123] | ||
| Marinobacter sp. MB1 (γ-Proteobacteria) | Unidentified | Antimicrobial | [123] | ||
| Pseudovibrio sp. PV1 (α-Proteobacteria) | Unidentified | Antimicrobial | [123] | ||
| Pseudovibrio sp. PV2 ( α-Proteobacteria) | Unidentified | Antimicrobial | [123] | ||
| Pseudovibrio sp. PV4 ( α-Proteobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM1 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM2 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM3 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM4 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM5 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM6 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM7 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM8 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM9 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM10 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM11 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM12 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM14 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM16 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM17 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM18 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Streptomyces sp. SM19 (Actinobacteria) | Unidentified | Antimicrobial | [123] | ||
| Bacillus sp. BC1 (Firmicutes) | Unidentified | Antimicrobial | [123] | ||
| Bacillus sp. BC2 (Firmicutes) | Unidentified | Antimicrobial | [123] | ||
| Haliclona sp. (Tomini Bay, North Sulawesi, Indonesia) | Unidentified fungus | Hirsutanol A | Antibiotic | [123] | |
| ent-gloeosteretriol | Antibiotic | [123] | |||
| Haliclona sp.(North Java Sea, Indonesia) | Unidentified bacterium 1 | Unidentified | Antibacterial | [2] | |
| Unidentified bacterium 2 | Unidentified | Antibacterial | [2] | ||
| Unidentified bacterium 3 | Unidentified | Antibacterial | [2] | ||
| Unidentified bacterium 4 | Unidentified | Antibacterial | [2] | ||
| Unidentified bacterium 5 | Unidentified | Antibacterial | [2] | ||
| Niphatidae | Niphates olemda (Indonesia) | Curvularia lunata (Ascomycota (fungus)) | Lunatin | Antibacterial | [124,125] |
| Cytoskyrin A | Antibacterial | [124,125] | |||
| Petrosiidae | Petrosia ficiformis (Capo S. Andrea, Elba, Italy) | Penicillium brevicompactum (Ascomycota (fungus)) | Mycophenolic acid | Immuno-suppressant | [128] |
| Aspergillus insuetus (Ascomycota (fungus)) | Terretonins E | Inhibit mammalian mitochondrial respiratory chain | [132] | ||
| Terretonins F | Inhibit mammalian mitochondrial respiratory chain | [132] | |||
| Petrosia sp. (Jeju Island, Korea) | Aspergillus versicolor (Ascomycota (fungus)) | Decumbenone A | Melanin inhibitor | [139] | |
| Fellutamide C | Cytotoxic | [140] | |||
| Xestospongia sp. (Off Noumea (New Caledonia, southwest Pacific)) | Micrococcus luteus R-1588-10 (Actinobacteria) | 2,4,4′-trichloro-2′-hydroxy-diphenylether (Triclosan) | Antimicrobial | [133] | |
| Acyl-1-(acyl-6′-mannobiosyl)-3-glycerol (Lutoside) | Antimicrobial | [133] | |||
| Xestospongia exigua (Bali Sea, Indonesia) | Penicillium cf. montanense (Ascomycota (fungus)) | Xestodecalactone B | Antifungal | [79,134] | |
| Xestospongia exigua (Indonesia) | Aspergillus versicolor (Ascomycota (fungus)) | Aspergillitine | Antibacterial | [124,135] | |
| Acantho-strongylophora sp. (Indonesia) | Micromonospora sp. (Actinobacteria) | Manzamine A | Antitumor, antimalarial | [82,137, 138] | |
| Order Lithistida Family Neopeltidae | Homophymia sp. (Off Touho, New Caledonia) | Pseudomonas sp. 1537-E7 (γ-Proteobacteria) | 2-undecyl-4-quinolone | Antimalarial Anti HIV | [142] |
| 2-undecen-1′-yl-4-quinolone | Cytotoxic | [142] | |||
| 2-nonyl-4-hydroxy-quinoline N-oxide | Antibacterial, cytotoxic | [142] | |||
| Theonellidae | Theonella swinhoei (Palau) | Unidentified bacterium | Swinholide A | Cytotoxic | [144,145] |
| Candidatus Entotheonella palauensis (δ-Proteobacteria) | Theopalauamide | Antifungal | [26,144,146] | ||
| Theonella swinhoei (Philippines) | Entotheonella palauenis (δ-Proteobacteria) | Theonegramide | Antifungal | [145,147] | |
| Theonella swinhoei (Hachijojima Island, Japan) | Uncultured bacterium | Onnamide A | Antitumor | [149,150] | |
| Order Poecilosclerida Family Acarnidae | Zyzzya sp. (Fiji) | Penicillium brocae (Ascomycota (fungus)) | Brocaenol A | Cytotoxic | [151,152] |
| Brocaenol B | Cytotoxic | [151,152] | |||
| Brocaenol C | Cytotoxic | [151,152] | |||
| Isodictyidae | Isodictya setifera (Hut Point and Danger Slopes, Ross Island, Antarctica) | Pseudomonas aeruginosa (γ-Proteobacteria) | Cyclo-(L-proline-L-methionine) | Antibacterial | [153] |
| Raspailiidae | Ectyoplasia ferox (Dominica, Carribean Island) | Coniothyrium sp. 193477 (Deuteromycota (fungus)) | (3S)-(3′,5′-dihydroxyphenyl) butan-2-one | Antimicrobial | [154] |
| 2-(1′(E)-propenyl)-octa-4(E),6(Z)-diene-1,2-Diol | Antimicrobial | [154] | |||
| (3R) 6-methoxymellein | Antimicrobial | [154] | |||
| (3R)-6-methoxy-7-chloromellein | Antimicrobial | [154] | |||
| Crypto-sporiopsinol | Antimicrobial | [154] | |||
| Phoma sp. (Ascomycota (fungus)) | Epoxyphomalin A | Antitumor | [155] | ||
| Spicellum roseum 193H15 (Deuteromycota (fungus)) | Trichodermol | Anticancer | [156,157] | ||
| 8-deoxytrichothecin | Anticancer | [156,157] | |||
| Mycalidae | Mycale plumose (Qingdao coast, China) | Saccharopolyspora sp. nov. (Actinobacteria) | Metacyclo-prodigiosin | Anticancer | [159] |
| Undecyl-prodigiosin | Anticancer | [159] | |||
| Penicillium auratiogriseum (Ascomycota (fungus)) | (S)-2,4-dihydroxy-1-butyl(4-hydroxy)-benzoate | Antitumor | [160] | ||
| Fructigenin A | Antitumor | [160] | |||
| Aurantiomide B | Cytotoxic | [161] | |||
| Aurantiomide C | Cytotoxic | [161] | |||
| Mycale adhaerens | Exophiala pisciphila N110102 (Ascomycota (fungus)) | Exophilin A | Antibacterial | [48,162] | |
| Myxillidae | Myxilla incrustance (Helgoland, Germany) | Microsphaeropsis sp. H5-50 (Anamorphic fungus) | Microsphaeropsisin | Antifungal | [79,154] |
| (R)-mellein | Antimicrobial | [154] | |||
| (3R,4S)-hydroxymellein | Antimicrobial | [154] | |||
| (3R,4R)-hydroxymellein | Antimicrobial | [154] | |||
| 4,8-dihydroxy-3,4-dihydro-2H-naphthalen-1-one | Antimicrobial | [154] | |||
| Order Spirophorida Family Tetillidae | Craniella australiensis (South China Sea) | Streptomyces sp. DA11 (Actinobacteria) | Chitinase | Antifungal | [163] |
| Order Verongida Family Aplysinellidae | Suberea clavata (Great Barrier Reef, Australia) | Salinospora sp. (Actinobacteria) | Rifamycin B | Antibiotic | [164,166] |
| Rifamycin SV | Antibiotic | [164,166] | |||
| Aplysinidae | Aplysina aerophoba (Mediterranean coast, France) | Bacillus subtilis A184 (Firmicutes) | Surfactin, iturin and fengycin | Antifungal, antibacterial, hemolytic | [170] |
| Bacillus subtilis A190 (Firmicutes) | Surfactin | Antifungal, hemolytic | [170] | ||
| Bacillus subtilis A202 (Firmicutes) | Iturin | Antifungal, hemolytic | [170] | ||
| Bacillus pumilus A586 (Firmicutes) | Pumilacidin containing β-hydroxy fatty-acid | Antibacterial | [170] | ||
| Aplysina aerophoba (Mediterranean Sea) | Microsphaeropsis sp. (Anamorphic fungus) | 10-Hydroxy-18-methoxyl-betaenone | Protein Kinase C ɛ inhibitor | [174] | |
| Aplysina aerophoba (Banyuls sur Mer) | Bacillus sp. SB8 (Firmicutes) | Unidentified compound | Antibacterial | [41] | |
| Bacillus sp. SB17 (Firmicutes) | Unidentified compound | Antibacterial | [41] | ||
| Micrococcus sp. SB58 (Actinobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Enterococcus sp. SB91 (Firmicutes) | Unidentified compound | Antibacterial | [41] | ||
| Arthrobacter sp. SB95 (Actinobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Unidentified bacteria SB122 | Unidentified compound | Antibacterial | [41] | ||
| Unidentified bacteria SB144 | Unidentified compound | Antibacterial | [41] | ||
| α-Proteobacteria SB6 | Unidentified compound | Antibacterial | [41] | ||
| α-Proteobacteria SB55 | Unidentified compound | Antibacterial | [41] | ||
| α-Proteobacteria SB63 | Unidentified compound | Antibacterial | [41] | ||
| α-Proteobacteria SB89 | Unidentified compound | Antibacterial | [41] | ||
| α-Proteobacteria SB156 | Unidentified compound | Antibacterial | [41] | ||
| α-Proteobacteria SB197 | Unidentified compound | Antibacterial | [41] | ||
| α-Proteobacteria SB202 | Unidentified compound | Antibacterial | [41] | ||
| α-Proteobacteria SB207 | Unidentified compound | Antibacterial | [41] | ||
| α-Proteobacteria SB214 | Unidentified compound | Antibacterial | [41] | ||
| Vibrio halioticoli SB177 (γ-Proteobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Pseudo-alteromonas sp. SB181 (γ-Proteobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Pseudo-alteromonas sp. SB182 (γ-Proteobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Pseudo-alteromonas sp. SB183 (γ-Proteobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Pseudo-alteromonas sp. SB185 (γ-Proteobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Pseud-oalteromonas sp. SB186 (γ-Proteobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Pseudo-alteromonas sp. SB192 (γ-Proteobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Pseudo-alteromonas sp. SB194 (γ-Proteobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Pseudo-alteromonas sp. SB200 (γ-Proteobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Pseudo-alteromonas sp. SB208 (γ-Proteobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Pseudo-alteromonas sp. SB213 (γ-Proteobacteria) | Unidentified compound | Antibacterial | [41] | ||
| Pseudo-ceratinidae | Pseudoceratina purpurea (Fiji) | Metarrhizium sp. 001103 (Ascomycota (fungus)) | Destruxin A | Antitumor | [90] |
| Destruxin B2 | Antitumor | [90] | |||
| Desmethyl B | Antitumor | [90] | |||
| E chlorohydrin | Antitumor | [90] | |||
| E2 chlorohydrin | Antitumor | [90] | |||
| Class: Calcarea | |||||
| Order Clathrinida Family Leucettidae | Leucetta microraphis (Great Barrier Reef, Australia) | Unidentified cyanobacteria | Leucamide A | Antitumor | [120] |
| Unidentified sponges | |||||
| Unidentified | Unidentified (Venezuela) | Emericella variecolor (Ascomycota (fungus)) | Varitriol | Anticancer | [176] |
| Varixanthone | Antimicrobial | [176] | |||
| Unidentified | Streptomyces sp. Ni-80 (Actinobacteria) | Urauchimycin A | Antibiotic | [177] | |
| Urauchimycin B | Antibiotic | [177] | |||
| Unidentified (Harrington Point, Otago Harbor, New Zealand) | Microascus longirostris SF-73 (Ascomycota (fungus)) | Cathestatin A | Cysteine protease inhibitor | [178] | |
| Cathestatin B | Cysteine protease inhibitor | [178] | |||
| Cathestatin C | Cysteine protease inhibitor | [178] | |||
| Unidentified (Pohnpei, The federated state of Micronesia) | Aspergillus ostianus 01F313 (Ascomycota (fungus)) | 8-chloro-9-hydroxy-8,9-deoxyasperlactone | Antibacterial | [179] | |
| 9-chloro-8-hydroxy-8,9-deoxyasperlactone | Antibacterial | [179] | |||
| 9-chloro-8-hydroxy-8,9-deoxyaspyrone | Antibacterial | [179] | |||
| Aspinonene | Antileukemic | [48,180] | |||
| Dihydroaspyrone | Antileukemic | [48,180] | |||
| Aspergillide A | Antileukemic | [48,181] | |||
| Aspergillide B | Antileukemic | [48,181] | |||
| Aspergillide C | Antileukemic | [48,181] | |||
| Cryptosphaeria eunomia (Ascomycota (fungus)) | Diaporthein A | Antibacterial | [48,182] | ||
| Diaporthein B | Antibacterial | [48,182] | |||
| Category | Chemical diversity |
|---|---|
| Antiandrogenic | Fattyacid esters, fatty acids |
| Antiangiogenic | Quinone |
| Anticancer | Quinone, steroid, fatty acid esters, fatty acids, diketopiperazine, alkaloid, terpenes, terpenoids, trichoverroids, prodigiosin derivative |
| AntiHIV | Quinolone derivative |
| Anti-inflammatory | Fatty acid esters, fatty acid |
| Antimalarial | Alkaloid, quinolone derivative |
| Antimicrobial | Polyketide, glycopeptides, α-pyrone derivative, peptide, protein, antimycin, lipopeptides, polybrominated biphenyl ether, cyclic depsipeptide, terpenes, pentaketides, furan carboxylic acid, alkaloid, diketopiperazine, anthraquinone, chromones, steroid, lactone, quinolone derivative, trisindole derivative, macrolactam, ethers, phenol derivative |
| Antiinfective | Polyketides |
| Antioxidant | Fatty acid esters, fatty acid, carotenoic acid |
| Anti-respiratory | Terpenoids |
| Antitumor | Diglucosyl-glycerol, polyketides, alkaloids, cyclopeptides, glycoglycerolipid, benzoic acid derivative, terpenoids, terpenes, trichoverroids |
| Hemolytic | Fatty acid ester, fatty acids |
| Hypocholesterolemic | Fatty acid ester, fatty acids |
| Immunosupressant | Mycophenolic acid |
| Melanin inhibitor | Polyketide |
| Nematicide | Fatty acid ester, fatty acids |
| Neuroactive | Unknown |
| Neuroprotective | Dihydropyridine |
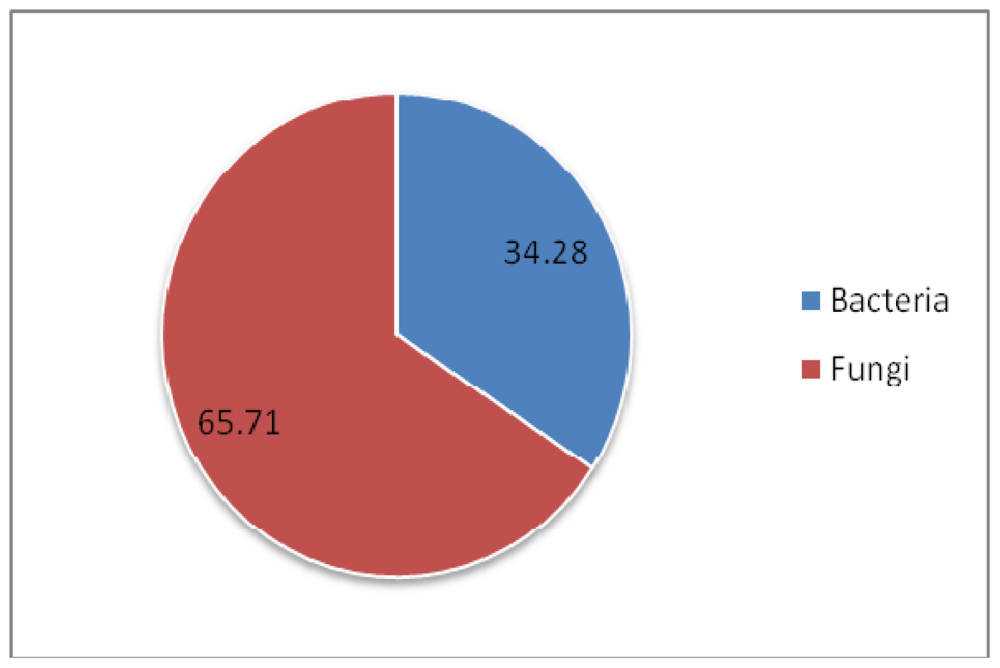
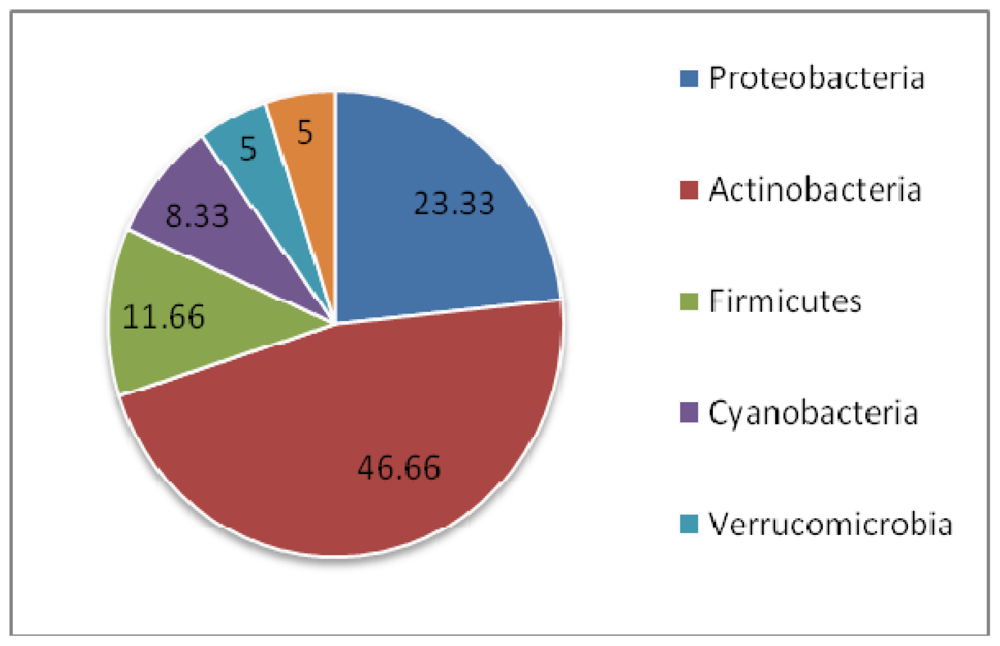
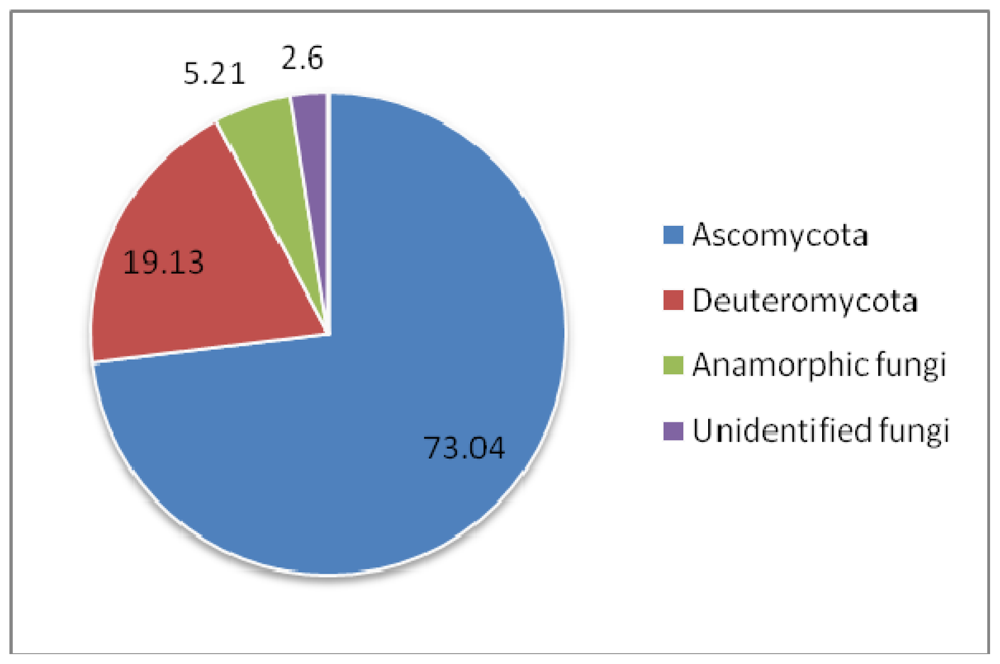
| Symbiont | Sponge order | Compound fuction | ||
|---|---|---|---|---|
| Bacteria | ↔ | Dendroceratida | → | Antiandrogenic |
| Bacteria | ↔ | Dictyoceratida, Hadromerida | → | Antiangiogenic |
| Bacteria | ↔ | Halichondrida, Dendroceratida, Poecilosclerida | → | Anticancer |
| Fungi | ↔ | Dictyoceratida, Halichondrida, Haplosclerida, Poecilosclerida | ||
| Bacteria | ↔ | Lithistida | → | AntiHIV |
| Fungi | ↔ | Dictyoceratida | ||
| Fungi | ↔ | Chondrosida | → | Anti-infective |
| Bacteria | ↔ | Dendroceratida | → | Ant-inflammatory |
| Bacteria | ↔ | Lithistida, Haplosclerida | → | Antimalarial |
| Bacteria | ↔ | Astrophorida, Dendroceratida Dictyoceratida, Hadromerida, Haplosclerida, Halichondrida, Lithistida, Poecilosclerida, Spirophorida, Verongida | → | Antimicrobial |
| Fungi | ↔ | Hadromerida, Halichondrida, Haplosclerida, Poecilosclerida | ||
| Bacteria | ↔ | Dendroceratida, Halichondrida | → | Antioxidant |
| Fungi | ↔ | Haplosclerida | → | Anti-respiratory |
| Bacteria | ↔ | Clathrinida, Halichondrida, Haplosclerida, Lithistida | → | Antitumor |
| Fungi | ↔ | Astrophorida, Dictyoceratida, Poecilosclerida, Verongida | ||
| Bacteria | ↔ | Hadromerida, Dendroceratida | → | Hemolytic |
| Bacteria | ↔ | Dendroceratida | → | Hypocholesterolemic |
| Fungi | ↔ | Haplosclerida | → | Immunosuppressant |
| Fungi | ↔ | Haplosclerida | → | Melanin inhibitor |
| Bacteria | ↔ | Dendroceratida | → | Nematicide |
| Bacteria | ↔ | Halichondrida | → | Neuroactive |
| Fungi | ↔ | Halichondrida | → | Neuroprotective |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Thomas, T.R.A.; Kavlekar, D.P.; LokaBharathi, P.A. Marine Drugs from Sponge-Microbe Association—A Review. Mar. Drugs 2010, 8, 1417-1468. https://doi.org/10.3390/md8041417
Thomas TRA, Kavlekar DP, LokaBharathi PA. Marine Drugs from Sponge-Microbe Association—A Review. Marine Drugs. 2010; 8(4):1417-1468. https://doi.org/10.3390/md8041417
Chicago/Turabian StyleThomas, Tresa Remya A., Devanand P. Kavlekar, and Ponnapakkam A. LokaBharathi. 2010. "Marine Drugs from Sponge-Microbe Association—A Review" Marine Drugs 8, no. 4: 1417-1468. https://doi.org/10.3390/md8041417