Natural Abundance 14C Content of Dibutyl Phthalate (DBP) from Three Marine Algae

1
Tohoku Pharmaceutical University, Aoba-ku, Sendai 981-8558, Japan
2
Department of Ocean Sciences, Tokyo University of Marine Science and Technology, Minato-ku, Tokyo 108-8477, Japan
3
The Nagoya University Museum, Chikusa-ku, Nagoya 464-8601, Japan
*
Author to whom correspondence should be addressed.
Mar. Drugs 2006, 4(4), 290-297; https://doi.org/10.3390/md404290
Submission received: 19 September 2006
/
Accepted: 31 October 2006
/
Published: 1 November 2006
Abstract
:Analysis of the natural abundance 14C content of dibutyl phthalate (DBP) from two edible brown algae, Undaria pinnatifida and Laminaria japonica, and a green alga, Ulva sp., revealed that the DBP was naturally produced. The natural abundance 14C content of di-(2-ethylhexyl) phthalate (DEHP) obtained from the same algae was about 50–80% of the standard sample and the 14C content of the petrochemical (industrial) products of DBP and DEHP were below the detection limit.
1. Introduction
Dialkyl phthalates, such as di-(2-ethylhexyl) phthalate (DEHP), diisononyl phthalate (DINP), diisodecyl phthalate (DIDP) and dibutyl phthalate (DBP), are petrochemicals used as plasticizers or solvents in a variety of industrial products. These chemicals have been thought to be environmental pollutants and detected in soils, sediments, terrestrial and marine waters, and also living organisms. DEHP has been isolated from terrestrial and marine organisms including plants [1], marine algae [2–5], and fungal and bacterial culture broths [6,7]. However, it is difficult to determine whether DEHP was produced by these organisms, the sample was contaminated during the separation process, or DEHP accumulated in the organisms, because DEHP has been used abundantly in petrochemical phthalate esters. More than 65% of all the phthalate esters used in Japan in 2003 were DEHPs followed by DINPs (>26%), DIDPs (about 3%), and DBPs (about 1%), and this relative ratio has changed little in recent years.
Interestingly, there has been no report of the isolation of DINP and DIDP from organisms, whereas DBP has been isolated from plants, marine algae, bacteria, and fungi [2, 8–11]. Diethyl phthalate (DEP), a less common petrochemical product, was obtained as a chemotactic factor from the culture medium of Helicobacter pylori[12] and also detected in the broth of Streptomyces sp. isolated from Antarctic sediment [13]. Moreover, di-(2-methylheptyl) phthalate, which is not a petrochemical used in the chemical industry, was isolated from a plant Hypericum hyssopifolium (Guttiferae) [14]. Chen reported the incorporation of 14C into DBP and DEHP by culturing the filamentous phase of a red alga, Bangia atropurpurea, with 14C-enriched NaHCO3[2]. DBP was enriched by about 171 times the background level and DEHP, by 5.7 times.
These reports led us to investigate the origin of the phthalate esters more closely. Recently, Teuten et al. demonstrated that polybrominated diphenyl esters isolated from a True’s beaked whale (Mesoplodon mirus) were naturally produced compounds and not derived from artificial products by analyzing the natural abundance 14C content of the isolated compounds and industrial products [15,16].
Therefore, we intended to investigate if DBP and DEHP obtained from marine algae are natural products or contaminants of petrochemical products using a similar method [17,18].
In this report, we suggest that DBP isolated from the CH2Cl2-methanol extracts of Undaria pinnatifida (Harvey) Suringar (Alariaceae, Phaeophyceae), Laminaria japonica Areschoug (Laminariaceae, Phaeophyceae) and Ulva sp. (Ulvaceae, Chlorophyceae) was a natural product based on an analysis of the natural abundance 14C content of the isolates.
2. Results and Discussion
Industrial products of DBP and DEHP contain very small amounts of 14C which cannot be detected even by the most sensitive method, accelerator mass spectrometry (AMS), because these compounds are made from fossil oils. If these phthalate esters are produced by living organisms, natural abundance 14C will be detected in the same amounts as modern carbon.
The isolation of DBP and DEHP was first examined using Ulva sp. collected in May 2005 to determine solvents suitable for the extraction process and procedures for concentrating lipophilic compounds. The conditions for HPLC and GC were established using authentic samples of DBP and DEHP.
The extraction and separation of U. pinnatifida were performed according to the procedures used for small amounts of Ulva sp. The aqueous residue obtained after the evaporation of organic solvents from the CH2Cl2-MeOH extract of the alga, was dissolved in MeOH and adsorbed on Hyflo Supercell ®. The adsorbent was placed in a glass column and lipophilic compounds were eluted with CH2Cl2. The lipophilic fraction was adsorbed on Hyflo Super-cell and applied to a silica gel column with hexane. The silica gel column was eluted successively with hexane and hexane-EtOAc (gradient). The eluate was monitored by TLC (hexane-EtOAc = 4:1) and three fractions containing DBP and/or DEHP were obtained. The first fraction (Fr. 1) contained DEHP and other compounds and the second (Fr. 2) and third (Fr. 3) fractions contained DBP, DEHP and other lipophilic compounds. DBP (1.24 mg) was separated by HPLC (ODS) with 80% MeOH-H2O from Frs. 2 and 3, and DEHP was recovered by changing the solvent to 100% MeOH. This DEHP fraction was combined with Fr. 1 and DEHP (2.14 mg) was isolated by HPLC with 100% MeOH.
Since the particles of Hyflo Super-cell were too small, dispersion and formation of masses occurred during the evaporation of solvents when large amounts of extract or lipophilic fraction were adsorbed. Therefore, silica gel was used in lieu of Hyflo Super-cell for the separation from L. japonica and Ulva sp. The other separation procedures for these algae were the same as those for U. pinnatifida.
DBP and DEHP obtained from three algae were identified and analyzed the purity by 1H NMR and GC-mass spectra. The 1H NMR spectra of DBP and DEHP from U. pinnatifida were clean and only a very few faint signals due to contaminants were detected. On the other hand, pigments and lipophilic compounds disturbed the separation of DBP and especially DEHP from L. japonica and Ulva sp. DEHP fractions obtained by the first HPLC separation from L. japonica and Ulva sp. contained a large quantity of contaminants, which were most probably unsaturated fatty acids. These fractions were separated repeatedly by HPLC. The 1H NMR spectra of the final fraction of DBP and DEHP from these two algae still showed the signals due to the contaminants. It is unclear if the difference in the amounts/species of contaminants between U. pinnatifida and the other two algae is due to the nature of these algae or the season when they were collected. L. japonica and Ulva sp. were harvested in the winter (cold water), when they usually contain more unsaturated fatty acids.
The concentration (w/w%) of DBP and DEHP in the final samples, analyzed by GC-MS utilizing the selected ion monitoring (SIM) method, was 60% (DBP) and 70% (DEHP) from L. japonica and 75% (DBP) and 60% (DEHP) from Ulva sp. It was not feasible to conduct further purification since the separation of contaminant(s) is difficult and would significantly reduce the amounts of DEHP. We thought that the measurement of 14C with these samples would give decipherable results because petrochemical compounds contain no measurable 14C and biogenetic products contain modern levels of 14C. Therefore, we submitted DBP and DEHP samples obtained from L. japonica and Ulva sp. without further separation, together with DBP and DEHP from U. pinnatifida.
Natural abundance 14C content was analyzed at the Tandetron AMS 14C Dating Laboratory, Center for Chronological Research at Nagoya University by the AMS method. The results are summarized in Table 1. The 14C concentration is shown as percent modern carbon (pMC) [19]. This unit represents a percentage value of the ratio of sample 14C concentration to the absolute 14C concentration, which is defined as corresponding to AD1950.
The industrial samples of DBP and DEHP did not give measurable 14C concentration as they are petrochemical products. The DBP samples from three algae contained modern 14C, which indicated that DBP was synthesized naturally and not of industrial origin. The higher 14C content in DBP can be ascribed to the enrichment of the 14C level caused by the testing of nuclear bombs during the 1950s and 1960s [20]. The enrichment of the atmospheric 14C is now 15% of the level before testing [21,22]. The 14C may have accumulated in marine sediments.
Significant amounts of 14C were detected in the DEHP samples from three algae. In the most reliable (highly purified) sample obtained from U. pinnatifida, about 50% of the 14C concentration was modern carbon. The DEHP samples from L. japonica and Ulva sp. showed even higher percentages. The contaminant(s) (unsaturated fatty acid(s)) in the former two samples would raise the 14C concentration. It may be that remarkable amounts of industrial DEHP were incorporated into the algae and/or contaminated the sample during the isolation procedures and that some amount of DEHP was biosynthesized to give the significant 14C content.
This study revealed that DBP obtained from two edible brown algae, U. pinnatifida and L. japonica, and a green alga, Ulva sp., was a natural product since modern 14C was detected in the isolated DBP samples.
DEHP may also be produced naturally, but the results of this study were not conclusive. The culture with 14C-enriched NaHCO3 gave no clear result for DEHP either [2]. Therefore, further studies will be required to confirm whether this compound is biosynthesized by these algae.
Di-(2-methylheptyl) phthalate isolated from H. hyssopifolium must be a biogenetic product of this plant [14], because it has not been synthesized. DBP and DEP detected in the culture media of microorganisms would also be produced by microorganisms [8–13]. These phthalate esters were in the environment long before their commercial manufacture and release.
Roles of phthalate esters are not known. It may be true that they are stored in the cell membrane and affect the flexibility of the cell [2]. The physiological functions of phthalate esters are interesting subjects for future studies.
3. Experimental Section
3.1. General experimental procedures
1H NMR (400 MHz) and 13C NMR (100 MHz) spectra were measured on a JEOL AL-400 spectrometer in CDCl3. GC-MS data were obtained with a Shimadzu GCMS-QP5050A spectrometer using Class-5000 software and a TC-70 column (0.25 μm x 30 m): carrier gas, He (0.4 ml/min); interface and ion source temperatures, 300 °C. For a typical elution, the column temperature was first set at 100 °C for 5 min, raised to 300 °C at 2 °C/min, and kept at 300 °C for 20 min. DBP and DEHP were eluted at about 47 and 60 min, respectively. HPLC was carried out with a Hitachi L-2130 Pump and L-2400 UV Detector (280 nm) equipped with a D-2500 data processor using a Senshu Pack Pegasil-ODS column (10 mm x 250 mm) at a flow rate of 3 ml/min. DBP was eluted at about 20 min with 80% MeOH-H2O. DEHP was eluted at about 10 min with 100% MeOH. Standard samples (petrochemical products) of DBP and DEHP were provided by SHOWA ETHER CO., LTD. and CG ESTER CORPORATION, respectively. All other solvents and reagents were of the highest grade commercially available and used without further purification.
3.2. Algal material and extraction
U. pinnatifida was collected at Kanazawahakkei, Yokohama, Japan in April 2005. The alga (about 13.5 kg) was cut into pieces and extracted four times with CH2Cl2-MeOH (2:1, each 10.5 l). The organic extract was evaporated to give the aqueous residue (772.6 g). L. japonica was collected at Hachinohe, Aomori Prefecture, Japan in January 2006. The alga (about 9.3 kg) was cut into pieces and extracted four times with CH2Cl2-MeOH (2:1, each 12 l). The organic extract was evaporated to give the aqueous residue (780.2 g). Ulva sp. was collected at Yokohama Marine Park, Yokohama, Japan in May and December 2005. The alga collected in December (about 13.5 kg) was cut into pieces and extracted four times with CH2Cl2-MeOH (2:1, each 12 l). The organic extract was evaporated to give the aqueous residue (1.23 kg).
3.3. Isolation of DBP and DEHP
The aqueous residue obtained from the organic extract of U. pinnatifida was dissolved in MeOH and the solution was mixed with Hyflo Super-cell® (1 kg). MeOH and water were evaporated and the dried adsorbent was placed in a glass column (diameter, 70 mm; length, 600 mm). The column was eluted with CH2Cl2 (4 l) to give a lipophilic fraction (32.5 g). The lipophilic fraction was dissolved in CH2Cl2, and the solution was mixed with 100 g of Hyflo Super-cell. After evaporation of the organic solvent, the dried adsorbent was mixed with hexane and poured on to a silica gel column (700 g) made up with hexane. The column was eluted with 2 l of hexane followed by hexane-EtOAc = 9:1 (2 l) and 4:1 (2 l). The eluates were monitored by TLC (hexane-EtOAc = 4:1) and divided into three fractions (Frs. 1–3). Fr. 1 contained DEHP and other compounds. DBP and DEHP were detected in two fractions (Frs. 2 and 3). Fr. 2 (120.2 mg) was separated by HPLC (ODS) with 80% MeOH-H2O to obtain a DBP fraction and then a DEHP fraction was recovered with 100% MeOH. Fr. 3 (12.1 mg) was subjected to a similar HPLC separation to give DBP and DEHP fractions. Two DBP fractions were combined and separated by HPLC (80% MeOH-H2O) to afford 1.24 mg of DBP. The DEHP fractions obtained from Frs. 2 and 3 were combined with Fr. 1, and DEHP (2.14 mg) was isolated by HPLC separation (100% MeOH).
The aqueous residue from Ulva sp. was dissolved in MeOH and the solution was mixed with silica gel (1.22 kg). MeOH and water were evaporated and the dried silica gel was placed in a glass column (diameter, 70 mm; length, 600 mm). The column was eluted with CH2Cl2 (3 l) to give a lipophilic fraction (216.8 g). The CH2Cl2 solution of the lipophilic fraction was mixed with 250 g of silica gel, and, after evaporation of the organic solvent, the adsorbed silica gel was applied to a silica gel column (800 g) with hexane. The column was eluted successively with 1 l each of hexane and hexane-CH2Cl2 = 9:1, 7:3, 1:1, 3:7, and 1:4. The eluates were monitored by TLC (hexane-EtOAc = 4:1) and divided into five fractions. DBP and DEHP were contained into two fractions (Frs. 3 and 4). Fr. 3 (127.1 mg) was separated by HPLC (ODS) with 80% MeOH-H2O to obtain a DBP fraction and then a DEHP fraction was recovered with 100% MeOH. Fr. 4 (240.7 mg) was subjected to a similar HPLC separation to give DBP and DEHP fractions. The combined DBP fraction (12.8 mg) was further separated by repeated HPLC (80% MeOH-H2O) to afford 1.78 mg of DBP. DEHP (3.87 mg) was isolated from the DEHP fraction (135.7 mg) by repeated HPLC separation (100% MeOH).
DBP (1.93 mg) and DEHP (2.47 mg) were isolated from the aqueous residue obtained from the organic extract of L. japonica by similar procedures.
DBP and DEHP isolated from these algae were identified based on the 1H NMR and GC-mass spectra with the standard samples of DBP and DEHP.
3.4. Quantitative analysis of DBP and DEHP
DBP and DEHP isolated from L. japonica and Ulva sp. were analyzed by GC-MS using the selected ion monitor (SIM) method. The base peak at m/z 149 was selected for monitoring. A calibration table was created by injecting standard solutions containing 10, 5, 2, 1, 0.5, and 0.1 μg of the authentic DBP or DEHP. The calibration curve was calculated by linear-least squares regression. The samples in the same solvent were injected and the chromatogram was integrated in terms of concentration. The quantitative analysis of samples was performed between 0.5 and 2 μg/injection.
The GC column temperature used for the analysis of DBP was first set at 100 °C for 6 min, raised to 300 °C at 2 °C/min, and held at 300 °C for 20 min. DBP was eluted at around 50 min. The amounts of DBP isolated from L. japonica and Ulva sp. were about 60 and 75 (w/w)%, respectively.
The column temperature for DEHP was set at 100 °C for 6 min and raised to 300 °C at 4 °C/min, and DEHP was eluted at around 38 min. The amounts of DEHP isolated from L. japonica and Ulva sp. were about 70 and 60 (w/w)%, respectively.
3.5. Analysis of natural abundance 14C content
Natural abundance 14C content was measured by the following three steps at the Tandetron AMS 14C Dating Laboratory, Center for Chronological Research, Nagoya University utilizing a model 4130- AMS system [18,23]: (1) each organic compound was first burned into CO2, (2) the CO2 was reduced to graphite, and (3) the graphite was analyzed by accelerator mass spectrometry (AMS) (Kitagawa, 1993; Nakamura et al., 2004). The results (Table 1) were expressed as percent modern carbon (pMC), a percentage value of the ratio of sample 14C concentrations to the absolute 14C concentration defined as corresponding to AD 1950 [17,19].
Acknowledgements
We thank Professor T. Nakamura and Ms. A. Ikeda of the Center for Chronological Research, Nagoya University for analysis of the natural abundance 14C content. We also thank SHOWA ETHER CO., LTD. and CG ESTER CORPORATION for the standard petrochemical products of DBP and DEHP, respectively.
References and Notes
- Lee, K. H.; Kim, J. H.; Lim, D. S.; Kim, C. H. Anti-leukemic and anti-mutagenic effects of di(2- ethylhexyl) phthalate isolated from Aloe vera Linne. J. Pharm. Pharmacol 2000, 52, 593–598. [Google Scholar]
- Chen, C. Y. Biosynthesis of di-(2-ethylhexyl) phthalate (DEHP) and di-n-butyl phthalate (DBP) from red alga––Bangia atropurpurea. Water Res 2004, 38, 1014–1018. [Google Scholar]
- Noguchi, T.; Ikawa, M.; Uebel, J. J.; Andersen, K. K. Lipid constituents of the red algae Ceramium rubrum. A search for antimicrobial and chemical defense substances. In Marine algae in pharmaceutical science; Hoppe, H. A., Levring, T., Tanaka, Y., Eds.; Walter de Gruyter & Co: New York, 1979; pp. 711–718. [Google Scholar]
- Sastry, V. M. V. S.; Rao, G. R. K. Dioctyl phthalate, and antibacterial compound from the marine brown alga—Sargassum wightii. J. Appl. Phycol 1995, 7, 185–186. [Google Scholar]
- Stefanov, K.; Konaklieva, M.; Brechany, E. Y.; Christie, W. W. Fatty acid composition of some algae from the black sea. Phytochemistry 1988, 27, 3495–3497. [Google Scholar]
- Al-Bari, M. A.; Bhuiyan, M. S.; Flores, M. E.; Petrosyan, P.; Garcia-Varela, M.; Islam, M. A. Streptomyces bangladeshensis sp. nov., isolated from soil, which produces bis-(2- ethylhexyl)phthalate. Int. J. Syst. Evol. Microbiol 2005, 55, 1973–1977. [Google Scholar]
- Amade, P.; Mallea, M.; Bouaicha, N. Isolation, structural identification and biological activity of two metabolites produced by Penicillium olsonii Bainier and Sartory. J. Antibiot 1994, 47, 201–207. [Google Scholar]
- El-Naggar, M. Y. M. Dibutyl phthalte and the antitumor agent F5A1, two metabolites produced by Streptomyces nasri submutant H35. Biomed. Lett 1997, 55, 125–131. [Google Scholar]
- Lee, D. -S. Dibutyl phthalate, an α-glucosidase inhibitor from Streptomyces melanosporofaciens. J. Biosci. Bioeng 2000, 89, 271–273. [Google Scholar]
- Roy, R. N.; Laskar, S.; Sen, S. K. Dibutyl phthalate, the bioactive compound produced by Streptomyces albidoflavus 321.2. Microbiol. Res 2006, 161, 121–126. [Google Scholar]
- Savard, M. E.; Miller, J. D.; Blais, L. A.; Seifert, K. A.; Samson, R. A. Secondary metabolites of Penicillium bilaii strain PB-50. Mycopathologia 1994, 127, 19–27. [Google Scholar]
- Keire, D. A.; Anton, P.; Faull, K. F.; Ruth, E.; Walsh, J. H.; Chew, P.; Quisimoro, D.; Territo, M.; Reeve, J. R., Jr. Diethyl phthalate, a chemotactic factor secreted by Helicobacter pylori. J. Biol. Chem 2001, 276, 48847–48853. [Google Scholar]
- Ivanova, V.; Oriol, M.; Montes, M. J.; Garcia, A.; Guinea, J. Secondary metabolites from a Streptomyces strain isolated from Livingston Island, Antarctica. Z. Naturforsh 2001, 56, 1–5. [Google Scholar]
- Cakir, A.; Mavi, A.; Yildirim, A.; Duru, M. E.; Harmandar, M.; Kazaz, C. Isolation and characterization of antioxidant phenolic compounds from the aerial parts of Hypericum hyssopifolium L. by activity-guided fractionation. J. Ethnopharm 2003, 87, 73–83. [Google Scholar]
- Teuten, E. L.; Xu, L.; Reddy, C. M. Two abundant bioaccumulated halogenated compounds are natural products. Science 2005, 307, 917–920. [Google Scholar]
- Reddy, C. M.; Xu, L.; Eglinton, T. I.; Boon, J. P.; Faulkner, D. J. Radiocarbon content of synthetic and natural semi-volatile halogenated organic compounds. Environ. Pollut 2002, 120, 163–168. [Google Scholar]
- Nakamura, T. Instruments for radiation measurement in life sciences (4). I. High sensitivity measurements of environmental and tracer radioisotopes with accelerator mass spectrometry. (Japanese). Radioisotopes 2003, 52, 145–171. [Google Scholar]
- Nakamura, T.; Niu, E.; Oda, H.; Ikeda, A.; Minami, M.; Ohta, T.; Oda, T. High precision 14C measurements with the HVEE Tandetron AMS system at Nagoya University. Nucl. Instr. Meth 2004, B223–224, 124–129. [Google Scholar]
- Stuiver, M.; Polach, H. A. Discussion: reporting of 14C data. Radiocarbon 1977, 19, 355–363. [Google Scholar]
- Nydal, R. Further investigation on the transfer of radiocarbon in nature. J. Geophys. Res 1968, 73, 3617–3635. [Google Scholar]
- Levin, I.; Kromer, B. Twenty years of atmospheric 14CO2 observations at Schauinsland station, Germany. Radiocarbon 1997, 39, 205–218. [Google Scholar]
- Levin, I.; Kromer, B.; Schoch-Fischer, H.; Bruns, M.; Münnich, M.; Berdau, D.; Vogel, J. C.; Münnich, K. O. 25 years of tropospheric 14C observations in central Europe. Radiocarbon 1985, 27, 1–19. [Google Scholar]
- Kitagawa, H.; Masuzawa, T.; Nakamura, T.; Matsumoto, E. A batch preparation method for graphite targets with low background for AMS 14C measurements. Radiocarbon 1993, 35, 295–300. [Google Scholar]
- Samples Availability: Not available.
Figure 1.
Structures of dibutyl phthalate (DBP) and di-(2-ethylhexyl) phthalate (DEHP).
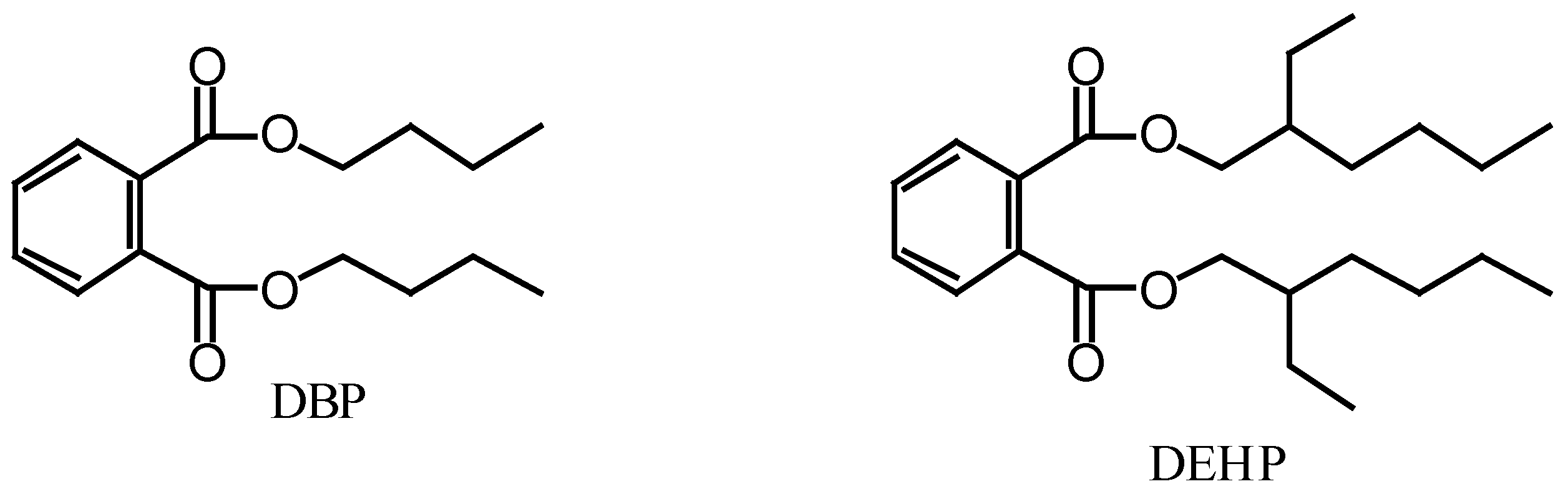

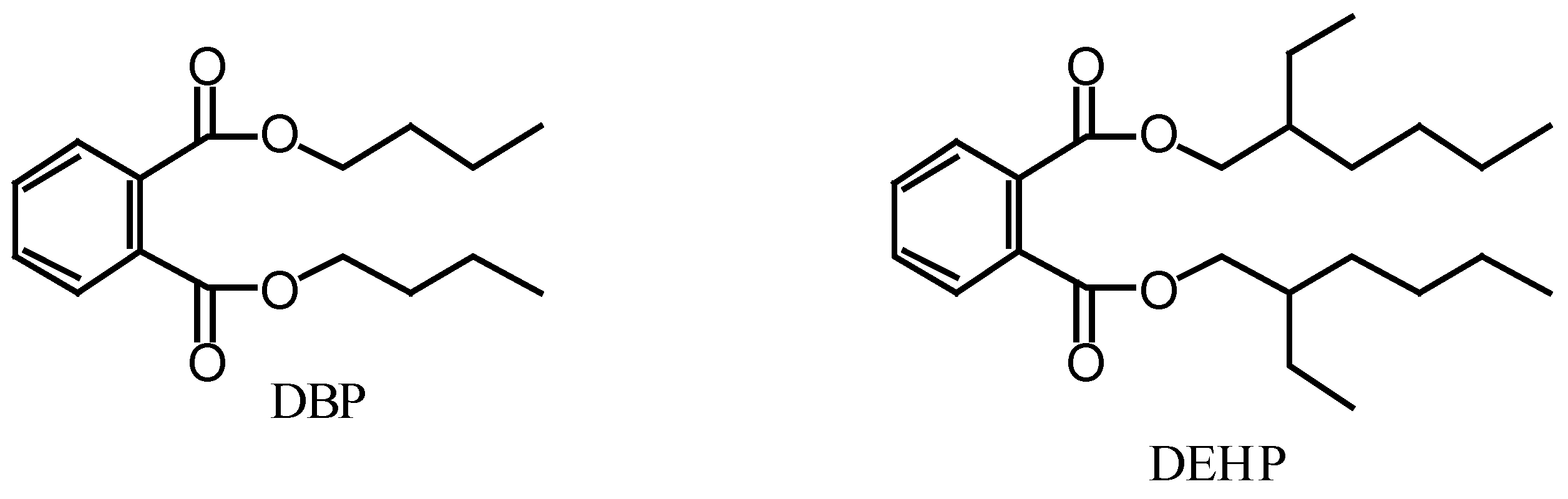{kind=link}
Table 1.
Natural abundance 14C content of dibutyl phthalate (DBP) and di-(2-ethylhexyl) phthalate (DEHP) of marine algal and industrial origins.
| compound | sourcea | 14C content (pMC)b | compound | sourcea | 14C content (pMC)b |
|---|---|---|---|---|---|
| DBP | U. pinnatifida | 249.4 ± 0.6 | DEHP | U. pinnatifida | 49.8 ± 0.2 |
| DBP | L. japonica | 281.2 ± 0.6 | DEHP | L. japonica | 85.6 ± 0.2 |
| DBP | Ulva sp. | 138.9 ± 0.4 | DEHP | Ulva sp. | 87.2 ± 0.2 |
| DBP | industrial | 0.21 ± 0.02 | DEHP | industrial | 0.11 ± 0.02 |
aU. pinnatifida: Undaria pinnatifida; L. japonica: Laminaria japonica; industrial: petrochemical product.
bPercent Modern Carbon: relative ratio (%) of the 14C concentration of the sample to that of the standard.
© 2006 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
MDPI and ACS Style
Namikoshi, M.; Fujiwara, T.; Nishikawa, T.; Ukai, K. Natural Abundance 14C Content of Dibutyl Phthalate (DBP) from Three Marine Algae. Mar. Drugs 2006, 4, 290-297. https://doi.org/10.3390/md404290
AMA Style
Namikoshi M, Fujiwara T, Nishikawa T, Ukai K. Natural Abundance 14C Content of Dibutyl Phthalate (DBP) from Three Marine Algae. Marine Drugs. 2006; 4(4):290-297. https://doi.org/10.3390/md404290
Chicago/Turabian StyleNamikoshi, Michio, Takeshi Fujiwara, Teruaki Nishikawa, and Kazuyo Ukai. 2006. "Natural Abundance 14C Content of Dibutyl Phthalate (DBP) from Three Marine Algae" Marine Drugs 4, no. 4: 290-297. https://doi.org/10.3390/md404290