Hypopigmenting Effects of Brown Algae-Derived Phytochemicals: A Review on Molecular Mechanisms


1
Department of Food Science and Nutrition, Pukyong National University, 45 Yongso-Ro, Nam-Gu, Busan 48513, Korea
2
Department of Foodservice Management, Woosong University, Daejeon 34606, Korea
*
Author to whom correspondence should be addressed.
Mar. Drugs 2017, 15(10), 297; https://doi.org/10.3390/md15100297
Submission received: 29 August 2017
/
Revised: 11 September 2017
/
Accepted: 20 September 2017
/
Published: 24 September 2017
(This article belongs to the Special Issue Marine Products for Health and Beauty)
Abstract
:There is a rapid increase in the demand for natural hypopigmenting agents from marine sources for cosmeceutical and pharmaceutical applications. Currently, marine macroalgae are considered as a safe and effective source of diverse bioactive compounds. Many research groups are exploring marine macroalgae to discover and characterize novel compounds for cosmeceutical, nutraceutical, and pharmaceutical applications. Many types of bioactive secondary metabolites from marine algae, including phlorotannins, sulfated polysaccharides, carotenoids, and meroterpenoids, have already been documented for their potential applications in the pharmaceutical industry. Among these metabolites, phlorotannins from brown algae have been widely screened for their pharmaceutical and hypopigmenting effects. Unfortunately, the majority of these articles did not have detailed investigations on molecular targets, which is critical to fulfilling the criteria for their cosmeceutical and pharmaceutical use. Very recently, a few meroterpenoids have been discovered from Sargassum sp., with the examination of their anti-melanogenic properties and mechanisms. Despite the scarcity of in vivo and clinical investigations of molecular mechanistic events of marine algae-derived hypopigmenting agents, identifying the therapeutic targets and their validation in humans has been a major challenge for future studies. In this review, we focused on available data representing molecular mechanisms underlying hypopigmenting properties of potential marine brown alga-derived compounds.
1. Introduction
Skin pigmentation is a complex process that includes synthesis of melanin pigments in melanocytes at the basal layer of epidermis and their distribution to neighboring keratinocytes. Skin color is primarily determined by the type, intensity and distribution of melanin pigments. The role of melanin is also essential to protect the skin from deleterious effects of solar ultraviolet (UV) radiation, which is responsible for the excess generation of reactive oxygen species (ROS) and leads to photocarcinogenesis [1], premature skin aging [2] and other damages. Although melanin pigments have an important role in protecting the skin against photocarcinogenesis, irregular hyperpigmentation is not desirable and is considered as a skin pigmentary disorder. Hyperpigmentation is caused by abnormal accumulation of melanin pigments in the skin with major skin hyperpigmentation disorders including melasma, freckles, lentigo, moles, leukoplakia, and so on [3,4]. Furthermore, periorbital hyperpigmentation, post-inflammatory hyperpigmentation, drugs and cosmetics-induced hyperpigmentation, age spots and hormonal factor-mediated hyperpigmentation are also common factors in causing imbalanced pigmentation in the skin [5,6]. There are various commercial depigmenting agents currently being used in skin-whitening cosmetics. However, most of them are not completely satisfactory due to their low efficiency or safety concerns [6,7,8]. Hydroquinone, which was introduced for clinical use as a depigmenting agent since 1961, has been widely used for years to treat skin hyperpigmentation disorders, especially against melasma [6,9]. However, it has been reported to cause contact dermatitis and exogenous ochronosis, which is darkening of the area of skin exposed to hydroquinone [9]. It was also reported to be unstable in cosmetics and is an easily-oxidized ingredient in cosmetic formulations [9]. Considering the consequences of long-term treatments [10], its application in cosmetics has been banned since 2001 by the European Commission [6]. Arbutin, a terrestrial plant originated derivative of hydroquinone, is another commercial skin-whitening agent. Although it showed strong tyrosinase (TYR) inhibitory activity, it was shown to lack efficiency in melanocyte pigmentation and in clinical trials [8]. As it is a glycosylated hydroquinone, arbutin may have similar risks as hydroquinone [7,11]. Kojic acid is an effective skin-lightening agent, but previous studies reported that kojic acid is associated with allergic contact dermatitis [12] and enhancement of carcinogenicity [13]. Therefore, there is growing demand for the examination of effective and safe agents from natural sources to treat hyperpigmentation disorders and to use in skin-whitening cosmetics in Asian countries.
More than 70% of the Earth’s surface consists of a marine habitat, which is a huge source of diversified population of flora and fauna [14]. Among marine biota, marine algae are rich in a diverse range of beneficial compounds with strong safety profiles [15,16,17]. Marine macroalgae are broadly divided into three major groups: brown algae (phaeophyceae); red algae (rhodophyceae), and; green algae (chlorophyceae). Brown algae are specifically rich sources of phenolic compounds with high antioxidant activities [18] and, thus, it could be a useful ingredient for nutricosmetics [19]. Marine phenolic compounds, such as phlorotannins, only exist in brown algae and possess a high potential for pharmaceutical and nutraceutical applications [18,20]. Brown algae are also rich in meroterpenoids that are proven for diverse health benefits due to their strong antioxidant activities [21,22,23,24,25,26,27]. Marine algae-derived sulfated polysaccharides, such as fucoidan, laminarans, ulvan and carrageenan, are being utilized in food, cosmetic and pharmaceutical industries [15]. It is important to note that cosmeceuticals from natural sources have higher market value compared to synthetic compounds [20]. There are many studies examining the hypopigmentation and skin protection roles of flavonoids, most of which are of terrestrial origin [28]. Until now, a considerable amount of effort has been given on searching hypopigmenting compounds from terrestrial sources. However, there are limited data on the hypopigmenting activities of marine alga-derived compounds focusing on their molecular mechanistic aspects.
Understanding the molecular mechanisms of any bioactive compound is very important for their proper application. A lack of clear understanding on therapeutic targets and effects results in unfortunate outcomes with huge economic losses during drug discovery. Therefore, after the initial screening, it is very important to know the detailed molecular events and identify specific targets of potential compounds in specific cell types and suitable animal models before entering the pre-clinical and clinical trials in the long process of drug development. In this review, we will summarize the potential hypopigmenting role of marine brown alga-derived compounds focusing on their molecular mechanistic approach. We will also focus on current status, major gaps and future directions on this issue.
2. Major Pathways Involved in Melanogenesis
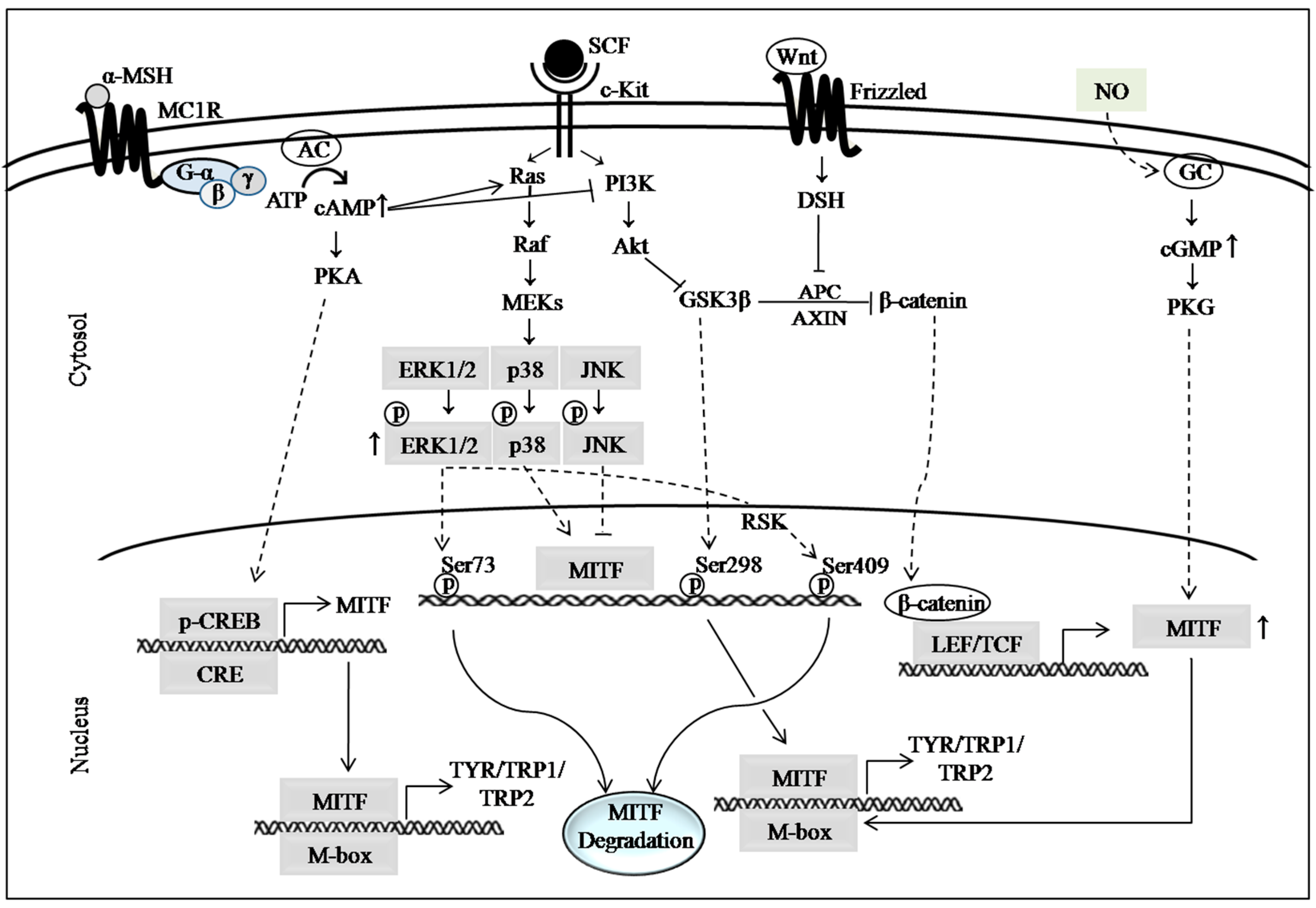
There are several well documented signaling pathways that are involved in the regulation of skin pigmentation. Among them, the cyclic adenosine monophosphate (cAMP) signaling pathway is considered as the classical signaling pathway that plays a pivotal role in melanogenesis [29]. Other major signaling pathways are also recognized for mediating melanogenic pathways, including mitogen-activated protein kinase (MAPK) signaling, phosphatidylinositol 3-kinase (PI3K)/Akt signaling and Wnt/β-catenin signaling pathways (Figure 1). The cAMP signaling pathway also initiates cross-talk with the extracellular signal-regulated kinase (ERK) 1/2, PI3K/Akt and Wnt/β-catenin signaling pathways. The molecular events in these pathways are briefly described in the following sections.
2.1. cAMP Signaling Pathway
The cAMP signaling pathway initiates when extracellular signals, such as α-melanocyte stimulating hormone (α-MSH) or adrenocorticotrophic hormone (ACTH), binds to the melanocortin 1 receptor (MC1R), which is a G-protein coupled seven transmembrane receptor. The conversion of cellular ATP to cAMP, the second messenger in melanogenic process, is catalyzed by adenylate cyclase (AC) once it is activated by G-protein. There are several stimuli, such as UV radiation, α-MSH, ACTH, forskolin, isobutylmethylxanthine or other cAMP elevating agents, which stimulate cellular responses to cAMP induction. Elevated levels of cAMP activate protein kinase A (PKA) through binding of cAMP to the binding domain of PKA. PKA consists of regulatory subunits and catalytic subunits. In the unstimulated state, the two catalytic subunits of PKA remain bound to the regulatory subunit dimer as an inactive holoenzyme, which has two cAMP-binding domains on each regulatory subunit [35]. Binding of cAMP to regulatory subunits of PKA results in the dissociation and activation of its catalytic subunits, which leads to the translocation of PKA to the nucleus and phosphorylation of its substrate cAMP-response element-binding protein (CREB) [5,36]. There are two reported phosphorylation residues in CREB, such as Ser129 and Ser133, which are associated with its activation. Ser133 is considered as the major site of phosphorylation for its activation [37]. Nonetheless, Jeong et al. [38] demonstrated the anti-melanogenic effects of baicalin through inhibition of CREB phosphorylation at Ser129, while phosphorylation level at Ser133 residue remained unchanged. CREB is one of the major transcription factors for the expression (or activation) of microphthalmia-associated transcription factor (MITF), which is responsible for the regulation of melanogenic enzymes [39]. Once CREB is activated, it binds to the CRE motif of the MITF promoter and transactivate this key regulator of melanogenic enzymes: TYR, tyrosinase-related protein (TRP)1, and TRP2 (Figure 1) [40,41]. Although these three enzymes are involved in melanin synthesis, the TYR is considered as the rate limiting enzyme for melanin synthesis [42]. In the cAMP signaling pathway, there are several critical points where skin-whitening compounds are involved in exerting their hypopigmenting effects. The major therapeutic target or regulatory points are: suppression of cAMP induction [30], inhibition of cAMP binding to PKA [36,43], inhibition of CREB activation and MITF downregulation [30], and inhibition of MC1R expression [33].
2.2. MAP Kinase Signaling Pathways
MAPK signaling is initiated by the activation of the c-Kit receptor, which is also known as tyrosine kinase receptor, with stem cell factor (SCF). The paracrine factor SCF is primarily produced by keratinocytes in response to several stimuli. After stimulation with SCF, the c-Kit transduces the signal that culminates in posttranslational modification of MITF, which alters its transcriptional activity or stability [32]. This signal can be mediated by three different kinases, such as extracellular signal regulated kinase (ERK), c-Jun N-terminal kinase (JNK) and p38 MAPK. Among them, ERK1/2 signaling is the most widely studied and strong signal, which is also considered a negative feedback mechanism for melanogenesis [44]. ERK1/2 becomes activated upon phosphorylation by Raf/MEK, before becoming translocated to the nucleus where they phosphorylate MITF at the Ser73 residue. This leads to subsequent ubiquitin-dependent proteasomal degradation of MITF [45,46]. Furthermore, they phosphorylate MITF at Ser409 through ribosomal S6 kinase (RSK), which also results in proteasome-mediated degradation of MITF [47]. ERK1/2-mediated degradation of MITF is recognized as one of the most attractive therapeutic target for anti-melanogenic compounds [48]. However, a previous study reported that ERK1/2 activation did not result in phosphorylation of MITF at the Ser73 residue, although they confirmed degradation of MITF [49]. Another study demonstrated that proteasomal degradation of MITF is not always followed by ERK activation [38]. ERK1/2 signaling also has cross-talk with cAMP signaling as the elevation of cellular cAMP can activate Ras/ERK signaling to induce MITF degradation [41,50].
The phosphorylation and activation of p38 MAPK in B16 melanoma cells was found to increase melanin synthesis through increased expression of MITF and TYR via CREB phosphorylation [51,52]. A previous investigation using an anti-melanogenic compound, c-phycocyanin, from spirulina, showed suppressed TYR expression through inhibition of p38 MAPK-mediated CREB activation [46]. Similarly, another study also reported the positive correlation between the suppression of p38 MAPK phosphorylation and the downregulation of MITF [33]. However, they did not clarify whether p38 directly acts on MITF phosphorylation or via CREB in this study. Phosphorylation and activation of stress-activated protein kinase, JNK, has also been reported to inhibit melanogenesis via downregulation of MITF [53]. On the other hand, Han et al. [54] demonstrated the positive correlation of suppression of total JNK with downregulation of MITF and TRP1 leading to attenuated melanogenesis and TYR activity in B16F10 cells. However, JNK-mediated suppression of pigmentation by marine algae-derived compounds has not yet been reported by any study.
2.3. PI3K/Akt Signaling Pathway
Similar to MAPK signaling, PI3K/Akt signaling cascade is also initiated by SCF/c-Kit interaction. Involvement of PI3K/Akt signaling in the melanogenic process was reported by several studies [55,56]. Akt, also known as protein kinase B, is a serine/threonine protein kinase. It is a downstream signaling molecule to PI3K. Akt is activated by phosphorylation on Ser473 and Thr308 residues via activation of PI3K [57]. After being activated, Akt phosphorylates the Ser9 residue of glycogen synthase kinase 3β (GSK3β), resulting in it being transformed into its inactive form [58]. However, a previous study in B16 cells reported that cAMP inhibits Akt phosphorylation and activation, which leads to activation of GSK3β by its dephosphorylation [29]. The activated form of GSK3β phosphorylates MITF at the Ser298 residue, which enhances the binding of MITF to the M-box sequence of the TYR promoter [29]. This cross-talk of cAMP with PI3K/Akt signaling cascade was reported in a previous study to occur via inhibition of PI3K activity in B16F10 cells [59]. Therefore, the phosphorylation and activation of PI3K/Akt could be a potential therapeutic approach for skin hyperpigmentation disorders. The Akt signaling pathway also has cross-talk with the Wnt/β-catenin signaling, which works via the Akt/GSK3β/β-catenin cascade [60].
2.4. Wnt/β-Catenin Signaling Pathway
The Wnt/β-catenin signaling pathway has been reported by several studies, which is involved in melanin synthesis [60,61]. There are several Wnt ligands that activate Wnt/β-catenin signaling, such as Wnt1, Wnt3a and Wnt8. Wnt-1 has been known to enhance melanocyte expansion and differentiation in mouse neural crest cells [62]. Activation of the frizzled receptor and low-density lipoprotein receptor-related protein 5/6 co-receptor by Wnt ligands phosphorylate and inactivate GSK3β, which results in increased cytoplasmic level of β-catenin. The accumulated β-catenin translocates into the nucleus and acts as a coactivator of MITF in association with lymphocyte enhancer factor 1 (LEF1), resulting in the transactivation of MITF. In the absence of Wnt signaling, a protein complex consisting of axin, adenomatous polyposis coli (APC), GSK3β and casein kinase 1 (CK1) leads to the phosphorylation of β-catenin in the N-terminal serine/threonine residues, which is followed by its proteasomal degradation. A previous study demonstrated that andrographolide, a plant-originated diterpenoid, inhibited melanin synthesis through decreased phosphorylation of GSK3β and enhanced degradation of β-catenin in B16F10 cells, human epidermis melanocytes (HEM), and ultraviolet B (UVB)-induced guinea-pig skin [60]. Suppression of pigmentation in normal human melanocytes (NHM) was caused by the enhancement of proteasome-dependent β-catenin degradation by cardamonin [61]. A previous study also reported cross-talk between cAMP signaling and Wnt/β-catenin signaling pathways in melanoma cells and NHM [63]. In this study, they showed α-MSH stimulated the activation of PKA and attenuated GSK3β. They also demonstrated that cAMP/PKA-mediated phosphorylation of β-catenin at Ser675 residue led to its stability and transcriptional activity. Therefore, the inhibition of GSK3β phosphorylation or enhancement of β-catenin degradation could be a potential therapeutic target for hypopigmenting compounds against skin hyperpigmentation disorders. However, the anti-melanogenic activity of marine macroalga-derived compounds via Wnt/β-catenin signaling has not yet been reported.
2.5. Autophagy, Nitric Oxide (NO) Signaling and Other Mechanistic Targets of Hypopigmenting Agents
Autophagy is a process through which damaged or defective organelles and unnecessary cellular aggregates are degraded by autophagosome-lysosomal machinery, which is known to facilitate the recycling of nutrients during starvation. Similarly, autophagy has a role in the destruction and removal of defective melanosomes [64]. Only a few investigations have reported the regulation of melanogenesis by the activation of autophagy in melanocytes, although the role of autophagy in melanogenic process is not well established [64,65]. Kim et al. [65] demonstrated ARP101(C20H26N2O5S)-induced activation of autophagy in α-MSH-stimulated Melan-a cells by punctate structure formation with autophagy marker green fluorescence protein fused-LC3 protein (GFP-LC3), and increased conversion of LC3I to LC3 II protein. Moreover, by electron microscopic analysis, they demonstrated the engulfment of melanosomes by autophagosomes, which resulted in the downregulation of pigmentation. In another study, Van Den Boorn et al. [66] reported suppression of pigmentation by monobenzone-induced polyubiquitination of TYR and macroautophagocytic degradation of melanosomes in human melanoma cells.
The NO signaling pathway was also reported to regulate melanogenesis [67]. Keratinocytes produce NO in response to UV-stimulation and NO interacts with guanylate cyclase (GC), which leads to increased levels of cyclic guanosine monophosphate (cGMP) in melanocytes. An elevated level of cGMP induces melanogenesis through the enhancement of MITF expression. UV-induced NO was also shown to enhance melanogenesis in alpaca skin melanocytes by stimulating MITF phosphorylation, while MITF levels remained unchanged [68]. Moreover, NO stimulated keratinocytes to produce excess α-MSH and also induced cAMP signaling by elevated expression of MC1R in melanocytes [69]. The inhibition of melanosome maturation [70] and trafficking to surrounding keratinocytes [71] were also reported to inhibit melanogenesis. However, these signaling mechanisms have not yet been reported in the case of marine macroalga-derived compounds.
3. Hypopigmenting Effects of Marine Brown Algae-Derived Phenolic Compounds
Phlorotannins, polymers of phloroglucinol (1,3,5-trihydroxybenzene), are marine polyphenols found only in brown algae (Figure 2 and Table 1) [20]. These compounds, also known as phloroglucinol derivatives, are markedly different from polyphenols of terrestrial origin. Algal phlorotannins have a wide range of applications, including anti-melanogenic [18,55,72], anti-skin aging [73], anti-oxidant [18,74,75], anti-inflammatory [76], hepatoprotective [74] and anti-allergic [77] activity. It is noteworthy that most of these properties of phlorotannins are valuable in the development of nutraceuticals and cosmeceuticals. Among brown alga-derived phlorotannins, phloroglucinol [78], eckol [78], dieckol [18,78], diphlorethohydroxycarmalol [79] and octaphlorethol A [72,80] were reported as potent TYR inhibitors, although there are a lack of in vitro and in vivo study on detailed molecular mechanisms of hypopigmenting action of these compounds. Heo et al. [18] isolated three types of phlorotannins, such as phloroglucinol, eckol and dieckol, from Ecklonia cava, and demonstrated the anti-melanogenic activity of eckol and dieckol on the basis of inhibition of mushroom TYR and melanin synthesis in B16F10 cells. In this study, they identified dieckol as a promising skin-whitening agent. Kang et al. [78] screened seventeen marine algae using mushroom TYR and identified phloroglucinol, eckstolonol, eckol, dieckol and phlorofucofuroeckol A from E. stolonifera as TYR inhibitors. They also reported dieckol as a potent TYR inhibitor (IC50 2.16 μg/mL), which showed activity three times higher than that of kojic acid. Our research team studied the hypopigmenting properties of another phlorotannin, dioxinodehydroeckol (isolated from E. stolonifera), in α-MSH-stimulated B16F10 cells and found PI3K/Akt-mediated transcriptional downregulation of MITF, leading to downregulation of melanogenic enzymes, which are namely TYR, TRP1 and TRP2 [55]. Cha et al. [7] screened aqueous extracts of 43 types of marine algae based on mushroom TYR inhibition and identified a red alga, Schizymenia dubyi (IC50 9.08 μg/mL), and three brown algae, Endarachne binghamiae (IC50 27.16 μg/mL), Sargassum siliquastrum (IC50 19.85 μg/mL) and E. cava (IC50 18.00 μg/mL) as potent TYR inhibitors. They further demonstrated the inhibitory effects of E. cava and S. siliquastrum on TYR activity and melanin synthesis in both B16F10 cells and Zebrafish model. Interestingly, in their investigations, the extracts of E. cava caused strong TYR inhibition (92%) in B16 cells, although it was much weaker (48%) in Zebrafish. However, they did not report any molecular event in this study. Jang et al. [81] isolated 4-hydroxyphenethyl alcohol from a brown alga, Hizikia fusiformis. They demonstrated inhibition of mushroom TYR activity and melanin content in B16F10 cells and remarkable reduction of UVB-induced hyperpigmented spots in brown guinea-pig skin after eight weeks of topical application. They also did not report any molecular mechanisms in hypopigmentation in their study.
4. Hypopigmenting Effects of Meroterpenoids
The brown algae are rich sources of meroterpenoid compounds, which are beneficial to general health [21,30,87]. In particular, the Sargassum genus was reported to contain high amount of meroterpenoids [21]. Algal meroterpenoids have anti-inflammatory [21,87,88,89,90], antioxidant [22], anti-ageing [23], anti-atherosclerotic [24,91], anti-adipogenic [25,92], anti-diabetic [26], anti-carcinogenic [93,94] and neuroprotective [95] activities. Recently, we demonstrated the hypopigmenting effects of ethanolic extract from S. serratifolium in B16F10 cells and identified three active meroterpenoid compounds, including sargahydroquinoic acid, sargaquinoic acid and sargachromenol (Figure 3), on the basis of their inhibitory activity on melanin synthesis in α-MSH-stimulated B16F10 cells [30]. We also elucidated that the extract from S. serratifolium inhibited hyperpigmentation in B16F10 cells through regulation of MITF via cAMP/CREB and ERK signaling pathways (Table 1). To the best of our knowledge, there was no examination of the anti-melanogenic activity of algal meroterpenoids before this report.
5. Hypopigmenting Effects of Fucoxanthin
Fucoxanthin is a group of carotenoids found in brown algae. The information on the effects of fucoxanthin on melanogenesis is very limited. Fucoxantin was reported to suppress TYR activity and melanogenesis in B16 murine melanoma cells. Furthermore, this has been seen in vivo in guinea pig and mouse skin [85]. In mice, the suppression of melanin biosynthesis was reported by both topical and oral treatments with fucoxanthin, although topical treatments resulted in better effects. This study has provided an important focus on the expression levels of melanogenic receptors in UV-irradiated mice and guinea pig skin. They found that topical treatment of 1% fucoxanthin significantly suppressed mRNA levels of endothelin receptor A (EDNRA), p75 neurotrophin receptor (p75NTR), prostaglandin E receptor 1 (EP1) and MC1R in mice. It also suppressed COX-2 expression, which downregulates prostaglandin (PG) in epidermis. Interestingly, although fucoxantin slightly suppressed TYR mRNA expression, there was no significant suppression. Therefore, they reported that fucoxanthin mainly suppressed TRP1 instead of the TYR. They suggested the suppression of PG and its receptor, EP1, in addition to MC1R by fucoxanthin, which has an inhibitory effect on melanogenesis. They also demonstrated the suppression of pigmentation in guinea pigs by a daily intake of low amount of fucoxanthin (0.001% in diet). Therefore, it can be a promising candidate for the formulation of cosmeceutical.
6. Hypopigmenting Effects of Non-Phenolic Compounds
Fucoidans, a fucose-rich sulfated polysaccharide, are predominantly found in marine brown algae and echinoderms [96]. Fucoidans have been shown to inhibit the activity of TYR [84,97], matrix metalloproteases (MMPs) and elastase [98]. Several studies indicated the high potential of fucoidan for anti-skin aging [96], anti-inflammatory [99] and antioxidant [100] activity, which demonstrates its cosmetic potential. Several previous articles reviewing bioactivity and therapeutic potential of fucoidans had a distinct lack of information regarding its hypopigmenting activity [101,102]. However, in a clinical test with 20 subjects, Fitton et al. [96] demonstrated that brown alga, F. vesiculosus extract (containing 58.6% fucoidan and 33.7% polyphenol) at 0.3% (w/v) attenuated the melanin index of age spots, increased brightness and reduced wrinkles. In another study, fucoidan, isolated from F. vesiculosus, was demonstrated to attenuate melanin content in spontaneously immortalized murine melanocyte Mel-Ab cells via ERK-mediated downregulation of MITF and TYR expression [84]. In contrast, a recent study had no inhibitory effects of fucoidan, isolated from the same alga, on TYR, TRP1, TRP2 and MITF expression in B16 murine melanoma cells, although it inhibited cellular melanin and TYR activity [42]. They suggested that fucoidan upregulated TYR expression, which was positively correlated to its apoptosis-stimulating activity in melanoma cells. Moreover, their data indicated that fucoidan suppressed the biological activity of TYR, which leads to inhibition of melanin synthesis. However, there is still very limited information on the depigmenting effects of fucoidans. Therefore, extensive studies will be needed regarding the screening of hypopigmenting fucoidans and characterization of their mechanistic events in vitro and in vivo.
7. Current Gaps and Future Directions
The necessity for identification and utilization of novel pharmaceuticals, cosmeceuticals and nutraceuticals from marine bioresources has been realized by many researchers. The world market for hypopigmentation is also huge for these marine-derived bioactive products. However, the sustainable use of marine resources, standardization of isolation process ensuring bioavailability and efficiency, safety profiling of isolated compounds and optimization of suitable doses are important. The stability of active compounds is also important for their commercial utilization. The development of an aquaculture technique for target algae could be a good option to ensure a sustainable supply for commercial utilization. The harvesting season and maturation stage are also important as there are seasonal and age variation in the biochemical composition and amount of some active components in marine algae.
The bioactivity of many compounds depends on their chemical structure and conformation, bioavailability, as well as type and position of functional groups. There are several novel extraction and separation techniques, such as supercritical CO2 extraction, membrane separation and ultrasonic-aided extraction. Depending on the properties of target compounds, the proper extraction and separation techniques should be selected to overcome the variances of source material. In some cases, the proper screening technique is also important for finding out the hypopigmentation compounds or extracts. In many cases, the activities of anti-melanogenic compounds are frequently verified against the inhibitory activity of mushroom TYR. However, this is not completely suitable, as there are inherent differences between mushroom and human TYR [103,104]. If human TYR is not readily available, the cellular TYR from mouse melanocytes can be much more reliable than mushroom TYR for initial screening of anti-melanogenic agents [86]. The phylogenetic analysis of TYR from various organisms on the basis of amino acid sequence of central catalytic domains suggests high level of homologies among human, gorilla and mouse TYR (representing the same cluster), while mushroom TYR was considerably more heterologous and placed in a distant cluster [103]. A previous study reported no effects of ethanolic extract of S. polycystum and its hexane fraction on mushroom TYR, but results in significant inhibition of cellular TYR when being tested on B16F10 mouse melanoma cells [86]. Han et al. [54] also reported that hydrolyzed ginseng extract could not inhibit mushroom TYR, but markedly suppressed melanogenesis and TYR activity in B16F10 cells. Similar findings were also reported by other studies [105]. Therefore, instead of the mushroom TYR inhibition assay, the human or mouse TYR inhibition assay could be a useful tool for screening potential anti-melanogenic agents.
In addition to screening techniques for hypopigmentation compounds, the safety of potential hypopigmenting compounds is essential for the application of human cosmetics or cosmeceuticals. Some commercial skin-whitening compounds, such as hydroquinone, have been used to treat skin hyperpigmentation disorders for a long time and were finally banned for their unwanted side effects. Therefore, long-term pre-clinical and clinical trials are necessary to verify the activity in humans focusing on molecular events, efficiency and side effects.
8. Conclusions
Anti-melanogenic compounds from marine brown algae have high potential as therapeutic agents against skin hyperpigmentation disorders. They also have huge demand in cosmetic industries as skin-lightening agents due to the increasing concern for lighter skin complexion by Asian women [106]. In association with efficiency, the safety profiles of hypopigmentation compounds are very important for human use. Marine algae are considered as being diversified and safe sources of bioactive compounds. Therefore, the identification of novel hypopigmentation compounds from marine algae and uncovering their molecular mechanisms using in vitro and in vivo systems has great importance in meeting the future challenges of pharmaceutical and cosmetic industry.
Acknowledgments
The study was a part of the project “Development of functional food products with natural materials derived from marine resources” funded by the Ministry of Oceans and Fisheries, Republic of Korea.
Author Contributions
M.S.A. screened and organized related articles, and wrote the manuscript. H.-R.K. corrected the manuscript. J.C. and M.-S.L. contributed to the analysis of data from the references.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Afaq, F.; Adhami, V.M.; Mukhtar, H. Photochemoprevention of ultraviolet B signaling and photocarcinogenesis. Mutat. Res. 2005, 571, 153–173. [Google Scholar] [CrossRef] [PubMed]
- Goihman-Yahr, M. Skin aging and photoaging: An outlook. Clin. Dermatol. 1996, 14, 153–160. [Google Scholar] [CrossRef]
- Han, E.; Chang, B.; Kim, D.; Cho, H.; Kim, S. Melanogenesis inhibitory effect of aerial part of Pueraria thunbergiana in vitro and in vivo. Arch. Dermatol. Res. 2015, 307, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Yoon, S.J.; Park, Y.J.; Lee, H.B. Inhibitory effect of ephedrannins A and B from roots of Ephedra sinica STAPF on melanogenesis. Biochim. Biophys. Acta 2015, 1850, 1389–1396. [Google Scholar] [CrossRef] [PubMed]
- Lehraiki, A.; Abbe, P.; Cerezo, M.; Rouaud, F.; Regazzetti, C.; Chignon-Sicard, B.; Passeron, T.; Bertolotto, C.; Ballotti, R.; Rocchi, S. Inhibition of melanogenesis by the antidiabetic metformin. J. Investig. Dermatol. 2014, 134, 2589–2597. [Google Scholar] [CrossRef] [PubMed]
- Briganti, S.; Camera, E.; Picardo, M. Chemical and instrumental approaches to treat hyperpigmentation. Pigment Cell Res. 2003, 16, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.H.; Ko, S.C.; Kim, D.; Jeon, Y.J. Screening of marine algae for potential tyrosinase inhibitor: Those inhibitors reduced tyrosinase activity and melanin synthesis in zebrafish. J. Dermatol. 2011, 38, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Curto, E.V.; Kwong, C.; Hermersdorfer, H.; Glatt, H.; Santis, C.; Virador, V.; Hearing, V.J., Jr.; Dooley, T.P. Inhibitors of mammalian melanocyte tyrosinase: In vitro comparisons of alkyl esters of gentisic acid with other putative inhibitors. Biochem. Pharmacol. 1999, 57, 663–672. [Google Scholar] [CrossRef]
- Gao, X.-H.; Zhang, L.; Wei, H.; Chen, H.-D. Efficacy and safety of innovative cosmeceuticals. Clin. Dermatol. 2008, 26, 367–374. [Google Scholar] [CrossRef] [PubMed]
- DeCaprio, A.P. The toxicology of hydroquinone—Relevance to occupational and environmental exposure. Crit. Rev. Toxicol. 1999, 29, 283–330. [Google Scholar] [CrossRef] [PubMed]
- O‘Donoghue, J.L. Hydroquinone and its analogues in dermatology—A risk-benefit viewpoint. J. Cosmet. Dermatol. 2006, 5, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Serra-Baldrich, E.; Tribo, M.J.; Camarasa, J.G. Allergic contact dermatitis from kojic acid. Contact Dermat. 1998, 39, 86–87. [Google Scholar] [CrossRef]
- Takizawa, T.; Imai, T.; Onose, J.; Ueda, M.; Tamura, T.; Mitsumori, K.; Izumi, K.; Hirose, M. Enhancement of hepatocarcinogenesis by kojic acid in rat two-stage models after initiation with N-bis(2-hydroxypropyl)nitrosamine or N-diethylnitrosamine. Toxicol. Sci. 2004, 81, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Pomponi, S.A. The bioprocess–technological potential of the sea. J. Biotechnol. 1999, 70, 5–13. [Google Scholar] [CrossRef]
- Wijesekara, I.; Pangestuti, R.; Kim, S.-K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar] [CrossRef]
- Heo, S.J.; Park, E.J.; Lee, K.W.; Jeon, Y.J. Antioxidant activities of enzymatic extracts from brown seaweeds. Bioresour. Technol. 2005, 96, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-K.; Ravichandran, Y.D.; Khan, S.B.; Kim, Y.T. Prospective of the cosmeceuticals derived from marine organisms. Biotechnol. Bioprocess Eng. 2008, 13, 511–523. [Google Scholar] [CrossRef]
- Heo, S.J.; Ko, S.C.; Cha, S.H.; Kang, D.H.; Park, H.S.; Choi, Y.U.; Kim, D.; Jung, W.K.; Jeon, Y.J. Effect of phlorotannins isolated from Ecklonia cava on melanogenesis and their protective effect against photo-oxidative stress induced by UV-B radiation. Toxicol. In Vitro 2009, 23, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Anunciato, T.P.; da Rocha Filho, P.A. Carotenoids and polyphenols in nutricosmetics, nutraceuticals, and cosmeceuticals. J. Cosmet. Dermatol. 2012, 11, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.A.; Kim, E.-A.; Son, K.-T.; Jeon, Y.-J. Bioactive properties and potentials cosmeceutical applications of phlorotannins isolated from brown seaweeds: A review. J. Photochem. Photobiol. B Biol. 2016, 162, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Joung, E.-J.; Gwon, W.-G.; Shin, T.; Jung, B.-M.; Choi, J.; Kim, H.-R. Anti-inflammatory action of the ethanolic extract from Sargassum serratifolium on lipopolysaccharide-stimulated mouse peritoneal macrophages and identification of active components. J. Appl. Phycol. 2017, 29, 563–573. [Google Scholar] [CrossRef]
- Seo, Y.; Park, K.E.; Kim, Y.A.; Lee, H.J.; Yoo, J.S.; Ahn, J.W.; Lee, B.J. Isolation of tetraprenyltoluquinols from the brown alga Sargassum thunbergii. Chem. Pharm. Bull. 2006, 54, 1730–1733. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A.; Ahn, B.N.; Kong, C.S.; Kim, S.K. Protective effect of chromene isolated from Sargassum horneri against UV-A-induced damage in skin dermal fibroblasts. Exp. Dermatol. 2012, 21, 630–631. [Google Scholar] [CrossRef] [PubMed]
- Gwon, W.G.; Lee, B.; Joung, E.J.; Choi, M.W.; Yoon, N.; Shin, T.; Oh, C.W.; Kim, H.R. Sargaquinoic Acid Inhibits TNF-alpha-Induced NF-kappaB Signaling, Thereby Contributing to Decreased Monocyte Adhesion to Human Umbilical Vein Endothelial Cells (HUVECs). J. Agric. Food Chem. 2015, 63, 9053–9061. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.N.; Choi, H.Y.; Lee, W.; Park, G.M.; Shin, W.S.; Kim, Y.K. Sargaquinoic acid and sargahydroquinoic acid from Sargassum yezoense stimulate adipocyte differentiation through PPARalpha/gamma activation in 3T3-L1 cells. FEBS Lett. 2008, 582, 3465–3472. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.N.; Lee, W.; Bae, G.U.; Kim, Y.K. Anti-diabetic and hypolipidemic effects of Sargassum yezoense in db/db mice. Biochem. Biophys. Res. Commun. 2012, 424, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Heinrich, M.; Myers, S.; Dworjanyn, S.A. Towards a better understanding of medicinal uses of the brown seaweed Sargassum in Traditional Chinese Medicine: A phytochemical and pharmacological review. J. Ethnopharmacol. 2012, 142, 591–619. [Google Scholar] [CrossRef] [PubMed]
- Liu-Smith, F.; Meyskens, F.L. Molecular mechanisms of flavonoids in melanin synthesis and the potential for the prevention and treatment of melanoma. Mol. Nutr. Food Res. 2016, 60, 1264–1274. [Google Scholar] [CrossRef] [PubMed]
- Khaled, M.; Larribere, L.; Bille, K.; Aberdam, E.; Ortonne, J.P.; Ballotti, R.; Bertolotto, C. Glycogen synthase kinase 3beta is activated by cAMP and plays an active role in the regulation of melanogenesis. J. Biol. Chem. 2002, 277, 33690–33697. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.S.; Joung, E.-J.; Choi, J.; Kim, H.-R. Ethanolic extract from Sargassum serratifolium attenuates hyperpigmentation through CREB/ERK signaling pathways in α-MSH-stimulated B16F10 melanoma cells. J. Appl. Phycol. 2017, 29, 2089–2096. [Google Scholar] [CrossRef]
- Garcia-Borron, J.C.; Abdel-Malek, Z.; Jimenez-Cervantes, C. MC1R, the cAMP pathway, and the response to solar UV: Extending the horizon beyond pigmentation. Pigment Cell Melanoma Res. 2014, 27, 699–720. [Google Scholar] [CrossRef] [PubMed]
- Widlund, H.R.; Fisher, D.E. Microphthalamia-associated transcription factor: A critical regulator of pigment cell development and survival. Oncogene 2003, 22, 3035–3041. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-J.; Dai, R.-Y.; Leu, Y.-L.; Tsai, T.-Y. Effects of the melanogenic inhibitor, uracil, derived from Lactobacillus plantarum TWK10-fermented soy milk on anti-melanogenesis in B16F0 mouse melanoma cells. J. Funct. Foods 2015, 17, 314–327. [Google Scholar] [CrossRef]
- Napolitano, A.; Micillo, R.; Monfrecola, G. Melanin pigmentation control by 1,3-thiazolidines: Does NO scavenging play a critical role? Exp. Dermatol. 2016, 25, 596–597. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Cheng, C.Y.; Saldanha, S.A.; Taylor, S.S. PKA-I holoenzyme structure reveals a mechanism for cAMP-dependent activation. Cell 2007, 130, 1032–1043. [Google Scholar] [CrossRef] [PubMed]
- Roh, E.; Yun, C.Y.; Young Yun, J.; Park, D.; Doo Kim, N.; Yeon Hwang, B.; Jung, S.H.; Park, S.K.; Kim, Y.B.; Han, S.B.; et al. cAMP-binding site of PKA as a molecular target of bisabolangelone against melanocyte-specific hyperpigmented disorder. J. Investig. Dermatol. 2013, 133, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Karelina, K.; Obrietan, K. CREB: A multifaceted regulator of neuronal plasticity and protection. J. Neurochem. 2011, 116, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.S.; Gu, G.E.; Jo, A.R.; Bang, J.S.; Yun, H.Y.; Baek, K.J.; Kwon, N.S.; Park, K.C.; Kim, D.S. Baicalin-induced Akt activation decreases melanogenesis through downregulation of microphthalmia-associated transcription factor and tyrosinase. Eur. J. Pharmacol. 2015, 761, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.G.; Choi, E.J.; Choi, Y.; Hwang, J.K. 5,7-dimethoxyflavone induces melanogenesis in B16F10 melanoma cells through cAMP-dependent signalling. Exp. Dermatol. 2011, 20, 445–447. [Google Scholar] [CrossRef] [PubMed]
- Bertolotto, C.; Bille, K.; Ortonne, J.P.; Ballotti, R. Regulation of tyrosinase gene expression by cAMP in B16 melanoma cells involves two CATGTG motifs surrounding the TATA box: Implication of the microphthalmia gene product. J. Cell Biol. 1996, 134, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Busca, R.; Ballotti, R. Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigment Cell Res. 2000, 13, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Xu, W.; Liang, J.W.; Wang, C.S.; Kang, Y. Effect of fucoidan on b16 murine melanoma cell melanin formation and apoptosis. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Hong, S.D.; Roh, E.; Jung, S.-H.; Cho, W.-J.; Hong Park, S.; Yoon, D.Y.; Ko, S.M.; Hwang, B.Y.; Hong, J.T.; et al. cAMP-dependent activation of protein kinase A as a therapeutic target of skin hyperpigmentation by diphenylmethylene hydrazinecarbothioamide. Br. J. Pharmacol. 2015, 172, 3434–3445. [Google Scholar] [CrossRef] [PubMed]
- Englaro, W.; Bertolotto, C.; Busca, R.; Brunet, A.; Pages, G.; Ortonne, J.P.; Ballotti, R. Inhibition of the mitogen-activated protein kinase pathway triggers B16 melanoma cell differentiation. J. Biol. Chem. 1998, 273, 9966–9970. [Google Scholar] [CrossRef] [PubMed]
- Hemesath, T.J.; Price, E.R.; Takemoto, C.; Badalian, T.; Fisher, D.E. MAP kinase links the transcription factor Microphthalmia to c-Kit signalling in melanocytes. Nature 1998, 391, 298–301. [Google Scholar] [PubMed]
- Wu, L.C.; Lin, Y.Y.; Yang, S.Y.; Weng, Y.T.; Tsai, Y.T. Antimelanogenic effect of c-phycocyanin through modulation of tyrosinase expression by upregulation of ERK and downregulation of p38 MAPK signaling pathways. J. Biomed. Sci. 2011, 18, 74. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Hemesath, T.J.; Takemoto, C.M.; Horstmann, M.A.; Wells, A.G.; Price, E.R.; Fisher, D.Z.; Fisher, D.E. c-Kit triggers dual phosphorylations, which couple activation and degradation of the essential melanocyte factor Mi. Genes Dev. 2000, 14, 301–312. [Google Scholar] [PubMed]
- Lee, H.J.; Lee, W.J.; Chang, S.E.; Lee, G.Y. Hesperidin, A Popular Antioxidant Inhibits Melanogenesis via Erk1/2 Mediated MITF Degradation. Int. J. Mol. Sci. 2015, 16, 18384–18395. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Park, S.H.; Kwon, S.B.; Kwon, N.S.; Park, K.C. Sphingosylphosphorylcholine inhibits melanin synthesis via pertussis toxin-sensitive MITF degradation. J. Pharm. Pharmacol. 2010, 62, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Englaro, W.; Rezzonico, R.; Durand-Clement, M.; Lallemand, D.; Ortonne, J.P.; Ballotti, R. Mitogen-activated protein kinase pathway and AP-1 are activated during cAMP-induced melanogenesis in B-16 melanoma cells. J. Biol. Chem. 1995, 270, 24315–24320. [Google Scholar] [CrossRef] [PubMed]
- Hirata, N.; Naruto, S.; Ohguchi, K.; Akao, Y.; Nozawa, Y.; Iinuma, M.; Matsuda, H. Mechanism of the melanogenesis stimulation activity of (–)-cubebin in murine B16 melanoma cells. Bioorg. Med. Chem. 2007, 15, 4897–4902. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, J.S.; Woo, J.T.; Lee, I.S.; Cha, B.Y. Hyperpigmentation mechanism of methyl 3,5-di-caffeoylquinate through activation of p38 and MITF induction of tyrosinase. Acta Biochim. Biophys. Sin. 2015, 47, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Bu, J.; Ma, P.C.; Chen, Z.Q.; Zhou, W.Q.; Fu, Y.J.; Li, L.J.; Li, C.R. Inhibition of MITF and tyrosinase by paeonol-stimulated JNK/SAPK to reduction of phosphorylated CREB. Am. J. Chin. Med. 2008, 36, 245–263. [Google Scholar] [CrossRef] [PubMed]
- Han, J.S.; Sung, J.H.; Lee, S.K. Antimelanogenesis Activity of Hydrolyzed Ginseng Extract (GINST) via Inhibition of JNK Mitogen-activated Protein Kinase in B16F10 Cells. J. Food Sci. 2016, 81, H2085–H2092. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Yoon, H.D.; Kim, J.I.; Choi, J.S.; Byun, D.S.; Kim, H.R. Dioxinodehydroeckol inhibits melanin synthesis through PI3K/Akt signalling pathway in alpha-melanocyte-stimulating hormone-treated B16F10 cells. Exp. Dermatol. 2012, 21, 471–473. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.X.; Lin, M.; Lu, S.S.; Qi, X.Y.; Zhang, R.X.; Zhang, Y.Y. Curcumin inhibits melanogenesis in human melanocytes. Phytother. Res. 2012, 26, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Alessi, D.R.; Andjelkovic, M.; Caudwell, B.; Cron, P.; Morrice, N.; Cohen, P.; Hemmings, B.A. Mechanism of activation of protein kinase B by insulin and IGF-1. EMBO J. 1996, 15, 6541–6551. [Google Scholar] [PubMed]
- Cross, D.A.E.; Alessi, D.R.; Cohen, P.; Andjelkovich, M.; Hemmings, B.A. Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature 1995, 378, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Busca, R.; Bertolotto, C.; Ortonne, J.P.; Ballotti, R. Inhibition of the phosphatidylinositol 3-kinase/p70(S6)-kinase pathway induces B16 melanoma cell differentiation. J. Biol. Chem. 1996, 271, 31824–31830. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.Y.; Yin, W.H.; Wang, M.R.; Dang, Y.Y.; Ye, X.Y. Andrographolide suppresses melanin synthesis through Akt/GSK3beta/beta-catenin signal pathway. J. Dermatol. Sci. 2015, 79, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.; Ryu, M.; Jeong, Y.; Chung, Y.H.; Kim, D.E.; Cho, H.S.; Kang, S.; Han, J.S.; Chang, M.Y.; Lee, C.K.; et al. Cardamonin suppresses melanogenesis by inhibition of Wnt/beta-catenin signaling. Biochem. Biophys. Res. Commun. 2009, 390, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Dunn, K.J.; Williams, B.O.; Li, Y.; Pavan, W.J. Neural crest-directed gene transfer demonstrates Wnt1 role in melanocyte expansion and differentiation during mouse development. Proc. Natl. Acad. Sci. USA 2000, 97, 10050–10055. [Google Scholar] [CrossRef] [PubMed]
- Bellei, B.; Pitisci, A.; Catricala, C.; Larue, L.; Picardo, M. Wnt/beta-catenin signaling is stimulated by alpha-melanocyte-stimulating hormone in melanoma and melanocyte cells: Implication in cell differentiation. Pigment Cell Melanoma Res. 2011, 24, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.; Ganesan, A.K. The pleiotropic roles of autophagy regulators in melanogenesis. Pigment Cell Melanoma Res. 2011, 24, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S.; Jo, Y.K.; Park, S.J.; Chang, H.; Shin, J.H.; Choi, E.S.; Kim, J.B.; Seok, S.H.; Kim, J.S.; Oh, J.S.; et al. ARP101 inhibits alpha-MSH-stimulated melanogenesis by regulation of autophagy in melanocytes. FEBS Lett. 2013, 587, 3955–3960. [Google Scholar] [CrossRef] [PubMed]
- Van den Boorn, J.G.; Picavet, D.I.; van Swieten, P.F.; van Veen, H.A.; Konijnenberg, D.; van Veelen, P.A.; van Capel, T.; Jong, E.C.; Reits, E.A.; Drijfhout, J.W.; et al. Skin-depigmenting agent monobenzone induces potent T-cell autoimmunity toward pigmented cells by tyrosinase haptenation and melanosome autophagy. J. Investig. Dermatol. 2011, 131, 1240–1251. [Google Scholar] [CrossRef] [PubMed]
- Romero-Graillet, C.; Aberdam, E.; Clement, M.; Ortonne, J.P.; Ballotti, R. Nitric oxide produced by ultraviolet-irradiated keratinocytes stimulates melanogenesis. J. Clin. Investig. 1997, 99, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wang, H.; Cao, J.; Ren, J.; Fan, R.; He, X.; Smith, G.W.; Dong, C. Nitric oxide enhances melanogenesis of alpaca skin melanocytes in vitro by activating the MITF phosphorylation. Mol. Cell. Biochem. 2011, 352, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Cao, J.; Wang, H.; Zhang, J.; Zhu, Z.; Bai, R.; Hao, H.; He, X.; Fan, R.; Dong, C. Nitric oxide enhances the sensitivity of alpaca melanocytes to respond to α-melanocyte-stimulating hormone by up-regulating melanocortin-1 receptor. Biochem. Biophys. Res. Commun. 2010, 396, 849–853. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Esparza, M.; Ferrer, C.; Castells, M.T.; Garcia-Borron, J.C.; Zuasti, A. Transforming growth factor beta1 mediates hypopigmentation of B16 mouse melanoma cells by inhibition of melanin formation and melanosome maturation. Int. J. Biochem. Cell Biol. 2001, 33, 971–983. [Google Scholar] [CrossRef]
- Minwalla, L.; Zhao, Y.; Cornelius, J.; Babcock, G.F.; Wickett, R.R.; Le Poole, I.C.; Boissy, R.E. Inhibition of melanosome transfer from melanocytes to keratinocytes by lectins and neoglycoproteins in an in vitro model system. Pigment Cell Res. 2001, 14, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.N.; Yang, H.M.; Kang, S.M.; Ahn, G.N.; Roh, S.W.; Lee, W.; Kim, D.K.; Jeon, Y.J. Whitening Effect of Octaphlorethol A Isolated from Ishige foliacea in an In Vivo Zebrafish Model. J. Microbiol. Biotechnol. 2015, 25, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Joe, M.J.; Kim, S.N.; Choi, H.Y.; Shin, W.S.; Park, G.M.; Kang, D.W.; Kim, Y.K. The inhibitory effects of eckol and dieckol from Ecklonia stolonifera on the expression of matrix metalloproteinase-1 in human dermal fibroblasts. Biol. Pharm. Bull. 2006, 29, 1735–1739. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Shin, T.; Utsuki, T.; Choi, J.S.; Byun, D.S.; Kim, H.R. Isolation and identification of phlorotannins from Ecklonia stolonifera with antioxidant and hepatoprotective properties in tacrine-treated HepG2 cells. J. Agric. Food Chem. 2012, 60, 5340–5349. [Google Scholar] [CrossRef] [PubMed]
- Jun, Y.J.; Lee, M.; Shin, T.; Yoon, N.; Kim, J.H.; Kim, H.R. Eckol enhances heme oxygenase-1 expression through activation of Nrf2/JNK pathway in HepG2 cells. Molecules 2014, 19, 15638–15652. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Lee, M.S.; Choi, J.W.; Utsuki, T.; Kim, J.I.; Jang, B.C.; Kim, H.R. Phlorofucofuroeckol A suppresses expression of inducible nitric oxide synthase, cyclooxygenase-2, and pro-inflammatory cytokines via inhibition of nuclear factor-kappaB, c-Jun NH2-terminal kinases, and Akt in microglial cells. Inflammation 2013, 36, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.-Y.; Choi, J.-S.; Byun, D.-S. Inhibitory effects of phloroglucinol derivatives isolated from Ecklonia stolonifera on FcεRI expression. Bioorg. Med. Chem. 2009, 17, 4734–4739. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.S.; Kim, H.R.; Byun, D.S.; Son, B.W.; Nam, T.J.; Choi, J.S. Tyrosinase inhibitors isolated from the edible brown alga Ecklonia stolonifera. Arch. Pharm. Res. 2004, 27, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.-J.; Ko, S.-C.; Kang, S.-M.; Cha, S.-H.; Lee, S.-H.; Kang, D.-H.; Jung, W.-K.; Affan, A.; Oh, C.; Jeon, Y.-J. Inhibitory effect of diphlorethohydroxycarmalol on melanogenesis and its protective effect against UV-B radiation-induced cell damage. Food Chem. Toxicol. 2010, 48, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.N.; Yang, H.M.; Kang, S.M.; Kim, D.; Ahn, G.; Jeon, Y.J. Octaphlorethol A isolated from Ishige foliacea inhibits alpha-MSH-stimulated induced melanogenesis via ERK pathway in B16F10 melanoma cells. Food Chem. Toxicol. 2013, 59, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.-S.; Park, H.-Y.; Nam, K.-H. Whitening effects of 4-hydroxyphenethyl alcohol isolated from water boiled with Hizikia fusiformis. Food Sci. Biotechnol. 2014, 23, 555–560. [Google Scholar] [CrossRef]
- Lee, S.-H.; Kang, S.-M.; Sok, C.H.; Hong, J.T.; Oh, J.-Y.; Jeon, Y.-J. Cellular activities and docking studies of eckol isolated from Ecklonia cava (Laminariales, Phaeophyceae) as potential tyrosinase inhibitor. Algae 2015, 30, 163–170. [Google Scholar] [CrossRef]
- Yoon, N.Y.; Eom, T.K.; Kim, M.M.; Kim, S.K. Inhibitory effect of phlorotannins isolated from Ecklonia cava on mushroom tyrosinase activity and melanin formation in mouse B16F10 melanoma cells. J. Agric. Food Chem. 2009, 57, 4124–4129. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.S.; Balcos, M.C.; Yun, H.Y.; Baek, K.J.; Kwon, N.S.; Kim, M.K.; Kim, D.S. ERK Activation by Fucoidan Leads to Inhibition of Melanogenesis in Mel-Ab Cells. Korean J. Physiol. Pharmacol. 2015, 19, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, H.; Tanaka, J.; Shan, S.J.; Maoka, T. Anti-pigmentary activity of fucoxanthin and its influence on skin mRNA expression of melanogenic molecules. J. Pharm. Pharmacol. 2010, 62, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.Y.; Kim, K.H.; Cheah, S.H. Inhibitory effects of Sargassum polycystum on tyrosinase activity and melanin formation in B16F10 murine melanoma cells. J. Ethnopharmacol. 2011, 137, 1183–1188. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, M.-S.; Lee, B.; Gwon, W.-G.; Joung, E.-J.; Yoon, N.-Y.; Kim, H.-R. Anti-inflammatory effects of sargachromenol-rich ethanolic extract of Myagropsis myagroides on lipopolysaccharide-stimulated BV-2 cells. BMC Complement. Altern. Med. 2014, 14, 231. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.; Jung, Y.; Kim, M.C.; Kwon, H.C.; Kang, K.S.; Kim, Y.K.; Kim, S.N. Sargahydroquinoic acid inhibits TNFalpha-induced AP-1 and NF-kappaB signaling in HaCaT cells through PPARalpha activation. Biochem. Biophys. Res. Commun. 2014, 450, 1553–1559. [Google Scholar] [CrossRef] [PubMed]
- Joung, E.J.; Lee, B.; Gwon, W.G.; Shin, T.; Jung, B.M.; Yoon, N.Y.; Choi, J.S.; Oh, C.W.; Kim, H.R. Sargaquinoic acid attenuates inflammatory responses by regulating NF-kappaB and Nrf2 pathways in lipopolysaccharide-stimulated RAW 264.7 cells. Int. Immunopharmacol. 2015, 29, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Joung, E.J.; Kwon, M.S.; Lee, B.; Utsuki, T.; Oh, C.W.; Kim, H.R. Anti-Inflammatory Effect of Ethanolic Extract of Sargassum serratifolium in Lipopolysaccharide-Stimulated BV2 Microglial Cells. J. Med. Food 2016, 19, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Gwon, W.G.; Joung, E.J.; Kwon, M.S.; Lim, S.J.; Utsuki, T.; Kim, H.R. Sargachromenol protects against vascular inflammation by preventing TNF-alpha-induced monocyte adhesion to primary endothelial cells via inhibition of NF-kappaB activation. Int. Immunopharmacol. 2017, 42, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Lim, S.-J.; Lee, B.; Shin, T.; Kim, H.-R. Ethanolic extract of Sargassum serratifolium inhibits adipogenesis in 3T3-L1 preadipocytes by cell cycle arrest. J. Appl. Phycol. 2017, 1–10. [Google Scholar] [CrossRef]
- De la Mare, J.A.; Lawson, J.C.; Chiwakata, M.T.; Beukes, D.R.; Edkins, A.L.; Blatch, G.L. Quinones and halogenated monoterpenes of algal origin show anti-proliferative effects against breast cancer cells in vitro. Investig. New Drugs 2012, 30, 2187–2200. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.W.; Park, M.S.; Kim, N.H.; Lee, J.H.; Oh, C.W.; Kim, H.R.; Kim, G.D. Hexane extract from Sargassum serratifolium inhibits the cell proliferation and metastatic ability of human glioblastoma U87MG cells. Oncol. Rep. 2015, 34, 2602–2608. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.K.; Kamei, Y. Sargaquinoic acid supports the survival of neuronal PC12D cells in a nerve growth factor-independent manner. Eur. J. Pharmacol. 2004, 488, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Fitton, J.H.; Dell’Acqua, G.; Gardiner, V.-A.; Karpiniec, S.S.; Stringer, D.N.; Davis, E. Topical Benefits of Two Fucoidan-Rich Extracts from Marine Macroalgae. Cosmetics 2015, 2, 66–81. [Google Scholar] [CrossRef]
- Wang, Z.J.; Si, Y.X.; Oh, S.; Yang, J.M.; Yin, S.J.; Park, Y.D.; Lee, J.; Qian, G.Y. The effect of fucoidan on tyrosinase: Computational molecular dynamics integrating inhibition kinetics. J. Biomol. Struct. Dyn. 2012, 30, 460–473. [Google Scholar] [CrossRef] [PubMed]
- Senni, K.; Gueniche, F.; Foucault-Bertaud, A.; Igondjo-Tchen, S.; Fioretti, F.; Colliec-Jouault, S.; Durand, P.; Guezennec, J.; Godeau, G.; Letourneur, D. Fucoidan a sulfated polysaccharide from brown algae is a potent modulator of connective tissue proteolysis. Arch. Biochem. Biophys. 2006, 445, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Semenov, A.V.; Mazurov, A.V.; Preobrazhenskaia, M.E.; Ushakova, N.A.; Mikhailov, V.I.; Berman, A.E.; Usov, A.I.; Nifant’ev, N.E.; Bovin, N.V. Sulfated polysaccharides as inhibitors of receptor activity of P-selectin and P-selectin-dependent inflammation. Vopr. Med. Khimii 1998, 44, 135–144. [Google Scholar] [PubMed]
- Wang, J.; Zhang, Q.; Zhang, Z.; Song, H.; Li, P. Potential antioxidant and anticoagulant capacity of low molecular weight fucoidan fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2010, 46, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Fitton, J.H. Therapies from Fucoidan; Multifunctional Marine Polymers. Mar. Drugs 2011, 9, 1731–1760. [Google Scholar] [CrossRef] [PubMed]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wichers, H.J.; Recourt, K.; Hendriks, M.; Ebbelaar, C.E.M.; Biancone, G.; Hoeberichts, F.A.; Mooibroek, H.; Soler-Rivas, C. Cloning, expression and characterisation of two tyrosinase cDNAs from Agaricus bisporus. Appl. Microbiol. Biotechnol. 2003, 61, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.Y.; Seok, J.K.; Suh, H.J.; Choi, Y.H.; Hong, S.S.; Kim, D.S.; Boo, Y.C. Antimelanogenic effects of luteolin 7-sulfate isolated from Phyllospadix iwatensis Makino. Br. J. Dermatol. 2016, 175, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.K.; Subedi, L.; Jeong, M.; Park, Y.U.; Kim, C.Y.; Kim, H.; Kim, S.Y. Gomisin N Inhibits Melanogenesis through Regulating the PI3K/Akt and MAPK/ERK Signaling Pathways in Melanocytes. Int. J. Mol. Sci. 2017, 18, 471. [Google Scholar] [CrossRef] [PubMed]
- Tengamnuay, P.; Pengrungruangwong, K.; Pheansri, I.; Likhitwitayawuid, K. Artocarpus lakoocha heartwood extract as a novel cosmetic ingredient: Evaluation of the in vitro anti-tyrosinase and in vivo skin whitening activities. Int. J. Cosmet. Sci. 2006, 28, 269–276. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic diagram representing major signaling pathways involved in the expression of melanogenic enzymes [29,30,31,32,33,34].

Figure 2.
Chemical structure of phlorotannins isolated from brown algae: (a) Eckol; (b) 2-phloroeckol; (c) 7-phloroeckol; (d) Diphlorethohydroxycarmalol; (e) Dieckol; (f) 6,6′-Bieckol; (g) Dioxinodehydroeckol; (h) Phloroglucinol; (i) Phlorofucofuroeckol A; (j) Phlorofucofuroeckol B; and (k) Octaphlorethol A.
Figure 2.
Chemical structure of phlorotannins isolated from brown algae: (a) Eckol; (b) 2-phloroeckol; (c) 7-phloroeckol; (d) Diphlorethohydroxycarmalol; (e) Dieckol; (f) 6,6′-Bieckol; (g) Dioxinodehydroeckol; (h) Phloroglucinol; (i) Phlorofucofuroeckol A; (j) Phlorofucofuroeckol B; and (k) Octaphlorethol A.
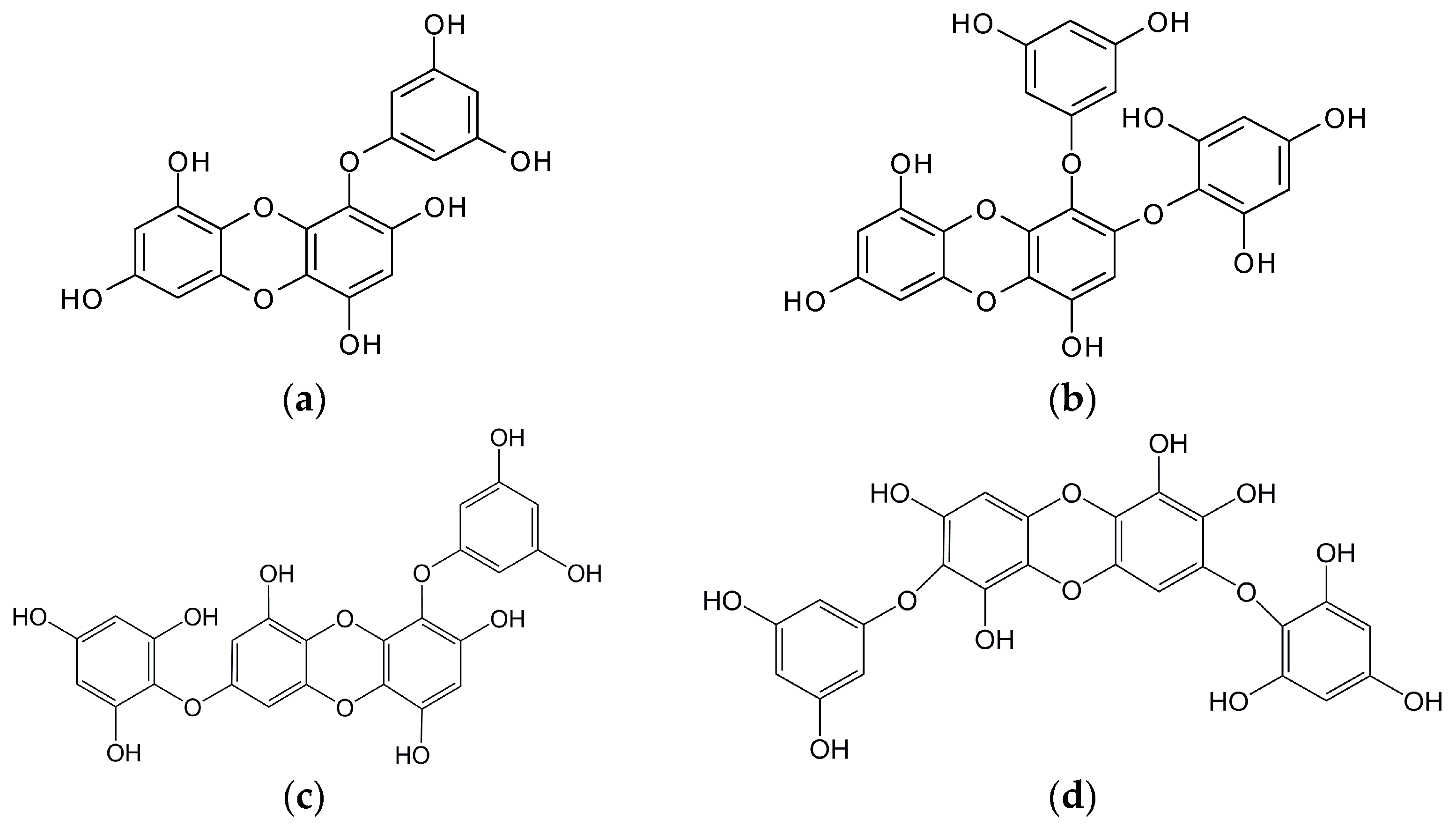
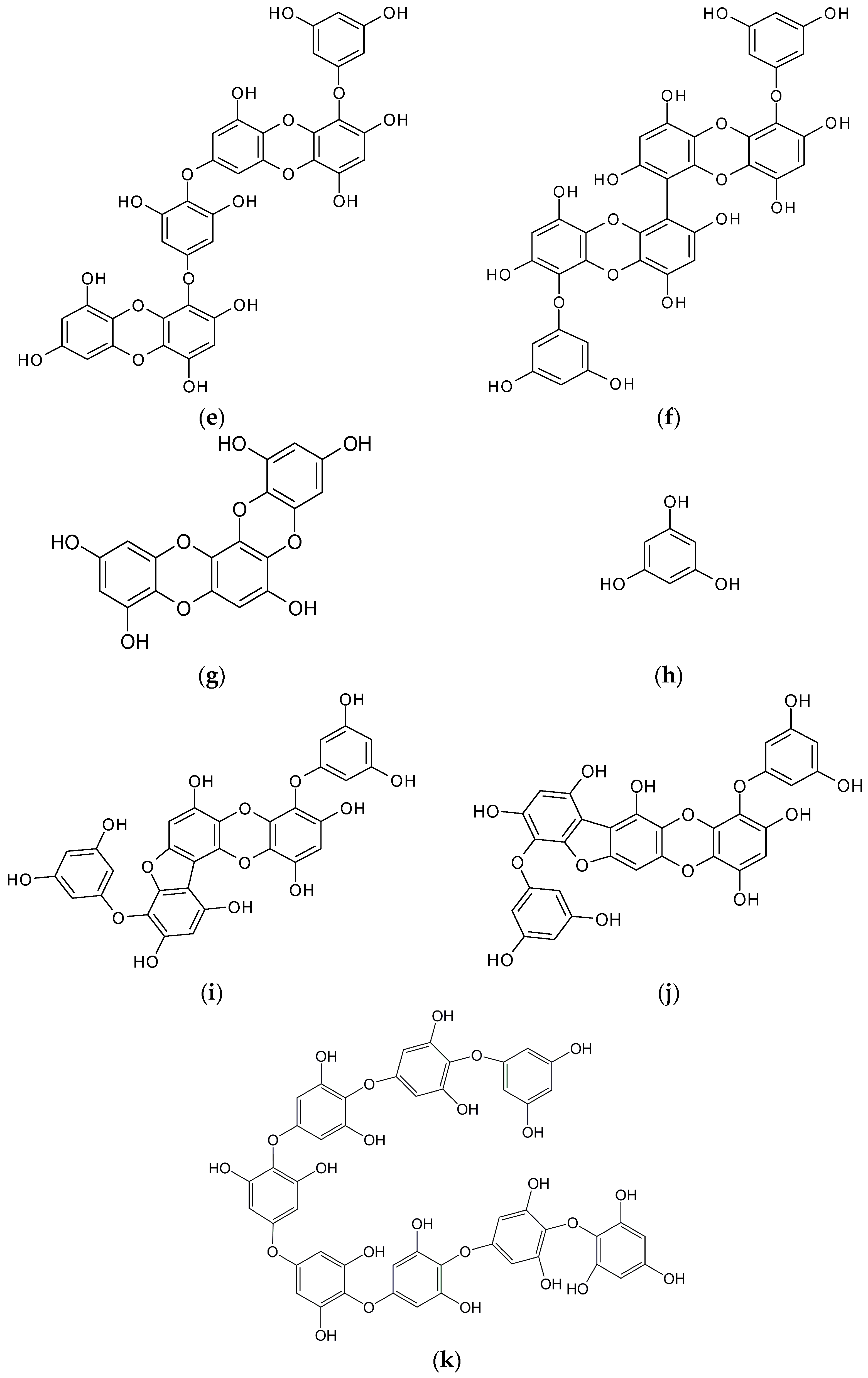
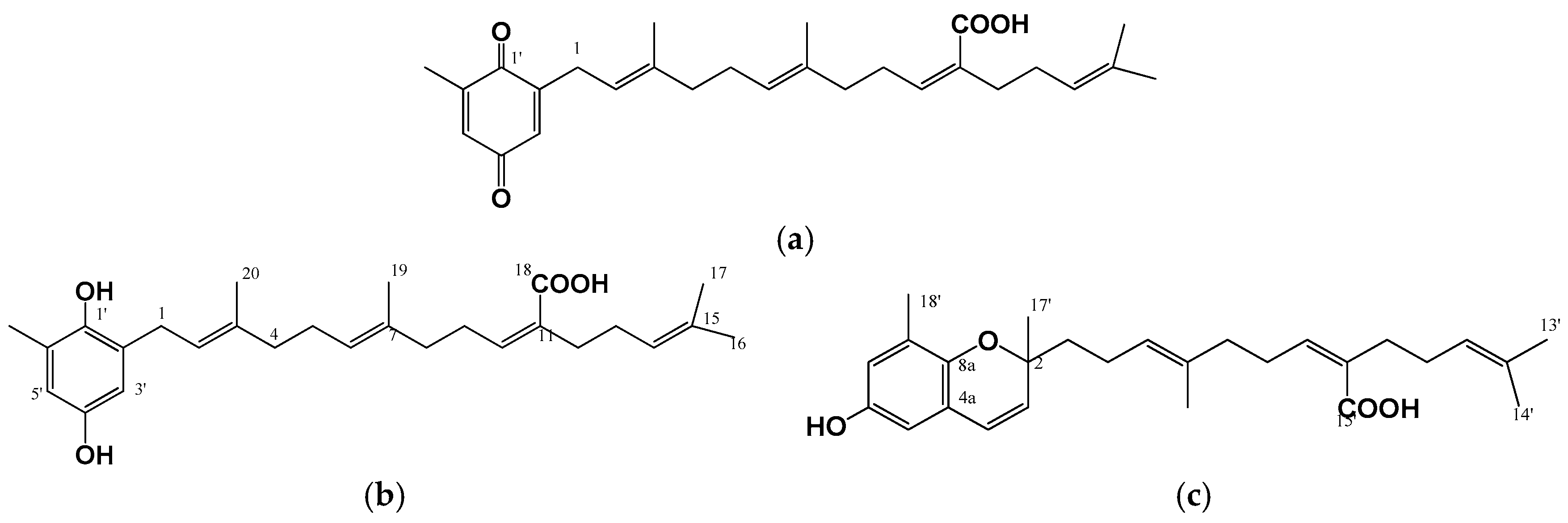
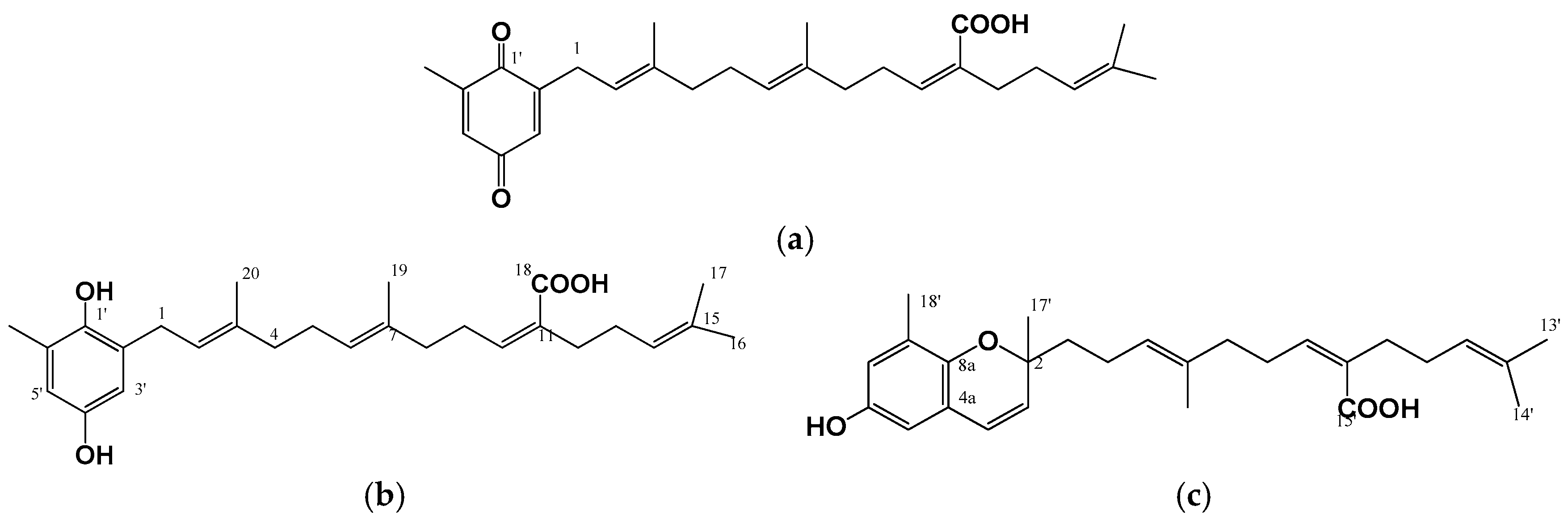
Figure 3.
Chemical structure of anti-melanogenic meroterpenoids isolated from the brown alga, Sargassum serratifolium [30]: (a) Sargaquinoic acid; (b) Sargahydroquinoic acid; and (c) Sargachromanol.
Figure 3.
Chemical structure of anti-melanogenic meroterpenoids isolated from the brown alga, Sargassum serratifolium [30]: (a) Sargaquinoic acid; (b) Sargahydroquinoic acid; and (c) Sargachromanol.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Overview of major hypopigmenting compounds from marine brown algae.
| Algae | Compounds/Extract | Type | Action Mechanism | Experimental System | Reference |
|---|---|---|---|---|---|
| Ecklonia stolonifera | Dioxinodehydroeckol | Phlorotannin | PI3K/Akt-mediated downregulation of MITF | B16F10 mouse melanoma cells | [55] |
| Ishige okamurae | Diphlorethohydroxycarmalol | Phlorotannin | Inhibition of mushroom TYR and melanin synthesis | B16F10 cells | [79] |
| E. cava | Eckol | Phlorotannin | Inhibition of cell free TYR (non-competitive) & cellular TYR, TRP1, and TRP2 | B16F10 cells | [82] |
| Dieckol | Inhibition of mushroom TYR & cellular melanin | B16F10 cells | [18] | ||
| Dioxinodehydroeckol | Mushroom TYR inhibition | Cell free | [83] | ||
| 7-phloroeckol | Inhibition of mushroom TYR (non-competitive) & cellular melanin | B16F10 cells | [83] | ||
| I. foliacea | Octaphlorethol A | Phlorotannin | ERK1/2-mediated downregulation of MITF, TYR, TRP1 & TRP2 in B16. Inhibition of in vivo TYR activity and melanin synthesis | B16F10 cells, Zebra fish embryo | [72,80] |
| Hizikia fusiformis | 4-hydroxyphenethyl alcohol | Non-flavonoid phenolic compound | Inhibition of mushroom TYR and melanin synthesis in B16. Reduction of pigmented spots in guinea-pig skin | B16F10 cells, Brown guinea-pig | [81] |
| Fucus vesiculosus | Fucoidan | Fucose-rich sulfated polysaccharide | ERK-mediated downregulation of MITF. | Mel-Ab cells | [84] |
| Inhibition of cellular TYR activity, melanin content & cell proliferation | B16 murine melanoma cells | [42] | |||
| Laminaria Japonica | Fucoxanthin | Carotenoid | Reduced TYR activity in B16 and melanin content in guinea-pigs & mice skin. Suppress PGE2, MSH, TRP1 & melanogenic stimulant receptors, NTR, EP1 & MC1R in vivo | B16 murine melanoma, UVB-induced mice, & guinea-pig | [85] |
| Sargassum serratifolium | Ethanolic extract containing sargaquinoic acid, sargahydroquinoic acid & sargachromenol | Meroterpenoid | cAMP and ERK1/2-mediated downregulation of MITF | B16F10 cells | [30] |
| S. polycystum | Ethanolic extract & its hexane fraction | NR | Inhibition of cellular TYR & melanin production | B16F10 cells | [86] |
EP1 = prostaglandin E receptor 1; MC1R = melanocortin 1 receptor; MSH = melanocyte stimulating hormone; NR = not reported; NTR = p75 neurotrophin receptor; PGE2 = prostaglandin E2; TYR = tyrosinase; TRP = tyrosinase-related protein; MITF = microphthalmia-associated transcription factor; ERK = extracellular signal-regulated kinase; cAMP = cyclic adenosine monophosphate.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Azam, M.S.; Choi, J.; Lee, M.-S.; Kim, H.-R. Hypopigmenting Effects of Brown Algae-Derived Phytochemicals: A Review on Molecular Mechanisms. Mar. Drugs 2017, 15, 297. https://doi.org/10.3390/md15100297
AMA Style
Azam MS, Choi J, Lee M-S, Kim H-R. Hypopigmenting Effects of Brown Algae-Derived Phytochemicals: A Review on Molecular Mechanisms. Marine Drugs. 2017; 15(10):297. https://doi.org/10.3390/md15100297
Chicago/Turabian StyleAzam, Mohammed Shariful, Jinkyung Choi, Min-Sup Lee, and Hyeung-Rak Kim. 2017. "Hypopigmenting Effects of Brown Algae-Derived Phytochemicals: A Review on Molecular Mechanisms" Marine Drugs 15, no. 10: 297. https://doi.org/10.3390/md15100297
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.