
Influence of Core Oligosaccharide of Lipopolysaccharide to Outer Membrane Behavior of Escherichia coli

Abstract
:1. Introduction
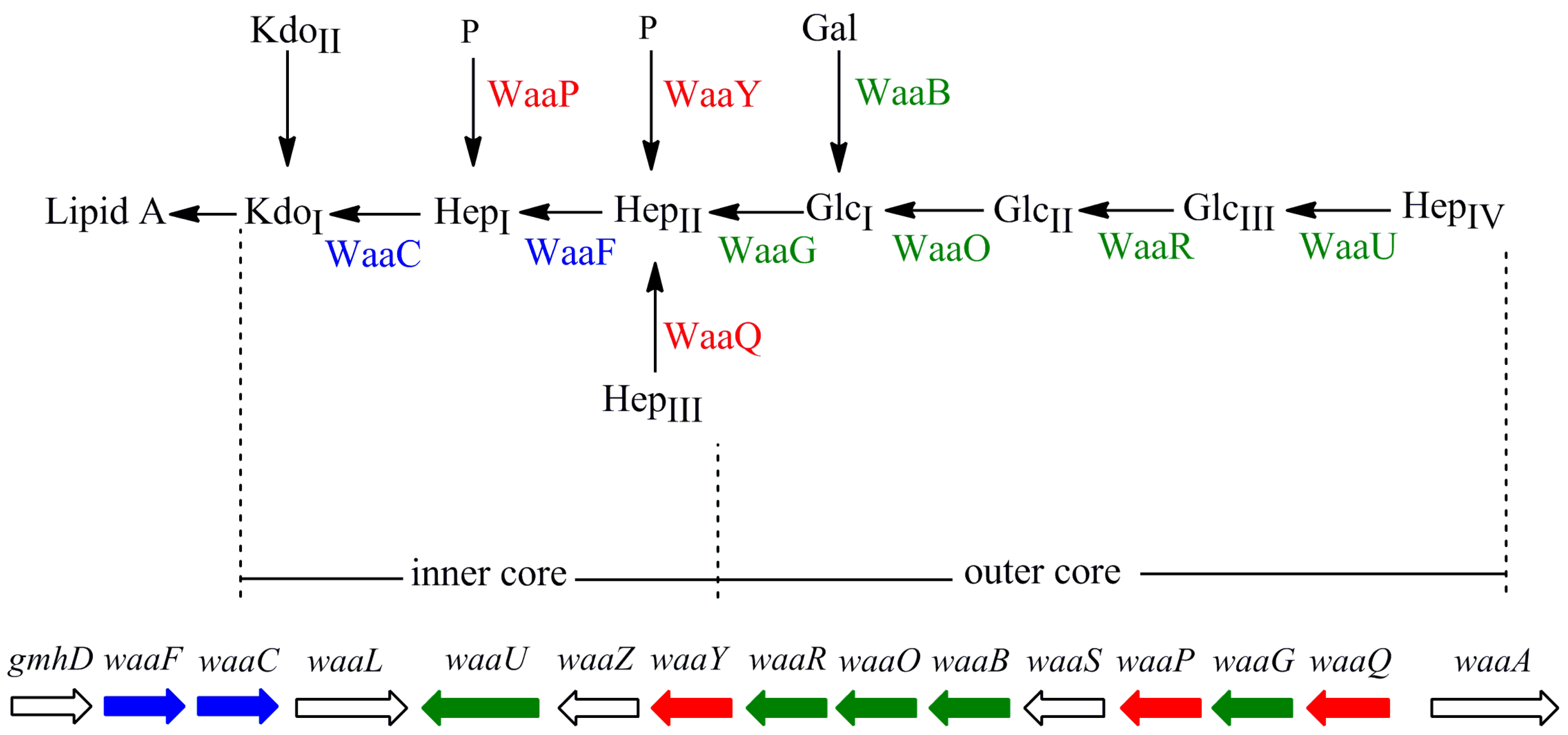
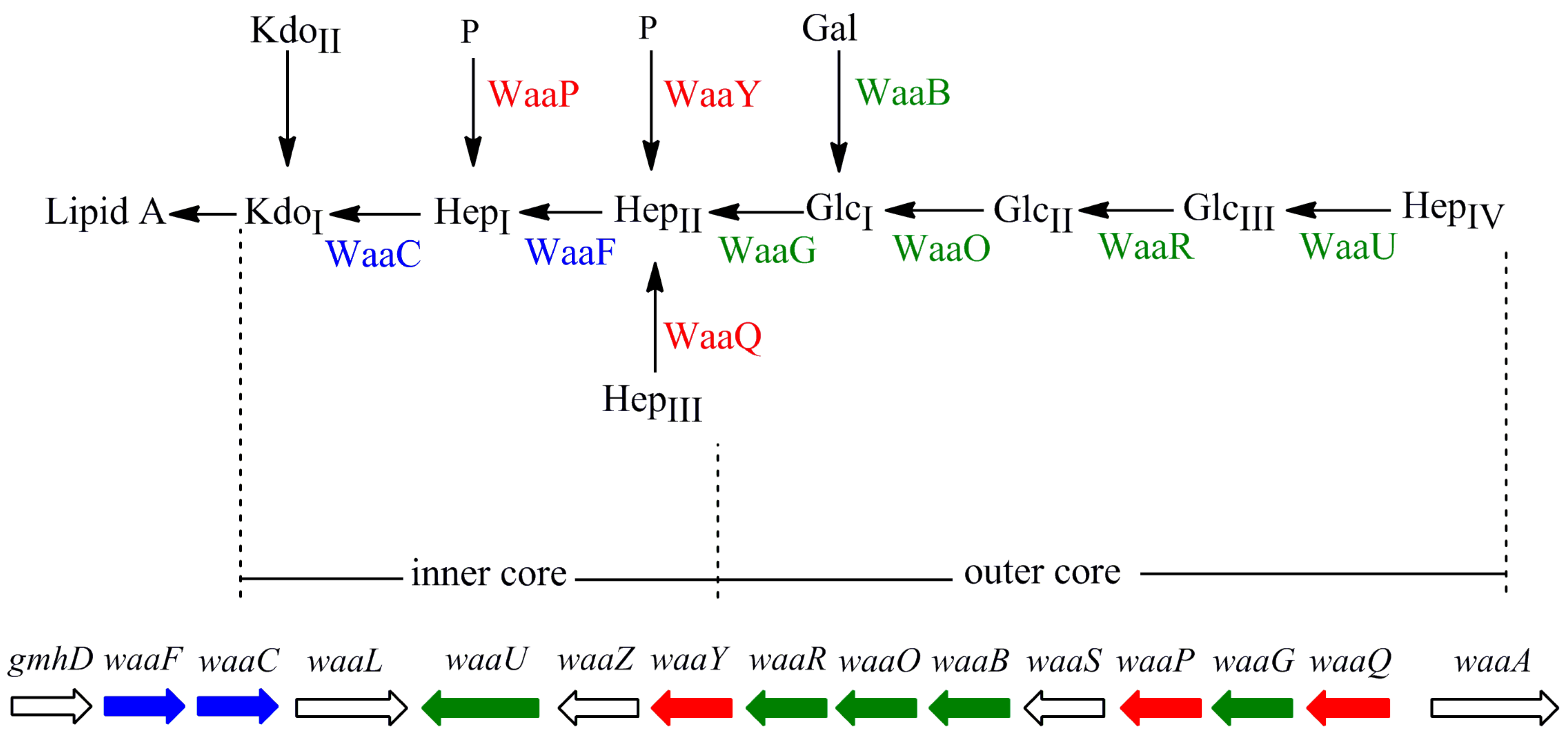
2. Results and Discussion
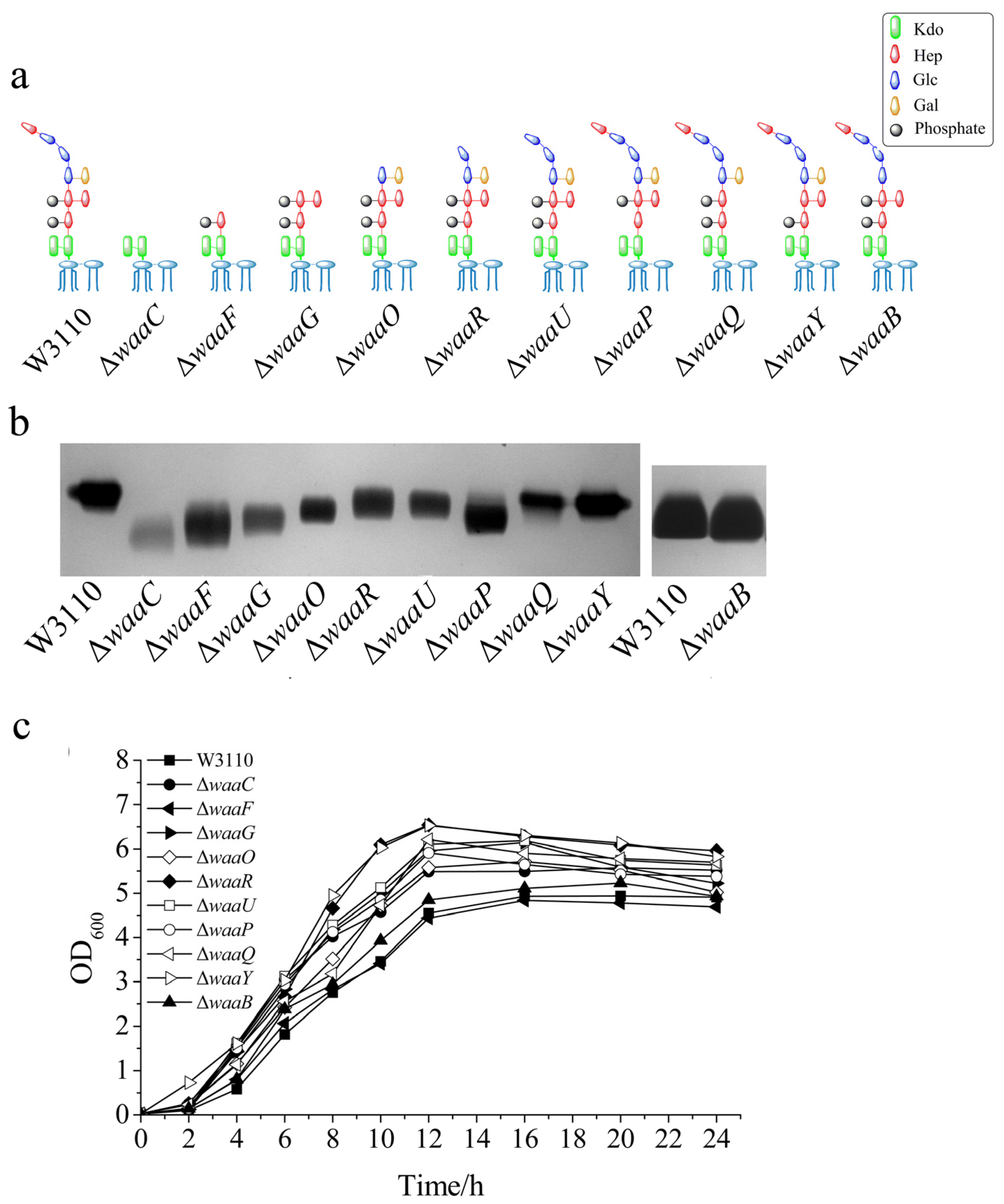
2.1. Construction of 10 E. coli LPS Core OS Mutant Strains and Comparison of Their LPS Structure and Cell Growth

2.2. The Effect of LPS Core OS on Cell Surface Hydrophobicity and Outer Membrane Permeability
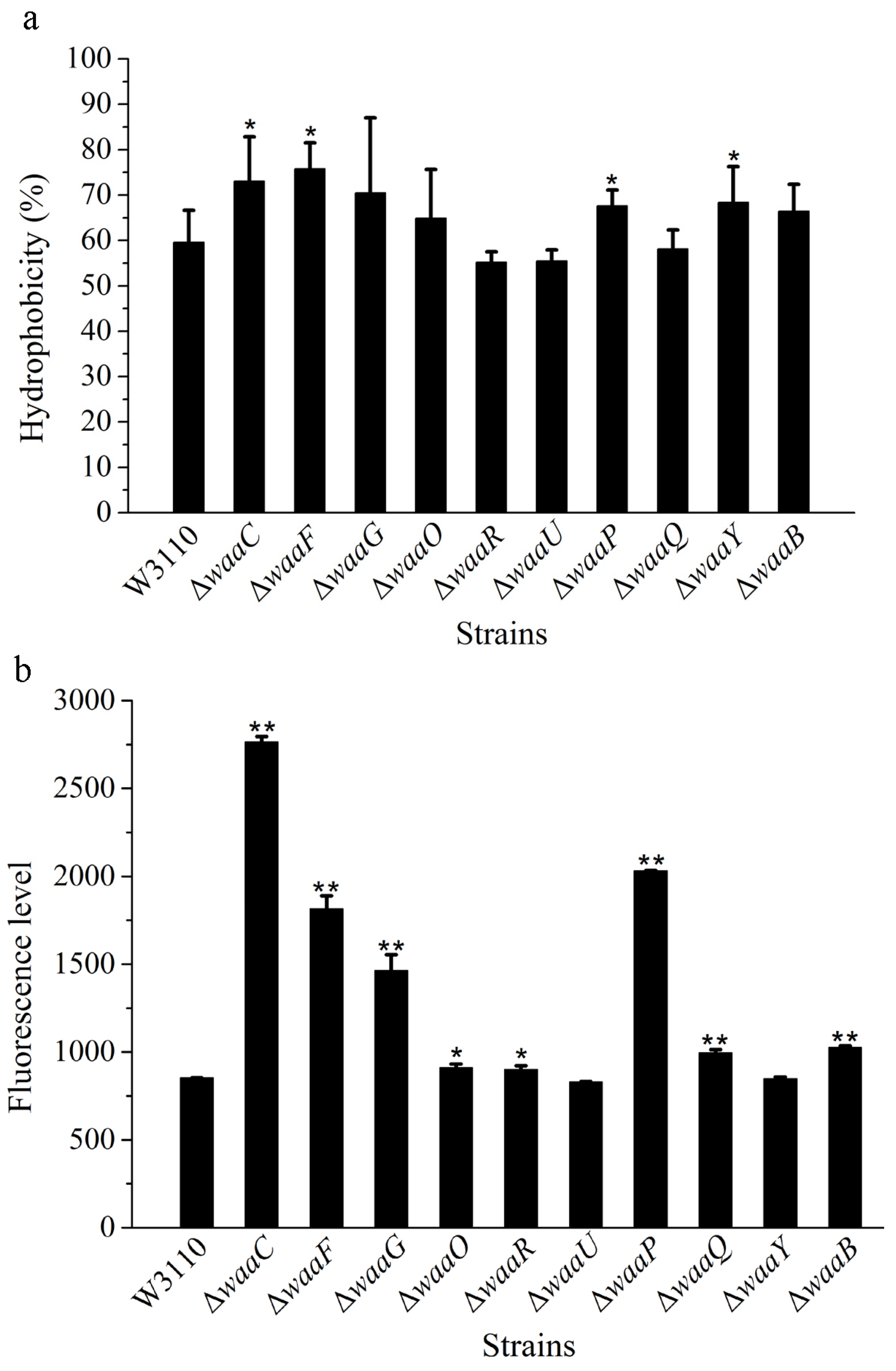

{kind=link}
{kind=link}
{kind=link}
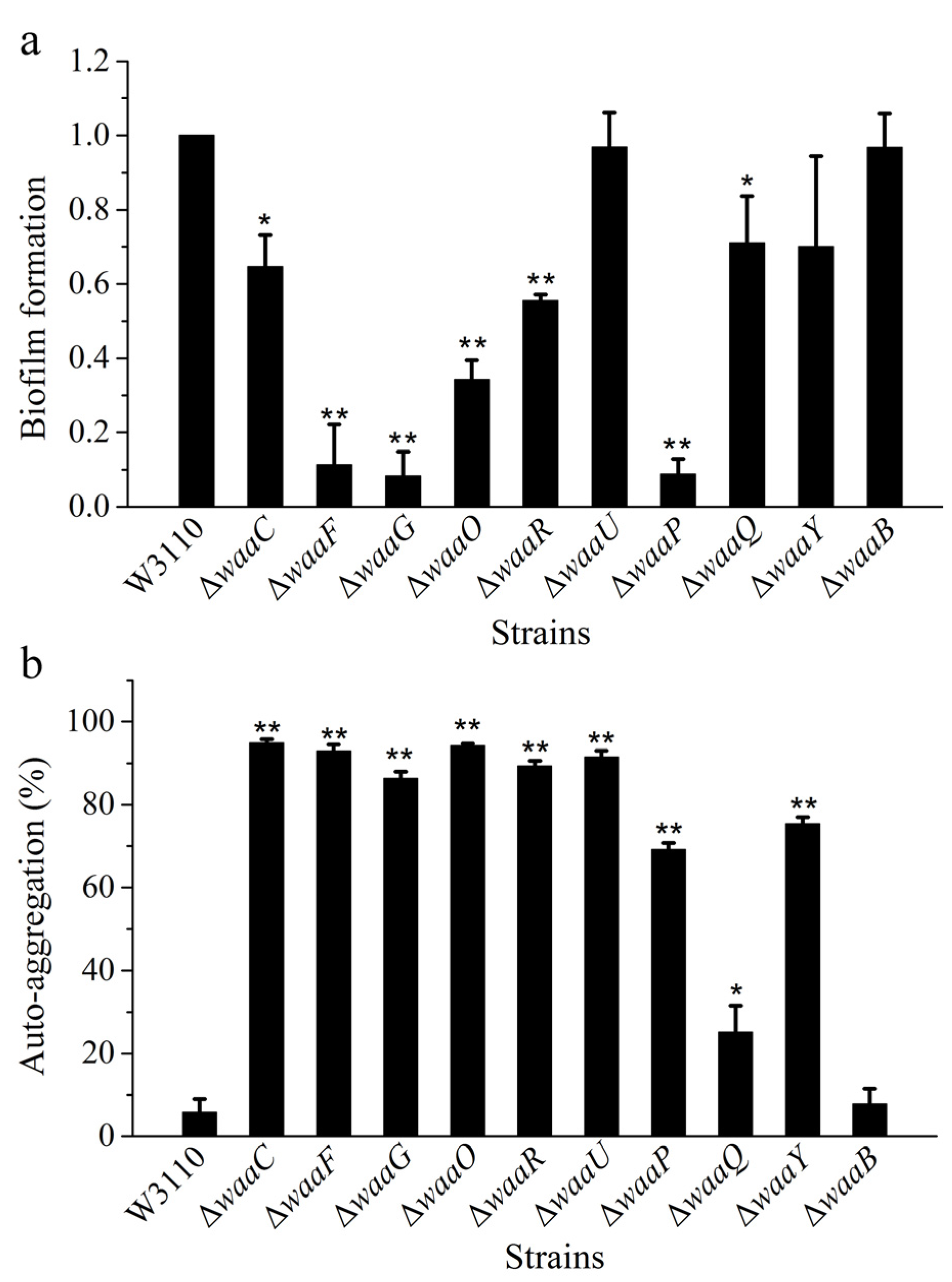{kind=link}
| Strain | MIC (μg/mL) | |
|---|---|---|
| Erythromycin | Novobiocin | |
| W3110 | >500 | >500 |
| ΔwaaC | 62.5 | 15.6 |
| ΔwaaF | 62.5 | 31.3 |
| ΔwaaG | 125 | 250 |
| ΔwaaO | 500 | >500 |
| ΔwaaR | >500 | >500 |
| ΔwaaU | >500 | >500 |
| ΔwaaP | 125 | 15.6 |
| ΔwaaQ | 500 | >500 |
| ΔwaaY | >500 | 250 |
| ΔwaaB | 500 | >500 |
2.3. Comparison of Biofilm Formation and Auto-Aggregation of 10 E. coli LPS Mutant Strains
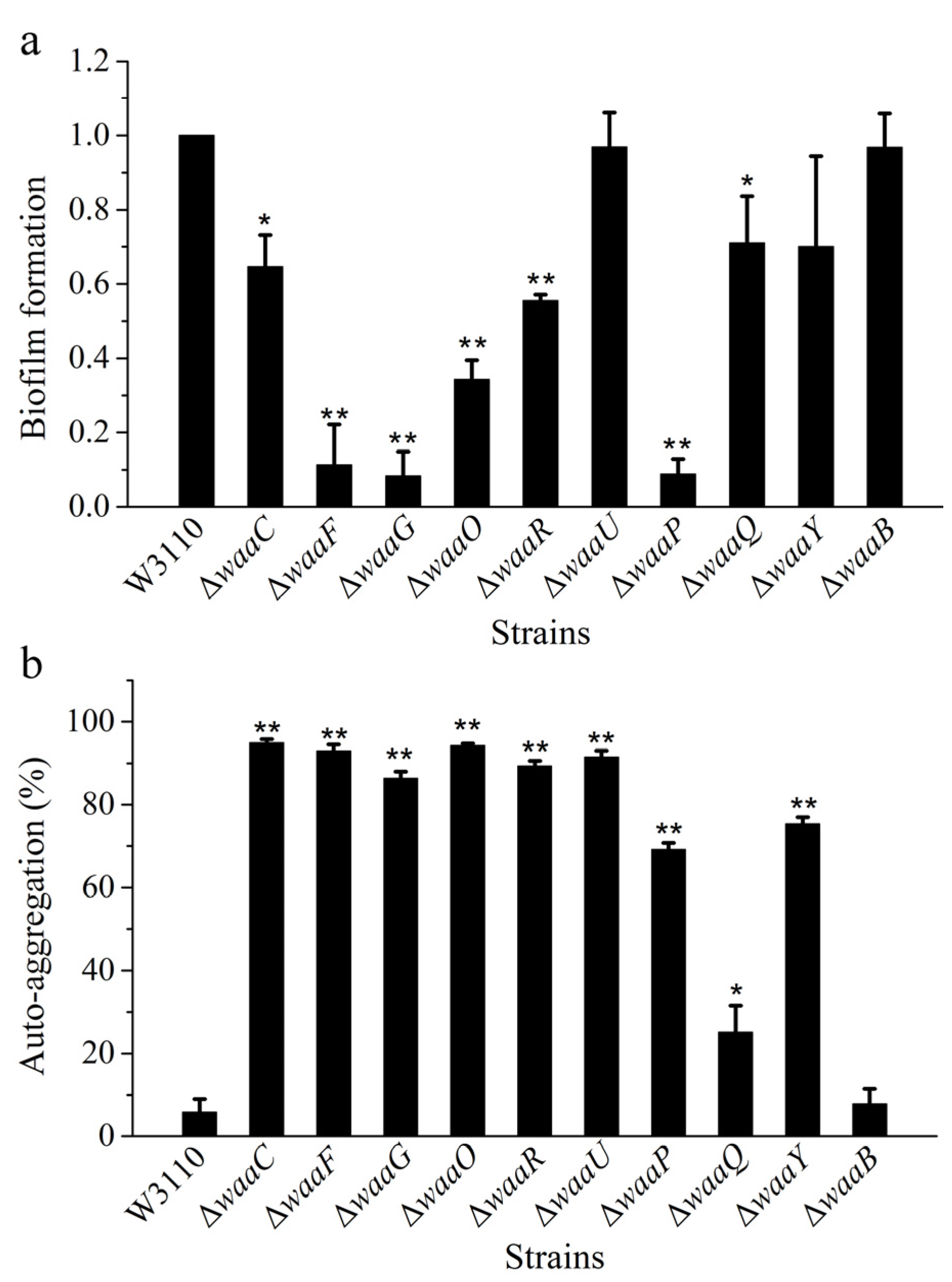
3. Experimental Section
3.1. Bacterial Strains and Growth Condition
3.2. DNA Manipulation
| Primers | Sequence (5′→3′) | Restriction Enzyme |
|---|---|---|
| waaC-U-F | CCGCTCGAGTAAATCAAGCAAGCCTAT | XhoI |
| waaC-U-R | CGGAATTCCAGTCAAGCAGTTTTGGA | EcoRI |
| waaC-D-F | CCCAAGCTTATCCGTCAGGCTTCCTCT | HindIII |
| waaC-D-R | AAAACTGCAGCTGGTTGCCCTGTATGGT | PstI |
| waaF-U-F | CCGCTCGAGAGAACCAGGCTTTAC | XhoI |
| waaF-U-R | CGGAATTCAGAGGAAGCCTGACGGAT | EcoRI |
| waaF-D-F | CCCAAGCTTAGCTCTTATGCGTCGCGATTCAG | HindIII |
| waaF-D-R | AAAACTGCAGTGCTACGCTGGCTTATC | PstI |
| waaG-U-F | CCGCTCGAGGCGAGGCTATCAGGTTGT | XhoI |
| waaG-U-R | CGGAATTCTATGGGACTTAACTGGCACCTG | EcoRI |
| waaG-D-F | CCCAAGCTTCTCGGCGTGCGGAGCAATGT | HindIII |
| waaG-D-R | AAAACTGCAGCACTCAGGCGATGAATAG | PstI |
| waaO-U-F | CCGCTCGAGTATCAGTGCCGATTGTGTC | XhoI |
| waaO-U-R | CGGAATTCTCCTGGAAAAACACCTGCT | EcoRI |
| waaO-D-F | CCCAAGCTTAGGATTTAGCAACTATC | HindIII |
| waaO-D-R | AAAACTGCAGTGGCAGGAAATGAGTCC | PstI |
| waaR-U-F | CCGCTCGAGAGGGTAGCATTGTGGACT | XhoI |
| waaR-U-R | CGGAATTCTGTGATGGAAACACCTAC | EcoRI |
| waaR-D-F | CCCAAGCTTCAACTAAACCGTGGCACAA | HindIII |
| waaR-D-R | AAAACTGCAGCTCCTGCTATAATTCCTG | PstI |
| waaU-U-F | CCGCTCGAGCTTCAAGACATCAGTGCAG | XhoI |
| waaU-U-R | CGGAATTCGTGAAAAGTTCCTAAGC | EcoRI |
| waaU-D-F | CCCAAGCTTTCCCTCGCATTTAATTTGGTCC | HindIII |
| waaU-D-R | AAAACTGCAGGGGGATTGGACTCAGTGATGTG | PstI |
| waaP-U-F | CCGCTCGAGGGTGGTTTAGATGGTTG | XhoI |
| waaP-U-R | CGGAATTCACTCAGGCGATGAATAG | EcoRI |
| waaP-D-F | CCCAAGCTTTTTTGGGATGCCTTTA | HindIII |
| waaP-D-R | AAAACTGCAGAATCCTTTGCGTTGTGTT | PstI |
| waaQ-U-F | CCGCTCGAGAGCACGTCAAAGTAAGT | XhoI |
| waaQ-U-R | CGGAATTCCTTTATGACCAGGATTT | EcoRI |
| waaQ-D-F | CCCAAGCTTAATTATGATCGTGGCG | HindIII |
| waaQ-D-R | AAAACTGCAGTGGTTGGTATGGGACTT | PstI |
| waaY-U-F | CCGCTCGAGCAGGAATTATAGCAGGA | XhoI |
| waaY-U-R | CGGAATTCCGGTAAAAACAACCAAGTC | EcoRI |
| waaY-D-F | CCCAAGCTTCCCATCGTGGTAACTTCA | HindIII |
| waaY-D-R | AAAACTGCAGCTTTCAAACGCCGCAT | PstI |
| waaB-U-F | CCGCTCGAGATAGCGTTTATCGGCGAAGC | XhoI |
| waaB-U-R | CGGAATTCGCCATCCCTTATCCATTTTG | EcoRI |
| waaB-D-F | CCCAAGCTTTTTCCCAATGACCCTACT | HindIII |
| waaB-D-R | AAAACTGCAGATCTCCCGGTTGATACAGA | PstI |
| Fkan-F | CGGAATTCGTGTAGGCTGGAGCTGCTTCG | EcoRI |
| Fkan-R | CCCAAGCTTGCCATTAATTCACTGATCAG | HindIII |
| Strains or Plasmids | Description | Source |
|---|---|---|
| Strains | ||
| W3110 | Wild-type E. coli, F−, λ− | Laboratory strain |
| W3110/pKD46 | W3110 transformed by pKD46 | This work |
| ΔwaaC | W3110 ΔwaaC | This work |
| ΔwaaF | W3110 ΔwaaF | This work |
| ΔwaaG | W3110 ΔwaaG | This work |
| ΔwaaO | W3110 ΔwaaO | This work |
| ΔwaaR | W3110 ΔwaaR | This work |
| ΔwaaU | W3110 ΔwaaU | This work |
| ΔwaaP | W3110 ΔwaaP | This work |
| ΔwaaQ | W3110 ΔwaaQ | This work |
| ΔwaaY | W3110 ΔwaaY | This work |
| ΔwaaB | W3110 ΔwaaB | This work |
| Plasmids | ||
| pKD46 | ParaBγβ exo, Repts,AmpR | [30] |
| pKD13 | oriR6K, FRT KanR FRT, AmpR | [30] |
| pCP20 | FLP+, λ cI857+, λpRRepts, CamR, AmpR | [30] |
| pBlueScript II SK+ | Cloning vector, ColE1, lacZ, AmpR | Stratagene |
3.3. LPS Preparation and Analysis
3.4. Cell Surface Hydrophobicity Assay
3.5. Membrane Permeability Assay
3.6. Antibiotic Susceptibility Assay
3.7. Quantification of Biofilm Formation
3.8. Auto-Aggregation Assay
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, X.; Quinn, P.J. Lipopolysaccharide: Biosynthetic pathway and structure modification. Prog. Lipid. Res. 2010, 49, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Reeves, P.R. Escherichia coli K12 regains its O antigen. Microbiology 1994, 140, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Yethon, J.A.; Heinrichs, D.E.; Monteiro, M.A.; Perry, M.B.; Whitfield, C. Involvement of waaY, waaQ, and waaP in the modification of Escherichia coli lipopolysaccharide and their role in the formation of a stable outer membrane. J. Biol. Chem. 1998, 273, 26310–26316. [Google Scholar] [CrossRef] [PubMed]
- Genevaux, P.; Bauda, P.; DuBow, M.S.; Oudega, B. Identification of Tn10 insertions in the rfaG, rfaP, and galU genes involved in lipopolysaccharide core biosynthesis that affect Escherichia coli adhesion. Arch. Microbiol. 1999, 172, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pradel, E.; Parker, C.T.; Schnaitman, C.A. Structures of the rfaB, rfaI, rfaJ, and rfaS genes of Escherichia coli K-12 and their roles in assembly of the lipopolysaccharide core. J. Bacteriol. 1992, 174, 4736–4745. [Google Scholar] [PubMed]
- Yethon, J.A.; Vinogradov, E.; Perry, M.B.; Whitfield, C. Mutation of the lipopolysaccharide core glucosyltransferase encoded by waaG destabilizes the outer membrane of Escherichia coli by interfering with core phosphorylation. J. Bacteriol. 2000, 182, 5620–5623. [Google Scholar] [CrossRef] [PubMed]
- Vaara, M. Agents that increase the permeability of the outer membrane. Microbiol. Rev. 1992, 56, 395–411. [Google Scholar] [PubMed]
- Dirienzo, J.M.; Inouye, M. Effect of reduced membrane lipid fluidity on the biosynthesis of lipopolysaccharide of Escherichia coli. Eur. J. Biochem. 1983, 135, 351–357. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, A.M.; Moran, A.P.; Würzner, R.; Orren, A. Complement-mediated lipopolysaccharide release and outer membrane damage in Escherichia coli J5: Requirement for C9. Immunology 2001, 102, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, M.; Mileykovskaya, E.; Dowhan, W. Lipids in the assembly of membrane proteins and organization of protein supercomplexs. Subcell Biochem. 2008, 49, 197–239. [Google Scholar] [PubMed]
- Bengoechea, J.A.; Brandenburg, K.; Arraiza, M.D.; Seydel, U.; Skurnik, M.; Moriyón, I. Pathogenic Yersinia enterocolitica strains increase the outer membrane permeability in response to environmental stimuli by modulating lipopolysaccharide fluidity and lipid A structure. Infect. Immun. 2003, 71, 2014–2021. [Google Scholar] [CrossRef] [PubMed]
- Bryan, L.E.; O’hara, K.; Wong, S. Lipopolysaccharide changes in impermeability-type aminoglycoside resistance in Pseudomonas aeruginosa. Antimicrob. Agents. Chemother. 1984, 26, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Galbraith, L.; Wilkinson, S.G.; Legakis, N.J.; Genimata, V.; Katsorchis, T.A.; Rietschel, E.T. Structural alternations in the envelope of a gentamicin-resistant rough mutant of Pseudomonas aeruginosa. Ann. Microbiol. 1984, 135, 121–136. [Google Scholar]
- Kadurugamuwa, J.L.; Lam, J.S.; Beveridge, T.J. Interaction of gentamicin with the A band and B band lipopolysaccharides of Pseudomonas aeruginosa and its possible lethal effect. Antimicrob. Agents. Chemother. 1993, 37, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Kropinski, A.M.; Kuzio, J.; Angus, B.L.; Hancock, R.E. Chemical and chromatographic analysis of lipopolysaccharide from an antibiotic-supersusceptible mutant of Pseudomonas aeruginosa. Antimicrob. Agents. Chemother. 1982, 21, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Shearer, B.G.; Legakis, N.J. Pseudomonas aeruginosa: evidence for the involvement of lipopolysaccharide in determining outer membrane permeability to carbenicillin and gentamicin. J. Infect. Dis. 1985, 152, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, T.; de Kievit, T.R.; Masoud, H.; Altman, E.; Richards, J.C.; Sadovskaya, I.; Speert, D.P.; Lam, J.S. Characterization of lipopolysaccharide-deficient mutants of Pseudomonas aeruginosa derived from serotypes O3, O5, and O6. Infect. Immun. 1994, 62, 809–817. [Google Scholar] [PubMed]
- Joanna, S.B.; Albert, V.; Patrick, W.; Nicholas, A.D. Mutation of a lipopolysaccharide synthesis gene results in increased biofilm of Stenotrophomonas maltophilia on plastic and glass surfaces. Ann. Microbiol. 2008, 58, 35–40. [Google Scholar] [CrossRef]
- Balestrino, D.; Ghigo, J.M.; Charbonnel, N.; Haagensen, J.A.; Forestier, C. The characterization of functions involved in the establishment and maturation of Klebsiella pneumoniae in vitro biofilm reveals dual roles for surface exopolysaccharides. Environ. Microbiol. 2008, 10, 685–701. [Google Scholar] [CrossRef] [PubMed]
- Shelud'ko, A.V.; Kulibiakina, O.V.; Shirokov, A.A.; Petrova, L.P.; Matora, Liu; Katsy, E.I. The effect of mutations in the synthesis of lipopolysaccharides and calcofluor-binding polysaccharides on biofilm formation by Azospirillum brasilense. Mikrobiologiia 2008, 77, 358–363. [Google Scholar]
- Karunakaran, E.; Mukherjee, J.; Ramalingam, B.; Biggs, C.A. “Biofilmology”: A multidisciplinary review of the study of microbial biofilms. Appl. Microbiol. Biotechnol. 2011, 90, 1869–1881. [Google Scholar] [CrossRef] [PubMed]
- Ciornei, C.D.; Novikov, A.; Beloin, C.; Fitting, C.; Caroff, M.; Ghigo, J.M.; Cavaillon, J.M.; Adib-Conquy, M. Biofilm-forming Pseudomonas aeruginosa bacteria undergo lipopolysaccharide structural modifications and induce enhanced inflammatory cytokine response in human monocytes. Innate. Immunity 2010, 16, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.H.; Li, P.L.; Liu, Z. The correlation between surface hydrophobicity and adherence of Bifidobacterium strains from centenarians’ faeces. Anaerobe 2006, 12, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Del Re, B.; Sgorbati, B.; Miglioli, M.; Palenzona, D. Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett. Appl. Microbiol. 2000, 31, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Perez, P.F.; Minnaard, Y.; Disalvo, E.A.; de Antoni, G.L. Surface properties of bifidobacterial strains of human origin. Appl. Environ. Microb. 1998, 64, 21–26. [Google Scholar]
- Austin, E.A.; Graves, J.F.; Hite, L.A.; Parker, C.T.; Schnaitman, C.A. Genetic analysis of lipopolysaccharide core biosynthesis by Escherichia coli K-12: Insertion mutagenesis of the rfa locus. J. Bacteriol. 1990, 172, 5312–5325. [Google Scholar] [PubMed]
- Serra-Moreno, R.; Acosta, S.; Hernalsteens, J.P.; Jofre, J.; Muniesa, M. Use of the lambda Red recombinase system to produce recombinant prophages carrying antibiotic resistance genes. BMC Mol. Biol. 2006, 7. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed]
- Westphal, O.; Jann, K. Bacterial Lipopolysaccharide-Extraction with Phenol Water and Further Application of Procedure. In Methods in Carbohydrate Chemistry; Whistler, R.L., Wolfrom, M.L., Eds.; Academic Press: New York, NY, USA, 1965; pp. 83–91. [Google Scholar]
- Gomez Zavaglia, A.; Kociubinski, G.; Perez, P.; Disalvo, E.; de Antoni, G. Effect of bile on the lipid composition and surface properties of bifidobacteria. J. Appl. Microbiol. 2002, 93, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Helander, I.M.; Mattila-Sandholm, T. Fluorometric assessment of gram-negative bacterial permeabilization. J. Appl. Microbiol. 2000, 88, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Kirov, S.M.; Castrisios, M.; Shaw, J.G. Aeromonas flagella (polar and lateral) are enterocyte adhesins that contribute to biofilm formation on surfaces. Infect. Immun. 2004, 72, 1939–1945. [Google Scholar] [CrossRef] [PubMed]
- Hasman, H.; Chakraborty, T.; Klemm, P. Antigen-43-mediated autoaggregation of Escherichia coli is blocked by fimbriation. J. Bacteriol. 1999, 181, 4834–4841. [Google Scholar] [PubMed]
- Rahman, M.M.; Kim, W.S.; Kumura, H.; Shimazaki, K. Autoaggregation and surface hydrophobicity of bifidobacteria. World J. Microb. Biotechnol. 2008, 24, 1593–1598. [Google Scholar] [CrossRef]
- Wang, J.; Ma, W.; Wang, Z.; Li, Y.; Wang, X. Construction and characterization of an Escherichia coli mutant producing Kdo2-lipid A. Mar. Drugs 2014, 12, 1495–1511. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Z.; Chen, J.; Ernst, R.K.; Wang, X. Influence of lipid A acylation pattern on membrane permeability and innate immune stimulation. Mar. Drugs 2013, 11, 3197–3208. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Li, Y.; Chen, J.; Tan, Y.; Guan, F.; Wang, X. Construction of monophosphoryl lipid A producing Escherichia coli mutants and comparison of immuno-stimulatory activities of their lipopolysaccharides. Mar. Drugs 2013, 11, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tao, G.; Wang, X. Construction of an Escherichia coli mutant producing monophosphoryl lipid A. Biotechnol. Lett. 2011, 33, 1013–1019. [Google Scholar] [CrossRef]
- Wang, X.; Quinn, P.J.; Yan, A. Kdo2-lipid A: Structural diversity and impact on immunopharmacology. Biol. Rev. Camb. Philos. Soc. 2015, 90, 408–427. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Wang, J.; Ren, G.; Li, Y.; Wang, X. Influence of Core Oligosaccharide of Lipopolysaccharide to Outer Membrane Behavior of Escherichia coli. Mar. Drugs 2015, 13, 3325-3339. https://doi.org/10.3390/md13063325
Wang Z, Wang J, Ren G, Li Y, Wang X. Influence of Core Oligosaccharide of Lipopolysaccharide to Outer Membrane Behavior of Escherichia coli. Marine Drugs. 2015; 13(6):3325-3339. https://doi.org/10.3390/md13063325
Chicago/Turabian StyleWang, Zhou, Jianli Wang, Ge Ren, Ye Li, and Xiaoyuan Wang. 2015. "Influence of Core Oligosaccharide of Lipopolysaccharide to Outer Membrane Behavior of Escherichia coli" Marine Drugs 13, no. 6: 3325-3339. https://doi.org/10.3390/md13063325