Cephalopod Ink: Production, Chemistry, Functions and Applications

Abstract
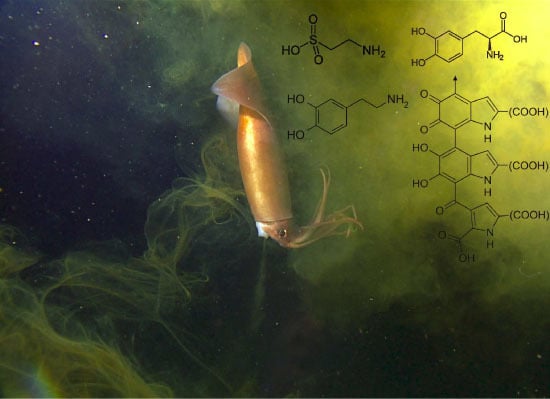
:
1. Introduction
2. Cephalopods: Who Are They and Who Inks
2.1. Cephalopod Systematics
2.2. Which Cephalopods Produce Ink?


3. What Is Cephalopod Ink?
3.1. Ink Is a Mixture of Two Glandular Secretions
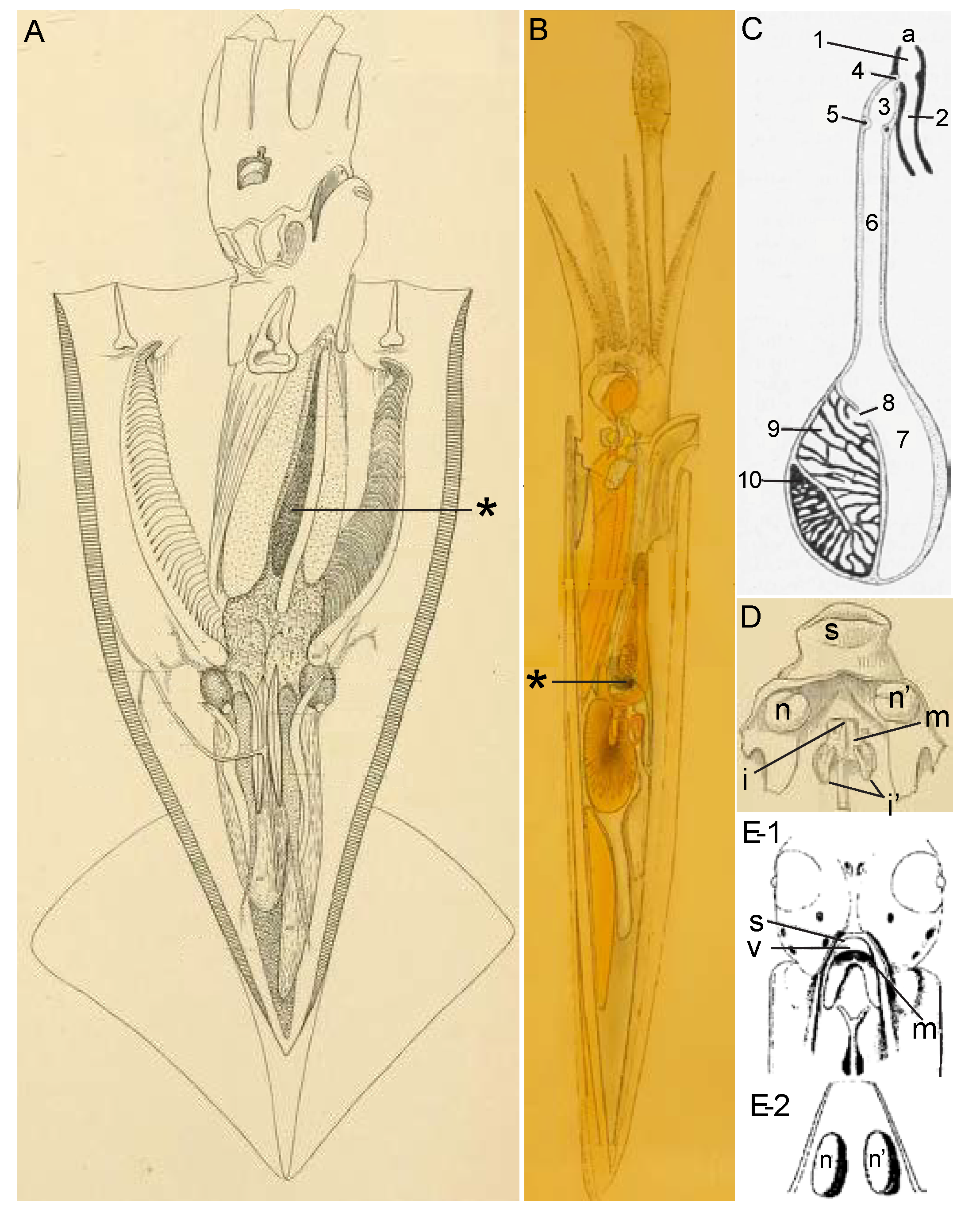
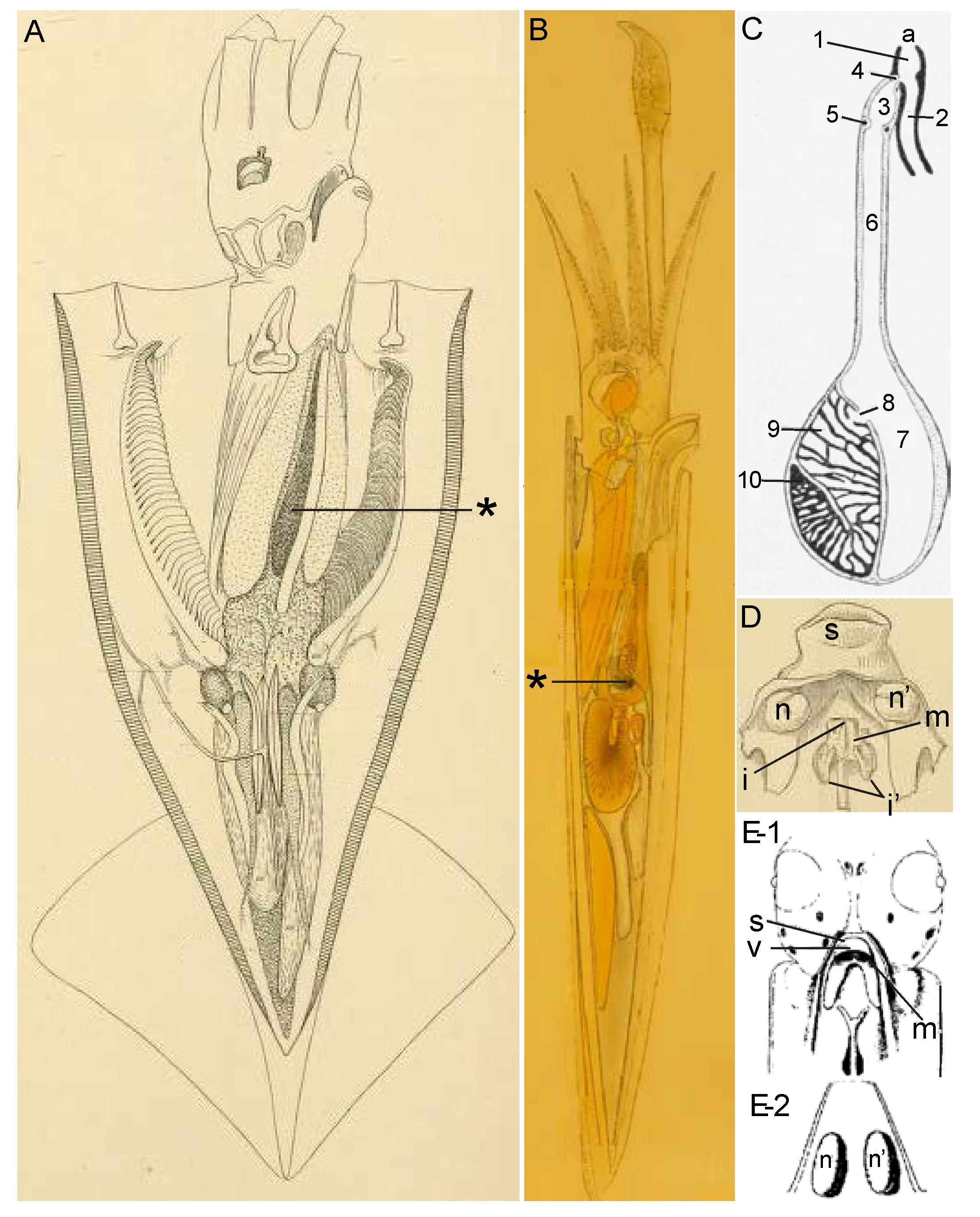
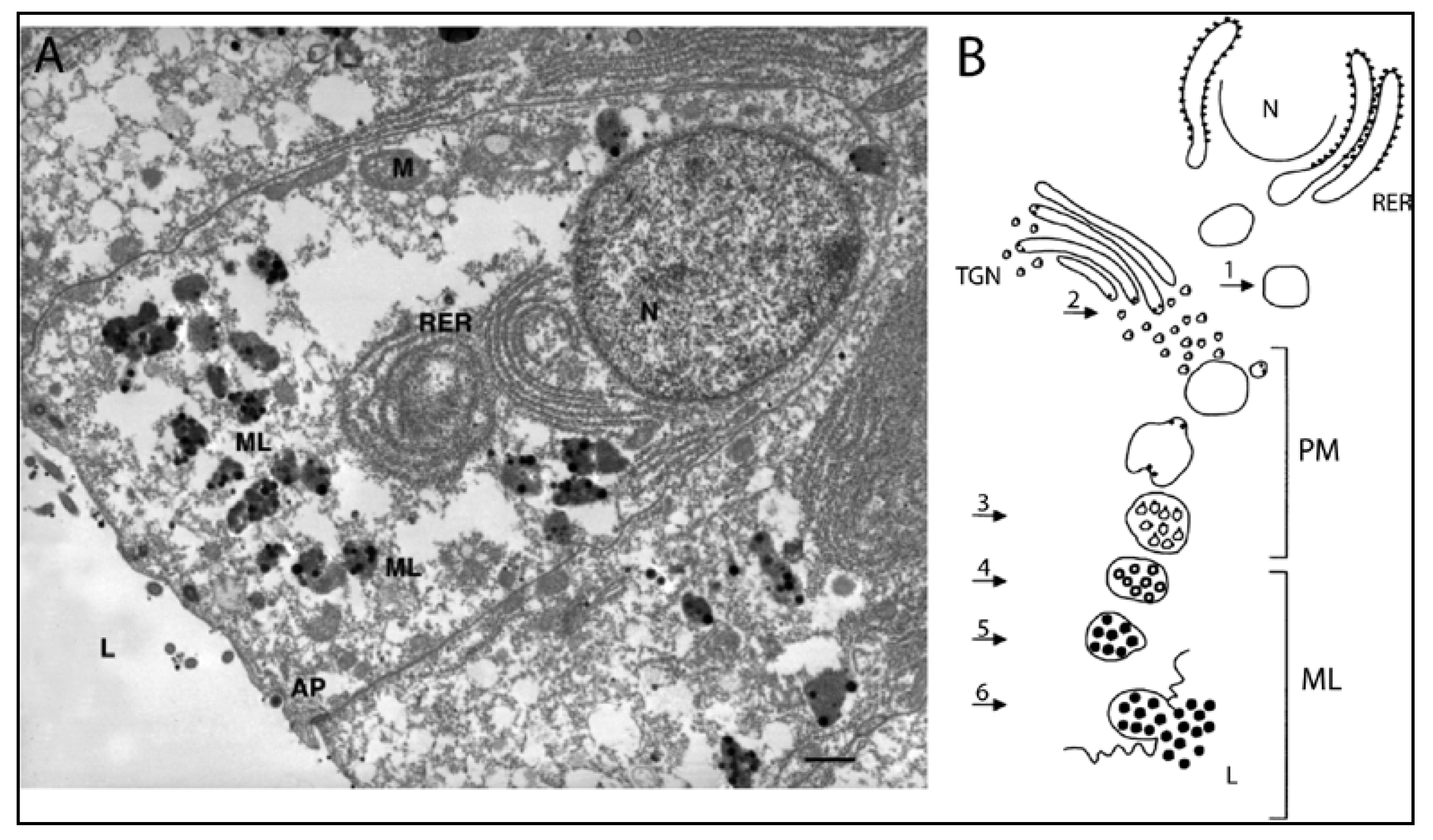
3.1.1. Ink Sac
3.1.2. Funnel Organ
3.1.3. Combining the Two Glandular Secretions
4. Chemical Constituents of Ink
4.1. Methodology Matters
4.2. General Composition of Ink
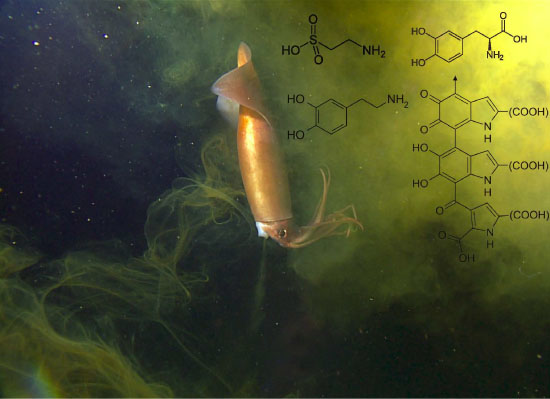
4.3. Melanin
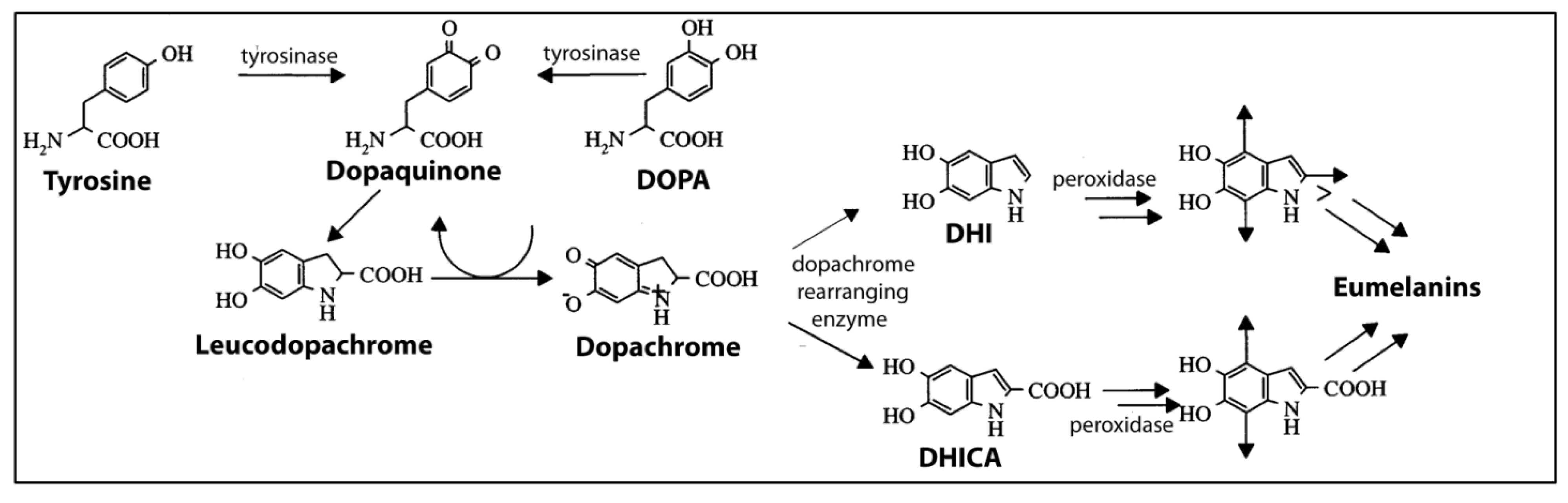
4.3.1. What Is Melanin?
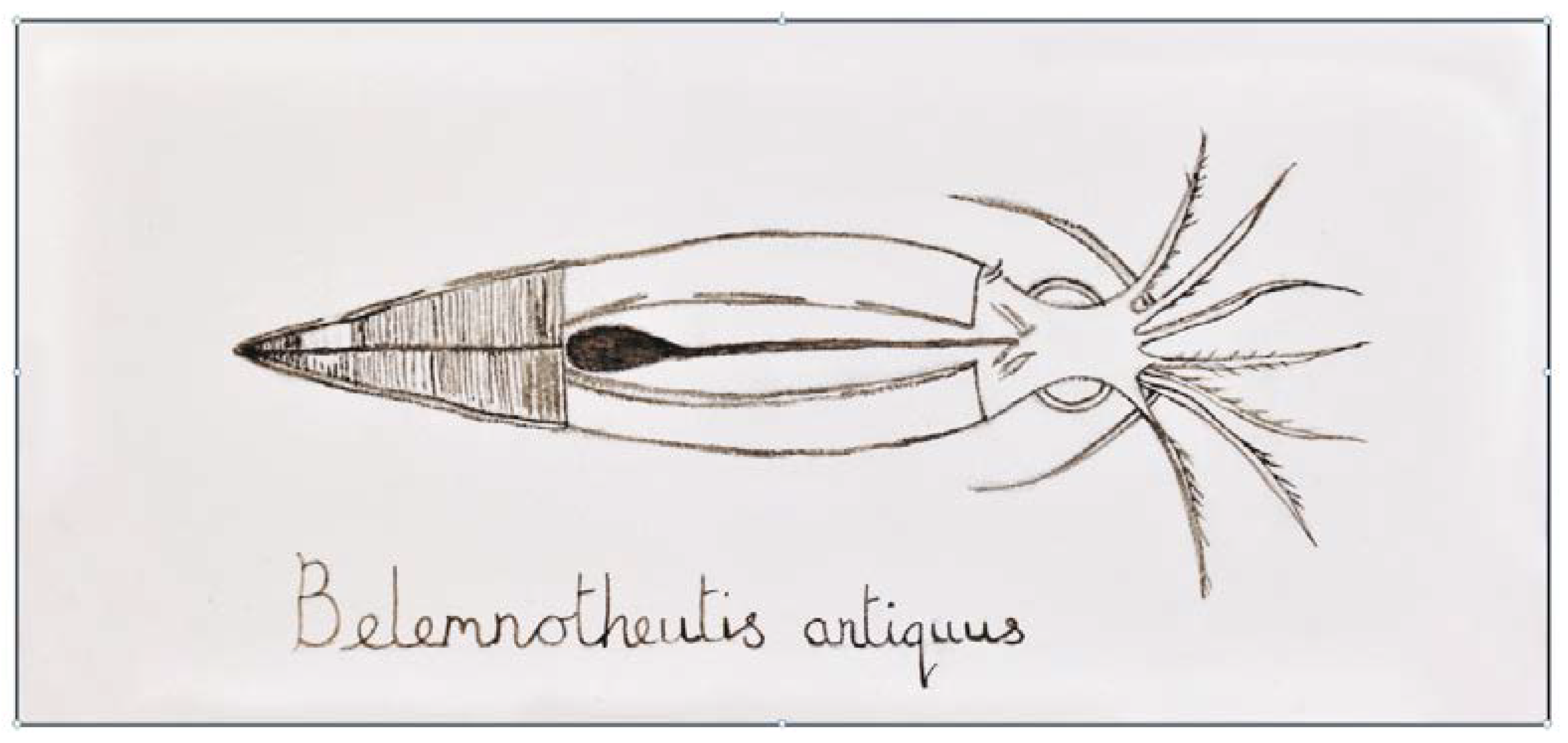
4.3.2. Production of Melanin in Cephalopods
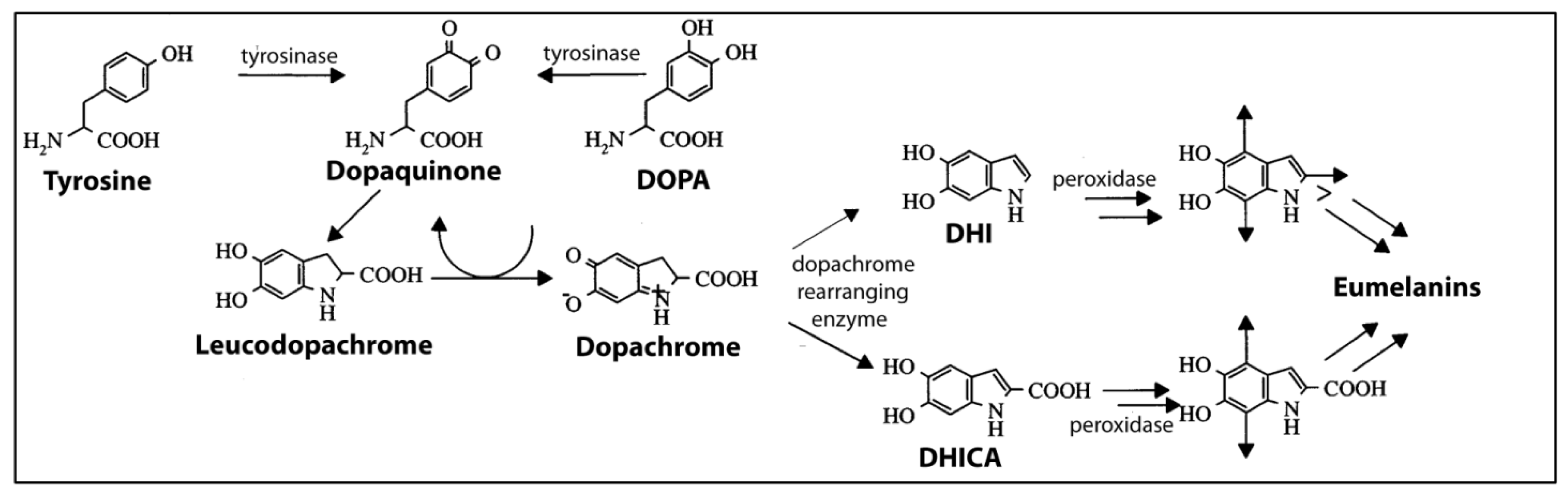
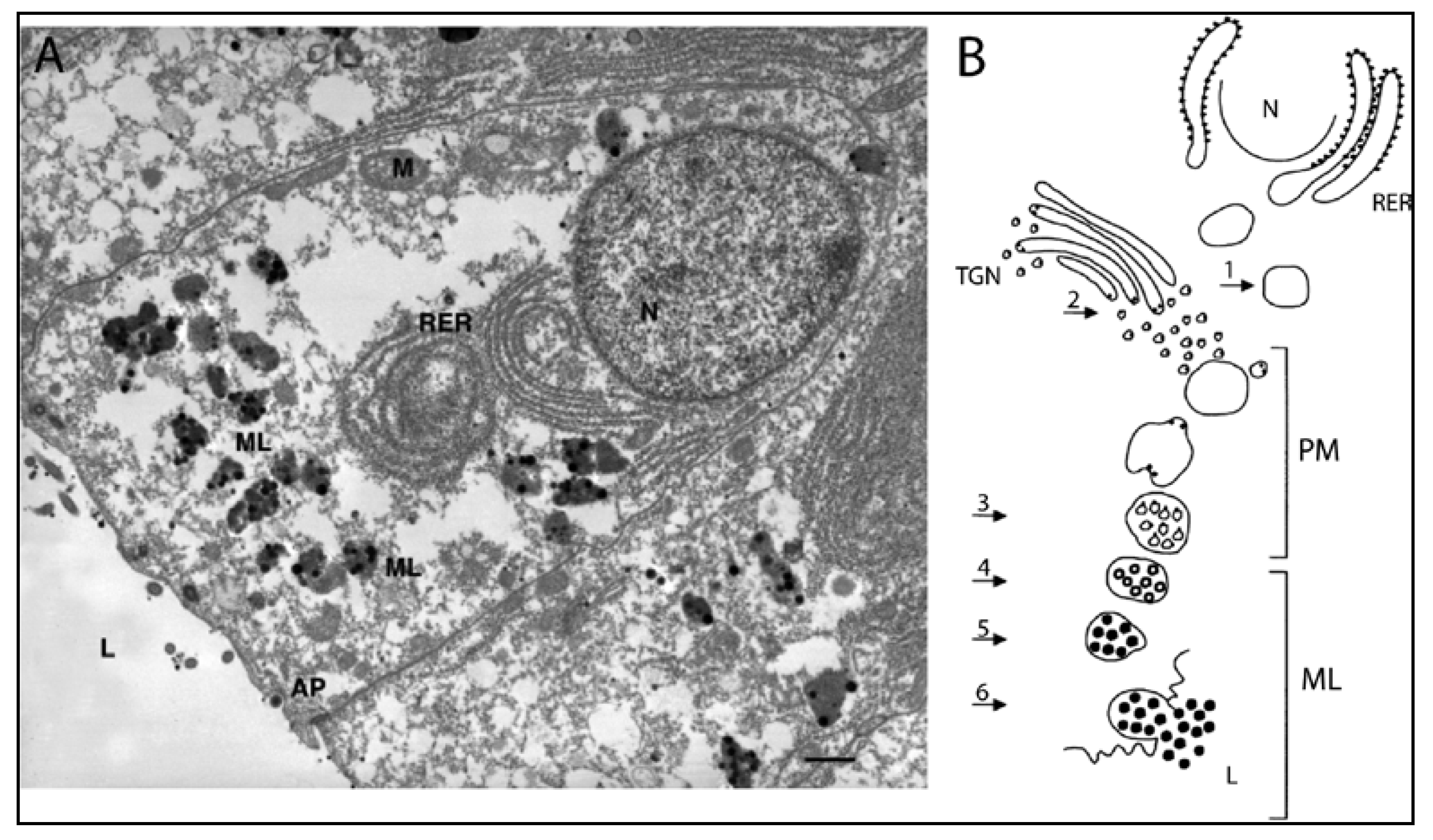
4.4. Melanin-Related Compounds
4.4.1. Tyrosinases
4.4.2. Dopachrome Rearranging Enzymes
4.4.3. Peroxidases
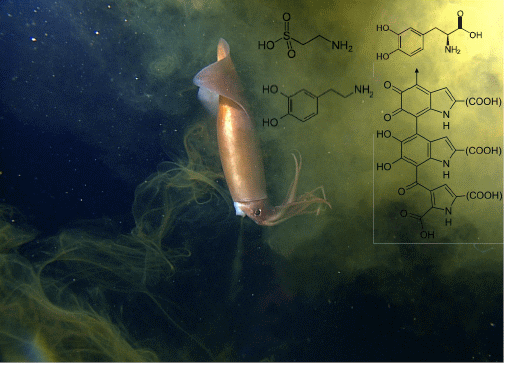
4.4.4. Catecholamines
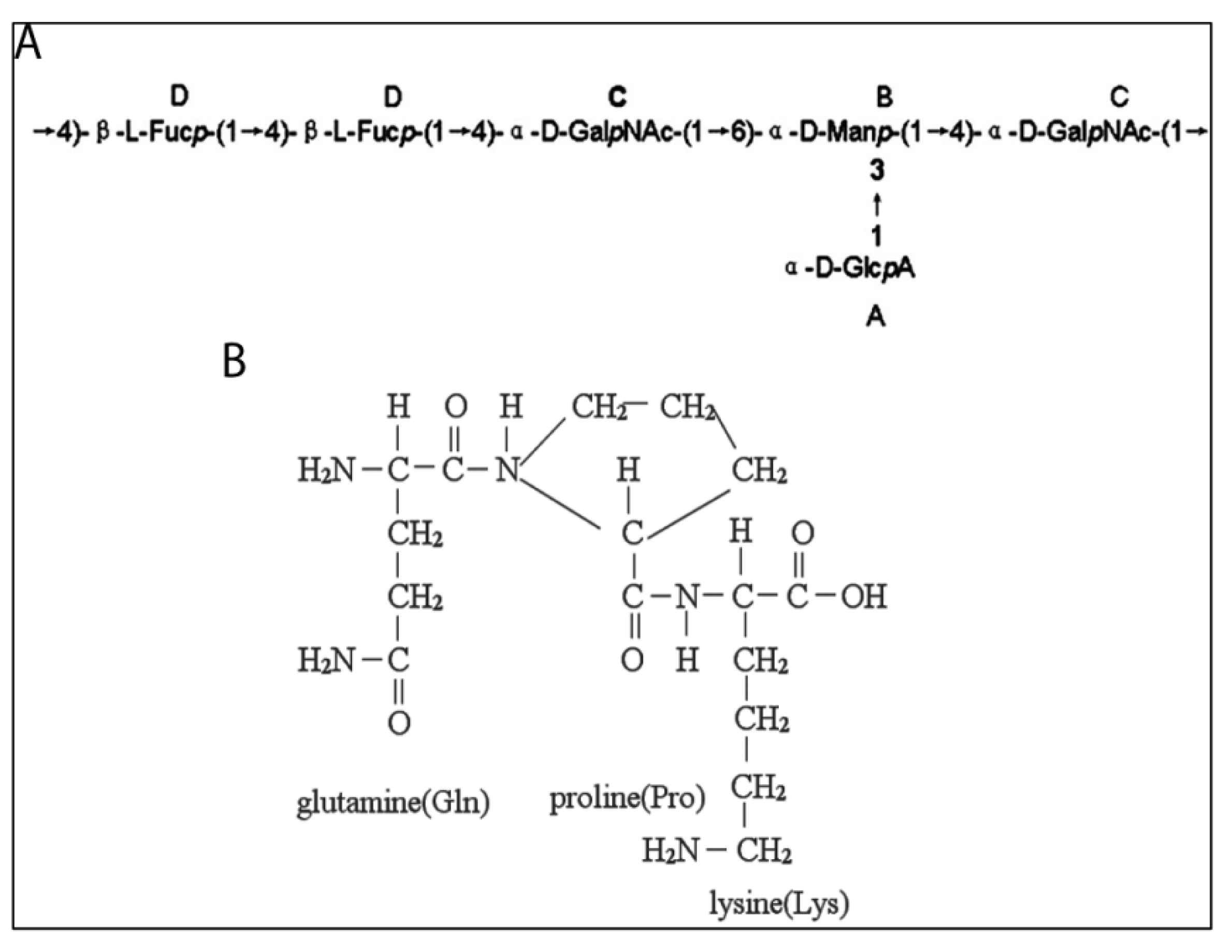
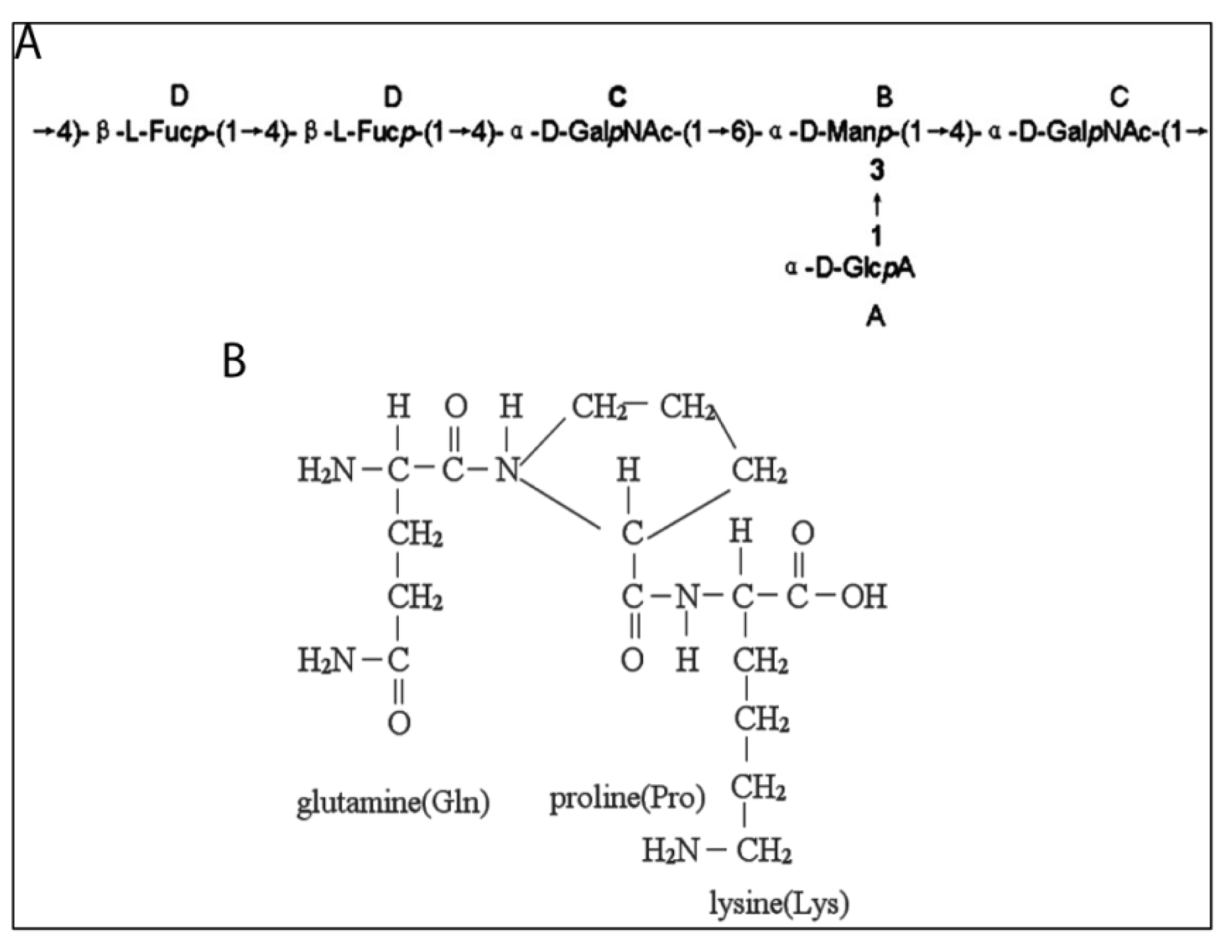
4.5. Peptidoglycans
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
4.6. Amino Acids
4.7. Metals
4.8. Toxins
5. Ink as an Anti-Predator Defense
5.1. Interspecific Effects: Ink as a Direct Deterrent of Predators
5.2. Intraspecific Effects: Ink as an Alarm Cue for Conspecifics

5.3. Evolution of Melanin as a Defense
6. Human Applications of Cephalopod Ink
6.1. Drugs
6.1.1. Antimicrobial Properties
6.1.2. Potential Anticancer Properties
6.1.3. Hematopoietic Effects
6.1.4. Anti-Hypertensive Actions
6.1.6. Potential Anti-Ulcerogenic Actions
6.1.7. Anti-Inflammatory Activity
6.1.8. Anti-Oxidant Activity
6.2. Sepia Ink as a Pigment in Writing, Art and Cosmetics

6.3. Cephalopod Ink as Food
7. Future Directions
8. Conclusions
Acknowledgments
Conflicts of Interest
References
- Schmitt, C.B. Aristotle as a cuttlefish: The origin and development of a Renaissance image. Stud. Renaiss. 1965, 12, 60–72. [Google Scholar] [CrossRef]
- Hanlon, R.T.; Messenger, J.B. Cephalopod Behaviour; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Voight, J. Cladistic analysis of the octopods based on anatomical characters. J. Molluscan Stud. 1997, 63, 311–325. [Google Scholar]
- Boyle, P.; Rodhouse, P. Cephalopods. Ecology and Fisheries; Blackwell Science: Oxford, UK, 2005. [Google Scholar]
- Norman, M.D. Ameloctopus litoralis, gen. et sp. Nov. (Cephalopoda: Octopodidae), a new shallow-water octopus from tropical waters. Invert. Taxon. 1992, 6, 567–582. [Google Scholar] [CrossRef]
- Bush, S.L.; Robison, B.H. Ink utilization by mesopelagic squid. Mar. Biol. 2007, 152, 485–494. [Google Scholar] [CrossRef]
- Von Boletzky, S. Juvenile behavior. In Cephalopod Life Cycles; Boyle, P.R., Ed.; Academic Press: London, UK, 1987; Volume II, pp. 45–84. [Google Scholar]
- Young, R.E.; Mangold, K.M. Cephalopod Pseudomorph Function. Tree of Life Web Project. Available online: http://tolweb.org/accessory/Cephalopod_Pseudomorph_Function?acc_id=1964 (accessed on 24 April 2014).
- Verrill, A.E. The cephalopods of the north-eastern coast of America. Part II. The smaller cephalopods, including the squids and the octopi, with other allied forms. Trans. Conn. Acad. Sci. 1880, 5, 259–446. [Google Scholar]
- Williams, L.W. The Anatomy of the Common Squid, Loligo Pealii Lasueur; Library and Printing-Office late E.J. Brill: Leiden, The Netherlands, 1909. [Google Scholar]
- Lang, A.; Hescheler, K. Lehrbuch der Vergleichenden Anatomie der Wirbellosen Thiere. Zweite Umgearbeitete Auflage. Erste Lieferung: Mollusca; Gustav Fisher: Jena, Germany, 1900. [Google Scholar]
- Laurie, M. The organ of Verrill in Loligo. Quart. J. Microsc. Soc. Lond. 1888, 29, 97–100. [Google Scholar]
- Budelmann, B.-U.; Schipp, R.; von Boletzky, S. Cephalopoda. In Microscopic Anatomy of Invertebrates, Mollusca II; Harrison, F.W., Kohn, A.J., Eds.; Wiley-Liss: New York, NY, USA, 1997; Volume 6A, pp. 119–414. [Google Scholar]
- Girod, P. Recherches sur la poche du noir des céphalopods des côte de France. Arch. Zool. Exp. Gén. 1882, 10, 1–100. [Google Scholar]
- Mangold, K.; Bidder, A.M.; Portmann, A. Structures cutaneés: La poche du noir. In Céphalopodes. Traité de Zoologie 5/4; Grassé, P.P., Ed.; Masson: Paris, France, 1989; pp. 154–162. [Google Scholar]
- Turchini, J. Étude histologique de la poche du noir des céphalopodes dibranchiaux. Les processus cytologiques de la sécrétion et de l’extrétion de l’encre. Arch. Anat. Microsc. Paris 1921, 18, 328–356. [Google Scholar]
- Turchini, J. Note histologique sur l’excrétion du noir de la seiche. Bull. Inst. Oceanogr. Monaco 1922, 412, 1–4. [Google Scholar]
- Turchini, J. Nature muqueuse des cellules a mélanine de la glande du noir de la Seiche (Sepia officinalis L.) et méchanism de l’excrétion du pigment. Compte Rendus l’Acad. Sci. Biol. 1922, 86, 480–482. [Google Scholar]
- Yung Ko Ching, M. Contribution à l'étude cytologique de l’ovogénèse, de développement et de quelques ogans chez les céphalopodes. Annal. l’Instit. l’Oceanogr. 1930, 7, 299–364. [Google Scholar]
- Graupner, G.; Fischer, I. Das Tintendrüsenepithel von Sepia vor, wahrend, und nach der Pigmentdildung. Z. Zellforsch. Mikrosk Anat. 1934, 21, 329–341. [Google Scholar] [CrossRef]
- Ries, E. Die Pigmentbildung in der Tintendrüse von Sepia officinalis. Z. Zellforsch. 1937, 25, 1–13. [Google Scholar] [CrossRef]
- Fioroni, P. Die embryonale Entwicklung der Köllikerschen Organe von Octopus vulgaris Lam. Revue Suisse Zool. 1962, 69, 497–511. [Google Scholar]
- Fioroni, P. Zur embryonalen und postembryonalen Entwicklung der Epidermis bei zehnarmigen Tintenfischen. Verh. Naturforsch. Ges. Basel 1963, 74, 149–160. [Google Scholar]
- Fioroni, P. Die embryonale Entwicklung der Hautdrüsen und des Trichterorganes von Octopus vulgaris Lam. Acta Anat. (Basel) 1962, 50, 264–295. [Google Scholar] [CrossRef]
- Szabo, G.; Fitzpatrick, T.B.; Wilgram, G. The pigmentary system of the squid (Loligo pealei). Biol. Bull. 1963, 125, 360. [Google Scholar]
- Vogel, F.S.; McGregor, D.H. The fine structure and some biochemical correlates of melanogenesis in the ink gland of the squid. Lab. Investig. 1964, 13, 767–778. [Google Scholar]
- Dilly, P.N.; Herring, P.J. The light organ and ink sac of Heteroteuthis dispar (Mollusca: Cephalopoda). J. Zool. 1978, 186, 47–59. [Google Scholar] [CrossRef]
- Wang, C.-L.; Fan, X.-X.; Yu, H.-W.; Miao, M. Histology of the ink sac of Sepiella maindroni and ultrastructure of the ink formation. Curr. Zool. 2008, 54, 366–372. [Google Scholar]
- Hoyle, W.E. Report of the Cephalopoda collected by H.M.S. Challenger during the years 1873–1876. Zoology 1886, 16, 1–245. [Google Scholar]
- Weiss, F.E. On some oigopsid cuttle fishes. Quart. J. Microsc. Soc. Lond. 1888, 29, 75–96. [Google Scholar]
- Müller, H. Bau der Cephaopoden. Z. Wiss. Zool. 1853, 4, 337–358. [Google Scholar]
- Voss, G.L. Function and comparative morphology of the funnel organ in cephalopods. In Proceedings of the XVI International Congress of Zoology, Washington, DC, USA, 20–27 August 1963; Volume 1, p. 74.
- Hu, M.Y.; Sucré, E.; Charmantier-Daures, M.; Charmantier, G.; Lucassen, M.; Himmerkus, N.; Melzner, F. Localization of ion-regulatory epithelia in embryos and hatchings of two cephalopods. Cell Tissue Res. 2010, 339, 571–583. [Google Scholar] [CrossRef]
- Von Boletzky, S. Puffing smoke-rings underwater: The functional morphology of cephalopod ink ejectors. Vie Milieu 1997, 471, 180–181. [Google Scholar]
- Herring, P.J. Luminescence in cephalopods and fish. Symp. Zool. Soc. Lond. 1977, 38, 127–159. [Google Scholar]
- Robison, B.H.; Reisenbichler, K.R.; Hunt, J.C.; Haddock, S.H. Light production by the arm tips of the deep-sea cephalopd Vampyroteuthis infernalis. Biol. Bull. 2003, 205, 102–109. [Google Scholar] [CrossRef]
- Young, J.Z. The visceral nerves of Octopus. Philos. Trans. R. Soc. Lond. Biol. 1967, 253, 1–22. [Google Scholar]
- Palumbo, A.; di Cosmo, A.; Poli, A.; di Cristo, C.; d’Ischia, M. A calcium/calmodulin-dependent nitric oxide synthase, NMDAR2/3 receptor subunits and glutamate in the CNS of the cuttlefish Sepia officinalis: Localization in specific neural pathways controlling the inking system. J. Neurochem. 1999, 73, 1254–1263. [Google Scholar]
- Palumbo, A.; Fiore, G.; di Cristo, C.; di Cosmo, A.; d’Ischia, M. NMDA receptor stimulation induces temporary α-tubulin degradation signaled by nitric oxide-mediated tyrosine nitration in the nervous system of Sepia officinalis. Biochem. Biophys. Res. Commun. 2002, 293, 1536–1543. [Google Scholar] [CrossRef]
- Shigeno, S.; Yamamoto, M. Organization of the nervous system in the pygmy cuttlefish, Idiosepius paradoxus Ortmann (Idiosepiidae, Cephalopoda). J. Morphol. 2002, 254, 65–80. [Google Scholar] [CrossRef]
- Nixon, M.; Young, J.Z. The Brains and Lives of Cephalopods; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Boycott, B.B. The functional organization of the brain of the cuttlefish Sepia officinalis. Proc. R. Soc. Lond. Biol. 1961, 153, 503–534. [Google Scholar] [CrossRef]
- Bone, Q.; Howarth, J.V. The role of l-glutamate in neuromuscular transmission in some molluscs. J. Mar. Biol. Assoc. UK 1980, 60, 619–626. [Google Scholar] [CrossRef]
- Di Cosmo, A.; di Cristo, C.; Messenger, J.B. l-Glutamate and its ionotropic receptors in the nervous system of cephalopods. Curr. Neuropharmacol. 2006, 4, 305–312. [Google Scholar] [CrossRef]
- Young, J.Z. The nervous system of Loligo. II. Suboesophageal centres. Philos. Trans. R. Soc. Lond. Biol. 1976, 274, 101–167. [Google Scholar] [CrossRef]
- Young, J.Z. The Anatomy of the Nervous System of Octopus Vulgaris; Clarendon Press: Oxford, UK, 1971. [Google Scholar]
- Shigeno, S.; Tsuchiya, K.; Segawa, S. Embryonic and paralarval development of the central nervous system of the loliginid squid Sepioteuthis lessoniana. J. Comp. Neurol. 2001, 437, 449–475. [Google Scholar] [CrossRef]
- Hay, M.E. Marine chemical ecology: What is known and what is next? J. Exp. Mar. Biol. Ecol. 1996, 200, 103–134. [Google Scholar] [CrossRef]
- Riffell, J.A.; Krug, P.J.; Zimmer, R.K. Fertilzation in the sea: The chemical identity of an abalone sperm attractant. J. Exp. Biol. 2002, 205, 1439–1450. [Google Scholar]
- Yambe, H.; Kitamura, S.; Kamio, M.; Yamada, M.; Matsunaga, S.; Fusetani, N.; Yamazaki, F. l-Kynurenine, an amino acid identified as a sex pheromone in the urine of ovulated female masu salmon. Proc. Natl. Acad. Sci. USA 2006, 103, 15370–15374. [Google Scholar]
- Prince, E.K.; Poulson, K.L.; Myers, T.L.; Sieg, R.D.; Kubanek, J. Characterization of allelopathic compounds from the red tide dinoflagellate. Karenia Brevis. Harmful Algae 2010, 10, 39–48. [Google Scholar] [CrossRef]
- Zimmer, R.K.; Zimmer, C.A. Dynamic scaling in chemical ecology. J. Chem. Ecol. 2008, 34, 822–836. [Google Scholar] [CrossRef]
- Madaras, F.; Gerber, J.P.; Peddie, F.; Kokkinn, M.J. The effect of sampling methods on the apparent constituents of ink from the squid Sepioteuthis australis. J. Chem. Ecol. 2010, 36, 1171–1179. [Google Scholar] [CrossRef]
- Wood, J.B.; Pennoyer, K.E.; Derby, C.D. Ink is a conspecific alarm cue in the Caribbean reef squid, Sepioteuthis sepioidea. J. Exp. Mar. Biol. Ecol. 2008, 367, 11–16. [Google Scholar] [CrossRef]
- Wood, J.B.; Maynard, A.; Lawlor, A.; Sawyer, E.K.; Simmons, D.; Pennoyer, K.E.; Derby, C.D. Caribbean reef squid, Sepioteuthis sepioidea, use ink as a defense against predatory French grunts, Haemulon flavolineatum. J. Exp. Mar. Biol. Ecol. 2010, 338, 20–27. [Google Scholar]
- Liu, Y.; Simon, J.D. The effect of preparation procedures on the morphology of melanin from the ink sac of Sepia officinalis. Pigment. Cell Res. 2003, 16, 72–80. [Google Scholar] [CrossRef]
- Palumbo, A. Melanogenesis in the ink gland of Sepia officinalis. Pigment. Cell Res. 2003, 16, 517–522. [Google Scholar] [CrossRef]
- Fiore, G.; Poli, A.; di Cosmo, A.; d’Ischia, M.; Palumbo, A. Dopamine in the ink defence system of Sepia officinalis: Biosynthesis, vesicular compartmentation in mature ink gland cells, nitric oxide (NO)/cGMP-induced depletion and fate in secreted ink. Biochem. J. 2004, 378, 785–791. [Google Scholar] [CrossRef]
- Lucero, M.T.; Farrington, H.; Gilly, W.F. Quantification of l-dopa and dopamine in squid ink: Implications for chemoreception. Biol. Bull. 1994, 187, 55–63. [Google Scholar] [CrossRef]
- Naraoka, T.; Chung, H.-S.; Uchisawa, H.; Sasaki, J.; Matsue, H. Tyrosinase activity in antitumor compounds of squid ink. Food Sci. Technol. Res. 2000, 6, 171–175. [Google Scholar] [CrossRef]
- Prota, G. Melanins, melanogenesis and melanocytes: Looking at their functional significance from a chemist’s viewpoint. Pigment. Cell Res. 2000, 13, 283–293. [Google Scholar]
- Prota, G.; Ortonne, J.P.; Voulot, C.; Khatchadourian, C.; Mardi, G.; Palumbo, A. Occurrence and properties of tyrosinase in the ejected ink of cephalopods. Comp. Biochem. Physiol. B 1981, 68, 415–419. [Google Scholar]
- Wang, F.R.; Xie, Z.G.; Ye, X.Q.; Deng, S.G.; Hu, Y.Q.; Guo, X.; Chen, S.G. Effectiveness of treatment of iron deficiency anemia in rats with squid ink melanin-Fe. Food Funct. 2014, 5, 123–128. [Google Scholar] [CrossRef]
- Pezzella, A.; d’Ischia, M.; Napolitano, A.; Palumbo, A.; Prota, G. An integrated approach to the structure of Sepia melanin. Evidence for a high proportion of degraded 5,6-dihydroxyindole-2-carboxylic acid units in the pigment backbone. Tetrahedron 1997, 53, 8281–8286. [Google Scholar] [CrossRef]
- Pezzella, A.; Napolitano, A.; d’Ischia, M.; Prota, G.; Seraglia, R.; Traldi, P. Identification of partially degraded oligomer of 5,6-dihydroxyindole-2-carboxylic acid in Sepia melanin by matrix-assisted laser desorption/ionization mass spectrometry. Rapid Comm. Mass Spectrom. 1997, 11, 368–372. [Google Scholar] [CrossRef]
- Clancy, C.M.; Simon, J.D. Ultrastructural organization of eumelanin from Sepia officinalis measured by atomic force microscopy. Biochemistry 2001, 40, 13353–13360. [Google Scholar]
- Matsuura, T.; Hino, M.; Akutagawa, S.; Shimoyama, Y.; Kobayashi, T.; Taya, Y.; Ueno, T. Optical and paramagnetic properties of size-controlled ink particles isolated from Sepia officinalis. Biosci. Biotechnol. Biochem. 2009, 73, 2790–2792. [Google Scholar] [CrossRef]
- Ortonne, J.P.; Voulot, C.; Khatchadourian, C.; Palumbo, A.; Prota, G. A reexamination of melanogenesis in the ink gland of cephalopds. In Phenotypic Expression of Pigment Cells; Seiji, M., Ed.; University of Tokyo Press: Tokyo, Japan, 1981; pp. 49–57. [Google Scholar]
- Schraermeyer, U. Fine structure of melanogenesis in the ink sac of Sepia officinalis. Pigment. Cell Res. 1994, 7, 52–60. [Google Scholar] [CrossRef]
- Fiore, G.; Mattiello, T.; Tedeschi, G.; Nonnis, S.; d’Ischia, M.; Palumbo, A. Protein nitration is specifically associated with melanin production and reveals redox imbalance as a new correlate of cell maturation in the ink gland of Sepia officinalis. Pigment. Cell Melanoma Res. 2009, 22, 857–859. [Google Scholar] [CrossRef]
- Palumbo, A.; Di Cosmo, A.; Gesualdo, I.; Hearing, V.J. Subcellular localization and function of melanogenic enzymes in the ink gland of Sepia officinalis. Biochem. J. 1997, 323, 749–756. [Google Scholar]
- Palumbo, A.; di Cosmo, A.; Gesualdo, I.; d’Ischia, M. A calcium-dependent nitric oxide synthase and NMDA R1 glutamate receptor in the ink gland of Sepia officinalis: A hint to a regulatory role of nitric oxide in melanogenesis? Biochem. Biophys. Res. Commun. 1997, 235, 429–432. [Google Scholar] [CrossRef]
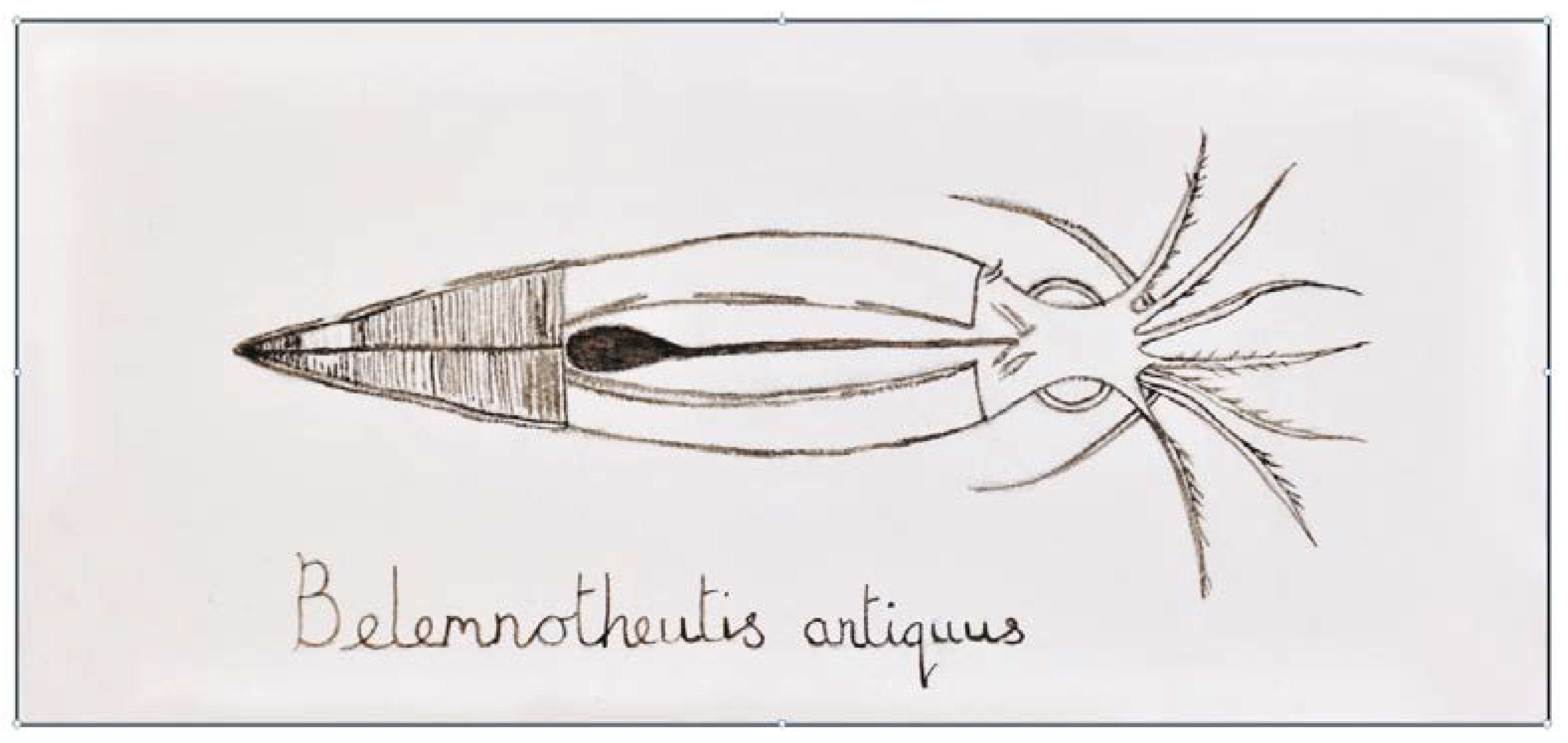
- Glass, K.; Shosuke, I.; Wilby, P.R.; Sota, T.; Nakamura, A.; Bowers, C.R.; Vinther, J.; Dutta, S.; Summons, R.; Briggs, D.E.G.; et al. Direct chemical evidence for eumelanin pigment from the Jurassic period. Proc. Natl. Acad. Sci. USA 2012, 109, 10218–10223. [Google Scholar] [CrossRef]
- Palumbo, A.; d’Ischia, M.; Misuraca, G.; de Martino, L.; Prota, G. A new dopachrome-rearranging enzyme from the ejected ink of the cuttlefish Sepia officinalis. Biochem. J. 1994, 299, 839–844. [Google Scholar]
- Shibata, T.; Prota, G.; Mishima, Y. Non-melanosomal regulatory factors in melanogenesis. J. Investig. Dermatol. 1993, 100, 274S–280S. [Google Scholar]
- D’ISCHIA, M.; Napolitano, A.; Prota, G. Peroxidase as an alternative to tyrosinase in the oxidative polymerization of 5,6-dihydroxyindoles to melanin(s). Biochim. Biophys. Acta 1991, 1073, 423–430. [Google Scholar] [CrossRef]
- Gesualdo, I.; Aniello, F.; Branno, M.; Palumbo, A. Molecular cloning of a peroxidase mRNA specifically expressed in the ink gland of Sepia officinalis. Biochim. Biophys. Acta 1997, 1353, 111–117. [Google Scholar] [CrossRef]
- Russo, G.L.; de Nisco, E.; Fiore, G.; di Donato, P.; d’Ischia, M.; Palumbo, A. Toxicity of melanin-free ink of Sepia officinalis to transformed cell lines: Identification of the active factor as tyrosinase. Biochem. Biophys. Res. Commun. 2003, 308, 293–299. [Google Scholar] [CrossRef]
- Siuda, J.F. Chemical defense mechanisms of marine organisms. Identification of 8-hydroxy-4-quinolone from the ink of the giant octopus, Octopus dofleini martini. Lloydia 1974, 37, 501–503. [Google Scholar]
- Takaya, Y.; Uchisawa, H.; Hanamatsu, K.; Narumi, F.; Okuzaki, B.; Matsue, H. Novel fucose-rich glycosaminoglycans from squid ink bearing repeating unit of trisaccharide structure (-6GalNAcα1-3GlcAβ1-3Fucα1-)n. Biochem. Biophys. Res. Commun. 1994, 198, 560–567. [Google Scholar]
- Takaya, Y.; Uchisawa, H.; Matsue, H.; Okuzaki, B.-I.; Narumi, F.; Sasaki, J.-I.; Ishida, K. An investigation of the antitumor peptidoglycan fraction from squid ink. Biol. Pharm. Bull. 1994, 17, 846–849. [Google Scholar] [CrossRef]
- Takaya, Y.; Uchisawa, H.; Narumi, F.; Matsue, H. Illexins A, B, and C from squid ink should have a branched structure. Biochem. Biophys. Res. Commun. 1996, 226, 335–338. [Google Scholar] [CrossRef]
- Chen, S.; Xu, J.; Xue, C.; Dong, P.; Sheng, W.; Yu, G.; Chai, W. Sequence determination of a non-sulfated glycosaminoglycan-like polysaccharide from melanin-free ink of the squid Ommastrephes bartrami by negative-ion electrospray tandem mass spectrometry and NMR spectroscopy. Glycoconj. J. 2008, 25, 481–492. [Google Scholar] [CrossRef]
- Sasaki, J.; Ishita, K.; Takaya, Y.; Uchisawa, H.; Matsue, H. Anti-tumor activity of squid ink. J. Nutr. Sci. Vitaminol. 1997, 43, 455–461. [Google Scholar]
- Wang, S.; Cheng, Y.; Wang, F.; Sun, L.; Liu, C.; Chen, G.; Li, Y.; Ward, S.G.; Qu, X. Inhibition activity of sulfated polysaccharide of Sepiella maindroni ink on matrix metalloproteinase (MMP)-2. Biomed. Pharmacother. 2008, 62, 297–302. [Google Scholar] [CrossRef]
- Liu, C.; Li, X.; Li, Y.; Feng, Y.; Zhou, S.; Wang, F. Structural characterisation and antimutagenic activity of a novel polysaccharide isolated from Sepiella maindroni ink. Food Chem. 2008, 110, 807–813. [Google Scholar]
- Zong, A.; Zhao, T.; Zhang, Y.; Song, X.; Shi, Y.; Cao, H.; Liu, C.; Cheng, Y.; Qu, X.; Cao, J.; et al. Anti-metastatic and anti-angiogenic activities of sufated polysaccharide of Sepiella maindroni ink. Carbohydr. Polym. 2013, 91, 403–409. [Google Scholar] [CrossRef]
- Ding, G.F.; Huang, F.F.; Yang, Z.S.; Yu, D.; Yang, Y.F. Anticancer activity of an oligopeptide isolated from hydrolysates of Sepia ink. Chin. J. Nat. Med. 2011, 9, 51–55. [Google Scholar]
- Huang, F.; Yang, Z.; Yu, D.; Wang, J.; Li, R.; Ding, G. Sepia ink oligopeptide induces apoptosis in prostate cancer cell lines via caspase-3 activation and elevation of Bax/Bcl-2 ratio. Mar. Drugs 2012, 10, 2153–2165. [Google Scholar] [CrossRef]
- Derby, C.D.; Kicklighter, C.E.; Johnson, P.M.; Zhang, X. Chemical composition of inks of diverse marine molluscs suggests convergent chemical defenses. J. Chem. Ecol. 2007, 33, 1105–1113. [Google Scholar] [CrossRef]
- Shirai, T.; Kikuchi, T.; Matsuo, S.; Inada, H.; Suzuki, T.; Hirano, T. Extractive components of the squid ink. Fish. Sci. 1997, 63, 939–944. [Google Scholar] [CrossRef]
- Palumbo, A.; Misuraca, G.; d’Ischia, M.; Prota, G. Effect of metal ions on the kinetics of tyrosine oxidation catalysed by tyrosinase. Biochem. J. 1985, 228, 647–651. [Google Scholar]
- Palumbo, A.; d’Ischia, M.; Misuraca, G.; Prota, G. Effect of metal ions on the rearrangement of dopachrome. Biochim. Biophys. Acta 1987, 925, 203–209. [Google Scholar] [CrossRef]
- Palumbo, A.; d’Ischia, M.; Misuraca, G.; Prota, G.; Schultz, T.M. Structural modifications in biosynthetic melanins induced by metal ions. Biochim. Biophys. Acta 1988, 964, 193–199. [Google Scholar] [CrossRef]
- Hong, L.; Simon, J.D. Insight into the binding of divalent cations to Sepia eumelanin from IR absorption spectroscopy. Photochem. Photobiol. 2006, 82, 1265–1269. [Google Scholar] [CrossRef]
- Raimundo, J.; Vale, C. Partitioning of Fe, Cu, Zn, Cd, and Pb concentrations among eleven tissues of Octopus vulgaris from the Portuguese coast. Cienc. Mar. 2008, 34, 297–305. [Google Scholar]
- Ichihashi, H.; Kohno, H.; Kannan, K.; Tsumura, A.; Yamasaki, S.I. Multielemental analysis of purpleback flying squid using high resolution inductively coupled plasma-mass spectrometry (HR ICP-MS). Environ. Sci. Technol. 2001, 35, 3103–3108. [Google Scholar] [CrossRef]
- Williams, B.L.; Caldwell, R.L. Intra-organismal distribution of tetrodotoxin in two species of blue-ringed octopuses (Hapalochlaena fasciata and H. lunulata). Toxicon 2009, 54, 345–353. [Google Scholar] [CrossRef]
- Williams, B.L. Behavioral and chemical ecology of marine organisms with respect to tetrodotoxin. Mar. Drugs 2010, 8, 381–398. [Google Scholar] [CrossRef]
- Costa, P.R.; Rosa, R.; Sampayo, M.A.M. Tissue distribution of the amnesic shellfish toxin, domoic acid, in Octopus vulgaris from the Portuguese coast. Mar. Biol. 2004, 144, 971–976. [Google Scholar] [CrossRef]
- Norman, M.D. Cephalopods. In A World Guide; ConchBooks: Hackenheim, Germany, 2000. [Google Scholar]
- Caldwell, R.L. An observation of inking behavior protecting adult Octopus bocki from predation by green turtles (Chelonia mydas) hatchlings. Pac. Sci. 2005, 59, 69–72. [Google Scholar] [CrossRef]
- Staudinger, M.D.; Hanlon, D.W.; Juanes, F. Primary and secondary defences of squid to cruising and ambush fish predators: Variable tactics and their survival value. Anim. Behav. 2011, 81, 585–594. [Google Scholar] [CrossRef]
- Derby, C.D.; Tottempudi, M.; Love-Chezem, T.; Wolfe, L.S. Ink from longfin inshore squid, Doryteuthis pealeii, as a chemical and visual defense against two predatory fishes, summer flounder, Paralichthys dentatus, and sea catfish, Ariopsis feils. Biol. Bull. 2013, 225, 152–160. [Google Scholar]
- Eibl-Eibesfeldt, I.; Scheer, G. Das Brutpflegeverhalten eines weiblichen Octopus aegina Gray. Z. Tierpsychol. 1962, 19, 257–261. [Google Scholar] [CrossRef]
- Guerra, A.; Gonzalez, J.L. First record of predation by a tompot blenny on the common cuttlefish Sepia officinalis eggs. Vie Milieu 2011, 61, 45–48. [Google Scholar]
- Eisner, T.; Grant, R.P. Toxicity, odor adversion, and “olfactory aposematism”. Science 1981, 213, 476. [Google Scholar] [CrossRef]
- Pasteels, J.M.; Grégoire, J.C.; Rowell-Rahier, M. The chemical ecology of defense in arthropods. Annu. Rev. Entomol. 1983, 28, 263–289. [Google Scholar] [CrossRef]
- Mithöfer, A.; Boland, W. Plant defense against herbivores: Chemical aspects. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2012, 63, 431–450. [Google Scholar] [CrossRef]
- MacGinitie, G.E.; MacGinitie, N. Natural History of Marine Animals; McGraw-Hill: New York, NY, USA, 1968. [Google Scholar]
- Fox, D.L. Biochromes: Occurrence, distribution and comparative biochemistry of prominent natural pigments in the marine world. In Biochemical and Biophysical Perspectives in Marine Biology; Malins, D.C., Sargent, J.R., Eds.; Academic Press: New York, NY, USA, 1974; Volume 1, pp. 169–211. [Google Scholar]
- Kittredge, J.S.; Takahashi, F.T.; Lindsey, J.; Lasker, R. Chemical signals in the sea: Marine allelochemics and evolution. Fish. Bull. 1974, 72, 1–11. [Google Scholar]
- Moynihan, M.; Rodaniche, A.F. The behavior and natural history of the Caribbean reef squid Sepioteuthis sepioidea. Adv. Ethol. 1982, 25, 1–151. [Google Scholar]
- Derby, C.D. Escape by inking and secreting: Marine molluscs avoid predators through a rich array of chemicals and mechanisms. Biol. Bull. 2007, 213, 274–289. [Google Scholar] [CrossRef]
- Kicklighter, C.E.; Shabani, S.; Johnson, P.M.; Derby, C.D. Sea hares use novel antipredatory chemical defenses. Curr. Biol. 2005, 15, 549–554. [Google Scholar] [CrossRef]
- Eisner, T.; Eisner, M.; Siegler, M. Secret Weapons: Defenses of Insects, Spiders, Scorpions, and Other Many-Legged Creatures; Harvard University Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Grüninger, T. The Predator-Prey Relationship between the Californian Moray Eel (Gymnothorax mordax) and the Two-Spotted Octopus (Octopus bimaculoides). Master’s Thesis, University of San Diego, San Diego, CA, USA, 1997. [Google Scholar]
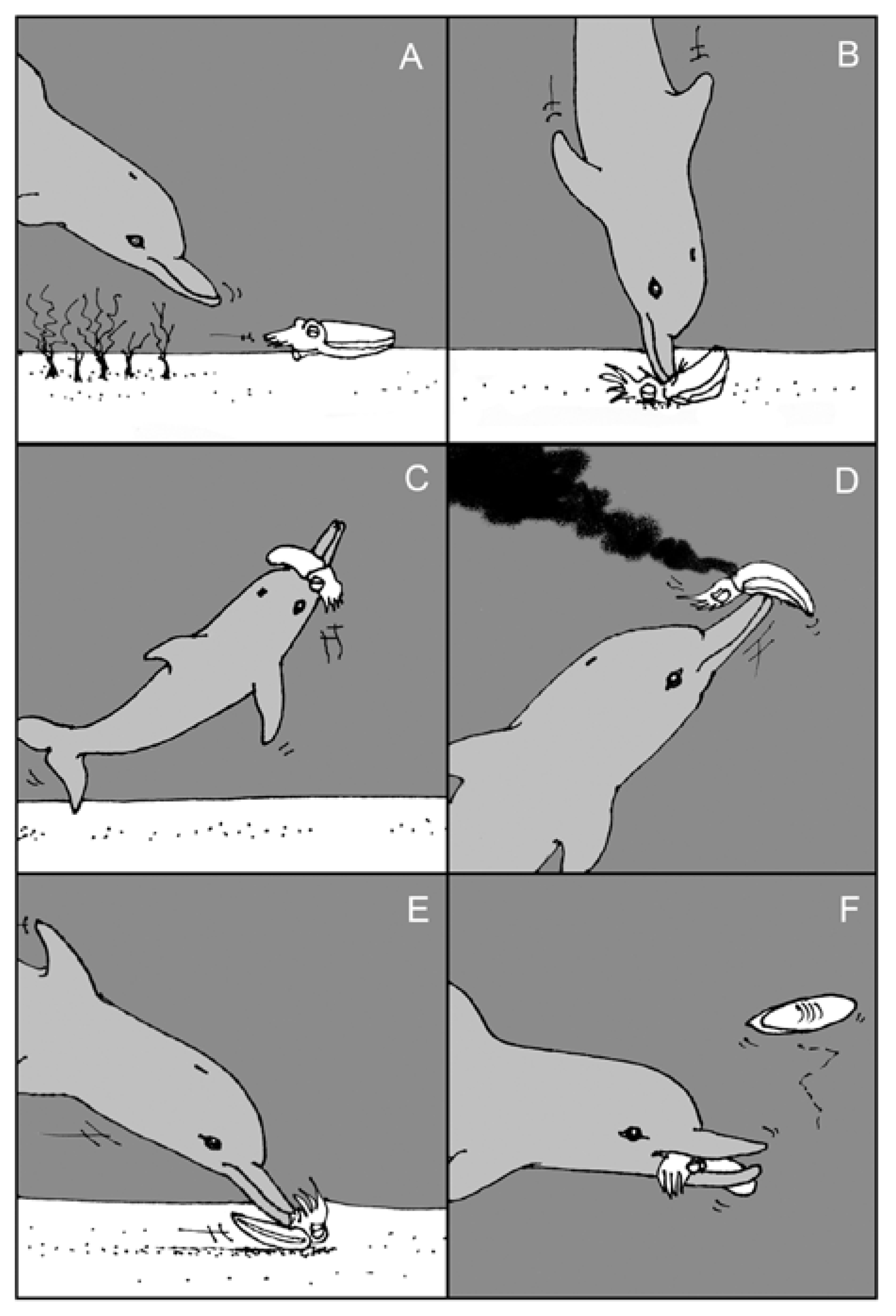
- Finn, J.; Tregenza, T.; Norman, M. Preparing the perfect cuttlefish meal: Complex prey handling by dolphins. PLoS One 2009, 4, e4217. [Google Scholar] [CrossRef]
- Eisner, T.; Meinwald, J. Defensive secretions of arthropods. Science 1966, 153, 1341–1350. [Google Scholar]
- Langley, W.M. The effect of prey defense on the attack behavior of the southern grasshopper mouse (Onychomys torridus). Z. Tierpsychol. 1981, 56, 115–127. [Google Scholar]
- Gilly, W.F.; Lucero, M.T. Behavioral responses to chemical stimulation of the olfactory organ in the squid Loligo opalescens. J. Exp. Biol. 1992, 162, 209–229. [Google Scholar]
- Seagle, B.-L.; Gasyna, E.M.; Mieler, W.F.; Norris, J.R., Jr. Photoprotection of human retinal pigment epithelium cells against blue light-induced apoptosis by melanin free radicals from Sepia officinalis. Proc. Natl. Acad. Sci. USA 2006, 103, 16644–16648. [Google Scholar] [CrossRef]
- Kamio, M.; Grimes, T.V.; Hutchins, M.H.; van Dam, R.; Derby, C.D. The purple pigment aplysioviolin in sea hare ink deters predatory blue crabs through their chemical senses. Anim. Behav. 2010, 80, 89–100. [Google Scholar] [CrossRef]
- Nair, J.R.; Pillai, D.; Joseph, S.M.; Gomathi, P.; Senan, P.V.; Sherief, P.M. Cephalopod research and bioactive substances. Indian J. Geo-Mar. Sci. 2011, 40, 13–27. [Google Scholar]
- Zhong, J.P.; Wang, G.; Shang, J.H.; Pan, J.Q.; Li, K.; Huang, Y.; Liu, H.Z. Protective effects of squid ink extract towards hemopoietic injuries induced by cyclophosphamine. Mar. Drugs 2009, 7, 9–18. [Google Scholar] [CrossRef]
- Mochizuki, A. An antiseptic effect of cuttlefish Sepioteuthis lessoniana ink. Nippon. Suisan Gakkaishi 1979, 45, 1401–1404. [Google Scholar] [CrossRef]
- Sheu, T.-Y.; Chou, C.C. Antimicrobial activity of squid ink. J. Chin. Agric. Chem. Soc. 1990, 28, 59–68. [Google Scholar]
- Takai, M.; Yamazaki, K.; Kawai, Y.; Inoue, N.; Shinano, H. Effects of squid liver, skin, and ink on chemical characteristics of “ika-shiokara” during ripening process. Bull. Jap. Soc. Sci. Fish. 1993, 59, 1609–1615. [Google Scholar] [CrossRef]
- Nirmale, V.; Nayak, B.B.; Kannappan, S.; Basu, S. Antibacterial effect of the Indian squid, Loligo duvauceli (d’Orbigny) ink. J. Indian Fish. Assoc. 2002, 29, 65–69. [Google Scholar]
- Chacko, D.; Patterson, J. Effect of Pharaoh cuttlefish, Sepia pharaonis ink against bacterial pathogens. Indian J. Microbiol. 2005, 45, 227–230. [Google Scholar]
- Vennila, R.; Rajesh, R.K.; Kanchana, S.; Arumugam, M.; Balasubramanian, T. Investigation of antimicrobial and plasma coagulation property of some molluscan ink extracts: Gastropods and cephalopods. Afr. J. Biochem. Res. 2011, 5, 14–21. [Google Scholar]
- Patterson, J.K.; Murugan, E.; Murugan, A. Screening of cephalopds for bio-activity. Spec. Publ. Phuket Mar. Biol. Cent. 2000, 21, 253–256. [Google Scholar]
- Nithya, M.; Ambikapathy, V.; Panneerselvam, A. Effect of pharaoh’s cuttlefish ink against bacterial pathogens. Asian J. Plant Sci. Res. 2011, 1, 49–55. [Google Scholar]
- Girija, S.A.S.; Priyadharshini, V.J.; Suba, P.K.; Hariprasad, P.; Raguraman, R. Antibacterial effect of squid ink on ESBL producing strains of Escherichia coli and Klebsiella pneumoniae. Indian J. Geo-Mar. Sci. 2012, 41, 338–343. [Google Scholar]
- Lin, L.-C.; Chen, W.-T. Antimicrobial and photoprotective properties of melanin extracted from various sources of animal. Taiwan. J. Agric. Chem. Food Sci. 2004, 42, 315–320. [Google Scholar]
- Vega Petkovic, M. Determination of the antimicrobial activity of purified melanin from the ink of Octopus mimus Gould, 1852 (Cephalopoda: Octopodidae). Lat. Amer. J. Aquat. Res. 2013, 41, 584–587. [Google Scholar]
- Fahmy, S.R.; Soliman, A.M. In vitro antioxidant, analgesic and cytotoxic activities of Sepia officinalis ink and Coelatura aegyptiaca extracts. Afr. J. Pharm. Pharmacol. 2013, 7, 1512–1522. [Google Scholar] [CrossRef]
- Roten, C.-A.H.; Karamata, D. Endogenous synthesis of peptidoglycan in eukaryotic cells; A novel concept involving its essential role in cell division, tumor formation and the biological clock. Experientia 1992, 48, 921–931. [Google Scholar] [CrossRef]
- Priya, S.V.; George, M.C.; Joseph, S.M.; Devika, P.; Nair, J.R.; and Sherief, P.M. Antiproliferative effect of cuttle fish ink extract on chicken embryo fibroblasts. In Proceedings of the National Seminar on New Frontiers in Marine Bioscience Research; Abidi, S.A.H., Ravindran, M., Venkatesan, R., Vijayakumaran, M., Eds.; National Institute of Ocean Technology, Chennai: Chennai, India, 2004; pp. 35–39. [Google Scholar]
- Lei, M.; Wang, J.; Wang, Y.; Pang, L.; Wang, Y.; Xu, W.; Xue, C. Study of the radio-protective effect of cuttlefish ink on hemopoietic injury. Asia Pac. J. Clin. Nutr. 2007, 16, 239–243. [Google Scholar]
- Kim, S.-Y.; Kim, S.-H.; Song, K.B. Characterization of an partial purification and angiotensin-converting enzyme inhibitor from squid ink. Agric. Chem. Biotechnol. 2003, 46, 122–123. [Google Scholar]
- Rajaganapathi, J.; Thyagarajan, S.P.; Patterson Edward, J.K. Study on cephalopod’s ink for anti-retroviral activity. Indian J. Exp. Biol. 2000, 38, 519–520. [Google Scholar]
- Mimura, T.; Maeda, K.; Hariyama, H.; Aonuma, S.; Satake, M.; Fujita, T. Studies on biological activities of melanin from marine animals. I. Purification of melanin from Ommastrephes bartrami Lesuel and its inhibitory activty on gastric secretion in rats. Chem. Pharmacol. Bull. 1982, 30, 1381–1386. [Google Scholar] [CrossRef]
- Mimura, T.; Maeda, K.; Tsujibo, H.; Satake, M.; Fujita, T. Studies on biological activities of melanin from marine animals. II. Purification of melanin from Octopus vulgaris Cuvier and its inhibitory activity on gastric juice secretion in rats. Chem. Pharm. Bull. 1982, 30, 1508–1512. [Google Scholar] [CrossRef]
- Mimura, T.; Maeda, K.; Terada, T.; Oda, Y.; Morishita, K.; Aonuma, S. Studies on biological activities of melanin from marine animals. III. Inhibitory effect of SM II (low molecular weight melanoprotein from squid) on phenylbutazone-induced ulceration in gastric mucosa in rats, and its mechanism of action. Chem. Pharm. Bull. 1985, 33, 2052–2060. [Google Scholar] [CrossRef]
- Mimura, T.; Itoh, S.; Tsujikawa, K.; Nakajima, H.; Satake, M.; Kohama, Y.; Okabe, M. Studies on biological activities of melanin from marine animals. V. Anti-inflammatory activity of low-molecular-weight melanoprotein from squid (Fr. SM II). Chem. Pharm. Bull. 1987, 35, 1144–1150. [Google Scholar] [CrossRef]
- Lin, L.-C.; Chen, W.-T. The study of antioxidant effects of melanins extracted from various tissues of animals. Asian-Australas. J. Anim. Sci. 2005, 18, 277–281. [Google Scholar]
- Liu, H.; Luo, P.; Chen, S.; Shang, J. Effects of squid ink on growth performace, antioxidant functions and immunity in growing broiler chickens. Asian-Australas. J. Anim. Sci. 2011, 24, 1752–1756. [Google Scholar] [CrossRef]
- Vate, N.K.; Benjakul, S. Antioxidative activity of melanin-free ink from splendid squid (Loligo formosana). Int. Aquat. Res. 2013, 5, 9. [Google Scholar] [CrossRef]
- Neifar, A.; Ben Adelmalek, I.; Bouajila, G.; Kolsi, R.; Nejmeddine Bradai, M.; Abdelmouleh, A.; Gargourib, A.; Ayed, N. Purification and incorporation of the black ink of cuttlefish Sepia officinalis in eye cosmetic products. Color. Technol. 2013, 129, 150–154. [Google Scholar] [CrossRef]
- Honda, S.; Takekoshi, Y.; Arai, Y. Cosmetics Based on Naturally Derived Melanin-Coated Pigments. U.S. Patent 5,380,359 A, 10 January 1995. [Google Scholar]
- Cagliani, I. Consigli e suggerimenti criteri di accettabilità. Cosmet. News 1999, 124, 25–27. [Google Scholar]
- Hazan, M. Essentials of Classic Italian Cooking; Knopf Inc.: New York, NY, USA, 1992. [Google Scholar]
- Marquinet, A.; Inaki, J. Process for Producing a Food Colorant, Colorant thus Obtained and Uses Thereof. U.S. Patent 6,329,010 B1, 11 December 2001. [Google Scholar]
- Xu, H.; Gou, J.; Choi, G.-P.; Lee, H.-Y.; Ahn, J. Functional properties of squid by-products fermented by probiotic bacteria. Food Sci. Biotechnol. 2009, 18, 761–765. [Google Scholar]
- Di Cosmo, A.; Winlow, W. Neuroecology and Neuroethology in Molluscs: The Interface Between Behavior and Environment; Nova Science Publishers Ltd.: New York, NY, USA, 2014. [Google Scholar]
- Nusnbaum, M.; Derby, C.D. Ink secretion protects sea hares by acting on the olfactory and non-olfactory chemical senses of a predatory fish. Anim. Behav. 2010, 79, 1067–1076. [Google Scholar] [CrossRef]
- Derby, C.D.; Aggio, J.F. Neuroecology of chemical defenses. Integr. Comp. Biol. 2011, 51, 771–780. [Google Scholar] [CrossRef]
- Naraoka, T.; Uchisawa, H.; Mori, H.; Matsue, H.; Chiba, S.; Kimura, A. Purification, characterization and molecular cloning of tyrosinase from the cephalopod mollusk, Illex argentinus. Eur. J. Biochem. 2003, 270, 4026–4038. [Google Scholar] [CrossRef]
- Scheinker, V.; Fiore, G.; Di Cristo, C.; Di Cosmo, A.; d’Ischia, M.; Enikolopov, G.; Palumbo, A. Nitric oxide synthase in the nervous system and ink gland of the cuttlefish Sepia officinalis: Molecular cloning and expression. Biochem. Biophys. Res. Comm. 2005, 338, 1204–1215. [Google Scholar] [CrossRef]
- Song, W.-W.; Mu, C.-K.; Shao, C.; Zhang, Y.-Y.; Zhan, P.-P.; Wang, C. Expression screening and annotation of Sepiella maindroni ink sac cDNA library. J. Genet. 2012, 28, e103–e108. [Google Scholar]
- Bassetti, M.; Merelli, M.; Temperoni, C.; Astilean, A. New antibiotics for bad bugs: Where are we? Ann. Clin. Microbiol. Antimicrob. 2013, 12, 22. [Google Scholar] [CrossRef]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. New horizons for old drugs and drug leads. J. Nat. Prod. 2014, 77, 703–723. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Derby, C.D. Cephalopod Ink: Production, Chemistry, Functions and Applications. Mar. Drugs 2014, 12, 2700-2730. https://doi.org/10.3390/md12052700
Derby CD. Cephalopod Ink: Production, Chemistry, Functions and Applications. Marine Drugs. 2014; 12(5):2700-2730. https://doi.org/10.3390/md12052700
Chicago/Turabian StyleDerby, Charles D. 2014. "Cephalopod Ink: Production, Chemistry, Functions and Applications" Marine Drugs 12, no. 5: 2700-2730. https://doi.org/10.3390/md12052700
APA StyleDerby, C. D. (2014). Cephalopod Ink: Production, Chemistry, Functions and Applications. Marine Drugs, 12(5), 2700-2730. https://doi.org/10.3390/md12052700