Gram-Negative Marine Bacteria: Structural Features of Lipopolysaccharides and Their Relevance for Economically Important Diseases

Abstract
:1. Introduction
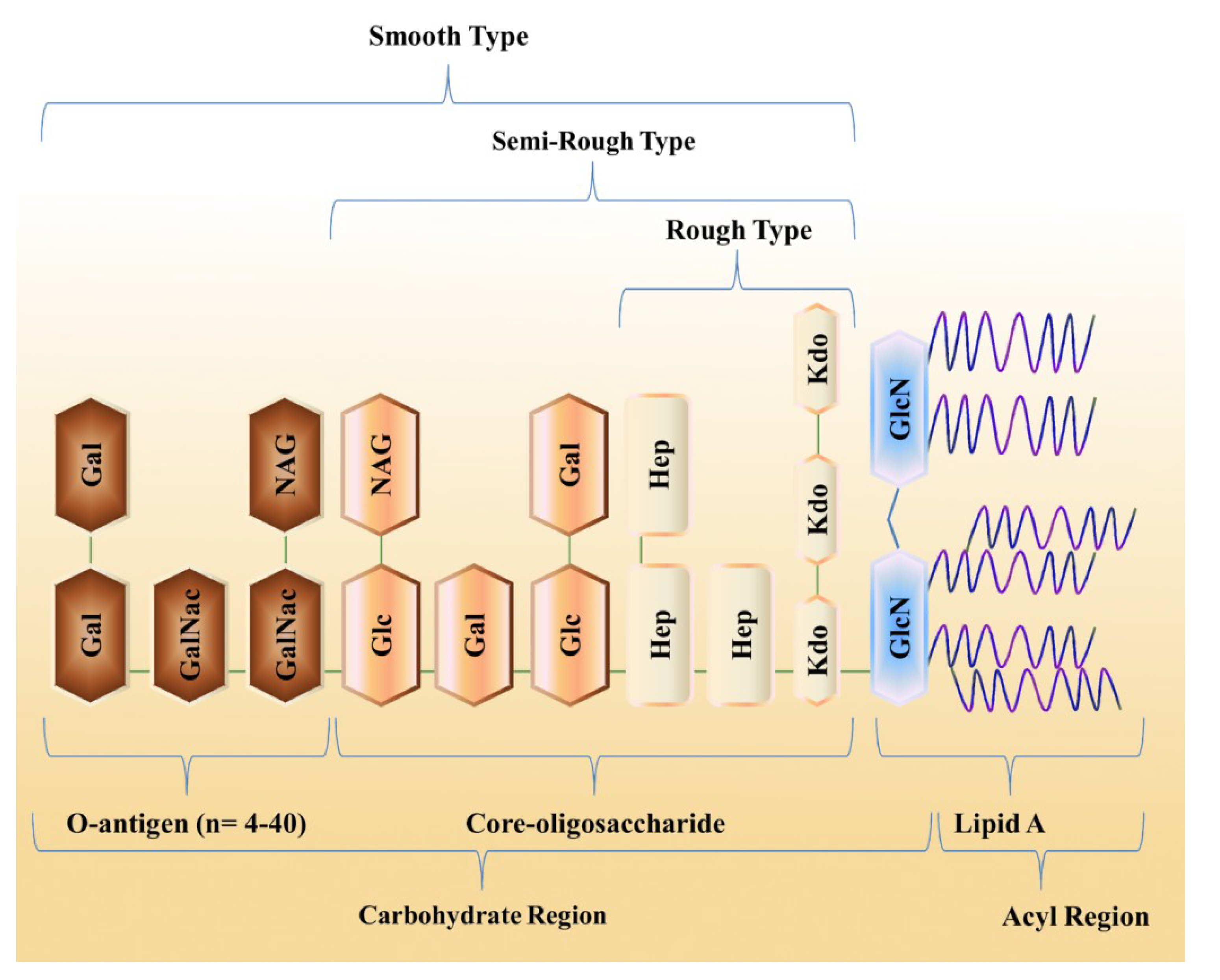
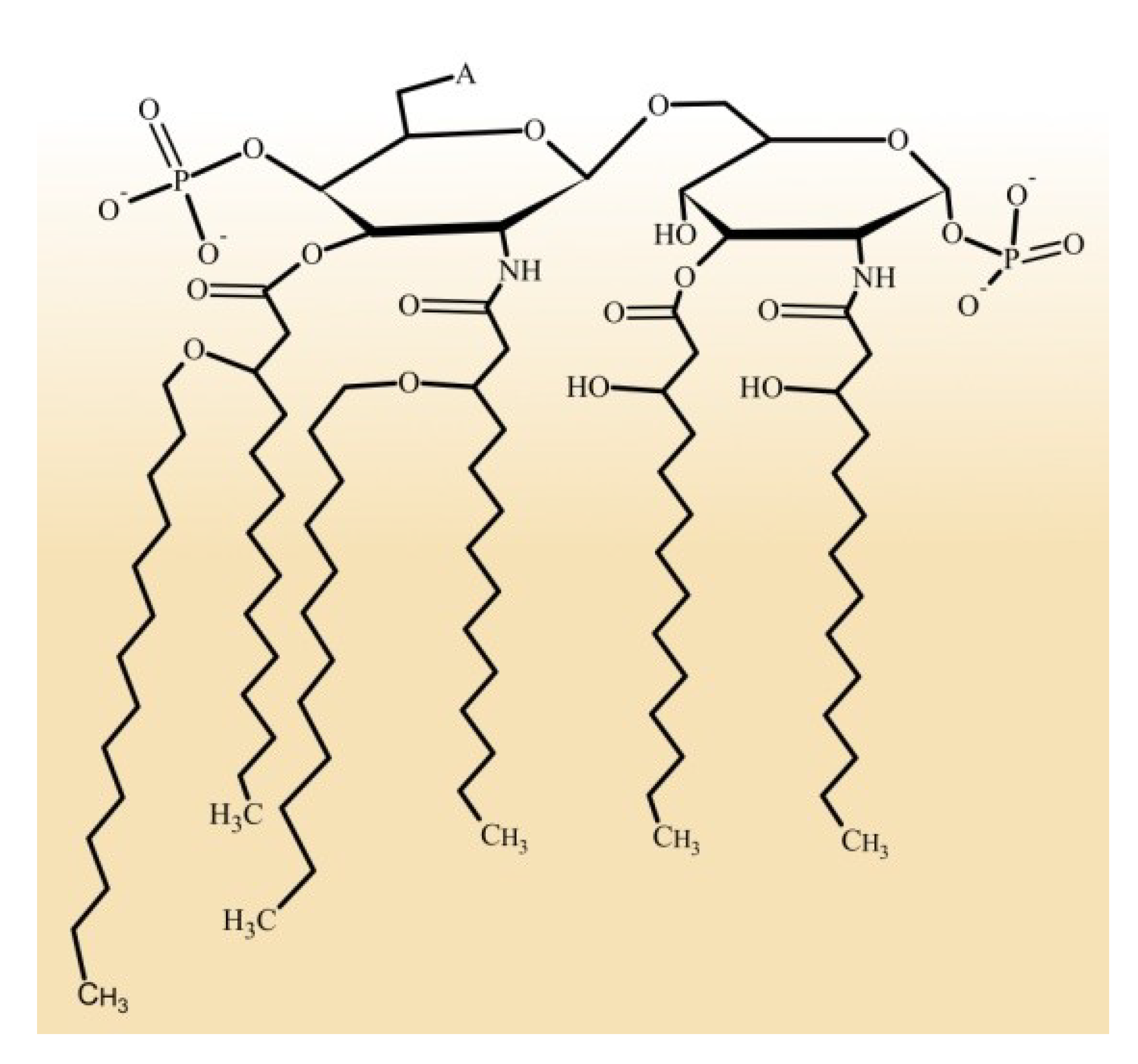
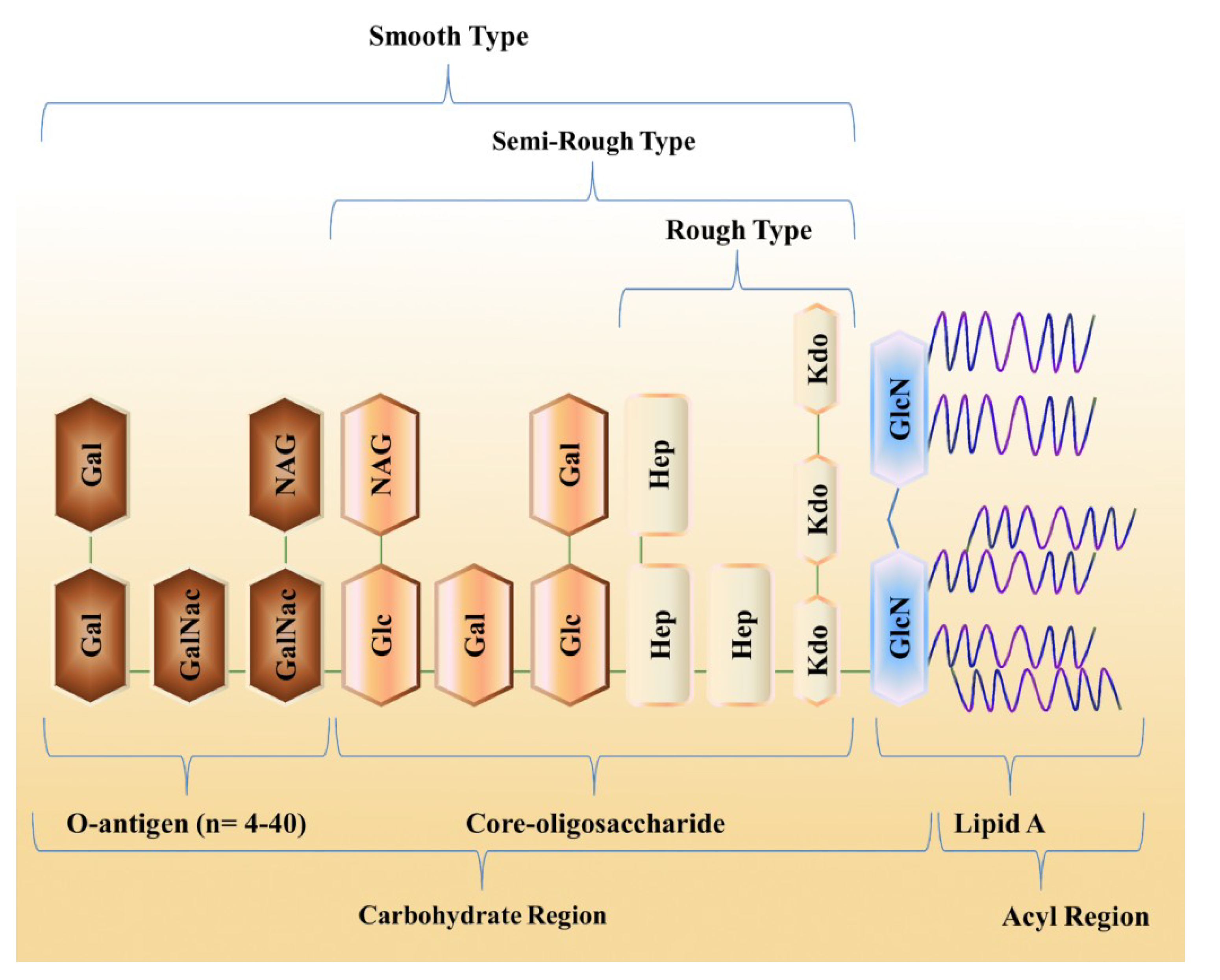
2. Ecological Diversity of Marine Bacteria
3. Structural Organization of LPS
3.1. Aeromonas

3.2. Pseudoalteromonas

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
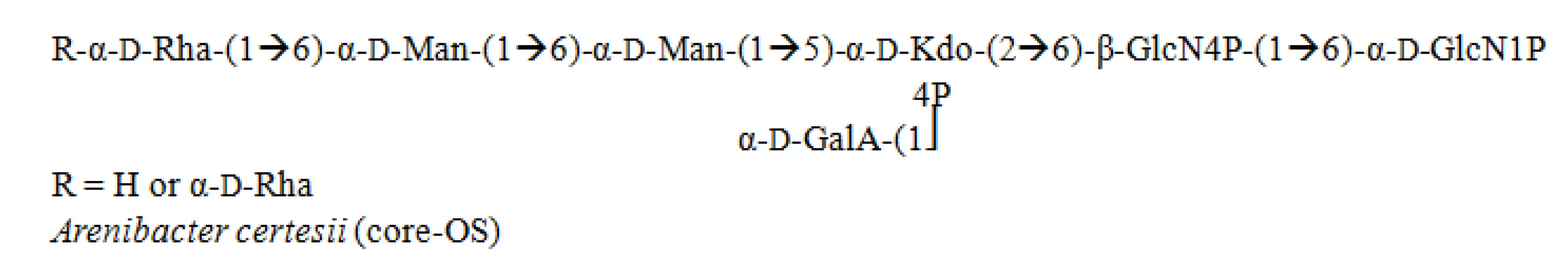{kind=link}
{kind=link}
| Bacterium | Type and linkage of acyl substituent | ||||
|---|---|---|---|---|---|
| GlcN II a | GlcN I | Reference | |||
| 3′ | 2′ | 3 | 2 | ||
| Pseudoalteromonas carrageenovora | 10:0(3-OH) b | 12:0(3-O-12:0) | - | 12:0(3-OH) | [64] |
| Pseudoalteromonas issachenkonii | 10:0(3-OH) b | 12:0(3-O-12:0) | - | 12:0(3-OH) | [73] |
| Alteromonas addita | 12:0(3-OH) | 14:0(3-O-12:0) | 10:0(3-OH) | 14:0(3-OH) | [77] |
| Alteromonas macleodii | 12:0(3-OH) b | 12:0(3-O-12:0) c | - | 12:0(3-OH) | [78] |
| Shewanella pacifica | 13:0(3-O-13:0) | 13:0(3-O-13:0) | 13:0(3-OH) | 13:0(3-OH) | [10] |
| Arenibacter certesii | 15:0(3-OH) | 15:0(3-O-15:0) | 15:0(3-OH) | 15:0(3-OH) | [11] |

3.3. Shewanella

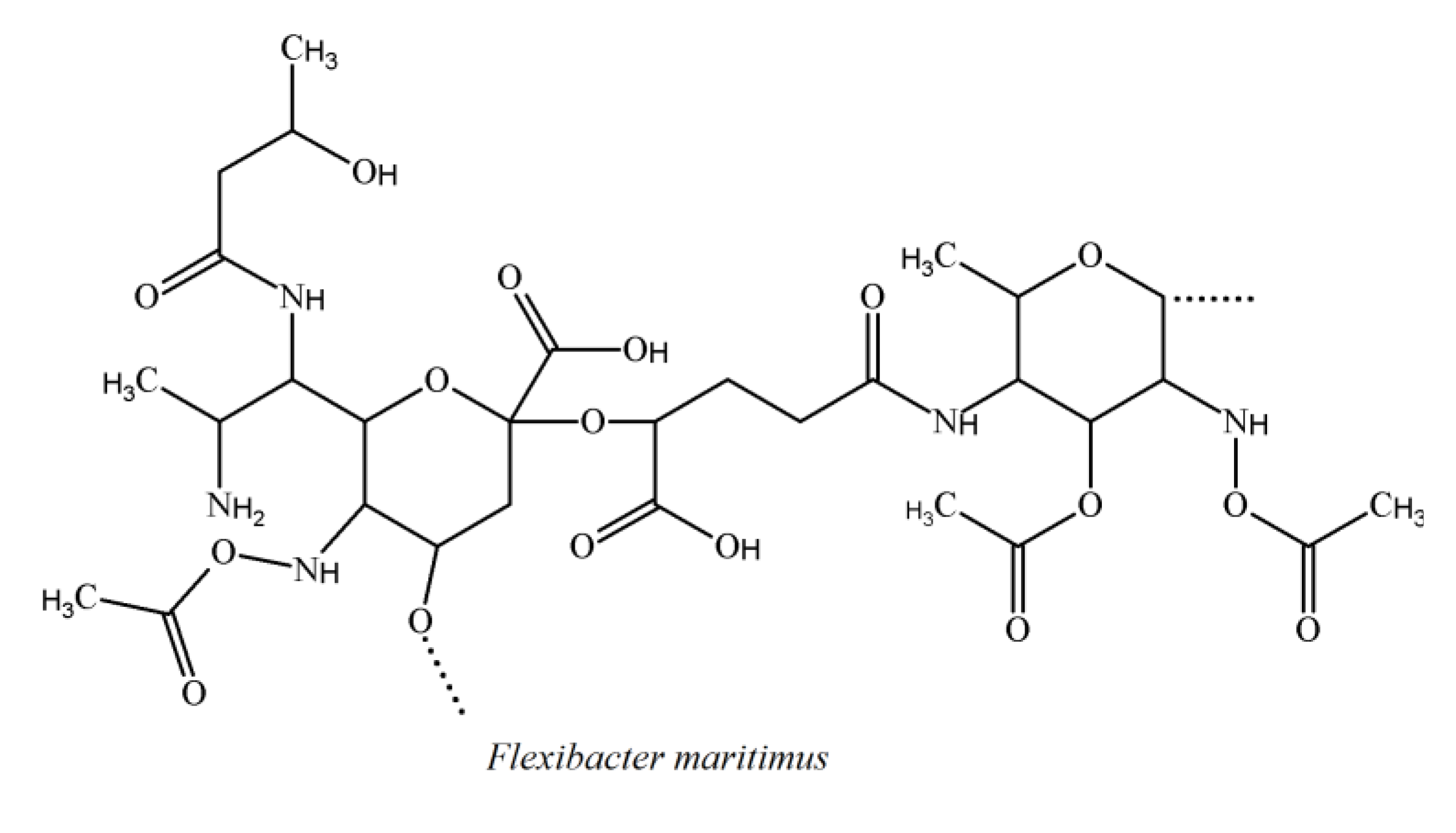
3.4. Cytophaga-Flavobacterium-Flexibacter Phylum
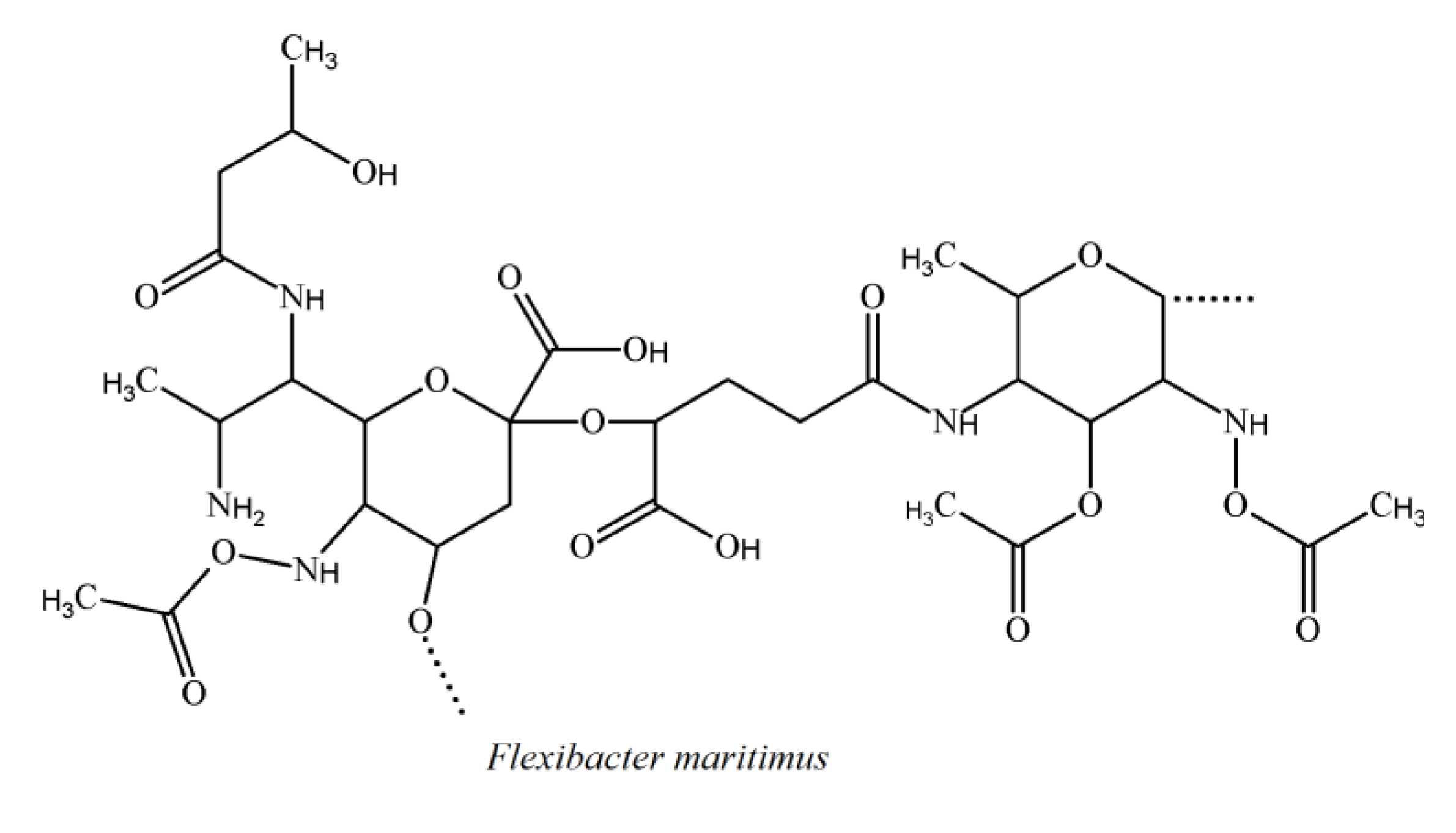
3.5. Alteromonas

3.6. Arenibacter
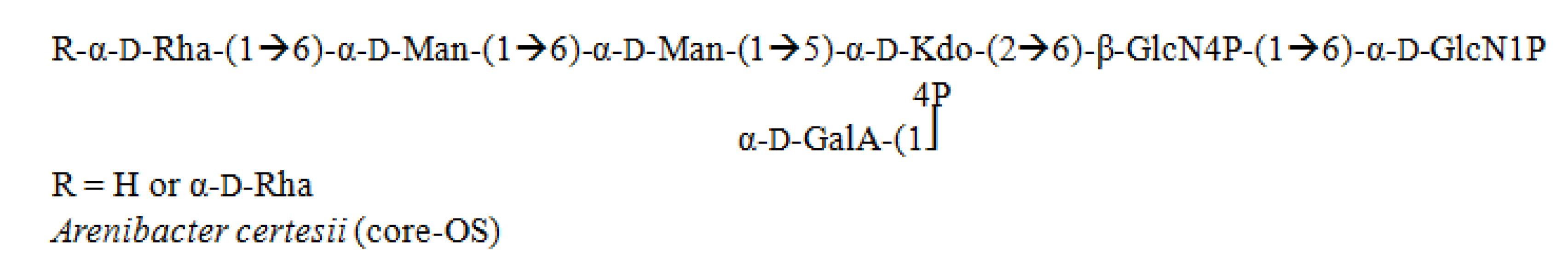
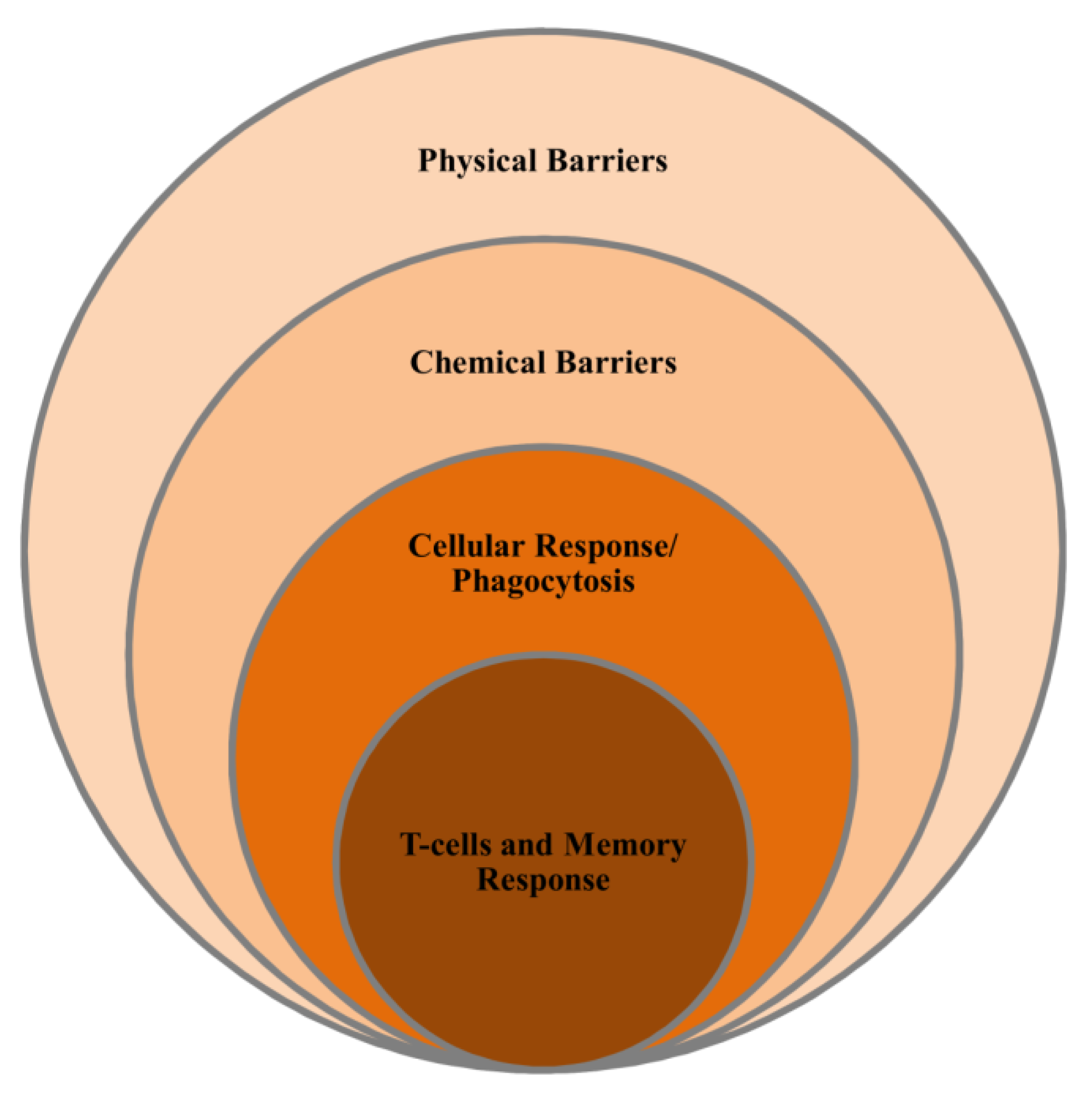
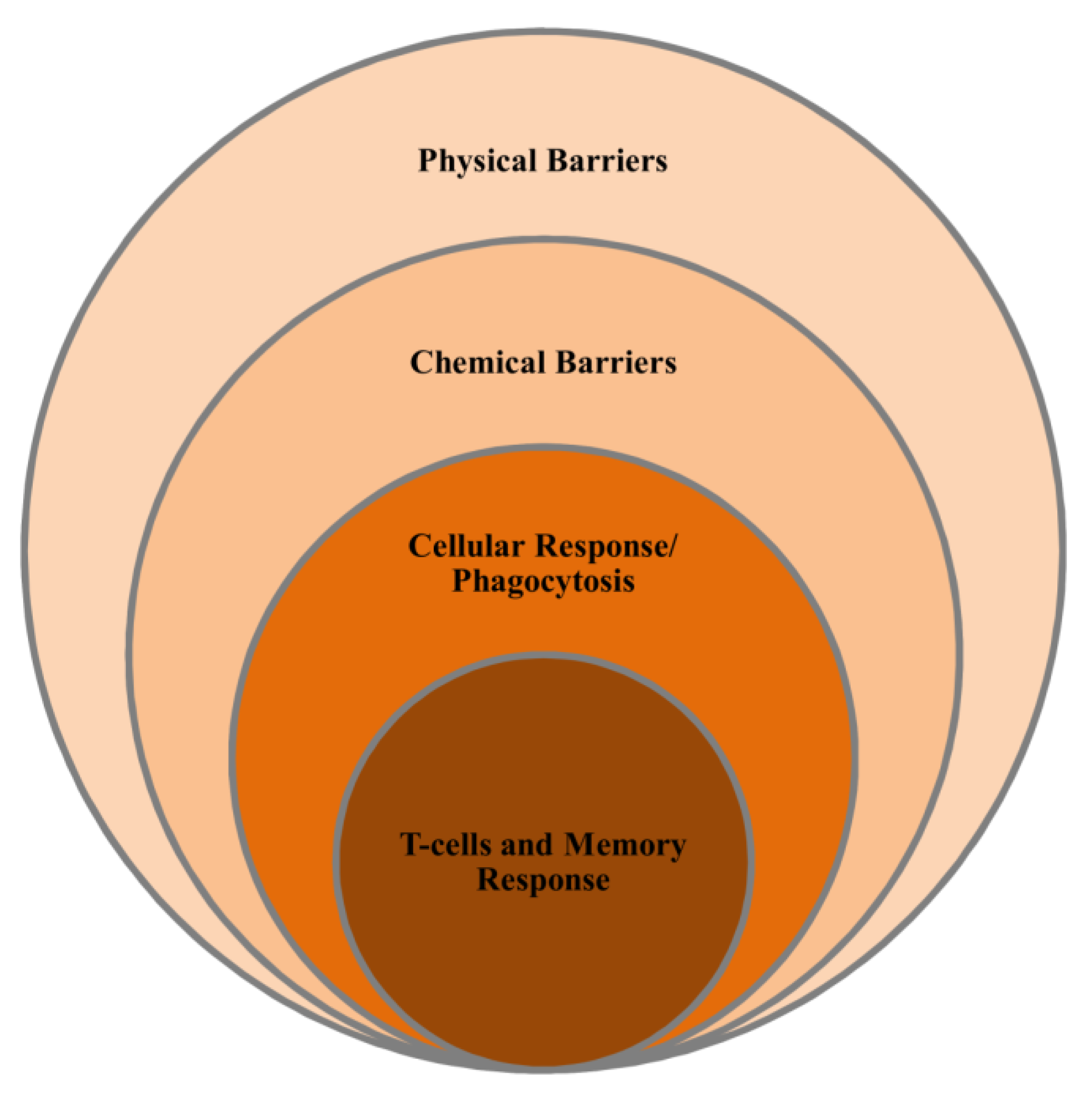
4. Marine Immune System
4.1. Non-Specific Immunity
4.2. Specific Immunity
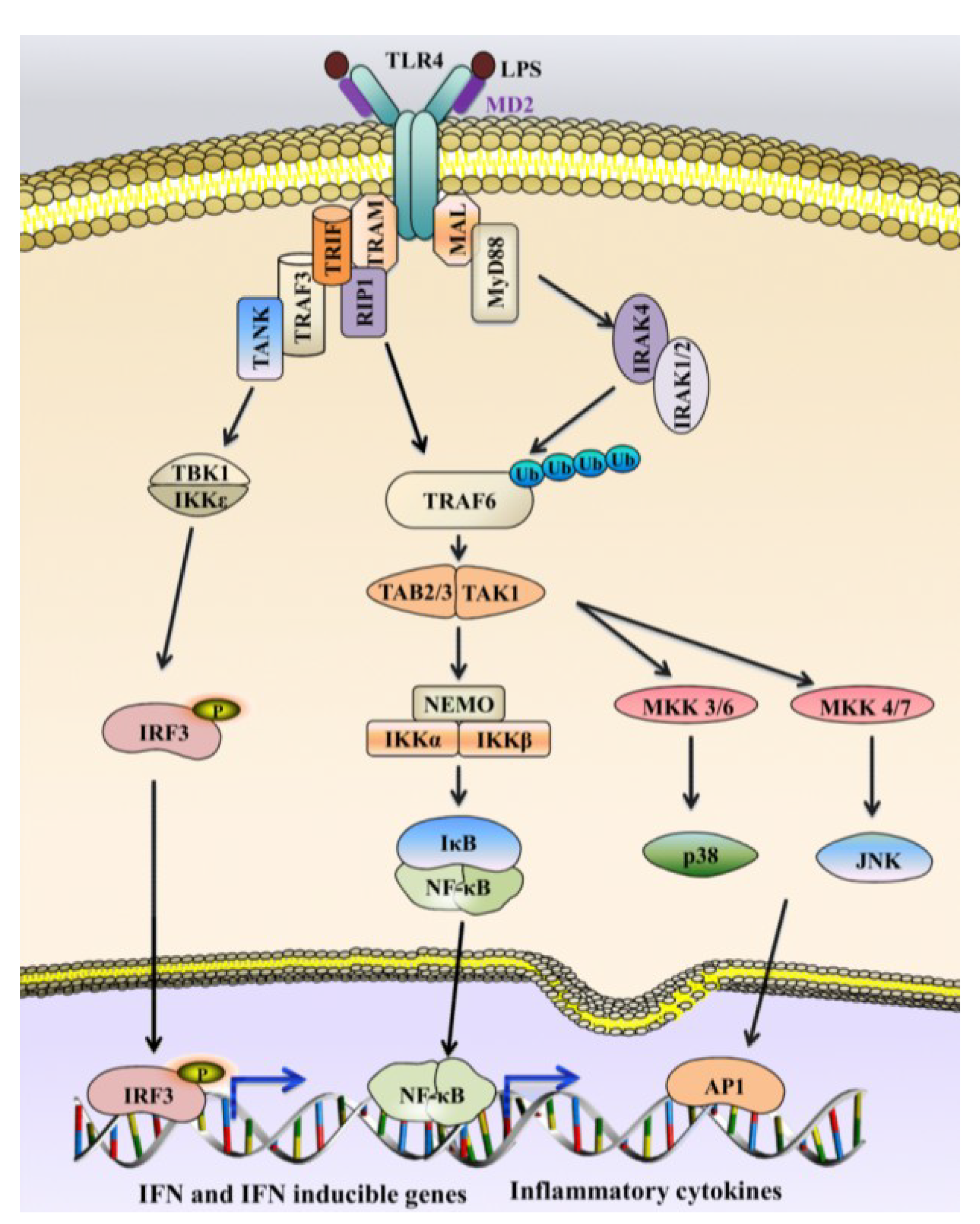
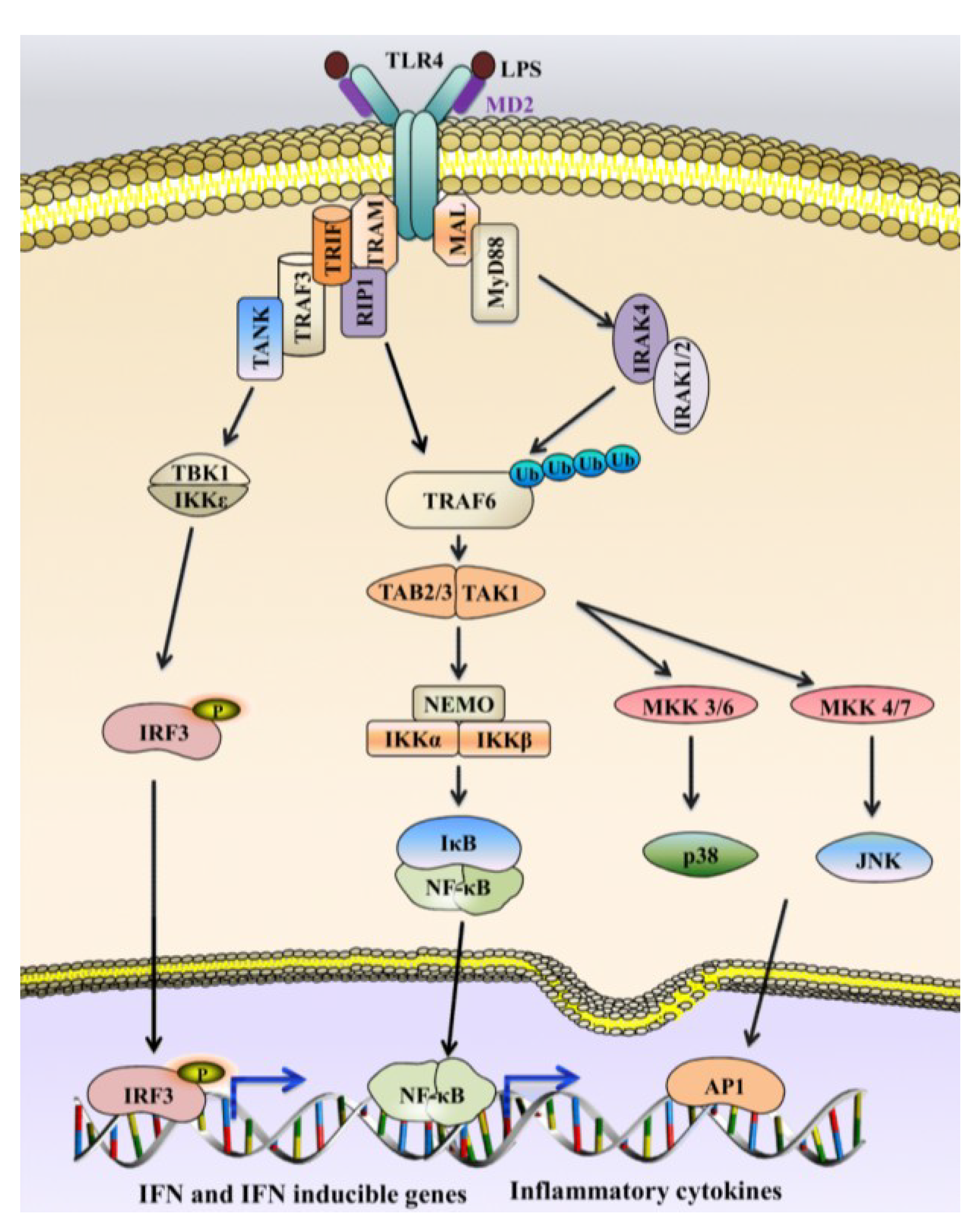
5. LPS Detection and Marine Immune Response
6. Structure-Activity Relationship of LPS
7. Marine LPS as a Drug Candidate for Human Diseases
8. Conclusions
Acknowledgments
Conflicts of Interest
References
- Head, I.M.; Jones, D.M.; Röling, W.F. Marine microorganisms make a meal of oil. Nat. Rev. Microbiol. 2006, 4, 173–182. [Google Scholar] [CrossRef]
- Hau, H.H.; Gralnick, J.A. Ecology and biotechnology of the genus shewanella. Annu. Rev. Microbiol. 2007, 61, 237–258. [Google Scholar] [CrossRef]
- Caroff, M.; Karibian, D. Structure of bacterial lipopolysaccharides. Carbohydr. Res. 2003, 338, 2431–2447. [Google Scholar] [CrossRef]
- Wilkinson, S.G. Bacterial lipopolysaccharides—Themes and variations. Prog. Lipid Res. 1996, 35, 283–343. [Google Scholar] [CrossRef]
- Jansson, P.E. The chemistry of O-polysaccharide chains in bacterial lipopolysaccharides. ChemInform 2000, 31, 155–178. [Google Scholar]
- Knirel, Y.A. O-specific Polysaccharides of Gram-negative Bacteria. In Microbial Glycobiology: Structures, Relevance and Applications; Elsevier: Amsterdam, The Netherlands, 2009; pp. 57–73. [Google Scholar]
- Holst, O. Chemical structure of the core region of lipopolysaccharides. Endotoxin Health Dis. 1999, 1, 115–154. [Google Scholar]
- Vinogradov, E.; Korenevsky, A.; Beveridge, T.J. The structure of the rough-type lipopolysaccharide from Shewanella oneidensis MR-1, containing 8-amino-8-deoxy-Kdo and an open-chain form of 2-acetamido-2-deoxy-d-galactose. Carbohydr. Res. 2003, 338, 1991–1997. [Google Scholar] [CrossRef]
- Vinogradov, E.; Korenevsky, A.; Beveridge, T.J. The structure of the core region of the lipopolysaccharide from Shewanella algae BrY, containing 8-amino-3,8-dideoxy-d-manno-oct-2-ulosonic acid. Carbohydr. Res. 2004, 339, 737–740. [Google Scholar] [CrossRef]
- Silipo, A.; Leone, S.; Molinaro, A.; Sturiale, L.; Garozzo, D.; Nazarenko, E.L.; Gorshkova, R.P.; Ivanova, E.P.; Lanzetta, R.; Parrilli, M. Complete structural elucidation of a novel lipooligosaccharide from the outer membrane of the marine bacterium Shewanella pacifica. Eur. J. Org. Chem. 2005, 2005, 2281–2291. [Google Scholar]
- Silipo, A.; Molinaro, A.; Nazarenko, E.L.; Sturiale, L.; Garozzo, D.; Gorshkova, R.P.; Nedashkovskaya, O.I.; Lanzetta, R.; Parrilli, M. Structural characterization of the carbohydrate backbone of the lipooligosaccharide of the marine bacterium Arenibacter certesii strain KMM 3941T. Carbohydr. Res. 2005, 340, 2540–2549. [Google Scholar] [CrossRef]
- Fahy, E.; Subramaniam, S.; Brown, H.A.; Glass, C.K.; Merrill, A.H., Jr.; Murphy, R.C.; Raetz, C.R.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A comprehensive classification system for lipids. J. Lipid Res. 2005, 46, 839–861. [Google Scholar] [CrossRef]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef]
- Steeghs, L.; de Cock, H.; Evers, E.; Zomer, B.; Tommassen, J.; van der Ley, P. Outer membrane composition of a lipopolysaccharide-deficient Neisseria meningitidis mutant. EMBO J. 2001, 20, 6937–6945. [Google Scholar] [CrossRef]
- Meredith, T.C.; Aggarwal, P.; Mamat, U.; Lindner, B.; Woodard, R.W. Redefining the requisite lipopolysaccharide structure in Escherichia coli. ACS Chem. Biol. 2006, 1, 33–42. [Google Scholar] [CrossRef]
- Galloway, S.M.; Raetz, C.R. A mutant of Escherichia coli defective in the first step of endotoxin biosynthesis. J. Biol. Chem. 1990, 265, 6394–6402. [Google Scholar]
- Belunis, C.J.; Clementz, T.; Carty, S.M.; Raetz, C.R. Inhibition of lipopolysaccharide biosynthesis and cell growth following inactivation of the kdta gene in Escherichia coli. J. Biol. Chem. 1995, 270, 27646–27652. [Google Scholar] [CrossRef]
- Onishi, H.R.; Pelak, B.A.; Gerckens, L.S.; Silver, L.L.; Kahan, F.M.; Chen, M.H.; Patchett, A.A.; Galloway, S.M.; Hyland, S.A.; Anderson, M.S.; et al. Antibacterial agents that inhibit lipid a biosynthesis. Science 1996, 274, 980–982. [Google Scholar] [CrossRef]
- Qureshi, N.; Takayama, K.; Mascagni, P.; Honovich, J.; Wong, R.; Cotter, R. Complete structural determination of lipopolysaccharide obtained from deep rough mutant of Escherichia coli. Purification by high performance liquid chromatography and direct analysis by plasma desorption mass spectrometry. J. Biol. Chem. 1988, 263, 11971–11976. [Google Scholar]
- Gibson, B.; Melaugh, W.; Phillips, N.; Apicella, M.; Campagnari, A.; Griffiss, J. Investigation of the structural heterogeneity of lipooligosaccharides from pathogenic Haemophilus and Neisseria species and of R-type lipopolysaccharides from Salmonella typhimurium by electrospray mass spectrometry. J. Bacteriol. 1993, 175, 2702–2712. [Google Scholar]
- Aderem, A.; Ulevitch, R.J. Toll-like receptors in the induction of the innate immune response. Nature 2000, 406, 782–787. [Google Scholar] [CrossRef]
- Magnadóttir, B. Innate immunity of fish (overview). Fish Shellfish Immunol. 2006, 20, 137–151. [Google Scholar] [CrossRef]
- Sogin, M.L.; Morrison, H.G.; Huber, J.A.; Welch, D.M.; Huse, S.M.; Neal, P.R.; Arrieta, J.M.; Herndl, G.J. Microbial diversity in the deep sea and the underexplored “rare biosphere”. Proc. Natl. Acad. Sci. USA 2006, 103, 12115–12120. [Google Scholar] [CrossRef]
- Toranzo, A.E.; Magariños, B.; Romalde, J.L. A review of the main bacterial fish diseases in mariculture systems. Aquaculture 2005, 246, 37–61. [Google Scholar] [CrossRef]
- Cervino, J.M.; Winiarski-Cervino, K.; Polson, S.W.; Goreau, T.; Smith, G.W. Identification of bacteria associated with a disease affecting the marine sponge Ianthella basta in new britain, papua new guinea. Mar. Ecol. Prog. Ser. 2006, 324, 139–150. [Google Scholar] [CrossRef]
- Pedersen, K.; Larsen, L. rRNA gene restriction patterns of Vibrio anguillarum serogroup O1. Dis. Aquat. Org. 1993, 16, 121–126. [Google Scholar] [CrossRef]
- Tiainen, T.; Pedersen, K.; Larsen, J.L. Ribotyping and plasmid profiling of Vibrio anguillarum serovar O2 and Vibrio ordalii. J. Appl. Bacteriol. 1995, 79, 384–392. [Google Scholar] [CrossRef]
- Bruno, D.W. Histopathology of bacterial kidney disease in laboratory infected rainbow trout, Salmo Gairdneri Richardson, and atlantic salmon, Salmo Salar L., with reference to naturally infected fish. J. Fish Dis. 1986, 9, 523–537. [Google Scholar] [CrossRef]
- Tison, D.L.; Nishibuchi, M.; Greenwood, J.D.; Seidler, R.J. Vibrio vulnificus biogroup 2: New biogroup pathogenic for eels. Appl. Environ. Microbiol. 1982, 44, 640–646. [Google Scholar]
- Lunder, T.; Sorum, H.; Holstad, G.; Steigerwalt, A.G.; Mowinckel, P.; Brenner, D.J. Phenotypic and genotypic characterization of Vibrio viscosus sp. nov. and Vibrio wodanis sp. nov. Isolated from Atlantic salmon (Salmo salar) with “winter ulcer”. Int. J. Syst. Evol. Microbiol. 2000, 50, 427–450. [Google Scholar] [CrossRef]
- Romalde, J.L. Photobacterium damselae subsp. piscicida: An integrated view of a bacterial fish pathogen. Int. Microbiol. 2002, 5, 3–9. [Google Scholar] [CrossRef]
- Wakabayashi, H.; Egusa, S. Characteristics of a Pseudomonas sp. From an epizootic of pond-cultured eels (Anguilla japonica). Nippon Suisan Gakkaishi 1972, 38, 577–587. [Google Scholar] [CrossRef]
- López-Romalde, S.; Magariños, B.; Núñez, S.; Toranzo, A.E.; Romalde, J.L. Phenotypic and genetic characterization of Pseudomonas anguilliseptica strains isolated from fish. J. Aquat. Anim. Health 2003, 15, 39–47. [Google Scholar] [CrossRef]
- Toranzo, A.E.; Barreiro, S.; Casal, J.F.; Figueras, A.; Barja, J.L. Pasteurellosis in cultured gilthead seabream (Sparus aurata): First report in spain. Aquaculture 1991, 99, 1–15. [Google Scholar] [CrossRef]
- Toranzo, A.E.; Santos, Y.; Nunez, S.; Barja, J.L. Biochemical and serological characteristics, drug resistance and plasmid profiles of spanish isolates of Aeromonas salmonicida. Fish Pathol. 1991, 26, 55–60. [Google Scholar] [CrossRef]
- Foster, G.; Osterman, B.S.; Godfroid, J.; Jacques, I.; Cloeckaert, A. Brucella ceti sp. nov. And Brucella pinnipedialis sp. nov. For brucella strains with cetaceans and seals as their preferred hosts. Int. J. Syst. Evol. Microbiol. 2007, 57, 2688–2693. [Google Scholar]
- Wakabayashi, H.; Hikida, M.; Masumura, K. Flexibacter maritimus sp. nov., a pathogen of marine fishes. Int. J. Syst. Bacteriol. 1986, 36, 396–398. [Google Scholar] [CrossRef]
- Bernardet, J.-F.; Grimont, P.A. Deoxyribonucleic acid relatedness and phenotypic characterization of Flexibacter columnaris sp. nov., nom. rev., Flexibacter psychrophilus sp. nov., nom. rev., and Flexibacter maritimus Wakabayashi, Hikida, and Masumura 1986. Int. J. Syst. Bacteriol. 1989, 39, 346–354. [Google Scholar] [CrossRef]
- Hedrick, R.P.; McDowell, T.; Groff, J. Mycobacteriosis in cultured striped bass from California. J. Wildl. Dis. 1987, 23, 391–395. [Google Scholar] [CrossRef]
- Bragg, R.R.; Huchzermeyer, H.F.; Hanisch, M.A. Mycobacterium fortuitum isolated from three species of fish in South Africa. Onderstepoort J. Vet. Res. 1990, 57, 101–102. [Google Scholar]
- Colorni, A. A systemic mycobacteriosis in the European sea bass Dicentrarchus labrax cultured in Eilat (Red Sea). Isr. J. Aquac. 1992, 44, 75–81. [Google Scholar]
- Lansdell, W.; Dixon, B.; Smith, N.; Benjamin, L. Communications: Isolation of several mycobacterium species from fish. J. Aquat. Anim. Health 1993, 5, 73–76. [Google Scholar] [CrossRef]
- Bruno, D.W.; Griffiths, J.; Mitchell, C.G.; Wood, B.P.; Fletcher, Z.J.; Drobniewski, F.A.; Hastings, T.S. Pathology attributed to Mycobacterium chelonae infection among farmed and laboratory-infected Atlantic salmon Salmo salar. Dis. Aquat. Organ. 1998, 33, 101–109. [Google Scholar] [CrossRef]
- Talaat, A.M.; Trucksis, M.; Kane, A.S.; Reimschuessel, R. Pathogenicity of Mycobacterium fortuitum and Mycobacterium smegmatis to goldfish, Carassius auratus. Vet. Microbiol. 1999, 66, 151–164. [Google Scholar] [CrossRef]
- Diamant, A.; Banet, A.; Ucko, M.; Colorni, A.; Knibb, W.; Kvitt, H. Mycobacteriosis in wild rabbitfish Siganus rivulatus associated with cage farming in the Gulf of Eilat, Red Sea. Dis. Aquat. Organ. 2000, 39, 211–219. [Google Scholar] [CrossRef]
- Rhodes, M.W.; Kator, H.; Kotob, S.; van Berkum, P.; Kaattari, I.; Vogelbein, W.; Floyd, M.M.; Butler, W.R.; Quinn, F.D.; Ottinger, C.; et al. A unique mycobacterium species isolated from an epizootic of striped bass (Morone saxatilis). Emerg. Infect. Dis. 2001, 7, 896–899. [Google Scholar] [CrossRef]
- Herbst, L.H.; Costa, S.F.; Weiss, L.M.; Johnson, L.K.; Bartell, J.; Davis, R.; Walsh, M.; Levi, M. Granulomatous skin lesions in moray eels caused by a novel mycobacterium species related to Mycobacterium triplex. Infect. Immun. 2001, 69, 4639–4646. [Google Scholar] [CrossRef]
- Dos Santos, N.M.; do Vale, A.; Sousa, M.J.; Silva, M.T. Mycobacterial infection in farmed turbot Scophthalmus maximus. Dis. Aquat. Organ. 2002, 52, 87–91. [Google Scholar] [CrossRef]
- Fryer, J.L.; Lannan, C.N.; Giovannoni, S.J.; Wood, N.D. Piscirickettsia salmonis gen. nov., sp. nov., the causative agent of an epizootic disease in salmonid fishes. Int. J. Syst. Bacteriol. 1992, 42, 120–126. [Google Scholar] [CrossRef]
- Bravo, S.; Campos, M. Coho salmon syndrome in chile. FHSAFS Newsl. 1989, 17, 3. [Google Scholar]
- García-Peña, F.J.; Pérez-Boto, D.; Jiménez, C.; San Miguel, E.; Echeita, A.; Rengifo-Herrera, C.; García-Párraga, D.; Ortega-Mora, L.M.; Pedraza-Díaz, S. Isolation and characterization of Campylobacter spp. From Antarctic fur seals (Arctocephalus gazella) at Deception Island, Antarctica. Appl. Environ. Microbiol. 2010, 76, 6013–6016. [Google Scholar] [CrossRef]
- Fenwick, S.G.; Duignan, P.J.; Nicol, C.M.; Leyland, M.J.; Hunter, J.E. A comparison of salmonella serotypes isolated from new Zealand sea lions and feral pigs on the Auckland Islands by pulsed-field gel electrophoresis. J. Wildl. Dis. 2004, 40, 566–570. [Google Scholar] [CrossRef]
- Stoddard, R.A.; Gulland, M.D.; Atwill, E.R.; Lawrence, J.; Jang, S.; Conrad, P.A. Salmonella and Campylobacter spp. In Northern Elephant Seals, California. Emerg. Infect. Dis. 2005, 11, 1967–1969. [Google Scholar]
- Palmgren, H.; McCafferty, D.; Aspan, A.; Broman, T.; Sellin, M.; Wollin, R.; Bergstrom, S.; Olsen, B. Salmonella in sub-Antarctica: Low heterogeneity in salmonella serotypes in South Georgian seals and birds. Epidemiol. Infect. 2000, 125, 257–262. [Google Scholar] [CrossRef]
- Sturm, N.; Abalos, P.; Fernandez, A.; Rodriguez, G.; Oviedo, P.; Arroyo, V.; Retamal, P. Salmonella enterica in Pinnipeds, Chile. Emerg. Infect. Dis. 2011, 17, 2377–2378. [Google Scholar] [CrossRef]
- Hansen, M.J.; Bertelsen, M.F.; Christensen, H.; Bojesen, A.M.; Bisgaard, M. Otariodibacter oris gen. nov., sp. nov., a member of the family pasteurellaceae isolated from the oral cavity of pinnipeds. Int. J. Syst. Evol. Microbiol. 2012, 62, 2572–2578. [Google Scholar] [CrossRef]
- Hansen, M.J.; Bertelsen, M.F.; Delaney, M.A.; Fravel, V.A.; Gulland, F.; Bojesen, A.M. Otariodibacter oris and Bisgaardia genomospecies 1 isolated from infections in pinnipeds. J. Wildl. Dis. 2013, 49, 661–665. [Google Scholar] [CrossRef]
- Foster, G.; Ross, H.; Patterson, I.; Hutson, R.; Collins, M. Actinobacillus scotiae sp. nov., a new member of the family Pasteurellaceae Pohl (1979) 1981 isolated from porpoises (Phocoena phocoena). Int. J. Syst. Bacteriol. 1998, 48, 929–933. [Google Scholar]
- Munro, R.; Ross, H.; Cornwell, C.; Gilmour, J. Disease conditions affecting common seals (Phoca vitulina) around the scottish mainland, september-november 1988. Sci. Total Environ. 1992, 115, 67–82. [Google Scholar] [CrossRef]
- Baker, J.R.; Ross, H.M. The role of bacteria in phocine distemper. Sci. Total Environ. 1992, 115, 9–14. [Google Scholar] [CrossRef]
- Holst, O.; Ulmer, A.J.; Brade, H.; Flad, H.D.; Rietschel, E.T. Biochemistry and cell biology of bacterial endotoxins. FEMS Immunol. Med. Microbiol. 1996, 16, 83–104. [Google Scholar] [CrossRef]
- Westphal, O.; Lüderitz, O.; Galanos, C.; Mayer, H.; Rietschel, E.T. The story of bacterial endotoxin. Adv. Immunopharmacol. 1986, 3, 13–34. [Google Scholar]
- Leone, S.; Silipo, A.; Nazarenko, E.L.; Lanzetta, R.; Parrilli, M.; Molinaro, A. Molecular structure of endotoxins from Gram-negative marine bacteria: An update. Mar. Drugs 2007, 5, 85–112. [Google Scholar] [CrossRef]
- Nazarenko, E.L.; Komandrova, N.A.; Gorshkova, R.P.; Tomshich, S.V.; Zubkov, V.A.; Kilcoyne, M.; Savage, A.V. Structures of polysaccharides and oligosaccharides of some Gram-negative marine proteobacteria. Carbohydr. Res. 2003, 338, 2449–2457. [Google Scholar] [CrossRef]
- Turska-Szewczuk, A.; Lindner, B.; Komaniecka, I.; Kozinska, A.; Pekala, A.; Choma, A.; Holst, O. Structural and immunochemical studies of the lipopolysaccharide from the fish pathogen, Aeromonas bestiarum strain K296, serotype O18. Mar. Drugs 2013, 11, 1235–1255. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Shashkov, A.S.; Senchenkova, S.y.N.; Merino, S.; Tomás, J.M. Structure of the O-polysaccharide of Aeromonashydrophila O:34; a case of random O-acetylation of 6-deoxy-l-talose. Carbohydr. Res. 2002, 337, 1381–1386. [Google Scholar]
- Cipriano, R.C.; Bullock, G.; Pyle, S. Aeromonas Hydrophila and Motile Aeromonad Septicemias of Fish; U.S. Fish and Wildlife Service: Washington, DC, USA, 1984; pp. 1–23. [Google Scholar]
- Janda, J.M. Recent advances in the study of the taxonomy, pathogenicity, and infectious syndromes associated with the genus aeromonas. Clin. Microbiol. Rev. 1991, 4, 397–410. [Google Scholar]
- Jensen, P.R.; Fenical, W. Strategies for the discovery of secondary metabolites from marine bacteria: Ecological perspectives. Annu. Rev. Microbiol. 1994, 48, 559–584. [Google Scholar] [CrossRef]
- Holmstrom, C.; Kjelleberg, S. Marine pseudoalteromonas species are associated with higher organisms and produce biologically active extracellular agents. FEMS Microbiol. Ecol. 1999, 30, 285–293. [Google Scholar] [CrossRef]
- Silipo, A.; Molinaro, A.; Nazarenko, E.L.; Gorshkova, R.P.; Ivanova, E.P.; Lanzetta, R.; Parrilli, M. The O-chain structure from the LPS of marine halophilic bacterium Pseudoalteromonas carrageenovora-type strain iam 12662T. Carbohydr. Res. 2005, 340, 2693–2697. [Google Scholar] [CrossRef]
- Silipo, A.; Leone, S.; Lanzetta, R.; Parrilli, M.; Sturiale, L.; Garozzo, D.; Nazarenko, E.L.; Gorshkova, R.P.; Ivanova, E.P.; Gorshkova, N.M.; et al. The complete structure of the lipooligosaccharide from the halophilic bacterium Pseudoalteromonas issachenkonii KMM 3549T. Carbohydr. Res. 2004, 339, 1985–1993. [Google Scholar]
- Corsaro, M.M.; Lanzetta, R.; Parrilli, E.; Parrilli, M.; Tutino, M.L. Structural investigation on the lipooligosaccharide fraction of psychrophilic Pseudoalteromonas haloplanktis TAC 125 bacterium. Eur. J. Biochem. FEBS 2001, 268, 5092–5097. [Google Scholar] [CrossRef]
- Krasikova, I.N.; Kapustina, N.V.; Isakov, V.V.; Gorshkova, N.M.; Solov’eva, T.F. Elucidation of structure of lipid a from the marine Gram-negative bacterium Pseudoalteromonas haloplanktis ATCC 14393T. Bioorganicheskaia Khimiia 2004, 30, 409–416. [Google Scholar]
- Corsaro, M.M.; Piaz, F.D.; Lanzetta, R.; Parrilli, M. Lipid a structure of Pseudoalteromonas haloplanktis TAC 125: Use of electrospray ionization tandem mass spectrometry for the determination of fatty acid distribution. J. Mass Spectrom. 2002, 37, 481–488. [Google Scholar] [CrossRef]
- Leone, S.; Molinaro, A.; Sturiale, L.; Garozzo, D.; Nazarenko, E.L.; Gorshkova, R.P.; Ivanova, E.P.; Shevchenko, L.S.; Lanzetta, R.; Parrilli, M. The outer membrane of the marine Gram-negative bacterium Alteromonas addita is composed of a very short-chain lipopolysaccharide with a high negative charge density. Eur. J. Org. Chem. 2007, 2007, 1113–1122. [Google Scholar]
- Liparoti, V.; Molinaro, A.; Sturiale, L.; Garozzo, D.; Nazarenko, E.L.; Gorshkova, R.P.; Ivanova, E.P.; Shevcenko, L.S.; Lanzetta, R.; Parrilli, M. Structural analysis of the deep rough lipopolysaccharide from gram negative bacterium Alteromonas macleodii atcc 27126T: The first finding of β-Kdo in the inner core of lipopolysaccharides. Eur. J. Org. Chem. 2006, 2006, 4710–4716. [Google Scholar] [CrossRef]
- Vinogradov, E.; Korenevsky, A.; Beveridge, T.J. The structure of the O-specific polysaccharide chain of the Shewanella algae BrY lipopolysaccharide. Carbohydr. Res. 2003, 338, 385–388. [Google Scholar] [CrossRef]
- Shashkov, A.S.; Torgov, V.I.; Nazarenko, E.L.; Zubkov, V.A.; Gorshkova, N.M.; Gorshkova, R.P.; Widmalm, G. Structure of the phenol-soluble polysaccharide from Shewanella putrefaciens strain A6. Carbohydr. Res. 2002, 337, 1119–1127. [Google Scholar]
- Vinogradov, E.V.; Bock, K.; Petersen, B.O.; Holst, O.; Brade, H. The structure of the carbohydrate backbone of the lipopolysaccharide from Acinetobacter strain ATCC 17905. Eur. J. Biochem. 1997, 243, 122–127. [Google Scholar]
- Vinogradov, E.; MacLean, L.L.; Crump, E.M.; Perry, M.B.; Kay, W.W. Structure of the polysaccharide chain of the lipopolysaccharide from Flexibacter maritimus. Eur. J. Biochem. 2003, 270, 1810–1815. [Google Scholar] [CrossRef]
- MacLean, L.L.; Vinogradov, E.; Crump, E.M.; Perry, M.B.; Kay, W.W. The structure of the lipopolysaccharide O-antigen produced by Flavobacterium psychrophilum (259–93). Eur. J. Biochem. 2001, 268, 2710–2716. [Google Scholar] [CrossRef]
- Perepelov, A.V.; Shashkov, A.S.; Tomshich, S.V.; Komandrova, N.A.; Nedashkovskaya, O.I. A pseudoaminic acid-containing O-specific polysaccharide from a marine bacterium Cellulophaga fucicola. Carbohydr. Res. 2007, 342, 1378–1381. [Google Scholar] [CrossRef]
- Baumann, L.; Baumann, P.; Mandel, M.; Allen, R.D. Taxonomy of aerobic marine eubacteria. J. Bacteriol. 1972, 110, 402–429. [Google Scholar]
- Gauthier, G.; Gauthier, M.; Christen, R. Phylogenetic analysis of the genera Alteromonas, Shewanella, and Moritella using genes coding for small-subunit rRNA sequences and division of the genus Alteromonas into two genera, Alteromonas (emended) and Pseudoalteromonas gen. nov., and proposal of twelve new species combinations. Int. J. Syst. Bacteriol. 1995, 45, 755–761. [Google Scholar] [CrossRef]
- De Castro, C.; Carannante, A.; Lanzetta, R.; Liparoti, V.; Molinaro, A.; Parrilli, M. Core oligosaccharide structure from the highly phytopathogenic Agrobacterium tumefaciens TT111 and conformational analysis of the putative rhamnan epitope. Glycobiology 2006, 16, 1272–1280. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Bowman, J.P.; Lysenko, A.M.; Zhukova, N.V.; Gorshkova, N.M.; Sergeev, A.F.; Mikhailov, V.V. Alteromonas addita sp. Nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 1065–1068. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Nedashkovskaya, O.I.; Chun, J.; Lysenko, A.M.; Frolova, G.M.; Svetashev, V.I.; Vysotskii, M.V.; Mikhailov, V.V.; Huq, A.; Colwell, R.R. Arenibacter gen. nov., new genus of the family Flavobacteriaceae and description of a new species, Arenibacter latericius sp. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 1987–1995. [Google Scholar] [CrossRef]
- Vorob’eva, E.V.; Krasikova, I.N.; Dmitrenok, A.S.; Dmitrenok, P.S.; Isakov, V.V.; Nedashkovskaia, O.I.; Solov’eva, T.F. An unusual lipid a from a marine bacterium Chryseobacterium scophtalmum CIP 104199T. Bioorganicheskaia Khimiia 2006, 32, 538–545. [Google Scholar]
- Nishijima, M.; Raetz, C.R. Membrane lipid biogenesis in Escherichia coli: Identification of genetic loci for phosphatidylglycerophosphate synthetase and construction of mutants lacking phosphatidylglycerol. J. Biol. Chem. 1979, 254, 7837–7844. [Google Scholar]
- Nishijima, M.; Bulawa, C.E.; Raetz, C.R. Two interacting mutations causing temperature-sensitive phosphatidylglycerol synthesis in Escherichia coli membranes. J. Bacteriol. 1981, 145, 113–121. [Google Scholar]
- Nishijima, M.; Raetz, C.R. Characterization of two membrane-associated glycolipids from an Escherichia coli mutant deficient in phosphatidylglycerol. J. Biol. Chem. 1981, 256, 10690–10696. [Google Scholar]
- Alexander, C.; Rietschel, E.T. Bacterial lipopolysaccharides and innate immunity. J. Endotoxin Res. 2001, 7, 167–202. [Google Scholar]
- Krasikova, I.N.; Kapustina, N.V.; Svetashev, V.I.; Gorshkova, R.P.; Tomshich, S.V.; Nazarenko, E.L.; Komandrova, N.A.; Ivanova, E.P.; Gorshkova, N.M.; Romanenko, L.A.; et al. Chemical characterization of lipid A from some marine proteobacteria. Biochem. Biokhimiia 2001, 66, 1047–1054. [Google Scholar] [CrossRef]
- Uribe, C.; Folch, H.; Enriquez, R.; Moran, G. Innate and adaptive immunity in teleost fish: A review. Vet. Med. 2011, 56, 486–503. [Google Scholar]
- Whyte, S.K. The innate immune response of finfish—A review of current knowledge. Fish Shellfish Immunol. 2007, 23, 1127–1151. [Google Scholar] [CrossRef]
- Magnadottir, B. Immunological control of fish diseases. Mar. Biotechnol. 2010, 12, 361–379. [Google Scholar] [CrossRef]
- Ingram, G.A. Substances involved in the natural resistance of fish to infection–—A review. J. Fish Biol. 1980, 16, 23–60. [Google Scholar] [CrossRef]
- Shephard, K.L. Functions for fish mucus. Rev. Fish Biol. Fish. 1994, 4, 401–429. [Google Scholar] [CrossRef]
- Ellis, A. Innate host defense mechanisms of fish against viruses and bacteria. Dev. Comp. Immunol. 2001, 25, 827–839. [Google Scholar] [CrossRef]
- Alexander, J.B.; Ingram, G.A. Noncellular nonspecific defence mechanisms of fish. Annu. Rev. Fish Dis. 1992, 2, 249–279. [Google Scholar] [CrossRef]
- Rombout, J.H.; Taverne, N.; van de Kamp, M.; Taverne-Thiele, A.J. Differences in mucus and serum immunoglobulin of carp (Cyprinus carpio L.). Dev. Comp. Immunol. 1993, 17, 309–317. [Google Scholar]
- Aranishi, F.; Nakane, M. Epidermal proteases of the Japanese eel. Fish Physiol. Biochem. 1997, 16, 471–478. [Google Scholar] [CrossRef]
- Boshra, H.; Li, J.; Sunyer, J. Recent advances on the complement system of teleost fish. Fish Shellfish Immunol. 2006, 20, 239–262. [Google Scholar] [CrossRef]
- Saurabh, S.; Sahoo, P. Lysozyme: An important defence molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Hibiya, T.; Yokote, M.; Oguri, M.; Sato, H.; Takashima, F.; Aida, K. An Atlas of Fish Histology. Normal and Pathological Features; Gustav Fischer Verlag: Stuttgart, Germany, 1982; Volume 69, p. 147. [Google Scholar]
- Evans, D.L.; Jaso-Friedmann, L. Nonspecific cytotoxic cells as effectors of immunity in fish. Annu. Rev. Fish Dis. 1992, 2, 109–121. [Google Scholar] [CrossRef]
- Hasegawa, S.; Nakayasu, C.; Yoshitomi, T.; Nakanishi, T.; Okamoto, N. Specific cell-mediated cytotoxicity against an allogeneic target cell line in isogeneic ginbuna crucian carp. Fish Shellfish Immunol. 1998, 8, 303–313. [Google Scholar] [CrossRef]
- Hellio, C.; Pons, A.M.; Beaupoil, C.; Bourgougnon, N.; Gal, Y.L. Antibacterial, antifungal and cytotoxic activities of extracts from fish epidermis and epidermal mucus. Int. J. Antimicrob. Agents 2002, 20, 214–219. [Google Scholar] [CrossRef]
- Chinchar, V.; Bryan, L.; Silphadaung, U.; Noga, E.; Wade, D.; Rollins-Smith, L. Inactivation of viruses infecting ectothermic animals by amphibian and piscine antimicrobial peptides. Virology 2004, 323, 268–275. [Google Scholar] [CrossRef]
- Maier, V.H.; Dorn, K.V.; Gudmundsdottir, B.K.; Gudmundsson, G.H. Characterisation of cathelicidin gene family members in divergent fish species. Mol. Immunol. 2008, 45, 3723–3730. [Google Scholar]
- Blazer, V.S. Piscine macrophage function and nutritional influences: A review. J. Aquat. Anim. Health 1991, 3, 77–86. [Google Scholar] [CrossRef]
- Lange, S.; Magnadóttir, B. Spontaneous haemolytic activity of atlantic halibut (Hippoglossus hippoglossus L.) and sea bass (Dicentrarchus labrax) serum. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 136, 99–106. [Google Scholar] [CrossRef]
- Magnadóttir, B.; Lange, S.; Gudmundsdottir, S.; Bøgwald, J.; Dalmo, R. Ontogeny of humoral immune parameters in fish. Fish Shellfish Immunol. 2005, 19, 429–439. [Google Scholar] [CrossRef]
- Secombes, C.; Fletcher, T. The role of phagocytes in the protective mechanisms of fish. Annu. Rev. Fish Dis. 1992, 2, 53–71. [Google Scholar]
- Fischer, U.; Utke, K.; Somamoto, T.; Köllner, B.; Ototake, M.; Nakanishi, T. Cytotoxic activities of fish leucocytes. Fish Shellfish Immunol. 2006, 20, 209–226. [Google Scholar] [CrossRef]
- Matsushita, M.; Endo, Y.; Nonaka, M.; Fujita, T. Complement-related serine proteases in tunicates and vertebrates. Curr. Opin. Immunol. 1998, 10, 29–35. [Google Scholar] [CrossRef]
- Nikoskelainen, S.; Lehtinen, J.; Lilius, E.-M. Bacteriolytic activity of rainbow trout (Oncorhynchus mykiss) complement. Dev. Comp. Immunol. 2002, 26, 797–804. [Google Scholar] [CrossRef]
- Yano, T. 3 the nonspecific immune system: Humoral defense. Fish Physiol. 1997, 15, 105–157. [Google Scholar] [CrossRef]
- Matsuyama, H.; Yano, T.; Yamakawa, T.; Nakao, M. Opsonic effect of the third complement component (c3) of carp (Cyprinus carpio) on phagocytosis by neutrophils. Fish Shellfish Immunol. 1992, 2, 69–78. [Google Scholar] [CrossRef]
- Jenkins, J.A.; Ourth, D.D. Opsonic effect of the alternative complement pathway on channel catfish peripheral blood phagocytes. Vet. Immunol. Immunopathol. 1993, 39, 447–459. [Google Scholar]
- Lammens, M.; Decostere, A.; Haesebrouck, F. Effect of flavobacterium psychrophilum strains and their metabolites on the oxidative activity of rainbow trout Oncorhynchus mykiss phagocytes. Dis. Aquat. Organ. 2000, 41, 173–179. [Google Scholar] [CrossRef]
- Lorenzen, N.; Lapatra, S.E. Immunity to rhabdoviruses in rainbow trout: The antibody response. Fish Shellfish Immunol. 1999, 9, 345–360. [Google Scholar] [CrossRef]
- Yin, Z.; Lam, T.; Sin, Y. Cytokine-mediated antimicrobial immune response of catfish, Clarias gariepinus, as a defence against Aeromonas hydrophil. Fish Shellfish Immunol. 1997, 7, 93–104. [Google Scholar] [CrossRef]
- Mulero, V.; Meseguer, J. Functional characterisation of a macrophage-activating factor produced by leucocytes of gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 1998, 8, 143–156. [Google Scholar] [CrossRef]
- Tafalla, C.; Figueras, A.; Novoa, B. Viral hemorrhagic septicemia virus alters turbot Scophthalmus maximus macrophage nitric oxide production. Dis. Aquat. Organ. 2001, 47, 101–107. [Google Scholar] [CrossRef]
- Robertsen, B. The interferon system of teleost fish. Fish Shellfish Immunol. 2006, 20, 172–191. [Google Scholar] [CrossRef]
- De Veer, M.J.; Holko, M.; Frevel, M.; Walker, E.; Der, S.; Paranjape, J.M.; Silverman, R.H.; Williams, B.R. Functional classification of interferon-stimulated genes identified using microarrays. J. Leukoc. Biol. 2001, 69, 912–920. [Google Scholar]
- Samuel, C.E. Antiviral actions of interferons. Clin. Microbiol. Rev. 2001, 14, 778–809. [Google Scholar]
- Savan, R.; Sakai, M. Genomics of fish cytokines. Comp. Biochem. Physiol. Part D Genomics Proteomics 2006, 1, 89–101. [Google Scholar] [CrossRef]
- Arma, N.; Hirono, I.; Aoki, T. Characterization of expressed genes in kidney cells of Japanese flounder Paralichthys olivaceus following treatment with ConA/PMA and LPS. Fish Pathol. 2004, 39, 189–196. [Google Scholar] [CrossRef]
- Hanington, P.C.; Wang, T.; Secombes, C.J.; Belosevic, M. Growth factors of lower vertebrates: Characterization of goldfish (Carassius auratus L.) macrophage colony-stimulating factor factor-1. J. Biol. Chem. 2007, 282, 31865–31872. [Google Scholar] [CrossRef]
- Kono, T.; Bird, S.; Sonoda, K.; Savan, R.; Secombes, C.J.; Sakai, M. Characterization and expression analysis of an interleukin-7 homologue in the Japanese pufferfish, Takifugu rubripes. FEBS J. 2008, 275, 1213–1226. [Google Scholar] [CrossRef]
- Bowden, T.; Butler, R.; Bricknell, I.; Ellis, A. Serum trypsin-inhibitory activity in five species of farmed fish. Fish Shellfish Immunol. 1997, 7, 377–385. [Google Scholar] [CrossRef]
- Bayne, C.J.; Gerwick, L. The acute phase response and innate immunity of fish. Dev. Comp. Immunol. 2001, 25, 725–743. [Google Scholar]
- Wu, Y.; Wang, S.; Peng, X. Serum acute phase response (APR)-related proteome of loach to trauma. Fish Shellfish Immunol. 2004, 16, 381–389. [Google Scholar] [CrossRef]
- Cook, M.T.; Hayball, P.J.; Birdseye, L.; Bagley, C.; Nowak, B.F.; Hayball, J.D. Isolation and partial characterization of a pentraxin-like protein with complement-fixing activity from snapper (Pagrus auratus, sparidae) serum. Dev. Comp. Immunol. 2003, 27, 579–588. [Google Scholar]
- De Haas, C.J.; van Leeuwen, E.M.; van Bommel, T.; Verhoef, J.; van Kessel, K.P.; van Strijp, J.A. Serum amyloid p component bound to Gram-negative bacteria prevents lipopolysaccharide-mediated classical pathway complement activation. Infect. Immun. 2000, 68, 1753–1759. [Google Scholar] [CrossRef]
- Nauta, A.J.; Daha, M.R.; van Kooten, C.; Roos, A. Recognition and clearance of apoptotic cells: A role for complement and pentraxins. Trends Immunol. 2003, 24, 148–154. [Google Scholar] [CrossRef]
- Grinde, B.; Lie, Ø.; Poppe, T.; Salte, R. Species and individual variation in lysozyme activity in fish of interest in aquaculture. Aquaculture 1988, 68, 299–304. [Google Scholar] [CrossRef]
- Sveinbjornsson, B.; Olsen, R.; Paulsen, S. Immunocytochemical localization of lysozyme in intestinal eosinophilic granule cells (EGCs) of atlantic salmon, Salmo salar L. J. Fish Dis. 1996, 19, 349–355. [Google Scholar] [CrossRef]
- Boes, M. Role of natural and immune IgM antibodies in immune responses. Mol. Immunol. 2000, 37, 1141–1149. [Google Scholar] [CrossRef]
- Acton, R.T.; Weinheimer, P.F.; Hall, S.J.; Niedermeier, W.; Shelton, E.; Bennett, J.C. Tetrameric immune macroglobulins in three orders of bony fishes. Proc. Natl. Acad. Sci. USA 1971, 68, 107–111. [Google Scholar] [CrossRef]
- Wilson, M.R.; Warr, G.W. Fish immunoglobulins and the genes that encode them. Annu. Rev. Fish Dis. 1992, 2, 201–221. [Google Scholar] [CrossRef]
- Elcombe, B.M.; Chang, R.J.; Taves, C.J.; Winkelhake, J.L. Evolution of antibody structure and effector functions: Comparative hemolytic activities of monomeric and tetrameric IgM from rainbow trout, Salmo gairdnerii. Comp. Biochem. Physiol. Part B Comp. Biochem. 1985, 80, 697–706. [Google Scholar] [CrossRef]
- Wilson, M.; Bengtén, E.; Miller, N.W.; Clem, L.W.; Du Pasquier, L.; Warr, G.W. A novel chimeric Ig heavy chain from a teleost fish shares similarities to IgD. Proc. Natl. Acad. Sci. USA 1997, 94, 4593–4597. [Google Scholar]
- Hatten, F.; Fredriksen, Å.; Hordvik, I.; Endresen, C. Presence of igm in cutaneous mucus, but not in gut mucus of atlantic salmon, Salmo salar. Serum IgM is rapidly degraded when added to gut mucus. Fish Shellfish Immunol. 2001, 11, 257–268. [Google Scholar] [CrossRef]
- Rombout, J.W.; Blok, L.J.; Lamers, C.H.; Egberts, E. Immunization of carp (Cyprinus carpio) with a vibrio anguillarum bacterin: Indications for a common mucosal immune system. Dev. Comp. Immunol. 1986, 10, 341–351. [Google Scholar]
- Lumsden, J.; Ostland, V.; Byrne, P.; Ferguson, H. Detection of a distinct gill-surface antibody response following horizontal infection and bath challenge of brook trout Salvelinus fontinalis with Flavobacterium branchiophilum, the causative agent of bacterial gill disease. Dis. Aquat. Organ. 1993, 16, 21–27. [Google Scholar] [CrossRef]
- Jenkins, P.; Wrathmell, A.; Harris, J.; Pulsford, A. Systemic and mucosal immune responses to enterically delivered antigen in Oreochromis mossambicus. Fish Shellfish Immunol. 1994, 4, 255–271. [Google Scholar] [CrossRef]
- Arkoosh, M.R.; Kaattari, S.L. Development of immunological memory in rainbow trout (Oncorhynchus mykiss). I. An immunochemical and cellular analysis of the B cell response. Dev. Comp. Immunol. 1991, 15, 279–293. [Google Scholar] [CrossRef]
- Whittington, R.; Munday, B.; Akhlaghi, M.; Reddacliff, G.; Carson, J. Humoral and peritoneal cell responses of rainbow trout (Oncorhynchus mykiss) to ovalbumin, Vibrio anguillarum and freund’s complete adjuvant following intraperitoneal and bath immunisation. Fish Shellfish Immunol. 1994, 4, 475–488. [Google Scholar] [CrossRef]
- Dos Santos, N.; Romano, N.; de Sousa, M.; Ellis, A.E.; Rombout, J.H. Ontogeny of B and T cells in sea bass (Dicentrarchus labrax, L.). Fish Shellfish Immunol. 2000, 10, 583–596. [Google Scholar] [CrossRef]
- Secombes, C. Will advances in fish immunology change vaccination strategies? Fish Shellfish Immunol. 2008, 25, 409–416. [Google Scholar] [CrossRef]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: Evidence for TLR4 as the Lps gene product. J. Immunol. 1999, 162, 3749–3752. [Google Scholar]
- Wiens, M.; Korzhev, M.; Krasko, A.; Thakur, N.L.; Perović-Ottstadt, S.; Breter, H.J.; Ushijima, H.; Diehl-Seifert, B.; Müller, I.M.; Müller, W.E.G. Innate immune defense of the sponge Suberites domuncula against bacteria involves a MyD88-dependent signaling pathway: Induction of a perforin-like molecule. J. Biol. Chem. 2005, 280, 27949–27959. [Google Scholar] [CrossRef]
- Sullivan, C.; Charette, J.; Catchen, J.; Lage, C.R.; Giasson, G.; Postlethwait, J.H.; Millard, P.J.; Kim, C.H. The gene history of zebrafish TLR4A and TLR4B is predictive of their divergent functions. J. Immunol. 2009, 183, 5896–5908. [Google Scholar] [CrossRef]
- Neumann, N.; Fagan, D.; Belosevic, M. Macrophage activating factor(s) secreted by mitogen stimulated goldfish kidney leukocytes synergize with bacterial lipopolysaccharide to induce nitric oxide production in teleost macrophages. Dev. Comp. Immunol. 1995, 19, 473–482. [Google Scholar] [CrossRef]
- Darawiroj, D.; Kondo, H.; Hirono, I.; Aoki, T. Immune-related gene expression profiling of yellowtail (Seriola quinqueradiata) kidney cells stimulated with ConA and LPS using microarray analysis. Fish Shellfish Immunol. 2008, 24, 260–266. [Google Scholar] [CrossRef]
- Purcell, M.K.; Smith, K.D.; Hood, L.; Winton, J.R.; Roach, J.C. Conservation of Toll-like receptor signaling pathways in teleost fish. Comp. Biochem. Physiol. Part D Genomics Proteomics 2006, 1, 77–88. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Chen, F.Y.; Thilagam, H.; Qiao, K.; Xu, W.F.; Wang, K.J. Modulation and interaction of immune-associated parameters with antioxidant in the immunocytes of crab Scylla paramamosain challenged with lipopolysaccharides. Evid. Based Complement. Alternat. Med. 2011, 2011, 824962. [Google Scholar]
- Palmer, C.V.; McGinty, E.S.; Cummings, D.J.; Smith, S.M.; Bartels, E.; Mydlarz, L.D. Patterns of coral ecological immunology: Variation in the responses of caribbean corals to elevated temperature and a pathogen elicitor. J. Exp. Biol. 2011, 214, 4240–4249. [Google Scholar] [CrossRef]
- Sepulcre, M.P.; Alcaraz-Perez, F.; Lopez-Munoz, A.; Roca, F.J.; Meseguer, J.; Cayuela, M.L.; Mulero, V. Evolution of lipopolysaccharide (LPS) recognition and signaling: Fish TLR4 does not recognize LPS and negatively regulates NF-κB activation. J. Immunol. 2009, 182, 1836–1845. [Google Scholar] [CrossRef]
- Swain, P.; Nayak, S.K.; Nanda, P.K.; Dash, S. Biological effects of bacterial lipopolysaccharide (endotoxin) in fish: A review. Fish Shellfish Immunol. 2008, 25, 191–201. [Google Scholar] [CrossRef]
- Iliev, D.B.; Roach, J.C.; Mackenzie, S.; Planas, J.V.; Goetz, F.W. Endotoxin recognition: In fish or not in fish? FEBS Lett. 2005, 579, 6519–6528. [Google Scholar] [CrossRef]
- Kabanov, D.S.; Prokhorenko, I.R. Structural analysis of lipopolysaccharides from gram-negative bacteria. Biochem. Biokhimiia 2010, 75, 383–404. [Google Scholar] [CrossRef]
- Loppnow, H.; Brade, H.; Durrbaum, I.; Dinarello, C.A.; Kusumoto, S.; Rietschel, E.T.; Flad, H.D. IL-1 induction-capacity of defined lipopolysaccharide partial structures. J. Immunol. 1989, 142, 3229–3238. [Google Scholar]
- Brandenburg, K.; Seydel, U. Investigation into the fluidity of lipopolysaccharide and free lipid A membrane systems by fourier-transform infrared spectroscopy and differential scanning calorimetry. Eur. J. Biochem. 1990, 191, 229–236. [Google Scholar] [CrossRef]
- Christ, W.J.; Asano, O.; Robidoux, A.L.; Perez, M.; Wang, Y.; Dubuc, G.R.; Gavin, W.E.; Hawkins, L.D.; McGuinness, P.D.; Mullarkey, M.A.; et al. E5531, a pure endotoxin antagonist of high potency. Science 1995, 268, 80–83. [Google Scholar]
- Raetz, C.R.H.; Garrett, T.A.; Reynolds, C.M.; Shaw, W.A.; Moore, J.D.; Smith, D.C.; Ribeiro, A.A.; Murphy, R.C.; Ulevitch, R.J.; Fearns, C.; et al. Kdo2-lipid A of Escherichia coli, a defined endotoxin that activates macrophages via TLR-4. J. Lipid Res. 2006, 47, 1097–1111. [Google Scholar] [CrossRef]
- Ulmer, A.; Heine, H.; Feist, W.; Kusumoto, S.; Kusama, T.; Brade, H.; Schade, U.; Rietschel, E.T.; Flad, H. Biological activity of synthetic phosphonooxyethyl analogs of lipid a and lipid a partial structures. Infect. Immun. 1992, 60, 3309–3314. [Google Scholar]
- Zahringer, U.; Knirel, Y.A.; Lindner, B.; Helbig, J.H.; Sonesson, A.; Marre, R.; Rietschel, E.T. The lipopolysaccharide of Legionella pneumophila serogroup 1 (strain Philadelphia 1): Chemical structure and biological significance. Prog. Clin. Biol. Res. 1995, 392, 113–139. [Google Scholar]
- Loppnow, H.; Libby, P.; Freudenberg, M.; Krauss, J.H.; Weckesser, J.; Mayer, H. Cytokine induction by lipopolysaccharide (LPS) corresponds to lethal toxicity and is inhibited by nontoxic rhodobacter capsulatus LPS. Infect. Immun. 1990, 58, 3743–3750. [Google Scholar]
- Tanamoto, K.; Azumi, S. Salmonella-type heptaacylated lipid a is inactive and acts as an antagonist of lipopolysaccharide action on human line cells. J. Immunol. 2000, 164, 3149–3156. [Google Scholar] [CrossRef]
- Que-Gewirth, N.L.; Ribeiro, A.A.; Kalb, S.R.; Cotter, R.J.; Bulach, D.M.; Adler, B.; Girons, I.S.; Werts, C.; Raetz, C.R. A methylated phosphate group and four amide-linked acyl chains in leptospira interrogans lipid A. The membrane anchor of an unusual lipopolysaccharide that activates TLR2. J. Biol. Chem. 2004, 279, 25420–25429. [Google Scholar]
- Caroff, M.; Karibian, D.; Cavaillon, J.M.; Haeffner-Cavaillon, N. Structural and functional analyses of bacterial lipopolysaccharides. Microbes Infect. Inst. Pasteur 2002, 4, 915–926. [Google Scholar] [CrossRef]
- Warabi, K.; Hamada, T.; Nakao, Y.; Matsunaga, S.; Hirota, H.; van Soest, R.W.; Fusetani, N. Axinelloside a, an unprecedented highly sulfated lipopolysaccharide inhibiting telomerase, from the marine sponge, axinella infundibula. J. Am. Chem. Soc. 2005, 127, 13262–13270. [Google Scholar]
- Vorobeva, E.V.; Krasikova, I.N.; Solov’eva, T.F. Influence of lipopolysaccharides and lipids A from some marine bacteria on spontaneous and Escherichia coli LPS-induced TNF-alpha release from peripheral human blood cells. Biochem. Biokhimiia 2006, 71, 759–766. [Google Scholar] [CrossRef]
- Maaetoft-Udsen, K.; Vynne, N.; Heegaard, P.M.; Gram, L.; Frokiaer, H. Pseudoalteromonas strains are potent immunomodulators owing to low-stimulatory LPS. Innate Immun. 2013, 19, 160–173. [Google Scholar] [CrossRef]
- Seppälä, I.; Mäkelä, O. Adjuvant effect of bacterial LPS and/or alum precipitation in responses to polysaccharide and protein antigens. Immunology 1984, 53, 827–836. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Anwar, M.A.; Choi, S. Gram-Negative Marine Bacteria: Structural Features of Lipopolysaccharides and Their Relevance for Economically Important Diseases. Mar. Drugs 2014, 12, 2485-2514. https://doi.org/10.3390/md12052485
Anwar MA, Choi S. Gram-Negative Marine Bacteria: Structural Features of Lipopolysaccharides and Their Relevance for Economically Important Diseases. Marine Drugs. 2014; 12(5):2485-2514. https://doi.org/10.3390/md12052485
Chicago/Turabian StyleAnwar, Muhammad Ayaz, and Sangdun Choi. 2014. "Gram-Negative Marine Bacteria: Structural Features of Lipopolysaccharides and Their Relevance for Economically Important Diseases" Marine Drugs 12, no. 5: 2485-2514. https://doi.org/10.3390/md12052485