Phylogenetic Diversity and Biological Activity of Actinobacteria Isolated from the Chukchi Shelf Marine Sediments in the Arctic Ocean

Abstract
:1. Introduction
2. Results and Discussion
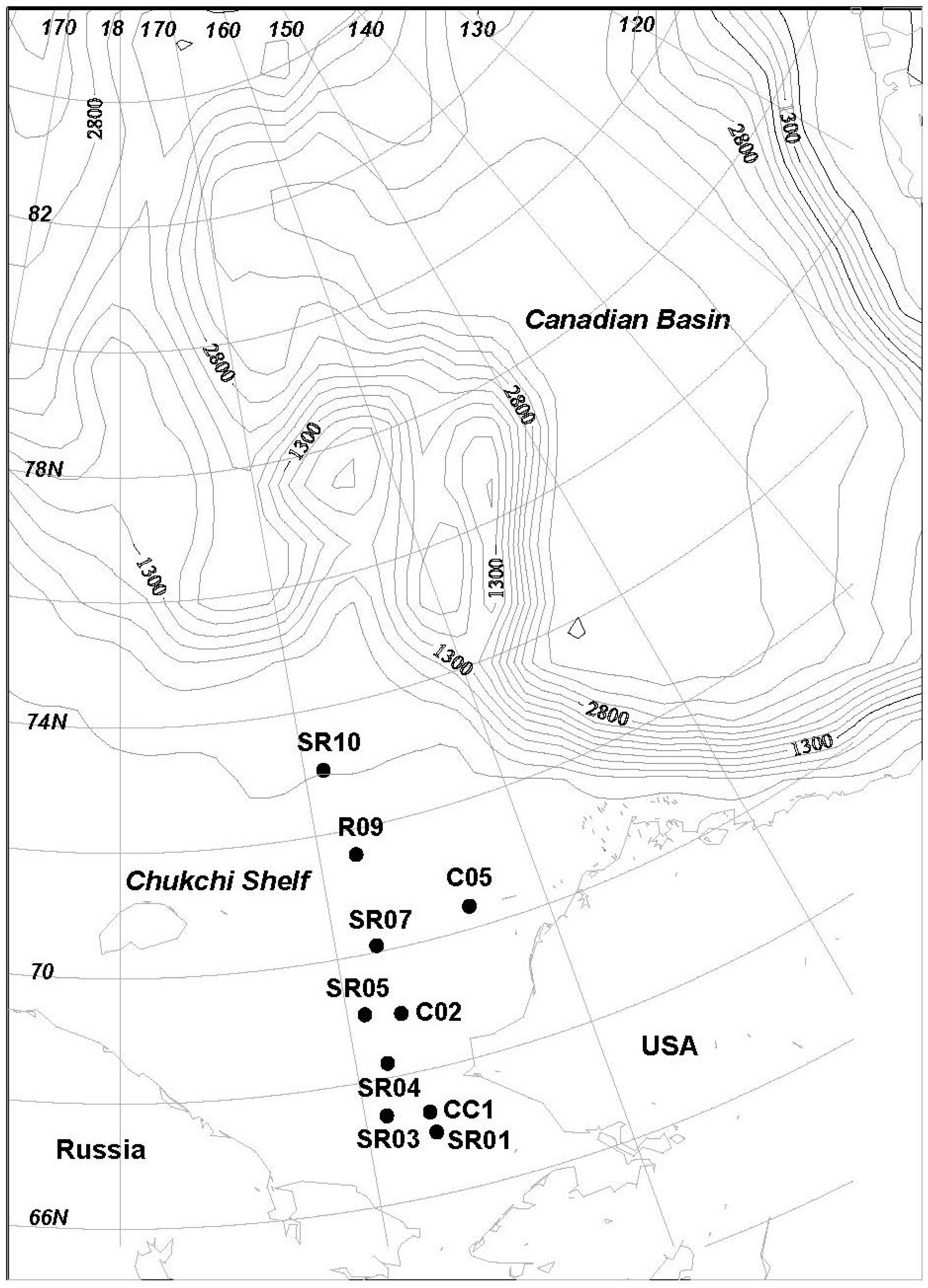
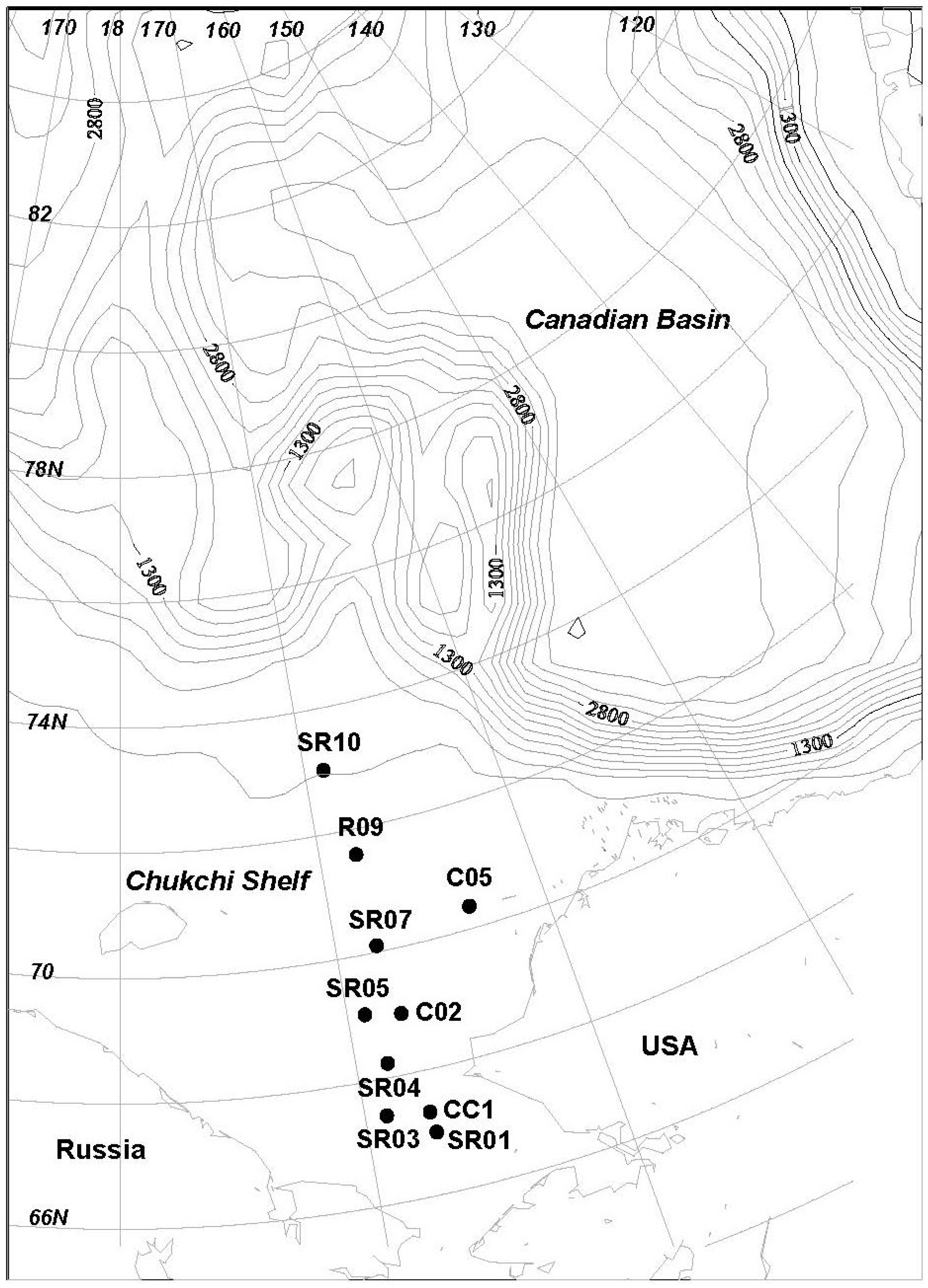
2.1. Isolation of Actinobacteria from the Chukchi Shelf Marine Sediment
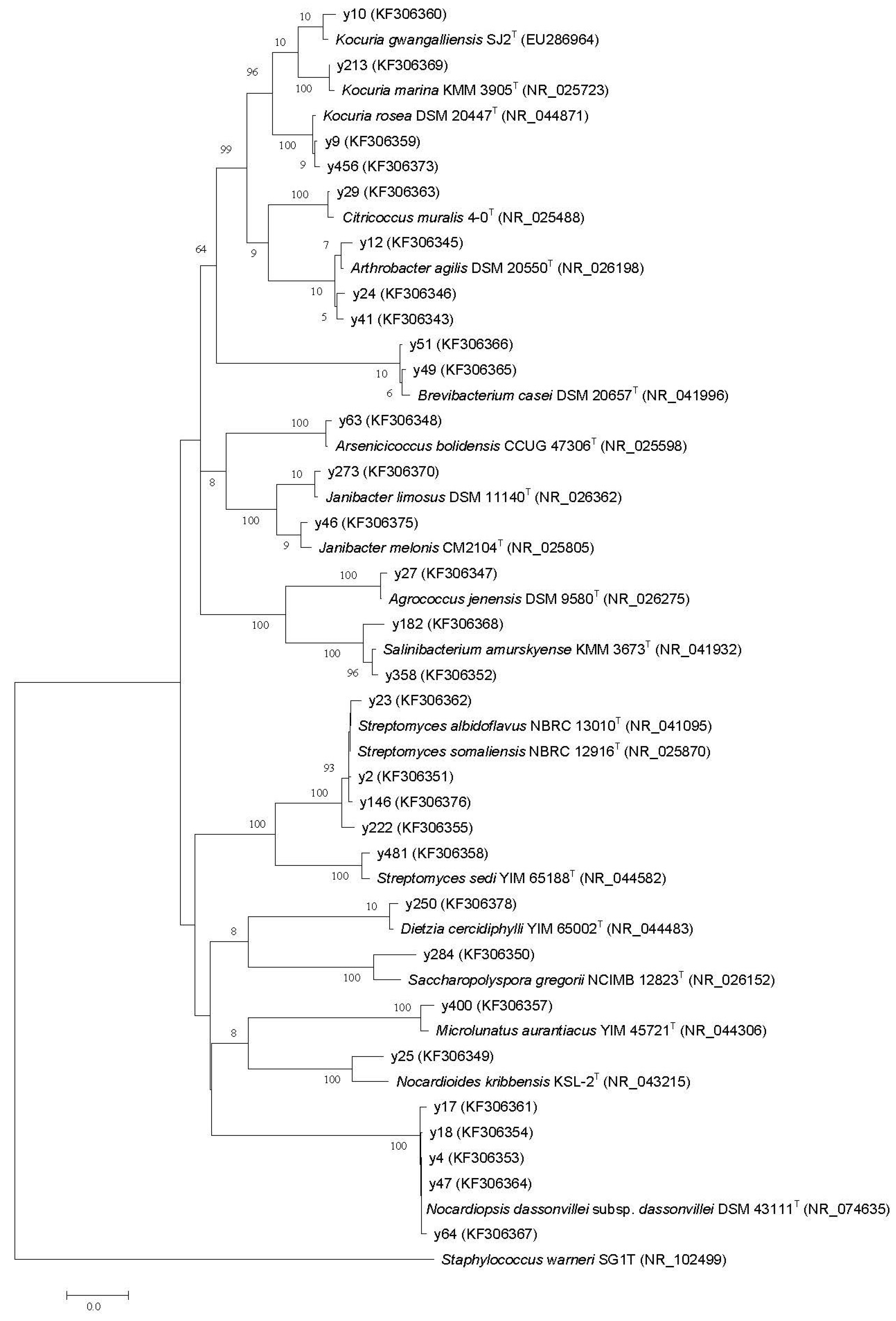
2.2. Phylogenetic Diversity

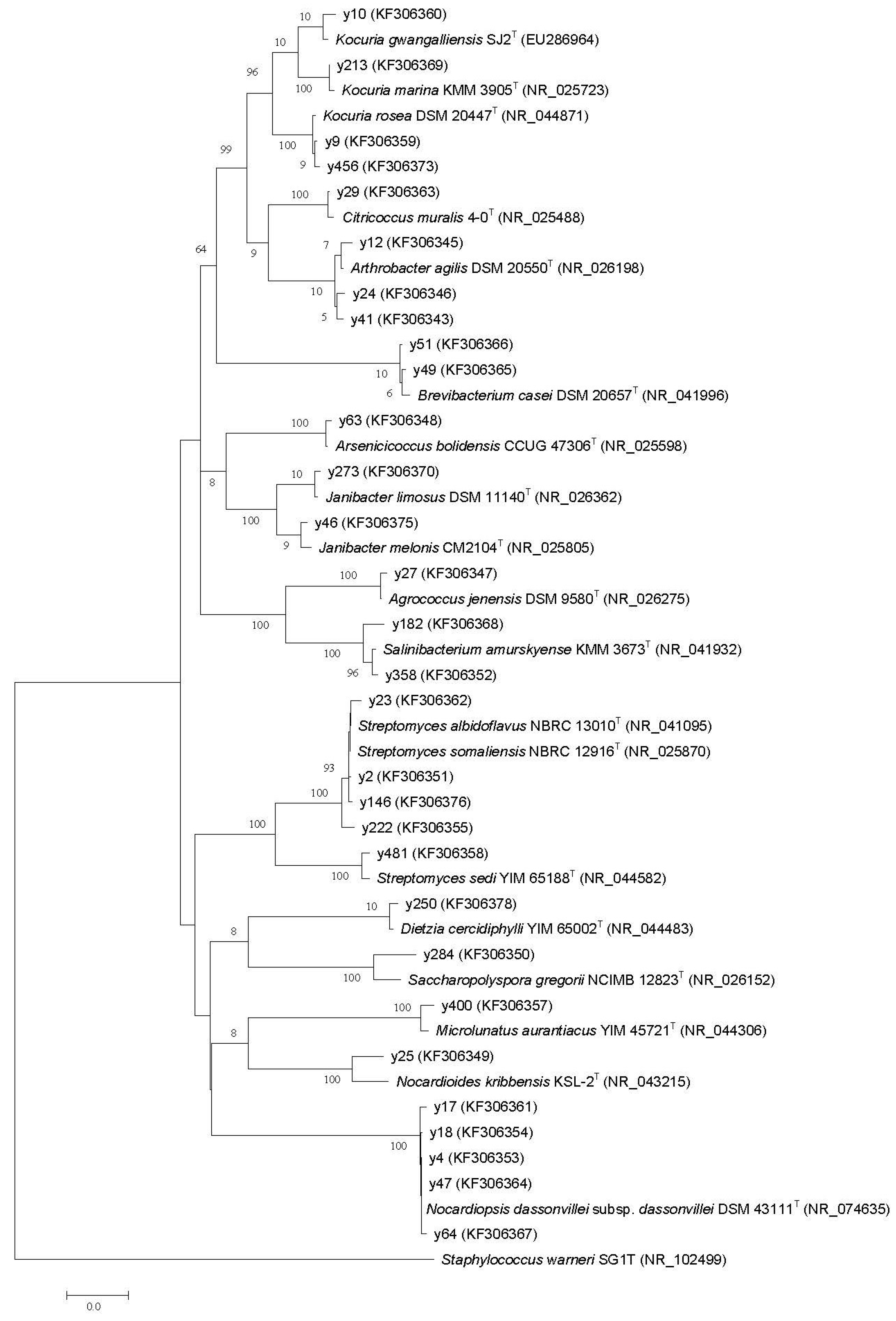{kind=link}
{kind=link}
| Genus | Isolate | The Closest Type Strain (% Identity) | Seawater Requirement | Biosynthetic Genes | Antimicrobial Activities * | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PKS I | PKS II | NRPS | phzE | dTGD | Halo | CYP | B. subtilis | C. albicans | ||||
| Brevibacteriaceae | ||||||||||||
| Brevibacterium | y49 | Brevibacterium casei NCDO 2048T (99.3) | − | + | − | + | − | + | + | − | − | − |
| y51 | Brevibacterium casei NCDO 2048T (99.4) | − | + | − | + | − | + | − | − | − | − | |
| Dietziaceae | ||||||||||||
| Dietzia | y250 | Dietzia cercidiphylli YIM 65002T (99.6) | + | + | − | + | − | − | + | − | − | − |
| Intrasporangiaceae | ||||||||||||
| Arsenicicoccus | y63 | Arsenicicoccus bolidensis CCUG 47306T (99.8) | − | − | − | − | − | + | + | − | − | − |
| Janibacter | y46 | Janibacter melonis CM2104T (99.5) | − | + | + | − | − | + | − | − | − | − |
| y273 | Janibacter limosus DSM 11140T (99.7) | − | + | − | + | − | + | + | + | − | − | |
| Microbacteriaceae | ||||||||||||
| Agrococcus | y27 | Agrococcus jenensis DSM 9580T (99.7) | − † | − | + | − | − | + | − | − | − | − |
| Salinibacterium | y182 | Salinibacterium amurskyense KMM 3673T (99.0) | − | + | − | − | + | + | − | + | − | − |
| y358 | Salinibacterium amurskyense KMM 3673T (99.7) | − † | + | − | − | − | + | − | − | − | − | |
| Micrococcaceae | ||||||||||||
| Arthrobacter | y12 | Arthrobacter agilis DSM 20550T (99.5) | − | + | − | + | − | + | − | − | − | − |
| y24 | Arthrobacter agilis DSM 20550T (99.3) | − † | + | − | − | − | + | − | + | − | − | |
| y41 | Arthrobacter agilis DSM 20550T (99.2) | − † | + | + | − | − | − | + | + | − | − | |
| Citricoccus | y29 | Citricoccus muralis 4-0T (99.7) | − † | + | − | − | − | + | + | + | − | − |
| Kocuria | y9 | Kocuria rosea DSM 20447T (99.7) | + | + | − | − | − | − | + | − | − | − |
| y456 | Kocuria rosea DSM 20447T (99.7) | − | + | − | + | − | + | + | − | − | − | |
| y10 | Kocuria gwangalliensis SJ2T (99.4) | − † | + | − | − | − | − | + | − | − | − | |
| y213 | Kocuria marina KMM 3905T (99.9) | − | + | − | − | + | − | − | − | − | − | |
| Nocardioidaceae | ||||||||||||
| Nocardioides | y25 | Nocardioides kribbensis KSL-2T (97.8) | + | − | − | − | − | + | + | − | − | − |
| Nocardiopsaceae | ||||||||||||
| Nocardiopsis | y4 | Nocardiopsis dassonvillei subsp. dassonvillei DSM 43111T (ABUI01000017) | + | − | + | + | + | + | − | − | + | − |
| y17 | Nocardiopsis dassonvillei subsp. dassonvillei DSM 43111T (99.7) | − † | − | + | + | − | + | + | − | + | + | |
| y18 | Nocardiopsis dassonvillei subsp. dassonvillei DSM 43111T (99.8) | + | − | + | + | + | + | − | − | + | + | |
| y47 | Nocardiopsis dassonvillei subsp. dassonvillei DSM 43111T (99.9) | − † | − | + | − | + | + | − | − | + | − | |
| y64 | Nocardiopsis dassonvillei subsp. dassonvillei DSM 43111T (99.8) | − † | − | + | − | − | + | − | − | + | + | |
| Propionibacteriaceae | ||||||||||||
| Microlunatus | y400 | Microlunatus aurantiacus YIM 45721T (99.2) | − | + | − | − | − | + | + | + | + | − |
| Pseudonocardiaceae | ||||||||||||
| Saccharopolyspora | y284 | Saccharopolyspora gregorii NCIMB 12823T (97.8) | − | − | + | − | − | + | − | − | − | − |
| Streptomycetaceae | ||||||||||||
| Streptomyces | y2 * | Streptomyces somaliensis NBRC 12916T (99.9) | − | + | − | + | − | + | − | + | + | + |
| y23 | Streptomyces somaliensis NBRC 12916T (99.7) | − | + | − | + | − | − | − | + | + | + | |
| y146 * | Streptomyces somaliensis NBRC 12916T (99.9) | − | + | − | − | + | + | − | + | + | + | |
| y222 | Streptomyces albidoflavus DSM 40455T (99.5) | − | + | + | + | − | + | − | + | + | + | |
| y481 | Streptomyces sedi YIM 65188T (99.4) | + | + | + | + | − | + | − | − | + | − | |
2.3. Antimicrobial Activities and Detection of Biosynthetic Genes
3. Experimental Section
3.1. Sediments Sample
| Samples | Water Depth (m) | Color | Composition | Benthos |
|---|---|---|---|---|
| CC1 | 45 | Light grey | Silty clay | Bivalves |
| C02 | 41 | Grey | Clay | Conch |
| C05 | 26 | Cinerous | Fine sand | Crab, conch, sand dollar, Polychaeta |
| SR01 | 41.5 | Cinerous | Silty sand | Conch, bivalve |
| SR03 | 50.7 | Cinerous | Clay | Polychaeta, conch, sipunculoid |
| SR04 | 48 | Grey | Silty clay | None |
| SR05 | 46.7 | Cinerous | Silty clay | Conch |
| SR07 | 29.7 | Cinerous | Silty mud | None |
| R09 | 43.5 | Grey | Mud soil | None |
| SR10 | 69.6 | Cinerous | Silty clay | Sea star, conch |
3.2. Isolation of Actinobacterial Strains
3.3. Dereplication by Rep-Polymerase Chain Reaction (PCR)
3.4. Seawater Requirements
3.5. 16S rRNA Gene Amplification and Phylogenetic Analysis
3.6. Antimicrobial Activity Testing by Agar Diffusion (Inhibition Zones)
3.7. Amplification of Biosynthetic Gene Fragments
| Gene | Primer | Length (bp) | Reference |
|---|---|---|---|
| PKS I (KSMA-F, KSMB-R) | 5′-TSGCSATGGACCCSCAGCAG-3′ 5′-CCSGTSCCGTGSGCCTCSAC-3′ | 700 | [47] |
| PKS II (540F, 1100R) | 5′-GGITGCACSTCIGGIMTSGAC-3′ 5′-CCGATSGCICCSAGIGAGTG-3′ | 554 | [48] |
| NRPS (A3F, A7R) | 5′-GCSTACSYSATSTACACSTCSGG-3′ 5′-SASGTCVCCSGTSCGGTAS-3′ | 700 | [49] |
| PhzE (phzEf, phzEr) | 5′-GAAGGCGCCAACTTCGTYATCAA-3′ 5′-GCCYTCGATGAAGTACTCGGTGTG-3′ | 450 | [50] |
| Halo (FW, RV) | 5′-TTCCCSCGSTACCASATCGGSGAG-3′ 5′-GSGGGATSWMCCAGWACCASCC-3′ | 500 | [32] |
| dTGD (dTGD-1, dTGD-2) | 5′-GSGGSGSSGCSGGSTTCATSGG-3′ 5′-GGGWRCTGGYRSGGSCCGTAGTTG-3′ | 600 | [51] |
| CYP (PEH-1, PEH-2) | 5′-TGGATCGGCGACGACCGSVYCGT-3′ 5′-CCGWASAGSAYSCCGTCGTACTT-3′ | 350 | [34] |
4. Conclusions
Abbreviations
| DDC | Dispersion and Differential Centrifugation |
| BSA | Albumin from Bovine Serum |
| NCBI | National Center of Biotechnology Information |
| BLASTN | Basic Local Alignment Search Tool of Nucleic acid |
| PKS | PolyKetide Synthase |
| NRPS | Nonribosomal Peptide Synthase |
| phzE | aminodeoxyisochorismate synthase |
| dTGD | dTDP-Glucose-4, 6-Dehydratase |
| Halo | Halogenase |
| CYP | Cytochrome P450 hydroxylase |
| FADH | Flavin Adenine Dinucleotide Hydrogen carrier |
Acknowledgments
Conflicts of Interest
References
- Ventura, M.; Canchaya, C.; Tauch, A.; Chandra, G.; Fitzgerald, G.F.; Chater, K.F.; van Sinderen, D. Genomics of Actinobacteria: Tracing the evolutionary history of an ancient phylum. Microbiol. Mol. Biol. Rev. 2007, 71, 495–548. [Google Scholar] [CrossRef]
- Bérdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef]
- Fiedler, H.-P.; Bruntner, C.; Bull, A.T.; Ward, A.C.; Goodfellow, M.; Potterat, O.; Puder, C.; Mihm, G. Marine actinomycetes as a source of novel secondary metabolites. Antonie van Leeuwenhoek 2005, 87, 37–42. [Google Scholar] [CrossRef]
- Lam, K.S. Discovery of novel metabolites from marine actinomycetes. Curr. Opin. Microbiol. 2006, 9, 245–251. [Google Scholar] [CrossRef]
- Bull, A.T.; Stach, J.E.M. Marine actinobacteria: New opportunities for natural product search and discovery. Trends Microbiol. 2007, 15, 491–499. [Google Scholar] [CrossRef]
- Stach, J.E.M.; Bull, A.T. Estimating and comparing the diversity of marine actinobacteria. Antonie van Leeuwenhoek 2005, 87, 3–9. [Google Scholar] [CrossRef]
- Goodfellow, M.; Fiedler, H.-P. A guide to successful bioprospecting: Informed by actinobacterial systematics. Antonie van Leeuwenhoek 2010, 98, 119–142. [Google Scholar] [CrossRef]
- Fenical1, W.; Jensen, P.R. Developing a new resource for drug discovery: Marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2, 666–673. [Google Scholar] [CrossRef]
- Olano, C.; Méndez, C.; Salas, J.A. Antitumor compounds from marine actinomycetes. Mar. Drugs 2009, 7, 210–248. [Google Scholar] [CrossRef]
- Fattorusso, E.; Taglialatela-Scafati, O. Marine antimalarials. Mar. Drugs 2009, 7, 130–152. [Google Scholar] [CrossRef]
- Rahman, H.; Austin, B.; Mitchell, W.J.; Morris, P.C.; Jamieson, D.J.; Adams, D.R.; Spragg, A.M.; Schweizer, M. Novel anti-infective compounds from marine bacteria. Mar. Drugs 2010, 8, 498–518. [Google Scholar] [CrossRef]
- Xiong, Z.-Q.; Wang, J.-F.; Hao, Y.-Y.; Wang, Y. Recent advances in the discovery and development of marine microbial natural products. Mar. Drugs 2013, 11, 700–717. [Google Scholar] [CrossRef]
- Weyland, H. Distribution of Actinomycetes on the Sea Floor. In Actinomycetes: Proceedings of the Fourth International Symposium on Actinomycete Biology, Cologne, 3–7 September 1979; Gustav Fischer Verlag: Stuttgart, Germany, 1981; pp. 185–193. [Google Scholar]
- Yu, Y.; Li, H.; Zeng, Y.; Chen, B. Isolation and phylogenetic assignation of actinomycetes in the marine sediments from the Arctic Ocean. Acta Oceanol. Sin. 2005, 24, 135–142. [Google Scholar]
- Bredholdt, H.; Galatenko, O.A.; Engelhardt, K.; Tjaervik, E.; Terekhova, L.P.; Zotchev, S.B. Rare actinomycete bacteria from the shallow water sediments of the Trondheim fjord, Norway: Isolation, diversity and biological activity. Environ. Microbiol. 2007, 9, 2756–2764. [Google Scholar] [CrossRef]
- Bredholdt, H.; Tjaervik, E.; Johnsen, G.; Zotchev, S.B. Actinomycetes from sediments in the Trondheim fjord, Norway: Diversity and biological activity. Mar. Drugs 2008, 6, 12–24. [Google Scholar] [CrossRef]
- Jørgensen, H.; Fjærvik, E.; Hakvåg, S.; Bruheim, P.; Bredholt, H.; Klinkenberg, G.; Ellingsen, T.E.; Zotchev, S.B. Candicidin biosynthesis gene cluster is widely distributed among Streptomyces spp. isolated from the sediments and the neuston layer of the Trondheim fjord, Norway. Appl. Environ. Microbiol. 2009, 75, 3296–3303. [Google Scholar] [CrossRef]
- Jørgensen, H.; Degnes, K.F.; Dikiy, A.; Fjærvik, E.; Klinkenberg, G.; Zotchev, S.B. Insights into the evolution of macrolactam biosynthesis through cloning and comparative analysis of the biosynthetic gene cluster for a novel macrocyclic lactam, ML-449. Appl. Environ. Microbiol. 2010, 76, 283–293. [Google Scholar] [CrossRef]
- Engelhardt, K.; Degnes, K.F.; Kemmler, M.; Bredholt, H.; Fjærvik, E.; Klinkenberg, G.; Sletta, H.; Ellingsen, T.E.; Zotchev, S.B. Production of a new thiopeptide antibiotic, TP-1161, by a marine Nocardiopsis sspecies. Appl. Environ. Microbiol. 2010, 76, 4969–4976. [Google Scholar] [CrossRef] [Green Version]
- Hakvåg, S.; Fjærvik, E.; Josefsen, K.D.; Ian, E.; Ellingsen, T.E.; Zotchev, S.B. Characterization of Streptomyces spp. isolated from the sea surface microlayer in the Trondheim fjord, Norway. Mar. Drugs 2008, 6, 620–635. [Google Scholar] [CrossRef]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O.; Krichevsky, M.I.; Moore, L.H.; Moore, W.E.C.; Murray, R.G.E.; Stackebrandt, E.; et al. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Evol. Microbiol. 1987, 37, 463–464. [Google Scholar]
- Stackebrandt, E.; Goebel, B.M. Taxonomic note: A place for DNA–DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar]
- Stackebrandt, E.; Ebers, J. Taxonomic parameters revisited: Tarnished gold standards. Microbiol. Today 2006, 33, 152–155. [Google Scholar]
- Wang, Y.X.; Cai, M.; Zhi, X.Y.; Zhang, Y.Q.; Tang, S.K.; Xu, L.H.; Cui, X.L.; Li, W.J. Microlunatus aurantiacus sp. nov., a novel actinobacterium isolated from a rhizosphere soil sample. Int. J. Syst. Evol. Microbiol. 2008, 58, 1873–1877. [Google Scholar] [CrossRef]
- Chen, L.; Li, S.U.; Xue, Y.F. Characterization of an alkaliphilic biosurfactant- producing strain of Dietzia. Microbiol. China 2012, 39, 1573–1579. [Google Scholar]
- Ratnakar, N.A.; Dirk, S.; Rolf, H.; Siegmund, L.; Irene, W.D.; Hartmut, L. Helquinoline, a new tetrahydroquinoline antibiotic from Janibacter limosus Hel 1+. J. Antibiot. 2004, 57, 17–23. [Google Scholar] [CrossRef]
- Rojas, J.L; Martín, J.; Tormo, J.R.; Vicente, F.; Brunati, M.; Ciciliato, I.; Losi, D.; Van, T.S.; Mergaert, J.; Swings, J.; et al. Bacterial diversity from benthic mats of Antarctic lakes as a source of new bioactive metabolites. Mar. Genomics 2009, 2, 33–41. [Google Scholar] [CrossRef]
- Fu, P.; Liu, P.; Gong, Q.H.; Wang, Y.; Wang, P.; Zhu, W.M. α-Pyrones from the marine-derived actinomycete Nocardiopsis dassonvillei subsp. dassonvillei XG-8-1. RSC Adv. 2013, 3, 20726–20731. [Google Scholar]
- Augustine, S.K.; Bhavsar, S.P.; Kapadnis, B.P. A non-polyene antifungal antibiotic from Streptomyces albidoflavus PU 23. J. Biosci. 2005, 30, 201–211. [Google Scholar] [CrossRef]
- Yan, L.L.; Han, N.N.; Zhang, Y.Q.; Yu, L.Y.; Chen, J.; Wei, Y.Z.; Li, Q.P.; Tao, L.; Zheng, G.H.; Yang, S.E.; et al. Antimycin A18 produced by an endophytic Streptomyces albidoflavus isolated from a mangrove plant. J. Antibiot. 2010, 63, 259–261. [Google Scholar] [CrossRef]
- Yuan, M.; Dong, N.; Li, H.R.; Yu, Y. Sequence-based analysis of secondary-metabolite biosynthesis in actinobacteria isolated from Chukchi shelf marine sediments. Adv. Polar Sci. 2014. submitted for publication. [Google Scholar]
- Hornung, A.; Bertazzo, M.; Dziarnowski, A.; Schneider, K.; Welzel, K.; Wohlert, S.E.; Holzenkämpfer, M.; Nicholson, G.J.; Bechthold, A.; Süssmuth, R.D.; et al. A Genomic screening approach to the structure-guided identification of drug candidates from natural sources. Chembiochem 2007, 8, 757–766. [Google Scholar] [CrossRef]
- Stockmann, M.; Piepersberg, W. Gene probes for the detection of 6-deoxyhexose metabolism in secondary metabolite-producing streptomycetes. FEMS Microbiol. Lett. 1992, 90, 185–189. [Google Scholar] [CrossRef]
- Lee, M.Y.; Myeong, J.S.; Park, H.J.; Han, K.; Kim, E.S. Isolation and partial characterization of a cryptic polyene gene cluster in Pseudonocardia autotrophica. J. Ind. Microbiol. Biotechnol. 2006, 33, 84–87. [Google Scholar] [CrossRef]
- Dairi, T.; Hamano, Y.; Igarashi, Y.; Furumai, T.; Oki, T. Cloning and nucleotide sequence of the putative polyketide synthase genes for pradimicin biosynthesis from Actinomadura hibisca. Biosci. Biotechnol. Biochem. 1997, 61, 1445–1453. [Google Scholar] [CrossRef]
- Kallifidas, D.; Kang, H.S.; Brady, S.F. Tetarimycin A, an MRSA-Active antibiotic identified through induced expression of environmental DNA gene clusters. J. Am. Chem. Soc. 2012, 134, 19552–19555. [Google Scholar] [CrossRef]
- Clardy, J.M.; Fischbach, A.; Walsh, C.T. New antibiotics from bacterial natural products. Nat. Biotechnol. 2006, 24, 1541–1550. [Google Scholar] [CrossRef]
- Hopkins, D.W.; McNaughton, S.J.; O’Donnell, A.G. A dispersion and differential centrifugation technique for representatively sampling microorganisms from soil. Soil Biol. Biochem. 1991, 23, 217–225. [Google Scholar] [CrossRef]
- Versalovic, J.; Schneider, M.; de Bruijn, F.J.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequencebased polymerase chain reaction. Methods Mol. Cell. Biol. 1994, 5, 24–40. [Google Scholar]
- Rademaker, J.L.W.; Hoste, B.; Louws, F.J.; Kersters, K.; Swings, J.; Vauterin, L.; Vauterin, P.; de Bruijn, F.J. Comparison of AFLP and rep-PCR genomic fingerprinting with DNA–DNA homology studies: Xanthomonas as a model system. Int. J. Syst. Evol. Microbiol. 2000, 50, 665–677. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Kim, O.S.; Cho, Y.J.; Lee, K.; Yoon, S.H.; Kim, M.; Na, H.; Park, S.C.; Jeon, Y.S.; Lee, J.H.; Yi, H.; et al. Introducing EzTaxon-e: A prokaryotic 16S rRNA Gene sequence database with phylotypes that represent uncultured species. Int. J. Syst. Evol. Microbiol. 2012, 62, 716–721. [Google Scholar] [CrossRef]
- EzTaxon Server. version 2.1. Available online: http://eztaxon-e ezbiocloud.net (accessed on 1 November 2013).
- NCBI database. Available online: http://www.ncbi.nlm.nlh.gov (accessed on 1 November 2013).
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 10, 2731–2739. [Google Scholar]
- Schneemann, I.; Nagel, K.; Kajahn, I.; Labes, A.; Wiese, J.; Imhoff, J.F. Comprehensive investigation of marine actinobacteria associated with the sponge Halichondria panacea. Appl. Environ. Microbiol. 2010, 76, 3702–3714. [Google Scholar] [CrossRef] [Green Version]
- Izumikawa, M.; Murata, M.; Tachibana, K.; Ebizuka, Y.; Fujii, I. Cloning of modular type I polyketide synthase genes from salinomycin producing strain of Streptomyces albus. Bioorg. Med. Chem. 2003, 11, 3401–3405. [Google Scholar] [CrossRef]
- Wawrik, B.; Kerkhof, L.; Zylstra, G.J.; Kukor, J.J. Identification of unique type II polyketide synthase genes in soil. Appl. Environ. Microbiol. 2005, 71, 2232–2238. [Google Scholar] [CrossRef]
- Ayuso-Sacido, A.; Genilloud, O. New PCR primers for the screening of NRPS and PKS-I systems in actinomycetes: Detection and distribution of these biosynthetic gene sequences in major taxonomic groups. Microb. Ecol. 2005, 49, 10–24. [Google Scholar] [CrossRef]
- Schneemann, I.; Wiese, J.; Kunz, A.L.; Imhoff, J.F. Genetic approach for the fast discovery of phenazine producing bacteria. Mar. Drugs 2011, 9, 772–789. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Li, T.; Wang, Y.G.; Xia, H. Identification and functional analysis of dTDP-glucose-4, 6-dehydratase gene and its linked gene cluster in an aminoglycoside antibiotics producer of Streptomyces tenebrarius H6. Curr. Microbiol. 2004, 49, 99–107. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yuan, M.; Yu, Y.; Li, H.-R.; Dong, N.; Zhang, X.-H. Phylogenetic Diversity and Biological Activity of Actinobacteria Isolated from the Chukchi Shelf Marine Sediments in the Arctic Ocean. Mar. Drugs 2014, 12, 1281-1297. https://doi.org/10.3390/md12031281
Yuan M, Yu Y, Li H-R, Dong N, Zhang X-H. Phylogenetic Diversity and Biological Activity of Actinobacteria Isolated from the Chukchi Shelf Marine Sediments in the Arctic Ocean. Marine Drugs. 2014; 12(3):1281-1297. https://doi.org/10.3390/md12031281
Chicago/Turabian StyleYuan, Meng, Yong Yu, Hui-Rong Li, Ning Dong, and Xiao-Hua Zhang. 2014. "Phylogenetic Diversity and Biological Activity of Actinobacteria Isolated from the Chukchi Shelf Marine Sediments in the Arctic Ocean" Marine Drugs 12, no. 3: 1281-1297. https://doi.org/10.3390/md12031281