Antimicrobial Peptides from Marine Proteobacteria

 ,
,
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum Proteobacteria | |||||
|---|---|---|---|---|---|
| Class | α-Proteobacteria | Class | β-Proteobacteria | ||
| Orders | Caulobacterales | Rhizobiales | Orders | Burkholderiales | Neisseriales |
| Kiloniellales | Rhodobacterales | Ferritrophicales | Nitrosomonadales | ||
| Kopriimonadales | Rhodospirillales | Gallionellales | Procabacteriales | ||
| Kordiimonadales | Rickettsiales | Hydrogenophilales | Rhodocyclales | ||
| Magnetococcales | Sneathiellales | Methylophilales | Unclassified taxa | ||
| Parvularculales | Sphingomonadales | ||||
| Unclassified taxa | |||||
| Class | γ-Proteobacteria | Class | δ-Proteobacteria | ||
| Orders | Acidithiobacillales | Orbales | Orders | Bdellovibrionales | Desulfuromonadales |
| Aeromonadales | Pasteurellales | Desulfarculales | Myxococcales | ||
| Alteromonadales | Pseudomonadales | Desulfobacterales | Syntrophobacterales | ||
| Cardiobacteriales | Salinisphaerales | Desulfovibrionales | Unclassified taxa | ||
| Chromatiales | Thiotrichales | Desulfurellales | |||
| Enterobacteriales | Vibrionales | ||||
| Legionellales | Xanthomonadales | ||||
| Methylococcales | Unclassified taxa | ||||
| Oceanospirillales | |||||
| Class | ε-Proteobacteria | Class | ζ-Proteobacteria (candidatus) | ||
| Orders | Campylobacterales | Order | Mariprofundales | ||
| Nautiliales | |||||
| Unclassified taxa | |||||
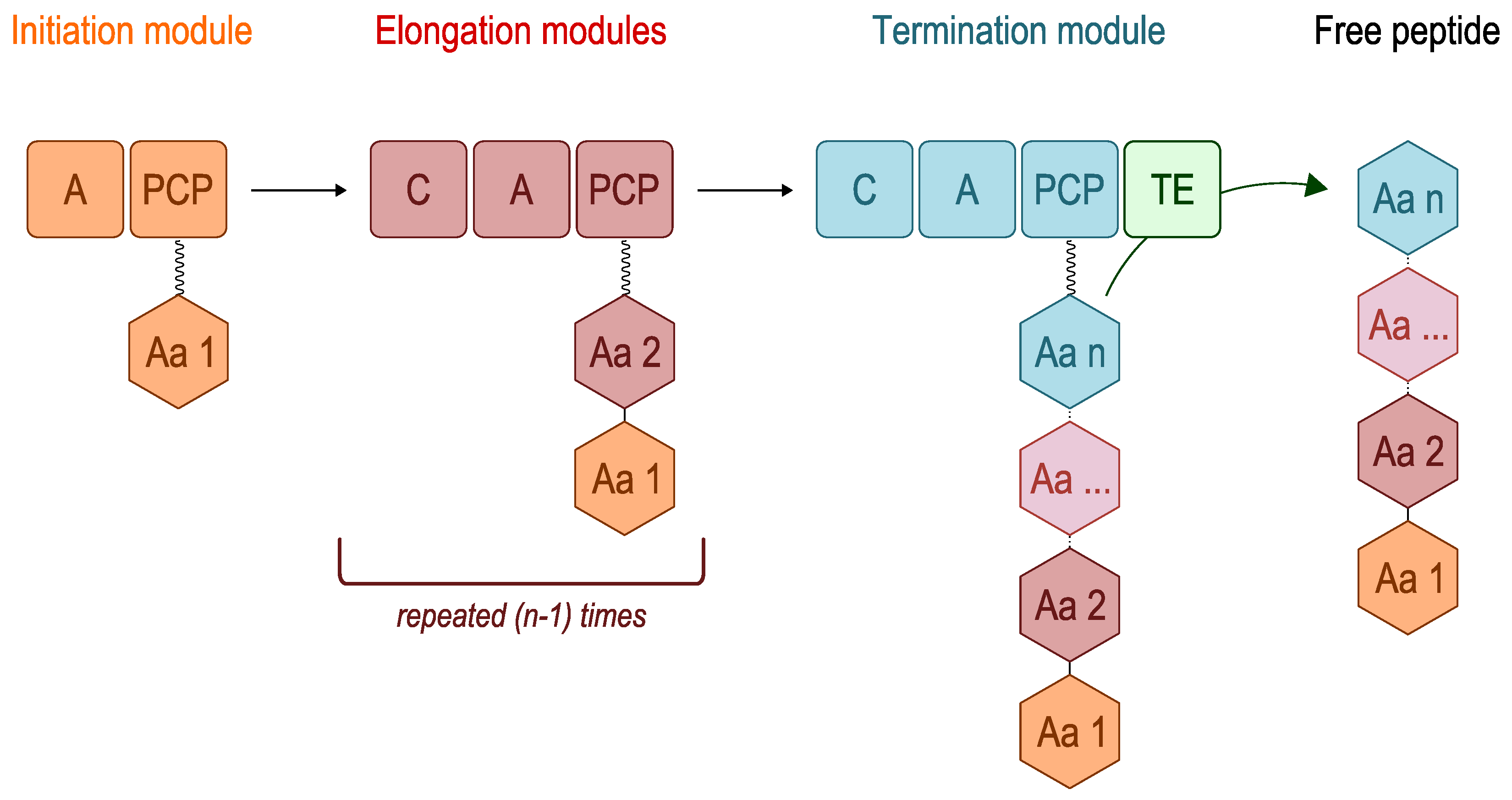
2. Overview of Nonribosomal Peptide Synthetases (NRPSs)
| Class/Order | Producers | Compounds | Activity | Biosynthesis |
|---|---|---|---|---|
| α-Proteobacteria | ||||
| Rhodobacterales | Oceanibulbus indolifex | Cyclic dipeptides | Antibacterial | Unknown |
| Phaeobacter sp. | Indigoidine | Antibacterial (Gram-negative) | NRPS | |
| γ-Proteobacteria | ||||
| Vibrionales | Vibrio sp. | Andrimid Moiramide (pseudopeptides) | Antibacterial | Hybrid NRPS-PKS (<100 kDa) |
| Kahalalides (depsipeptides) | Antifungal | Unknown | ||
| Photobacterium halotolerans | Holomycin (pyrrothine) | Bacteriostatic | NRPS described in Streptomyces clavuligerus | |
| Photobacterium sp. | Unnarmicins (depsipeptides) | Antibacterial (Gram-negative) | Unknown | |
| Ngercheumicins (depsipeptides) | Antibacterial (Gram-negative) | Unknown | ||
| Solonamides (depsipeptides) | Antibacterial (Gram-negative) | Unknown | ||
| Alteromonadales | Pseudoalteromonas sp. | Thiomarinols (pyrrothine) | Antibacterial | NRPS (pTLM1) |
| Cyclopeptides | Antibacterial | Unknown | ||
| Lipopeptides | Antibacterial | NRPS | ||
| Pseudomonadales | Pseudomonas sp. | Massetolides (cyclic lipopeptides) | Antimycobacterial | NRPS described in Pseudomonas fluorescens |
| δ-Proteobacteria | ||||
| Myxococcales | Myxococcus fulvus | Myxovalargins (polyketide/polypeptide hybrid) | Antibacterial | Unknown |
| Althiomycin (polyketide/polypeptide hybrid) | Antibacterial | Hybrid NRPS-PKS described in Mycococcus xanthus | ||
| Myxothiazols | Antifungal | NRPS described in Stigmatella aurantiaca | ||
| Paraliomyxa miuraensis | Miuraenamides (depsipeptides) | Antifungal | Unknown |
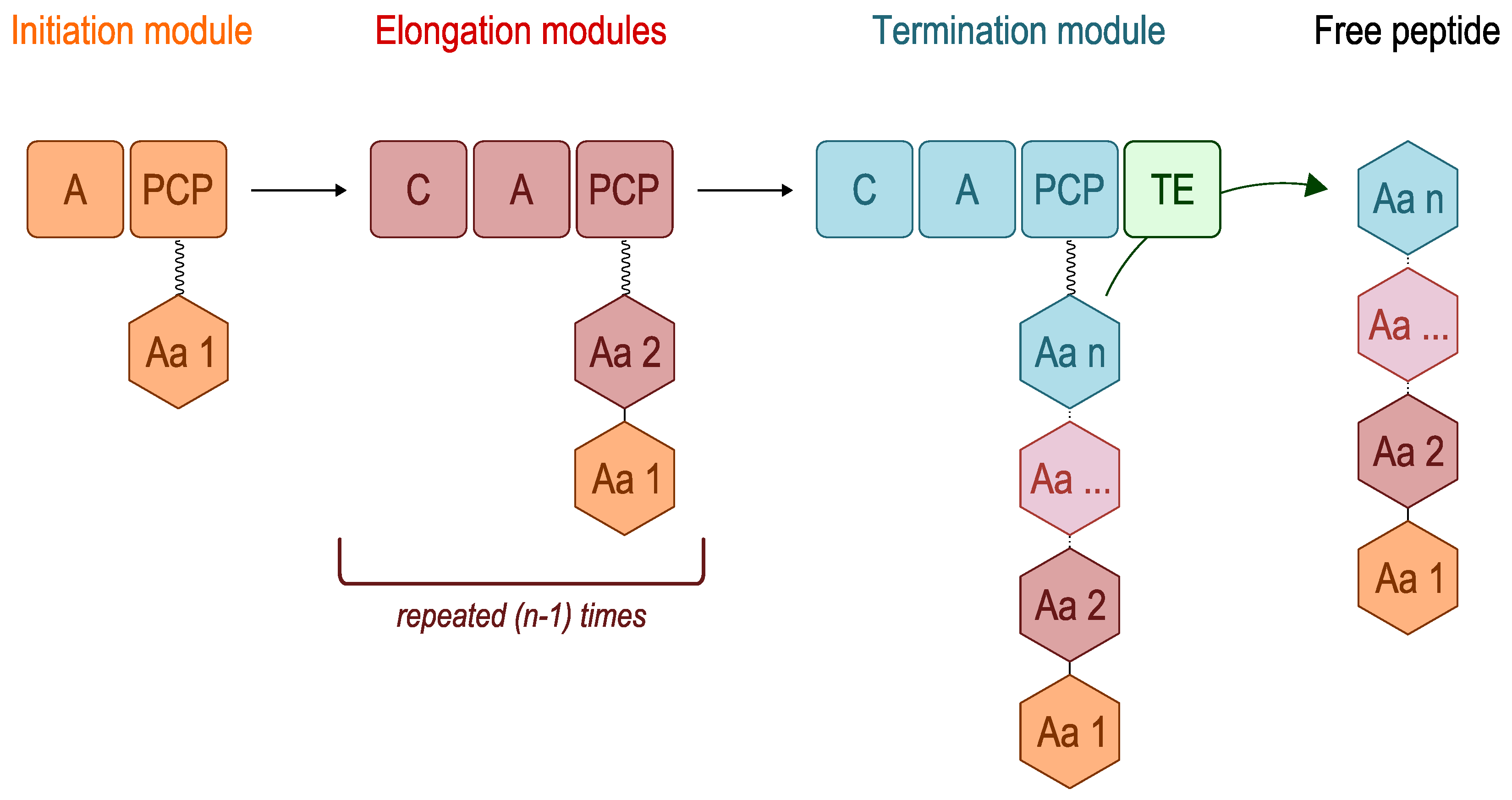
2.1. Genome-Based Strategy
2.2. Proteomics Strategy
3. Antimicrobial NRPs from Marine Proteobacteria
3.1. α-Proteobacteria
3.1.1. Cyclic Dipeptides
3.1.2. Indigoidine
3.2. γ-Proteobacteria
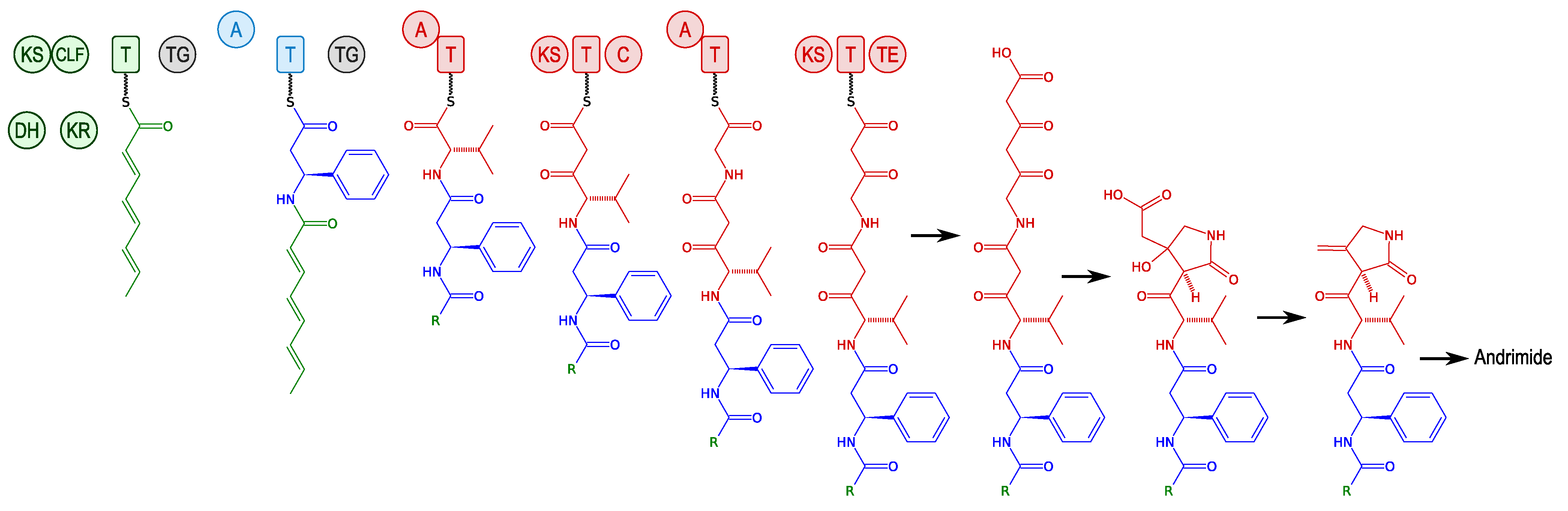
3.2.1. Andrimid and Moiramide
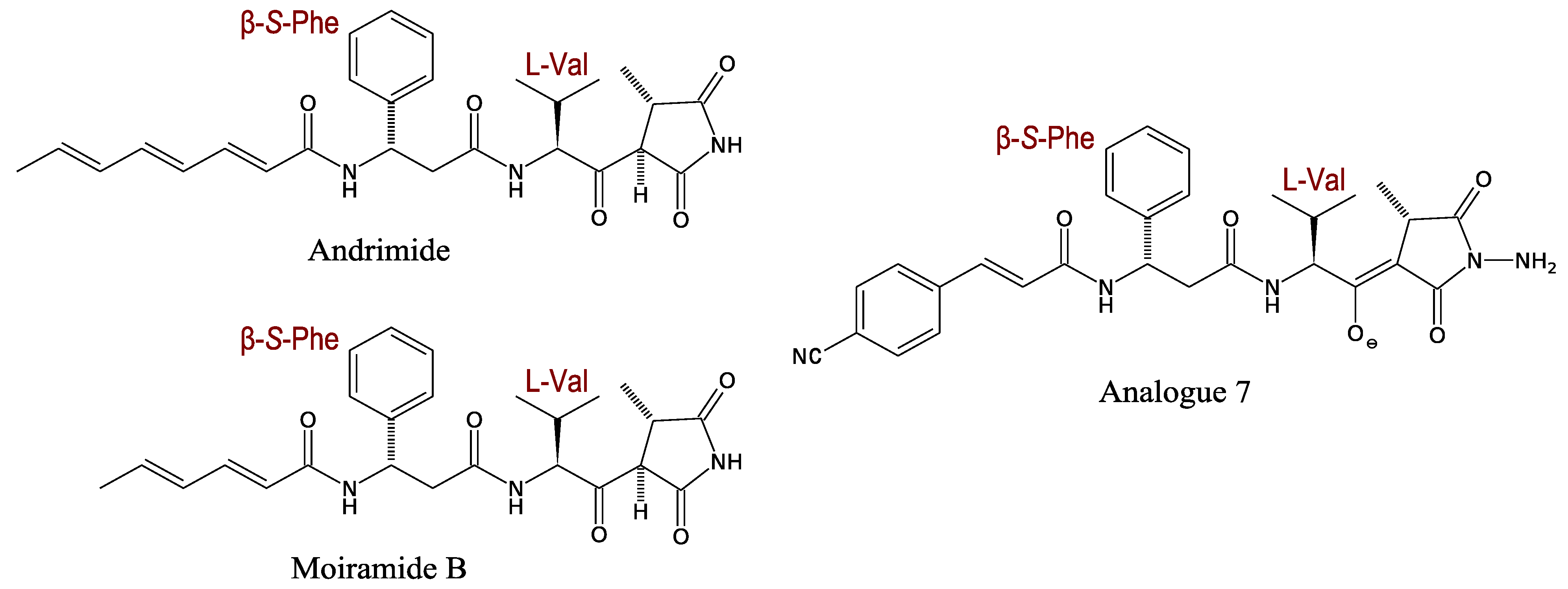
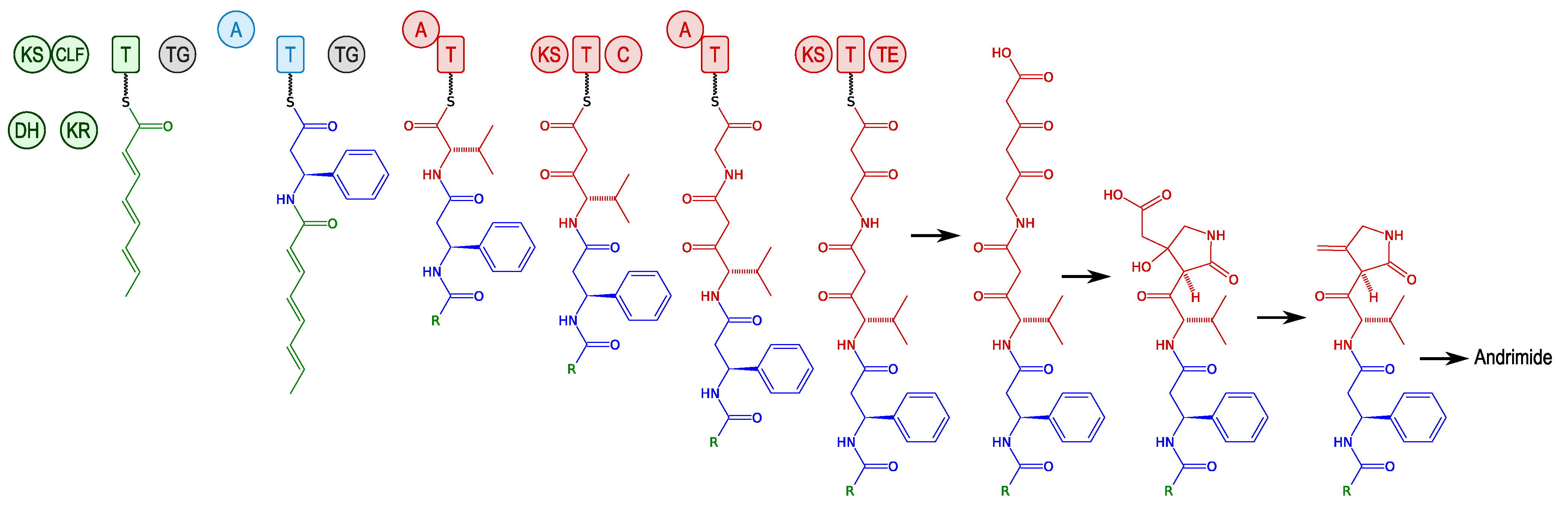
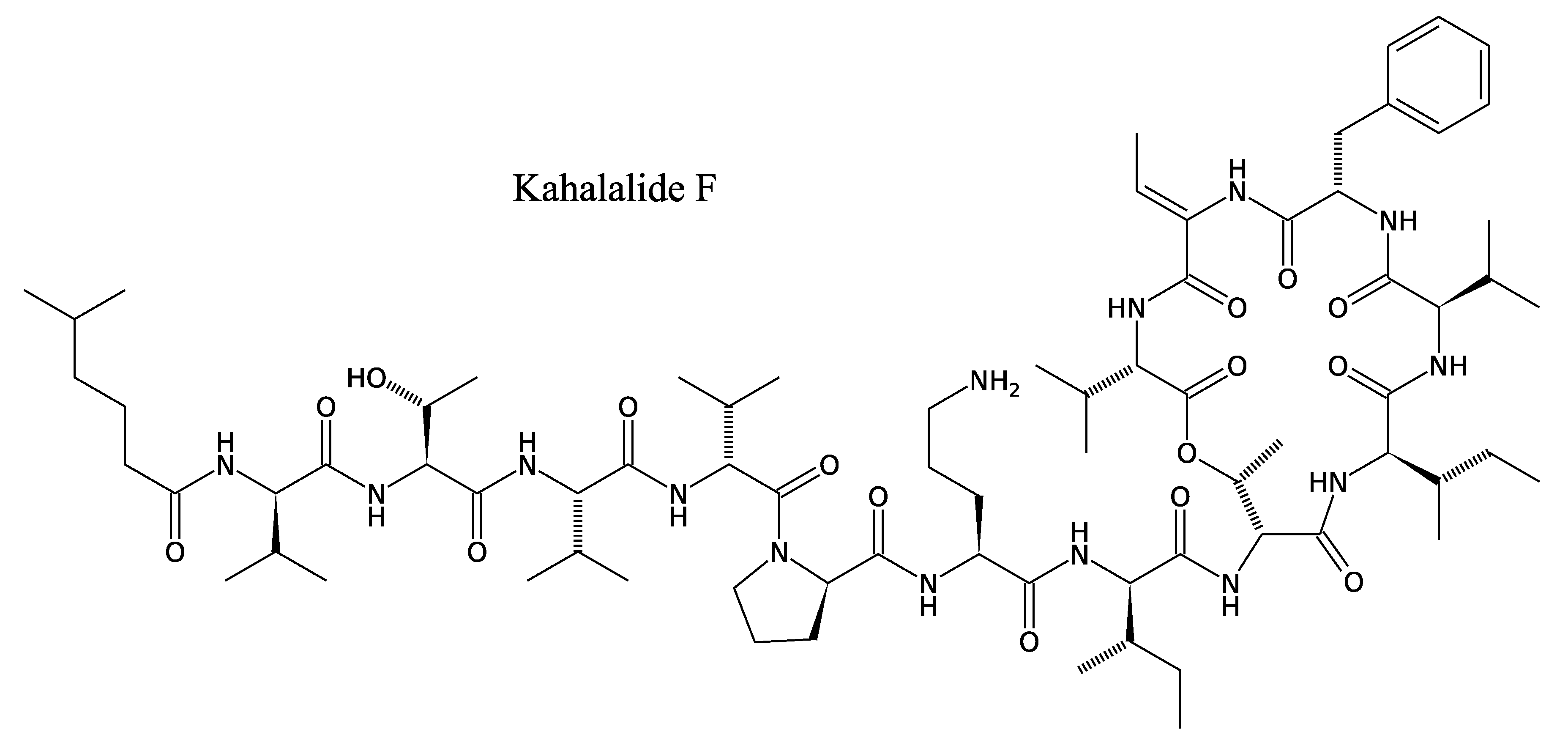
3.2.2. Kahalalides
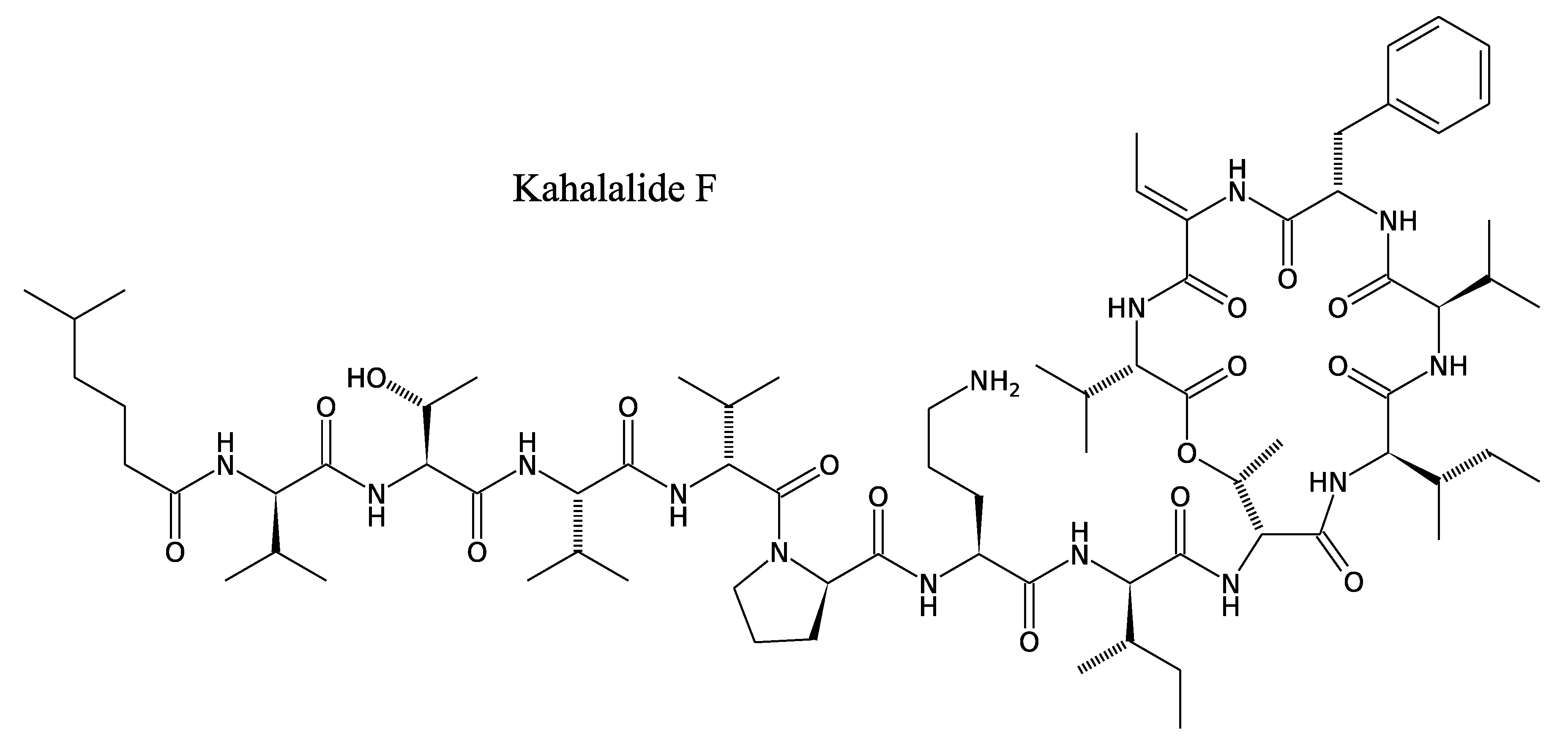
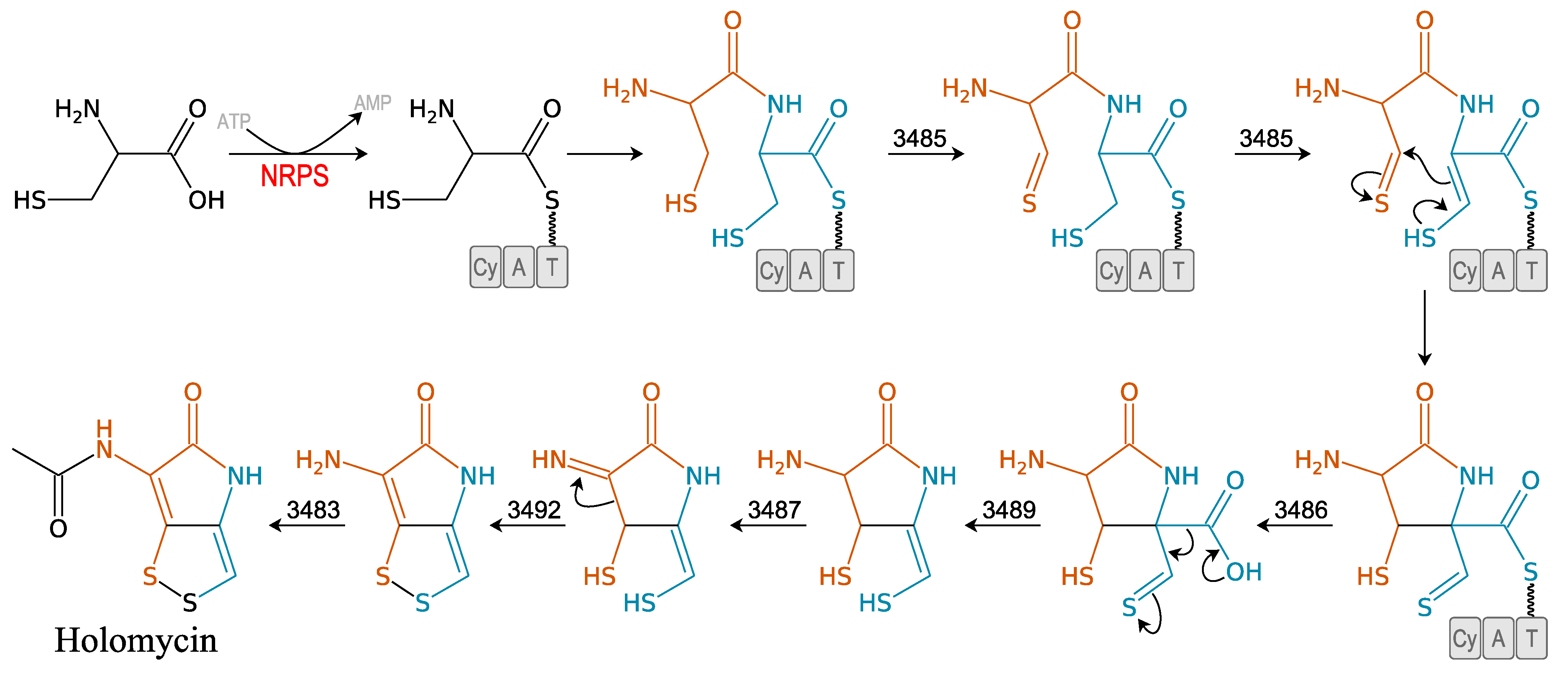
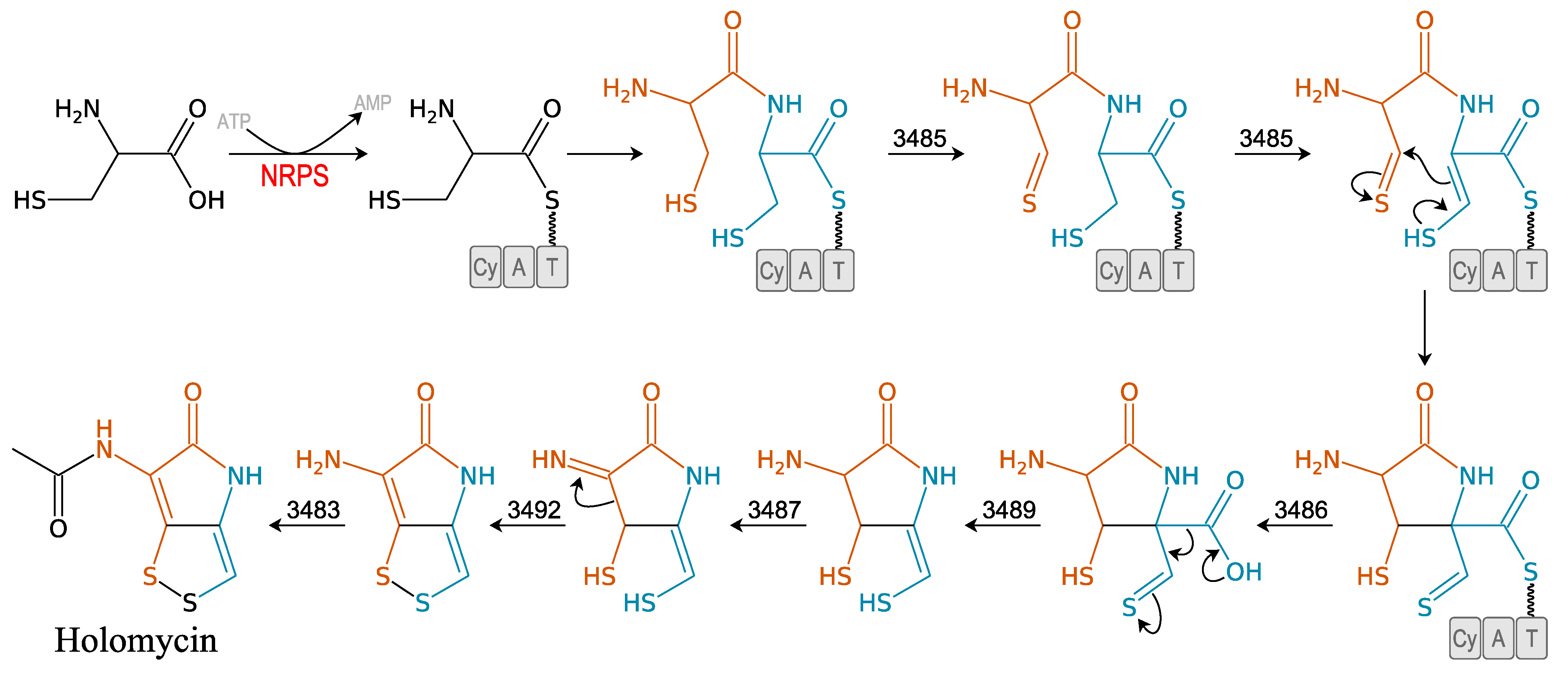
3.2.3. Holomycin
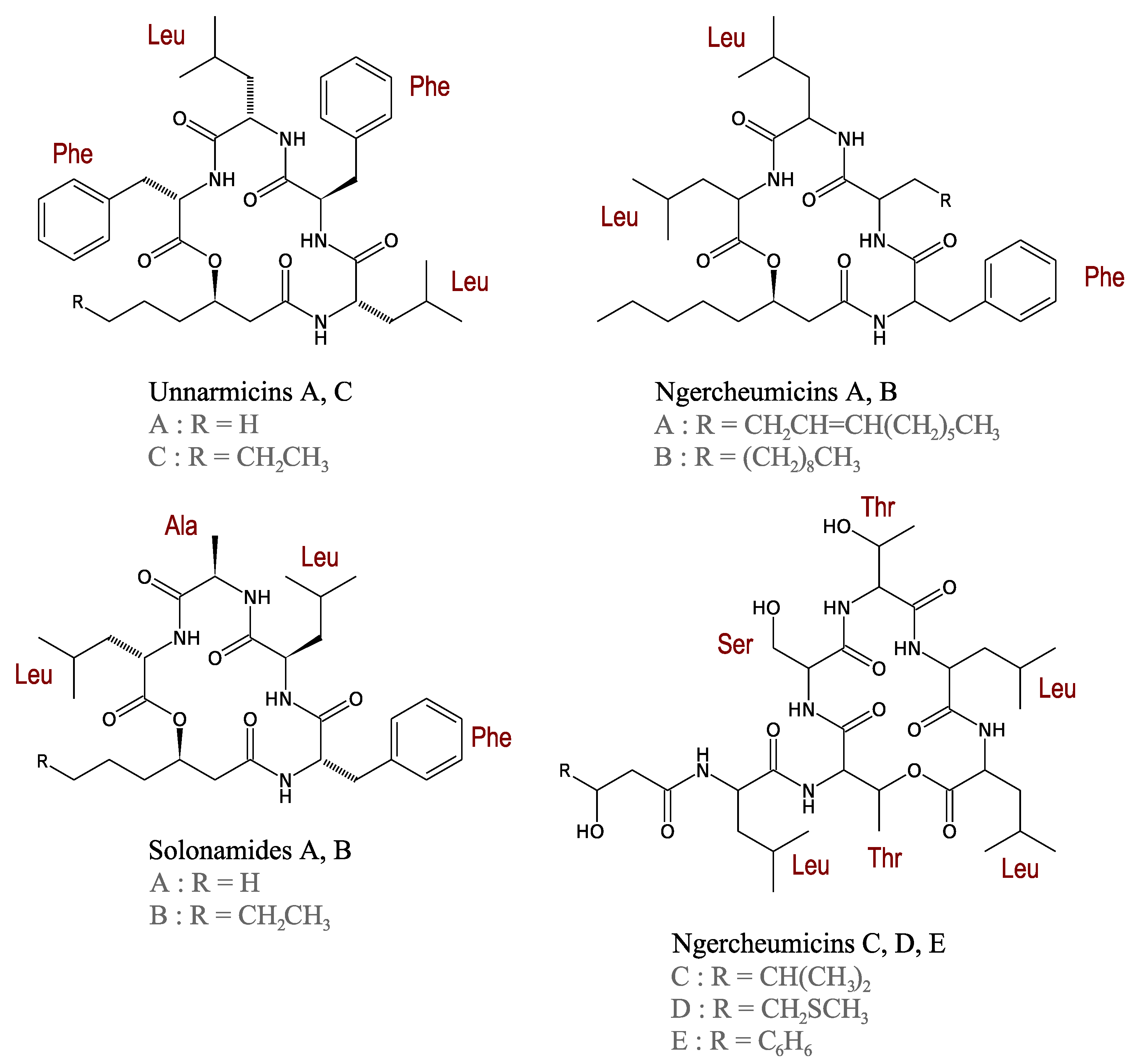
3.2.4. Unnarmicins
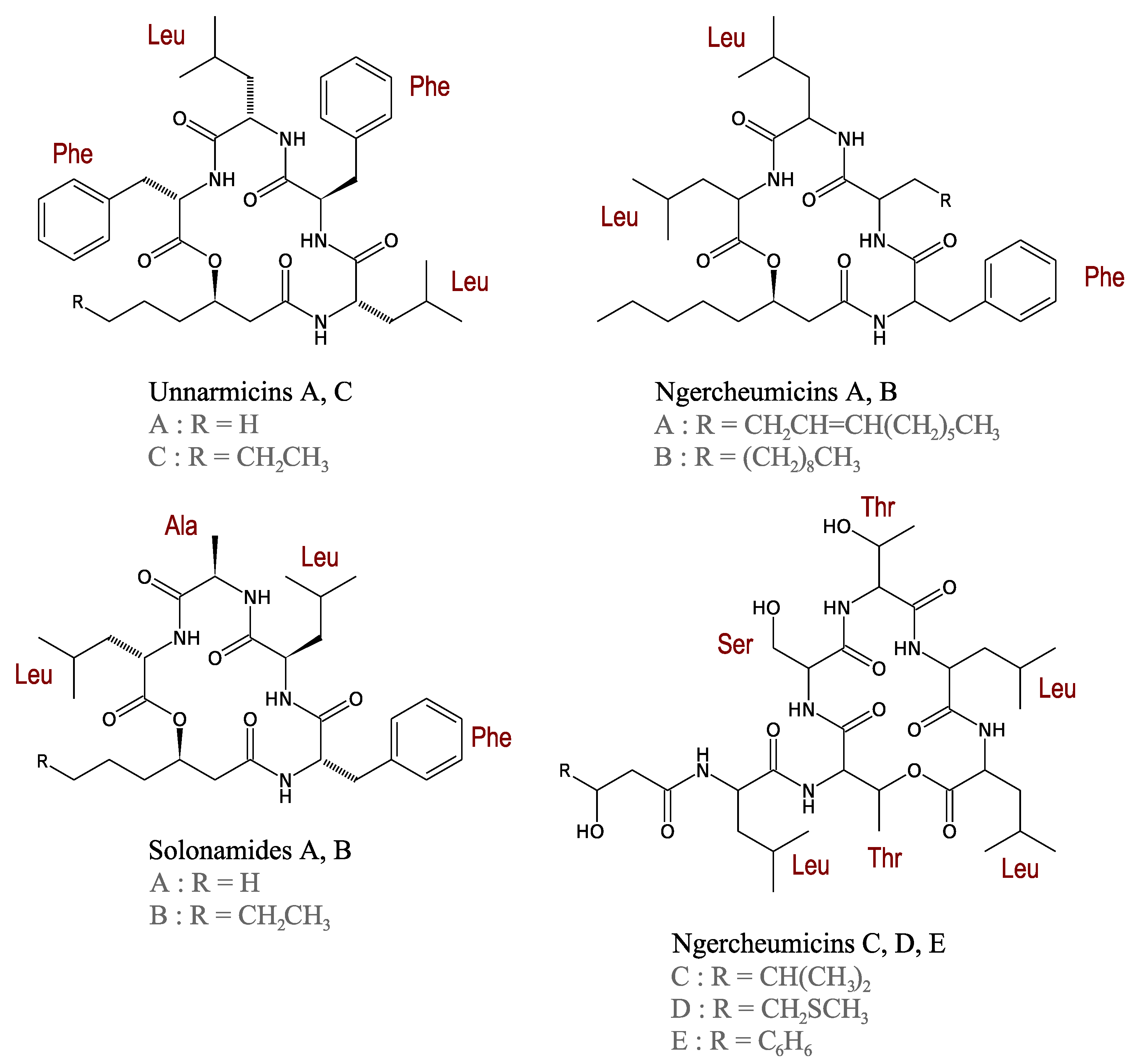
3.2.5. Ngercheumicins
3.2.6. Solonamides
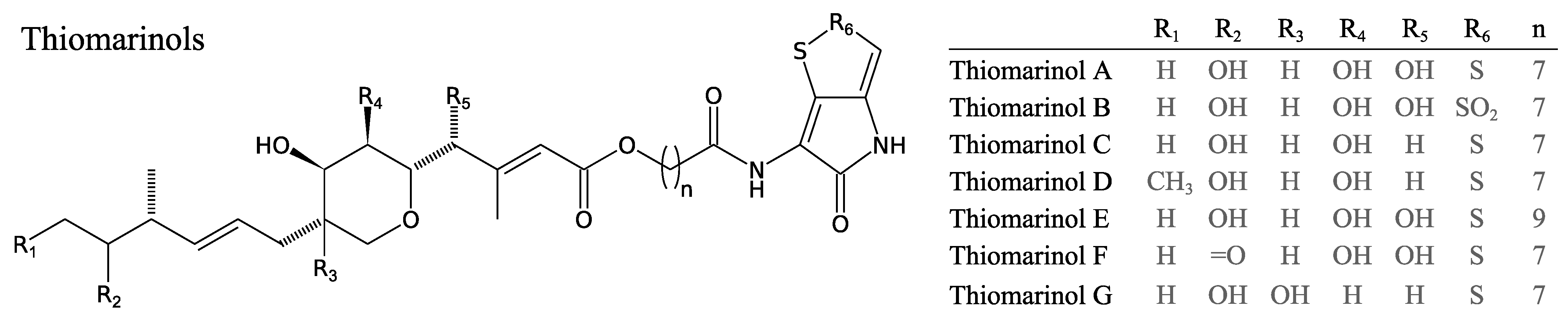
3.2.7. Thiomarinols
3.2.8. Cyclo-Peptides

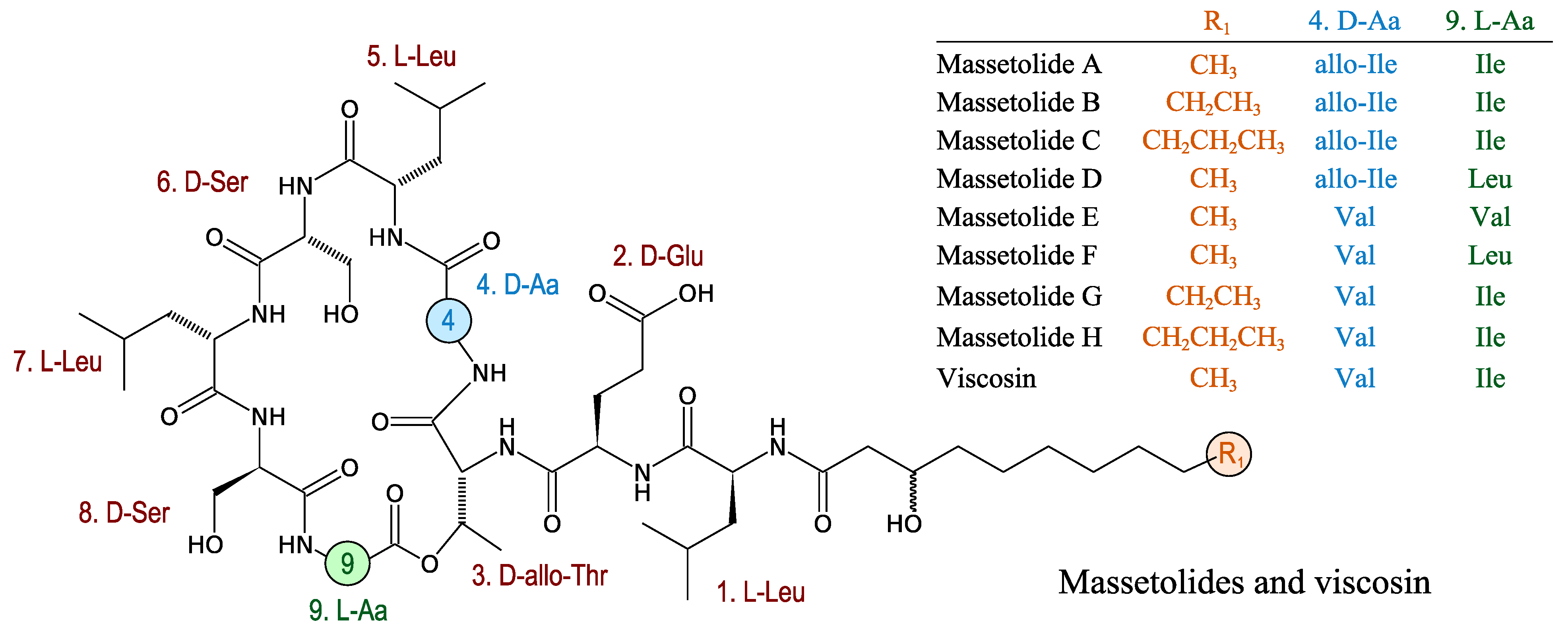
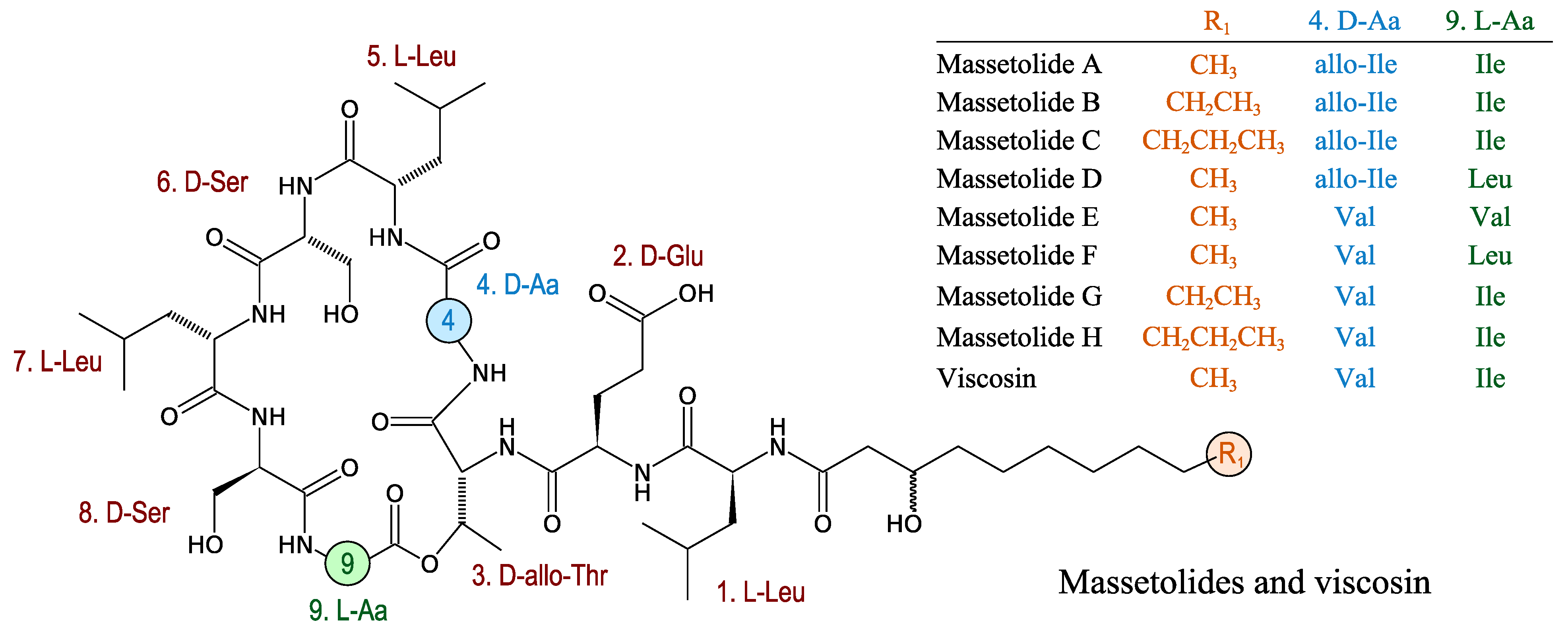
3.2.9. Massetolides
3.2.10. Predicted NRPs
3.3. δ-Proteobacteria
3.3.1. Myxovalargins
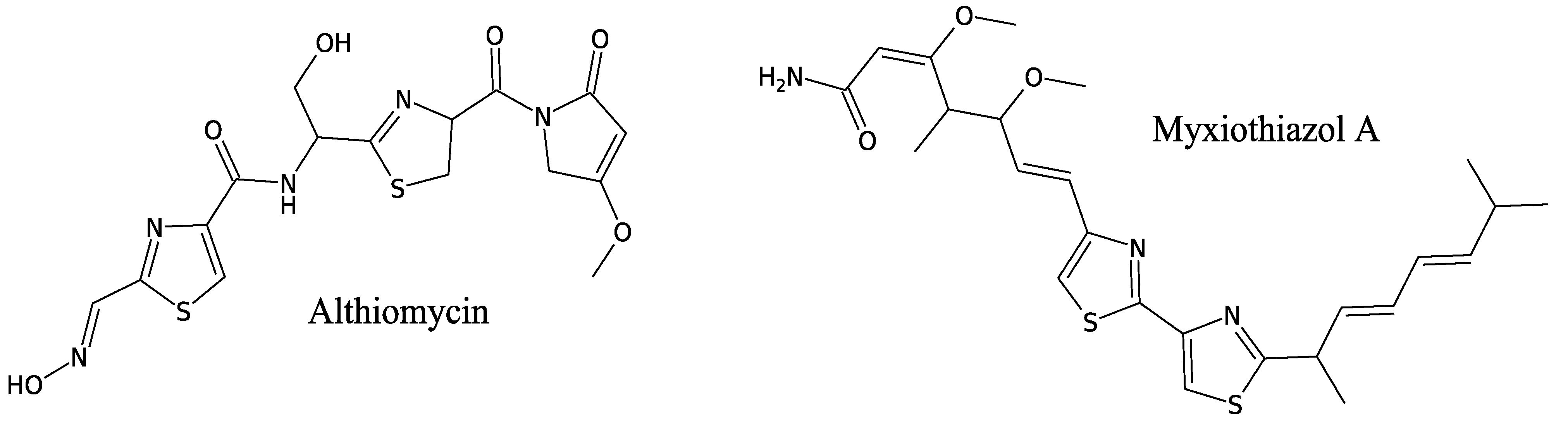
3.3.2. Althiomycin

3.3.3. Myxothiazols
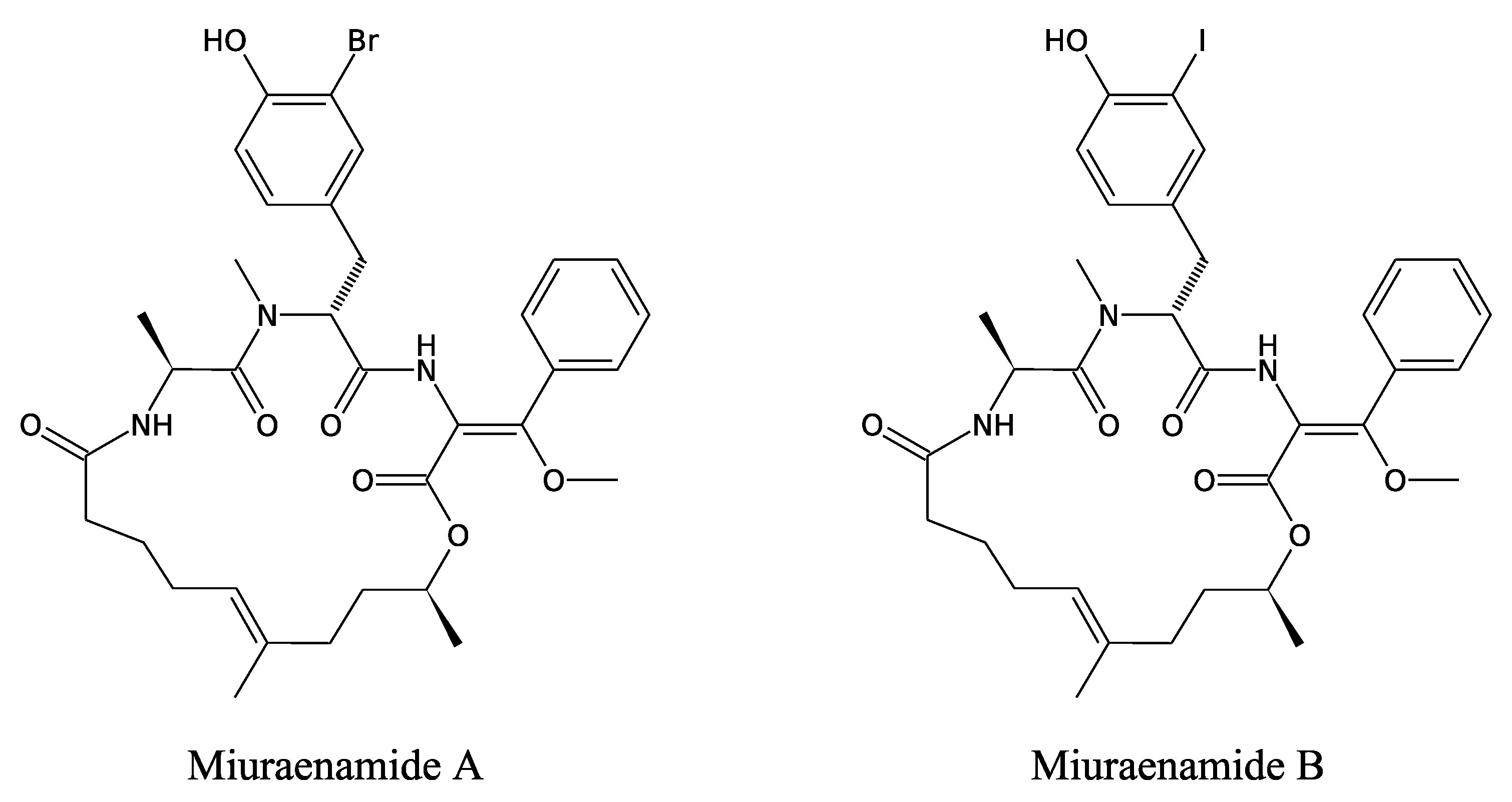
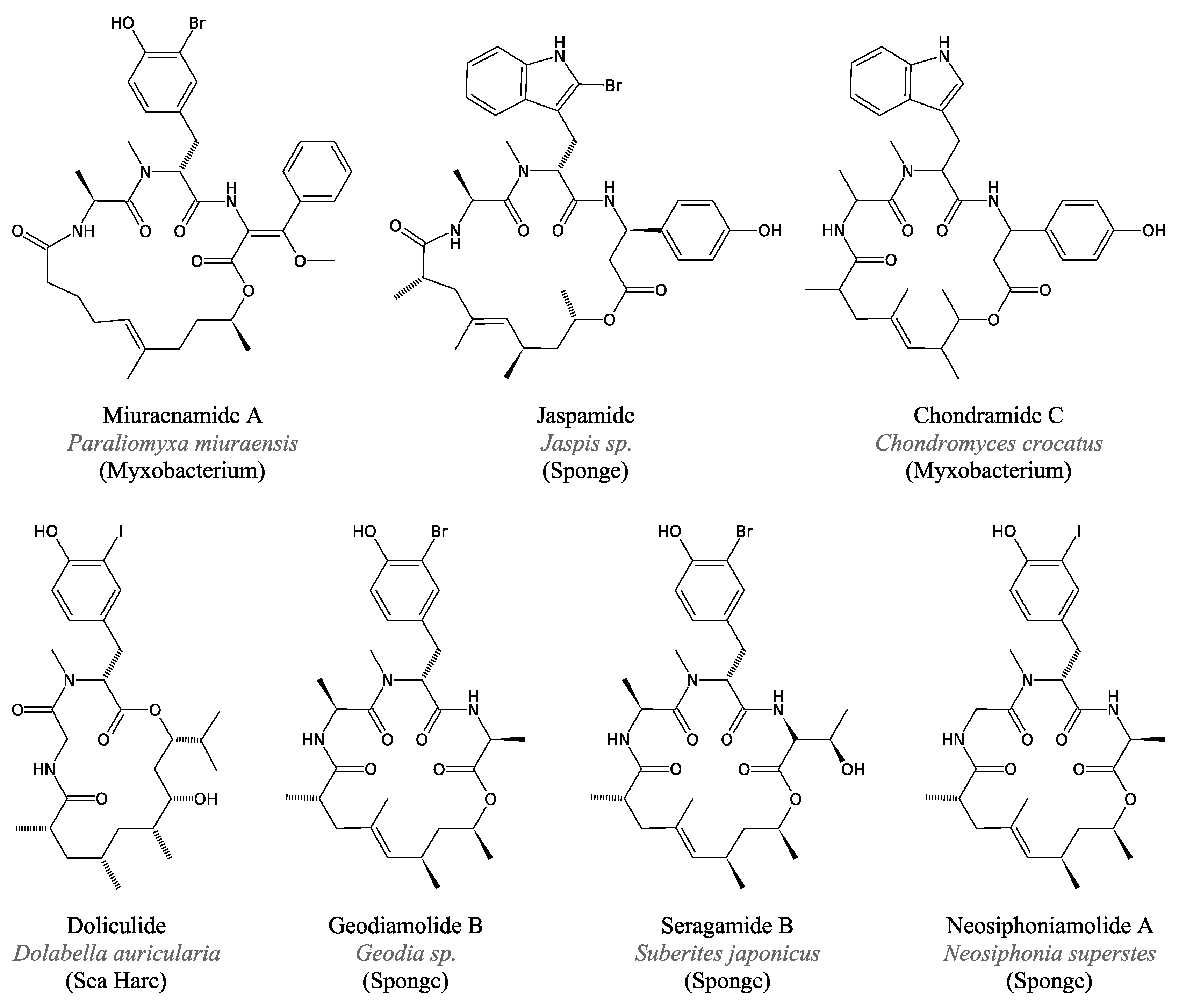
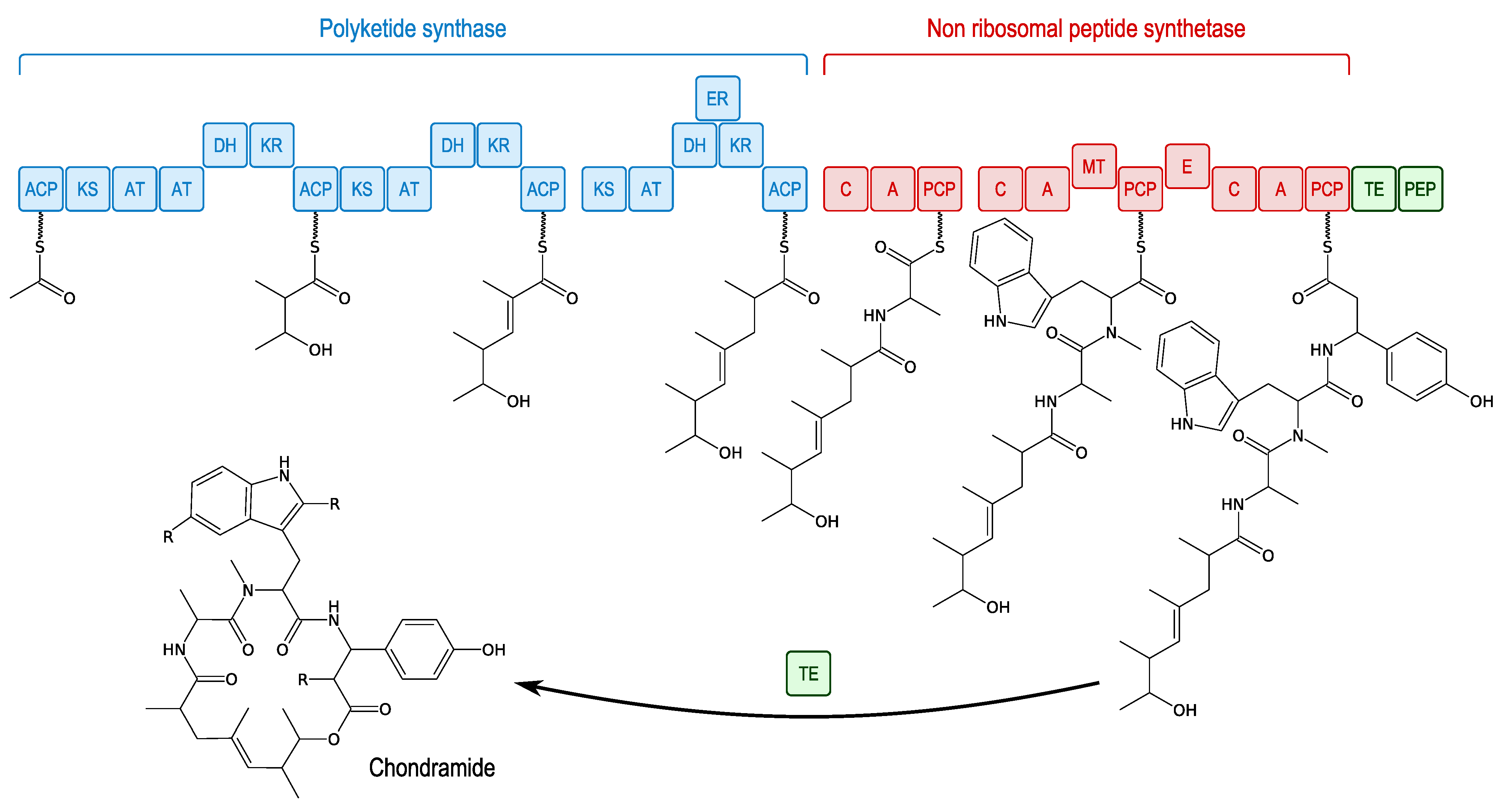
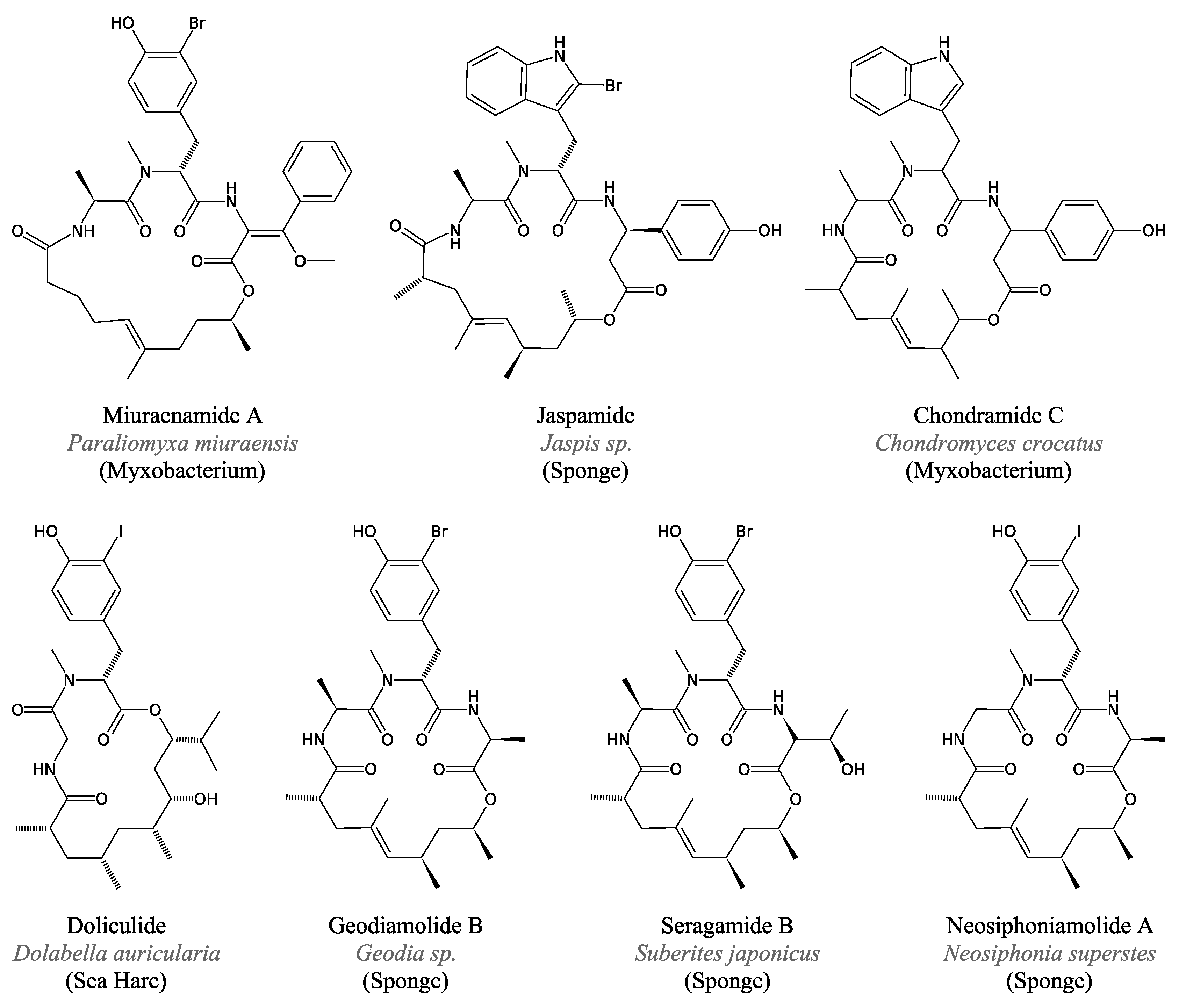
3.3.4. Miuraenamides

3.4. Molecules of Suspected Bacterial Origin
3.4.1. Theopalauamide
3.4.2. Jaspamide and Related Peptides
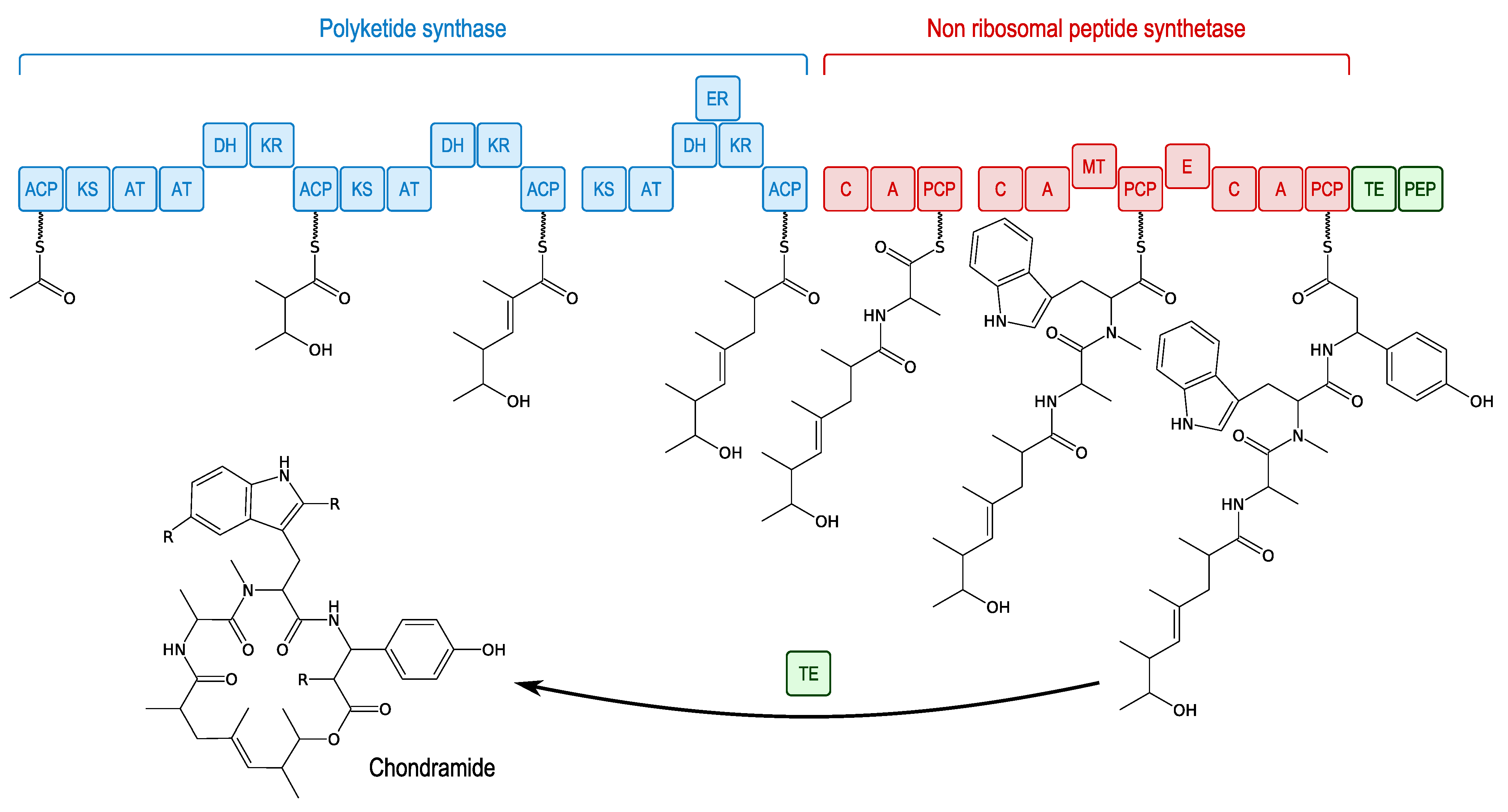
3.4.3. Microsclerodermins and Pedeins
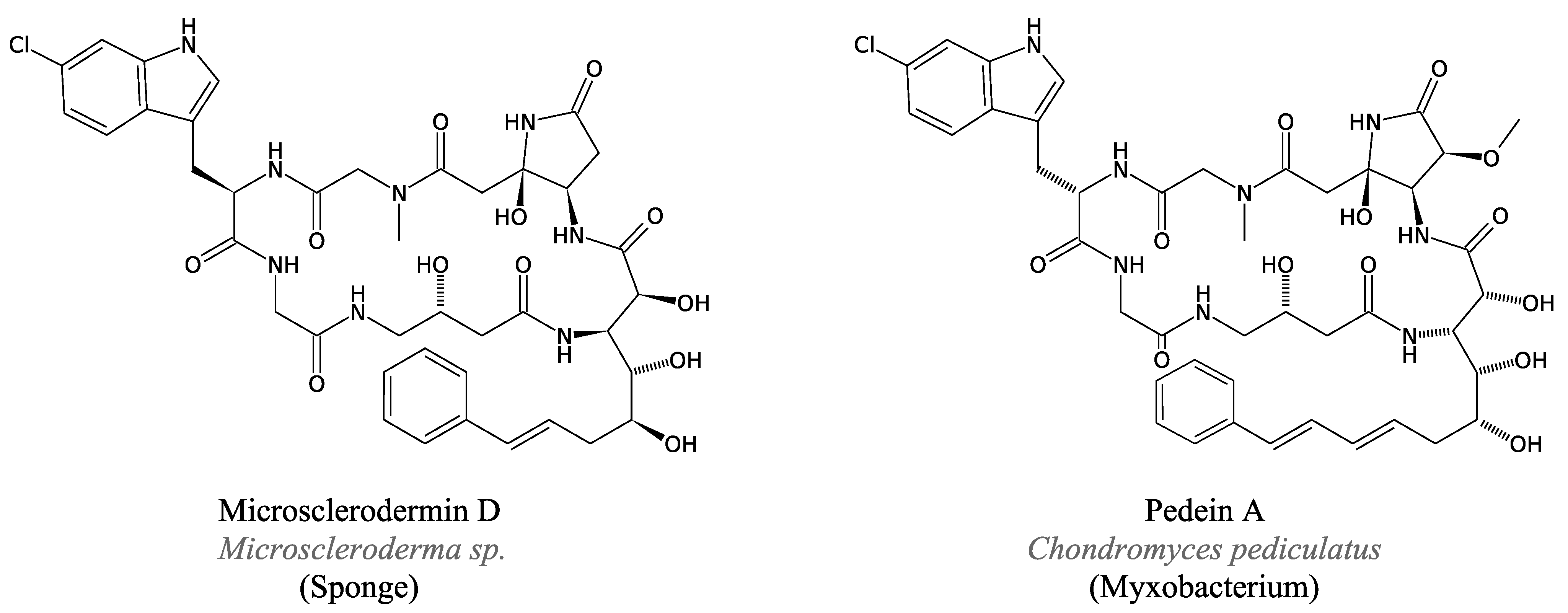
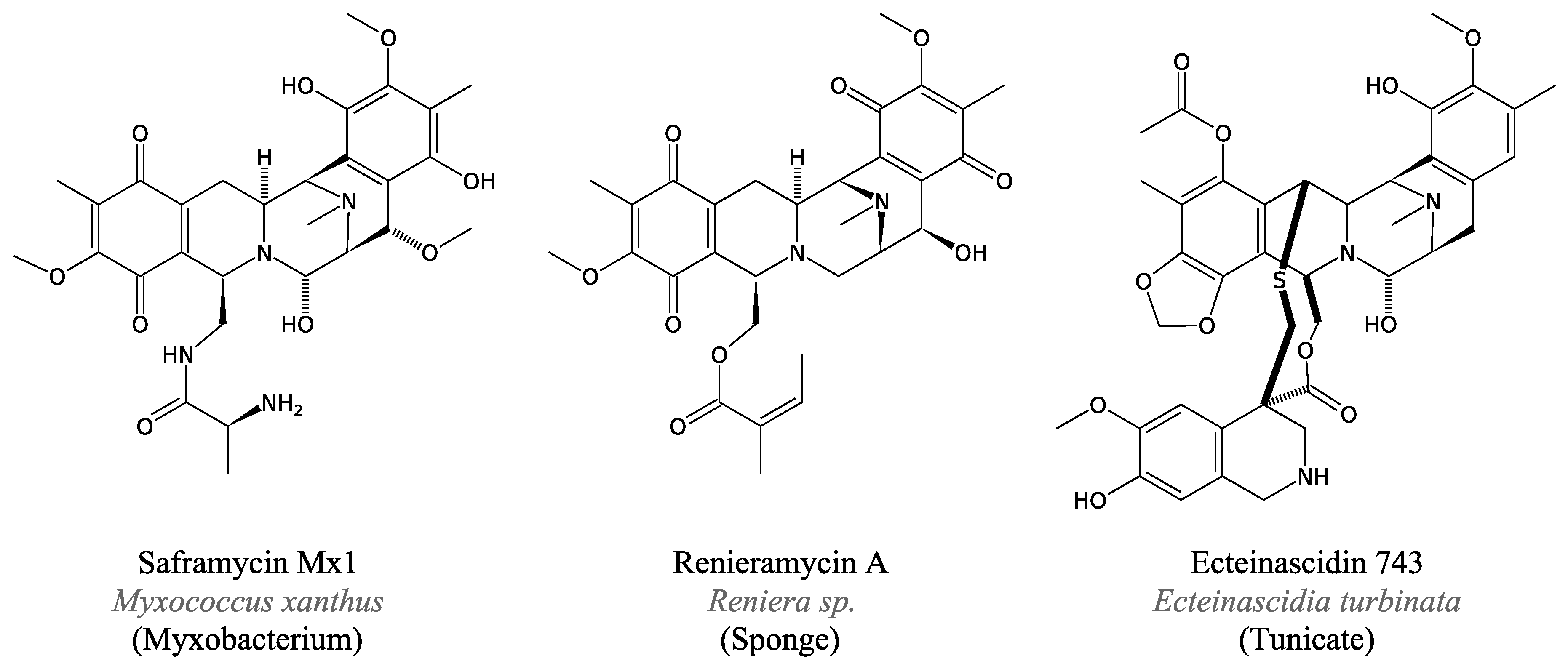
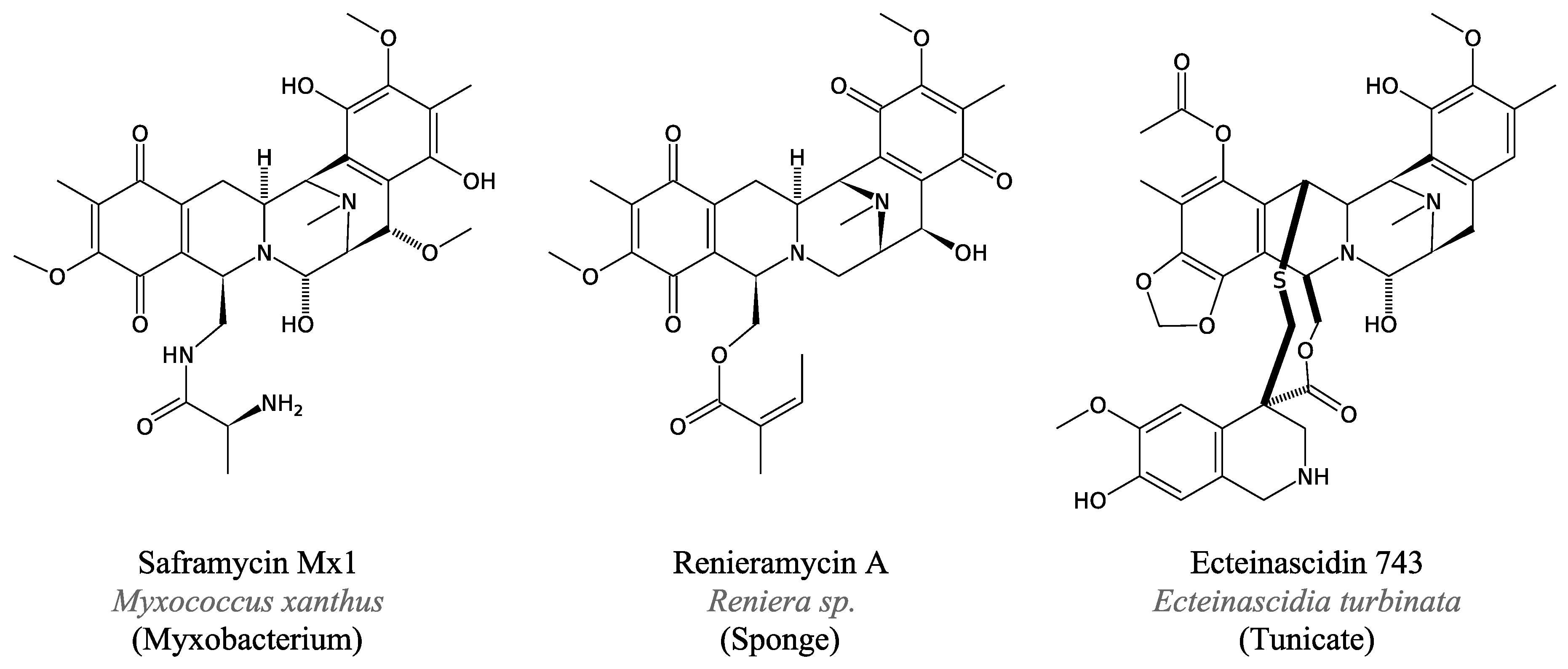
3.4.4. Saframycin and Related Peptides
4. Conclusions
Acknowledgements
Conflicts of Interest
References
- Singh, S.B.; Barrett, J.F. Empirical antibacterial drug discovery-foundation in natural products. Biochem. Pharmacol. 2006, 71, 1006–1015. [Google Scholar] [CrossRef]
- Joint, I.; Mühling, M.; Querellou, J. Culturing marine bacteria—An essential prerequisite for biodiscovery. Microb. Biotechnol. 2010, 3, 564–575. [Google Scholar] [CrossRef] [Green Version]
- Fortman, J.L.; Sherman, D.H. Utilizing the power of microbial genetics to bridge the gap between the promise and the application of marine natural products. Chembiochem 2005, 6, 960–978. [Google Scholar] [CrossRef]
- Giovannoni, S.; Stingl, U. The importance of culturing bacterioplankton in the “omics” age. Nat. Rev. Microbiol. 2007, 5, 820–826. [Google Scholar] [CrossRef]
- King, G.M.; Smith, C.B.; Tolar, B.; Hollibaugh, J.T. Analysis of composition and structure of coastal to mesopelagic bacterioplankton communities in the northern gulf of Mexico. Front. Microbiol. 2012, 3, 438–451. [Google Scholar]
- Jamieson, R.E.; Rogers, A.D.; Billett, D.S.M.; Smale, D.A.; Pearce, D.A. Patterns of marine bacterioplankton biodiversity in the surface waters of the Scotia Arc, Southern Ocean. FEMS Microbiol. Ecol. 2012, 80, 452–468. [Google Scholar] [CrossRef]
- Lau, S.C.K.; Zhang, R.; Brodie, E.L.; Piceno, Y.M.; Andersen, G.; Liu, W.-T. Biogeography of bacterioplankton in the tropical seawaters of Singapore. FEMS Microbiol. Ecol. 2013, 84, 259–269. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Fischer, A.; Roggentin, T.; Wehmeyer, U.; Bomar, D.; Smida, J. A phylogenetic survey of budding, and/or prosthecate, non-phototrophic eubacteria: Membership of Hyphomicrobium, Hyphomonas, Pedomicrobium, Filomicrobium, Caulobacter and “Dichotomicrobium” to the alpha-subdivision of purple non-sulfur bacteria. Arch. Microbiol. 1988, 149, 547–556. [Google Scholar] [CrossRef]
- Emerson, D.; Rentz, J.A.; Lilburn, T.G.; Davis, R.E.; Aldrich, H.; Chan, C.; Moyer, C.L. A novel lineage of proteobacteria involved in formation of marine Fe-oxidizing microbial mat communities. PLoS One 2007, 2, e667. [Google Scholar] [CrossRef]
- Bhatnagar, I.; Se-Kwon, K. Immense essence of excellence: Marine microbial bioactive compounds. Mar. Drugs 2010, 8, 2673–2701. [Google Scholar] [CrossRef]
- Gilbert, J.A.; Field, D.; Swift, P.; Newbold, L.; Oliver, A.; Smyth, T.; Somerfield, P.J.; Huse, S.; Joint, I. The seasonal structure of microbial communities in the Western English Channel. Environ. Microbiol. 2009, 11, 3132–3139. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2004, 21, 1–49. [Google Scholar] [CrossRef]
- Koglin, A.; Walsh, C.T. Structural insights into nonribosomal peptide enzymatic assembly lines. Nat. Prod. Rep. 2009, 26, 987–1000. [Google Scholar] [CrossRef]
- Lipmann, F.; Gevers, W.; Kleinkauf, H.; Roskoski, R., Jr. Polypeptide synthesis on protein templates: The enzymatic synthesis of gramicidin S and tyrocidine. Adv. Enzymol. Relat. Areas Mol. Biol. 1971, 35, 1–34. [Google Scholar]
- Lipmann, F. Nonribosomal polypeptide synthesis on polyenzyme templates. Acc. Chem. Res. 1973, 6, 361–367. [Google Scholar] [CrossRef]
- Kawakami, T.; Murakami, H. Genetically encoded libraries of nonstandard peptides. J. Nucleic Acids 2012, 2012. [Google Scholar] [CrossRef]
- Schwarzer, D.; Finking, R.; Marahiel, M.A. Nonribosomal peptides: From genes to products. Nat. Prod. Rep. 2003, 20, 275–287. [Google Scholar] [CrossRef]
- Stachelhaus, T.; Mootz, H.D.; Bergendahl, V.; Marahiel, M.A. Peptide bond formation in nonribosomal peptide biosynthesis catalytic role of the condensation domain. J. Biol. Chem. 1998, 273, 22773–22781. [Google Scholar] [CrossRef]
- Hur, G.H.; Vickery, C.R.; Burkart, M.D. Explorations of catalytic domains in non-ribosomal peptide synthetase enzymology. Nat. Prod. Rep. 2012, 29, 1074–1098. [Google Scholar] [CrossRef]
- Weber, T.; Marahiel, M.A. Exploring the domain structure of modular nonribosomal peptide synthetases. Structure 2001, 9, 3–9. [Google Scholar] [CrossRef]
- Schwarzer, D.; Marahiel, M.A. Multimodular biocatalysts for natural product assembly. Naturwissenschaften 2001, 88, 93–101. [Google Scholar] [CrossRef]
- Lambalot, R.H.; Gehring, A.M.; Flugel, R.S.; Zuber, P.; LaCelle, M.; Marahiel, M.A.; Reid, R.; Khosla, C.; Walsh, C.T. A new enzyme superfamily—The phosphopantetheinyl transferases. Chem. Biol. 1996, 3, 923–936. [Google Scholar] [CrossRef]
- Nikolouli, K.; Mossialos, D. Bioactive compounds synthesized by non-ribosomal peptide synthetases and type-I polyketide synthases discovered through genome-mining and metagenomics. Biotechnol. Lett. 2012, 34, 1393–1403. [Google Scholar] [CrossRef]
- Caboche, S.; Pupin, M.; Leclère, V.; Fontaine, A.; Jacques, P.; Kucherov, G. NORINE: A database of nonribosomal peptides. Nucleic Acids Res. 2008, 36, 326–331. [Google Scholar]
- Mootz, H.D.; Schwarzer, D.; Marahiel, M.A. Ways of assembling complex natural products on modular nonribosomal peptide synthetases. Chembiochem 2002, 3, 490–504. [Google Scholar] [CrossRef]
- Blasiak, L.C.; Clardy, J. Discovery of 3-formyl-tyrosine metabolites from Pseudoalteromonas tunicata through heterologous expression. J. Am. Chem. Soc. 2010, 132, 926–927. [Google Scholar] [CrossRef]
- Bachmann, B.O.; Ravel, J. Methods for in silico prediction of microbial polyketide and nonribosomal peptide biosynthetic pathways from DNA sequence data. Meth. Enzymol. 2009, 458, 181–217. [Google Scholar] [CrossRef]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. AntiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, 339–346. [Google Scholar] [CrossRef]
- Ziemert, N.; Podell, S.; Penn, K.; Badger, J.H.; Allen, E.; Jensen, P.R. The natural product domain seeker NaPDoS: A phylogeny based bioinformatic tool to classify secondary metabolite gene diversity. PLoS One 2012, 7, e34064. [Google Scholar]
- De Bruijn, I.; de Kock, M.J.D.; Yang, M.; de Waard, P.; van Beek, T.A.; Raaijmakers, J.M. Genome-based discovery, structure prediction and functional analysis of cyclic lipopeptide antibiotics in Pseudomonas species. Mol. Microbiol. 2007, 63, 417–428. [Google Scholar] [CrossRef]
- Li, B.; Walsh, C.T. Identification of the gene cluster for the dithiolopyrrolone antibiotic holomycin in Streptomyces clavuligerus. Proc. Natl. Acad. Sci. USA 2010, 107, 19731–19735. [Google Scholar] [CrossRef]
- Park, H.-M.; Kim, B.-G.; Chang, D.; Malla, S.; Joo, H.-S.; Kim, E.-J.; Park, S.-J.; Sohng, J.K.; Kim, P.I. Genome-based cryptic gene discovery and functional identification of NRPS siderophore peptide in Streptomyces peucetius. Appl. Microbiol. Biotechnol. 2013, 97, 1213–1222. [Google Scholar] [CrossRef]
- Bumpus, S.B.; Evans, B.S.; Thomas, P.M.; Ntai, I.; Kelleher, N.L. A proteomics approach to discovering natural products and their biosynthetic pathways. Nat. Biotechnol. 2009, 27, 951–956. [Google Scholar] [CrossRef]
- Stein, T.; Vater, J.; Kruft, V.; Wittmann-Liebold, B.; Franke, P.; Panico, M.; Mc Dowell, R.; Morris, H.R. Detection of 4′-phosphopantetheine at the thioester binding site for l-valine of gramicidin S synthetase 2. FEBS Lett. 1994, 340, 39–44. [Google Scholar] [CrossRef]
- Wagner-Döbler, I.; Rheims, H.; Felske, A.; El-Ghezal, A.; Flade-Schröder, D.; Laatsch, H.; Lang, S.; Pukall, R.; Tindall, B.J. Oceanibulbus indolifex gen. nov., sp. nov., a North Sea alphaproteobacterium that produces bioactive metabolites. Int. J. Syst. Evol. Microbiol. 2004, 54, 1177–1184. [Google Scholar] [CrossRef]
- Milne, P.J.; Hunt, A.L.; Rostoll, K.; van der Walt, J.J.; Graz, C.J. The biological activity of selected cyclic dipeptides. J. Pharm. Pharmacol. 1998, 50, 1331–1337. [Google Scholar] [CrossRef]
- Slightom, R.N.; Buchan, A. Surface colonization by marine roseobacters: Integrating genotype and phenotype. Appl. Environ. Microbiol. 2009, 75, 6027–6037. [Google Scholar] [CrossRef]
- Cude, W.N.; Mooney, J.; Tavanaei, A.A.; Hadden, M.K.; Frank, A.M.; Gulvik, C.A.; May, A.L.; Buchan, A. Production of the antimicrobial secondary metabolite indigoidine contributes to competitive surface colonization by the marine roseobacter Phaeobacter sp. strain Y4I. Appl. Environ. Microbiol. 2012, 78, 4771–4780. [Google Scholar] [CrossRef]
- Oclarit, J.M.; Okada, H.; Ohta, S.; Kaminura, K.; Yamaoka, Y.; Iizuka, T.; Miyashiro, S.; Ikegami, S. Anti-bacillus substance in the marine sponge, Hyatella species, produced by an associated Vibrio species bacterium. Microbios 1994, 78, 7–16. [Google Scholar]
- Graff, J.R.; Forschner-Dancause, S.R.; Menden-Deuer, S.; Long, R.A.; Rowley, D.C. Vibrio cholerae exploits sub-lethal concentrations of a competitor-produced antibiotic to avoid toxic interactions. Front. Microbiol. 2013, 4, 8–18. [Google Scholar]
- Wietz, M.; Mansson, M.; Gotfredsen, C.H.; Larsen, T.O.; Gram, L. Antibacterial compounds from marine Vibrionaceae isolated on a global expedition. Mar. Drugs 2010, 8, 2946–2960. [Google Scholar] [CrossRef]
- Singh, M.P.; Mroczenski-Wildey, M.J.; Steinberg, D.A.; Andersen, R.J.; Maiese, W.M.; Greenstein, M. Biological activity and mechanistic studies of andrimid. J. Antibiot. 1997, 50, 270–273. [Google Scholar] [CrossRef]
- Jin, M.; Fischbach, M.A.; Clardy, J. A biosynthetic gene cluster for the acetyl-CoA carboxylase inhibitor andrimid. J. Am. Chem. Soc. 2006, 128, 10660–10661. [Google Scholar] [CrossRef]
- Fredenhagen, A.; Tamura, S.Y.; Kenny, P.T.M.; Komura, H.; Naya, Y.; Nakanishi, K.; Nishiyama, K.; Sugiura, M.; Kita, H. Andrimid, a new peptide antibiotic produced by an intracellular bacterial symbiont isolated from a brown planthopper. J. Am. Chem. Soc. 1987, 109, 4409–4411. [Google Scholar] [CrossRef]
- Freiberg, C.; Brunner, N.A.; Schiffer, G.; Lampe, T.; Pohlmann, J.; Brands, M.; Raabe, M.; Häbich, D.; Ziegelbauer, K. Identification and characterization of the first class of potent bacterial acetyl-CoA carboxylase inhibitors with antibacterial activity. J. Biol. Chem. 2004, 279, 26066–26073. [Google Scholar] [CrossRef]
- Pohlmann, J.; Lampe, T.; Shimada, M.; Nell, P.G.; Pernerstorfer, J.; Svenstrup, N.; Brunner, N.A.; Schiffer, G.; Freiberg, C. Pyrrolidinedione derivatives as antibacterial agents with a novel mode of action. Bioorg. Med. Chem. Lett. 2005, 15, 1189–1192. [Google Scholar] [CrossRef]
- Freiberg, C.; Pohlmann, J.; Nell, P.G.; Endermann, R.; Schuhmacher, J.; Newton, B.; Otteneder, M.; Lampe, T.; Häbich, D.; Ziegelbauer, K. Novel bacterial acetyl coenzyme A carboxylase inhibitors with antibiotic efficacy in vivo. Antimicrob. Agents Chemother. 2006, 50, 2707–2712. [Google Scholar] [CrossRef]
- Evans, B.S.; Chen, Y.; Metcalf, W.W.; Zhao, H.; Kelleher, N.L. Directed evolution of the nonribosomal peptide synthetase AdmK generates new andrimid derivatives in vivo. Chem. Biol. 2011, 18, 601–607. [Google Scholar] [CrossRef]
- Magarvey, N.A.; Fortin, P.D.; Thomas, P.M.; Kelleher, N.L.; Walsh, C.T. Gatekeeping versus promiscuity in the early stages of the andrimid biosynthetic assembly line. ACS Chem. Biol. 2008, 3, 542–554. [Google Scholar] [CrossRef]
- Gao, J.; Hamann, M.T. Chemistry and biology of kahalalides. Chem. Rev. 2011, 111, 3208–3235. [Google Scholar] [CrossRef]
- Hill, R.T.; Enticknap, J.; Rao, K.V.; Hamann, M.T. Kahalalide-Producing Bacteria 2004. Eur. Pat. Appl. EP1689848, 2004. [Google Scholar]
- Shilabin, A.G.; Hamann, M.T. In vitro and in vivo evaluation of select kahalalide F analogs with antitumor and antifungal activities. Bioorg. Med. Chem. 2011, 19, 6628–6632. [Google Scholar] [CrossRef]
- Kenig, M.; Reading, C. Holomycin and an antibiotic (MM 19290) related to tunicamycin, metabolites of Streptomyces clavuligerus. J. Antibiot. 1979, 32, 549–554. [Google Scholar] [CrossRef]
- Oliva, B.; O’Neill, A.; Wilson, J.M.; O’Hanlon, P.J.; Chopra, I. Antimicrobial properties and mode of action of the pyrrothine holomycin. Antimicrob. Agents Chemother. 2001, 45, 532–539. [Google Scholar] [CrossRef]
- Khachatourians, G.G.; Tipper, D.J. In vivo effect of thiolutin on cell growth and macromolecular synthesis in Escherichia coli. Antimicrob. Agents Chemother. 1974, 6, 304–310. [Google Scholar] [CrossRef]
- Urbanczyk, H.; Ast, J.C.; Dunlap, P.V. Phylogeny, genomics, and symbiosis of Photobacterium. FEMS Microbiol. Rev. 2011, 35, 324–342. [Google Scholar] [CrossRef]
- Hazen, T.H.; Pan, L.; Gu, J.-D.; Sobecky, P.A. The contribution of mobile genetic elements to the evolution and ecology of Vibrios. FEMS Microbiol. Ecol. 2010, 74, 485–499. [Google Scholar] [CrossRef]
- Oku, N.; Kawabata, K.; Adachi, K.; Katsuta, A.; Shizuri, Y. Unnarmicins A and C, new antibacterial depsipeptides produced by marine bacterium Photobacterium sp. MBIC06485. J. Antibiot. 2008, 61, 11–17. [Google Scholar] [CrossRef]
- Yoshikawa, K.; Adachi, K.; Nishida, F.; Mochida, K. Planar structure and antibacterial activity of korormicin derivatives isolated from Pseudoalteromonas sp. F-420. J. Antibiot. 2003, 56, 866–870. [Google Scholar] [CrossRef]
- Adachi, K.; Kawabata, Y.; Kasai, H.; Katsuta, M.; Shizuri, Y. Novel Ngercheumicin or Its Salt Useful for Treating Infection Caused by Pseudovibrio denitrificans. Japanese Patent JP2007230911-A, 2007. [Google Scholar]
- Mansson, M.; Gram, L.; Larsen, T.O. Production of bioactive secondary metabolites by marine Vibrionaceae. Mar. Drugs 2011, 9, 1440–1468. [Google Scholar] [CrossRef]
- Finch, R.G.; Pritchard, D.I.; Bycroft, B.W.; Williams, P.; Stewart, G.S. Quorum sensing: A novel target for anti-infective therapy. J. Antimicrob. Chemother. 1998, 42, 569–571. [Google Scholar] [CrossRef]
- Wise, M.P.; Williams, D.W.; Lewis, M.A.O.; Frost, P.J. Macrolides and community-acquired pneumonia: Is quorum sensing the key? Crit. Care 2010, 14, 181–183. [Google Scholar] [CrossRef]
- Defoirdt, T.; Boon, N.; Bossier, P. Can bacteria evolve resistance to quorum sensing disruption? PLoS Pathog. 2010, 6, e1000989. [Google Scholar] [CrossRef] [Green Version]
- Shiozawa, H.; Kagasaki, T.; Kinoshita, T.; Haruyama, H.; Domon, H.; Utsui, Y.; Kodama, K.; Takahashi, S. Thiomarinol, a new hybrid antimicrobial antibiotic produced by a marine bacterium: fermentation, isolation, structure, and antimicrobial activity. J. Antibiot. 1993, 46, 1834–1842. [Google Scholar] [CrossRef]
- Shiozawa, H.; Kagasaki, T.; Torikata, A.; Tanaka, N.; Fujimoto, K.; Hata, T.; Furukawa, Y.; Takahashi, S. Thiomarinols B and C, new antimicrobial antibiotics produced by a marine bacterium. J. Antibiot. 1995, 48, 907–909. [Google Scholar] [CrossRef]
- Shiozawa, H.; Shimada, A.; Takahashi, S. Thiomarinols D, E, F and G, new hybrid antimicrobial antibiotics produced by a marine bacterium; isolation, structure, and antimicrobial activity. J. Antibiot. 1997, 50, 449–452. [Google Scholar] [CrossRef]
- Fukuda, D.; Haines, A.S.; Song, Z.; Murphy, A.C.; Hothersall, J.; Stephens, E.R.; Gurney, R.; Cox, R.J.; Crosby, J.; Willis, C.L.; et al. A natural plasmid uniquely encodes two biosynthetic pathways creating a potent anti-MRSA antibiotic. PLoS One 2011, 6, e18031. [Google Scholar] [CrossRef]
- Murphy, A.C.; Fukuda, D.; Song, Z.; Hothersall, J.; Cox, R.J.; Willis, C.L.; Thomas, C.M.; Simpson, T.J. Engineered thiomarinol antibiotics active against MRSA are generated by mutagenesis and mutasynthesis of Pseudoalteromonas SANK73390. Angew. Chem. Int. Ed. Engl. 2011, 50, 3271–3274. [Google Scholar] [CrossRef]
- Rungprom, W.; Siwu, E.R.O.; Lambert, L.K.; Dechsakulwatana, C.; Barden, M.C.; Kokpol, U.; Blanchfield, J.T.; Kita, M.; Garson, M.J. Cyclic tetrapeptides from marine bacteria associated with the seaweed Diginea sp. and the sponge Halisarca ectofibrosa. Tetrahedron 2008, 64, 3147–3152. [Google Scholar] [CrossRef]
- Dahiya, R.; Gautam, H. Toward the synthesis and biological screening of a cyclotetrapeptide from marine bacteria. Mar. Drugs 2011, 9, 71–81. [Google Scholar] [CrossRef]
- Binz, T.M.; Maffioli, S.I.; Sosio, M.; Donadio, S.; Muller, R. Insights into an unusual nonribosomal peptide synthetase biosynthesis. J. Biol. Chem. 2010, 285, 32710–32719. [Google Scholar] [CrossRef]
- Gerard, J.M. Antibiotic Secondary Metabolites of Bacteria Isolated from the Marine Environment. Ph.D. Thesis, University of British Columbia, Vancouver, Canada, 1992. [Google Scholar]
- De Bruijn, I.; de Kock, M.J.D.; de Waard, P.; van Beek, T.A.; Raaijmakers, J.M. Massetolide A biosynthesis in Pseudomonas fluorescens. J. Bacteriol. 2008, 190, 2777–2789. [Google Scholar] [CrossRef]
- Zheng, L.; Han, X.; Chen, H.; Lin, W.; Yan, X. Marine bacteria associated with marine macroorganisms: The potential antimicrobial resources. Ann. microbiol. 2005, 55, 119–124. [Google Scholar]
- Zhu, P.; Zheng, Y.; You, Y.; Yan, X.; Shao, J. Sequencing and modular analysis of the hybrid non-ribosomal peptide synthase-polyketide synthase gene cluster from the marine sponge Hymeniacidon perleve-associated bacterium Pseudoalteromonas sp. strain NJ631. Can. J. Microbiol. 2009, 55, 219–227. [Google Scholar] [CrossRef]
- Caboche, S.; Leclère, V.; Pupin, M.; Kucherov, G.; Jacques, P. Diversity of monomers in nonribosomal peptides: Towards the prediction of origin and biological activity. J. Bacteriol. 2010, 192, 5143–5150. [Google Scholar] [CrossRef] [Green Version]
- Wade, D.; Boman, A.; Wåhlin, B.; Drain, C.M.; Andreu, D.; Boman, H.G.; Merrifield, R.B. All-d amino acid-containing channel-forming antibiotic peptides. Proc. Natl. Acad. Sci. USA 1990, 87, 4761–4765. [Google Scholar] [CrossRef]
- Li, Z.-F.; Li, X.; Liu, H.; Liu, X.; Han, K.; Wu, Z.-H.; Hu, W.; Li, F.; Li, Y.-Z. Genome sequence of the halotolerant marine bacterium Myxococcus fulvus HW-1. J. Bacteriol. 2011, 193, 5015–5016. [Google Scholar] [CrossRef]
- Irschik, H.; Gerth, K.; Kemmer, T.; Steinmetz, H.; Reichenbach, H. The myxovalargins, new peptide antibiotics from Myxococcus fulvus (Myxobacterales). I. Cultivation, isolation, and some chemical and biological properties. J. Antibiot. 1983, 36, 6–12. [Google Scholar] [CrossRef]
- Irschik, H.; Reichenbach, H. The mechanism of action of myxovalargin A, a peptide antibiotic from Myxococcus fulvus. J. Antibiot. 1985, 38, 1237–1245. [Google Scholar] [CrossRef]
- Irschik, H.; Gerth, K.; Höfle, G.; Kohl, W.; Reichenbach, H. The myxopyronins, new inhibitors of bacterial RNA synthesis from Myxococcus fulvus (Myxobacterales). J. Antibiot. 1983, 36, 1651–1658. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Nakayama, Y.; Takeda, K.; Tawara, K.; Maeda, K.; Takeuchi, T.; Umezawa, H. A new antibiotic, althiomycin. J. Antibiot. 1957, 10, 195–200. [Google Scholar]
- Fujimoto, H.; Kinoshita, T.; Suzuki, H.; Umezawa, H. Studies on the mode of action of althiomycin. J. Antibiot. 1970, 23, 271–275. [Google Scholar] [CrossRef]
- Cortina, N.S.; Revermann, O.; Krug, D.; Müller, R. Identification and characterization of the althiomycin biosynthetic gene cluster in Myxococcus xanthus DK897. Chembiochem 2011, 12, 1411–1416. [Google Scholar] [CrossRef]
- Gerc, A.J.; Song, L.; Challis, G.L.; Stanley-Wall, N.R.; Coulthurst, S.J. The insect pathogen Serratia marcescens Db10 uses a hybrid non-ribosomal peptide synthetase-polyketide synthase to produce the antibiotic althiomycin. PLoS One 2012, 7, e44673. [Google Scholar]
- Trowitzsch, W.; Reifenstahl, G.; Wray, V.; Gerth, K. Myxothiazol, an antibiotic from Myxococcus fulvus (Myxobacterales). II. Structure elucidation. J. Antibiot. 1980, 33, 1480–1490. [Google Scholar] [CrossRef]
- Silakowski, B.; Schairer, H.U.; Ehret, H.; Kunze, B.; Weinig, S.; Nordsiek, G.; Brandt, P.; Blöcker, H.; Höfle, G.; Beyer, S.; et al. New lessons for combinatorial biosynthesis from myxobacteria. The myxothiazol biosynthetic gene cluster of Stigmatella aurantiaca DW4/3-1. J. Biol. Chem. 1999, 274, 37391–37399. [Google Scholar] [CrossRef]
- Thierbach, G.; Reichenbach, H. Myxothiazol, a new antibiotic interfering with respiration. Antimicrob. Agents Chemother. 1981, 19, 504–507. [Google Scholar] [CrossRef]
- Ahn, J.-W.; Jang, K.H.; Yang, H.-C.; Oh, K.-B.; Lee, H.-S.; Shin, J. Bithiazole metabolites from the myxobacterium Myxococcus fulvus. Chem. Pharm. Bull. 2007, 55, 477–479. [Google Scholar] [CrossRef]
- Perlova, O.; Fu, J.; Kuhlmann, S.; Krug, D.; Stewart, A.F.; Zhang, Y.; Müller, R. Reconstitution of the myxothiazol biosynthetic gene cluster by Red/ET recombination and heterologous expression in Myxococcus xanthus. Appl. Environ. Microbiol. 2006, 72, 7485–7494. [Google Scholar] [CrossRef]
- Iizuka, T.; Fudou, R.; Jojima, Y.; Ogawa, S.; Yamanaka, S.; Inukai, Y.; Ojika, M. Miuraenamides A and B, novel antimicrobial cyclic depsipeptides from a new slightly halophilic myxobacterium: Taxonomy, production, and biological properties. J. Antibiot. 2006, 59, 385–391. [Google Scholar] [CrossRef]
- Ojika, M.; Inukai, Y.; Kito, Y.; Hirata, M.; Iizuka, T.; Fudou, R. Miuraenamides: Antimicrobial cyclic depsipeptides isolated from a rare and slightly halophilic myxobacterium. Chem. Asian J. 2008, 3, 126–133. [Google Scholar] [CrossRef]
- Bewley, C.A.; Faulkner, D.J. Lithistid sponges: Star performers or hosts to the stars. Angew. Chem. Int. Ed. 1998, 37, 2162–2178. [Google Scholar] [CrossRef]
- Schmidt, E.W.; Obraztsova, A.Y.; Davidson, S.K.; Faulkner, D.J.; Haygood, M.G. Identification of the antifungal peptide-containing symbiont of the marine sponge Theonella swinhoei as a novel δ-Proteobacterium, “Candidatus Entotheonella palauensis”. Mar. Biol. 2000, 136, 969–977. [Google Scholar] [CrossRef]
- Chan, W.R.; Tinto, W.F.; Manchand, P.S.; Todaro, L.J. Stereostructures of geodiamolides A and B, novel cyclodepsipeptides from the marine sponge Geodia sp. J. Org. Chem. 1987, 52, 3091–3093. [Google Scholar] [CrossRef]
- Tanaka, C.; Tanaka, J.; Bolland, R.F.; Marriott, G.; Higa, T. Seragamides A–F, new actin-targeting depsipeptides from the sponge Suberites japonicus Thiele. Tetrahedron 2006, 62, 3536–3542. [Google Scholar] [CrossRef]
- Ishiwata, H.; Nemoto, T.; Ojika, M.; Yamada, K. Isolation and stereostructure of doliculide, a cytotoxic cyclodepsipeptide from the Japanese sea hare Dolabella auricularia. J. Org. Chem. 1994, 59, 4710–4711. [Google Scholar] [CrossRef]
- Zabriskie, T.M.; Klocke, J.A.; Ireland, C.M.; Marcus, A.H.; Molinski, T.F.; Faulkner, D.J.; Xu, C.; Clardy, J. Jaspamide, a modified peptide from a Jaspis sponge, with insecticidal and antifungal activity. J. Am. Chem. Soc. 1986, 108, 3123–3124. [Google Scholar] [CrossRef]
- D’Auria, M.V.; Gomez Paloma, L.; Minale, L.; Zampella, A.; Debitus, C.; Perez, J. Neosiphoniamolide A, a novel cyclodepsipeptide, with antifungal activity from the marine sponge Neosiphonia superstes. J. Nat. Prod. 1995, 58, 121–123. [Google Scholar] [CrossRef]
- Kunze, B.; Jansen, R.; Sasse, F.; Höfle, G.; Reichenbach, H. Chondramides A approximately D, new antifungal and cytostatic depsipeptides from Chondromyces crocatus (Myxobacteria). Production, physico-chemical and biological properties. J. Antibiot. 1995, 48, 1262–1266. [Google Scholar] [CrossRef]
- Weissman, K.J.; Müller, R. A brief tour of myxobacterial secondary metabolism. Bioorg. Med. Chem. 2009, 17, 2121–2136. [Google Scholar] [CrossRef]
- Rachid, S.; Krug, D.; Kunze, B.; Kochems, I.; Scharfe, M.; Zabriskie, T.M.; Blöcker, H.; Müller, R. Molecular and biochemical studies of chondramide formation-highly cytotoxic natural products from Chondromyces crocatus Cm c5. Chem. Biol. 2006, 13, 667–681. [Google Scholar] [CrossRef]
- Ebada, S.S.; Wray, V.; de Voogd, N.J.; Deng, Z.; Lin, W.; Proksch, P. Two new jaspamide derivatives from the marine sponge Jaspis splendens. Mar. Drugs 2009, 7, 434–444. [Google Scholar]
- Sasse, F.; Kunze, B.; Gronewold, T.M.; Reichenbach, H. The chondramides: Cytostatic agents from myxobacteria acting on the actin cytoskeleton. J. Natl. Cancer Inst. 1998, 90, 1559–1563. [Google Scholar] [CrossRef]
- Sumiya, E.; Shimogawa, H.; Sasaki, H.; Tsutsumi, M.; Yoshita, K.; Ojika, M.; Suenaga, K.; Uesugi, M. Cell-morphology profiling of a natural product library identifies bisebromoamide and miuraenamide A as actin filament stabilizers. ACS Chem. Biol. 2011, 6, 425–431. [Google Scholar] [CrossRef]
- Matcha, K.; Madduri, A.V.R.; Roy, S.; Ziegler, S.; Waldmann, H.; Hirsch, A.K.H.; Minnaard, A.J. Total synthesis of (−)-doliculide, structure-activity relationship studies and its binding to F-actin. Chembiochem 2012, 13, 2537–2548. [Google Scholar] [CrossRef]
- Freitas, V.M.; Rangel, M.; Bisson, L.F.; Jaeger, R.G.; Machado-Santelli, G.M. The geodiamolide H, derived from Brazilian sponge Geodia corticostylifera, regulates actin cytoskeleton, migration and invasion of breast cancer cells cultured in three-dimensional environment. J. Cell. Physiol. 2008, 216, 583–594. [Google Scholar] [CrossRef]
- Bewley, C.A.; Debitus, C.; Faulkner, D.J. Microsclerodermins A and B antifungal cyclic peptides from the lithistid sponge Microscleroderma sp. J. Am. Chem. Soc. 1994, 116, 7631–7636. [Google Scholar] [CrossRef]
- Kunze, B.; Böhlendorf, B.; Reichenbach, H.; Höfle, G. Pedein A and B: Production, isolation, structure elucidation and biological properties of new antifungal cyclopeptides from Chondromyces pediculatus (Myxobacteria). J. Antibiot. 2008, 61, 18–26. [Google Scholar] [CrossRef]
- Arai, T.; Takahashi, K.; Nakahara, S.; Kubo, A. The structure of a novel antitumor antibiotic, saframycin A. Experientia 1980, 36, 1025–1027. [Google Scholar] [CrossRef]
- Frincke, J.M.; Faulkner, D.J. Antimicrobial metabolites of the sponge Reniera sp. J. Am. Chem. Soc. 1982, 104, 265–269. [Google Scholar] [CrossRef]
- Irschik, H.; Trowitzsch-Kienast, W.; Gerth, K.; Höfle, G.; Reichenbach, H. Saframycin Mx1, a new natural saframycin isolated from a myxobacterium. J. Antibiot. 1988, 41, 993–998. [Google Scholar] [CrossRef]
- Pospiech, A.; Cluzel, B.; Bietenhader, J.; Schupp, T. A new Myxococcus xanthus gene cluster for the biosynthesis of the antibiotic saframycin Mx1 encoding a peptide synthetase. Microbiology 1995, 141, 1793–1803. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Holt, T.G.; Fregeau, N.L.; Stroh, J.G.; Keifer, P.A.; Sun, F.; Li, L.H.; Martin, D.G. Ecteinascidins 729, 743, 745, 759A, 759B, and 770: Potent antitumor agents from the Caribbean tunicate Ecteinascidia turbinata. J. Org. Chem. 1990, 55, 4512–4515. [Google Scholar] [CrossRef]
- Grohar, P.J.; Griffin, L.B.; Yeung, C.; Chen, Q.-R.; Pommier, Y.; Khanna, C.; Khan, J.; Helman, L.J. Ecteinascidin 743 interferes with the activity of EWS-FLI1 in Ewing sarcoma cells. Neoplasia 2011, 13, 145–153. [Google Scholar]
- Erba, E.; Bergamaschi, D.; Bassano, L.; Damia, G.; Ronzoni, S.; Faircloth, G.T.; D’Incalci, M. Ecteinascidin-743 (ET-743), a natural marine compound, with a unique mechanism of action. Eur. J. Cancer 2001, 37, 97–105. [Google Scholar]
- Garcia, R.O.; Krug, D.; Müller, R. Discovering natural products from myxobacteria with emphasis on rare producer strains in combination with improved analytical methods. Meth. Enzymol. 2009, 458, 59–91. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Desriac, F.; Jégou, C.; Balnois, E.; Brillet, B.; Chevalier, P.L.; Fleury, Y. Antimicrobial Peptides from Marine Proteobacteria. Mar. Drugs 2013, 11, 3632-3660. https://doi.org/10.3390/md11103632
Desriac F, Jégou C, Balnois E, Brillet B, Chevalier PL, Fleury Y. Antimicrobial Peptides from Marine Proteobacteria. Marine Drugs. 2013; 11(10):3632-3660. https://doi.org/10.3390/md11103632
Chicago/Turabian StyleDesriac, Florie, Camille Jégou, Eric Balnois, Benjamin Brillet, Patrick Le Chevalier, and Yannick Fleury. 2013. "Antimicrobial Peptides from Marine Proteobacteria" Marine Drugs 11, no. 10: 3632-3660. https://doi.org/10.3390/md11103632