Delivery of RNAi-Based Oligonucleotides by Electropermeabilization

{kind=link}
Abstract
:1. Introduction
2. How to Perform Electrotransfer of RNAi-Based Oligonucleotides
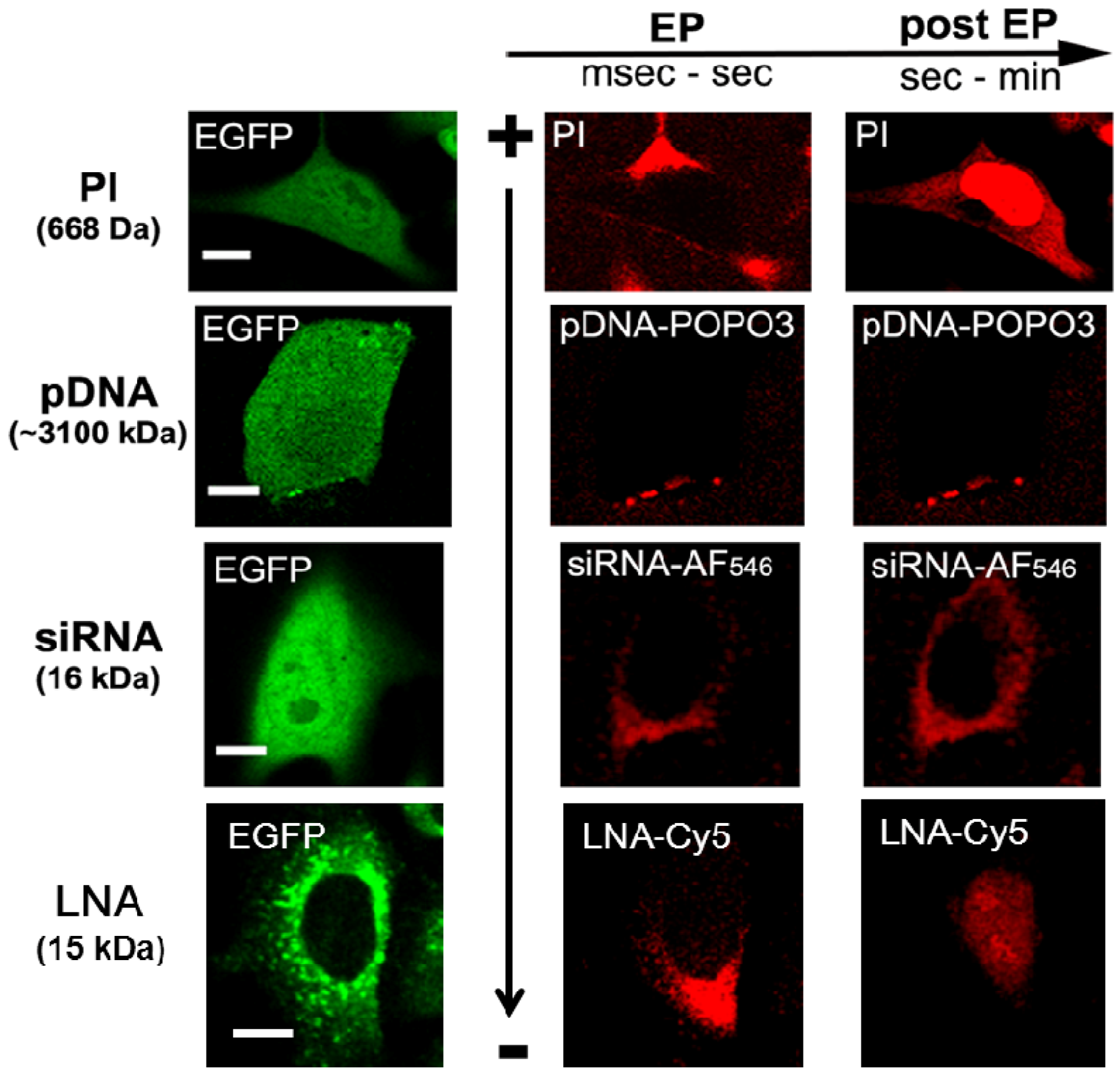
3. Electrotransfer of RNAi-Based Oligonucleotides and Mechanisms

4. Conclusions
Acknowledgments
Conflict of Interest
References
- Bumcrot, D.; Manoharan, M.; Koteliansky, V.; Sah, D.W. RNAi therapeutics: A potential new class of pharmaceutical drugs. Nat. Chem. Biol. 2006, 2, 711–719. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar]
- Tong, A.W.; Zhang, Y.A.; Nemunaitis, J. Small interfering RNA for experimental cancer therapy. Curr. Opin. Mol. Ther. 2005, 7, 114–124. [Google Scholar]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef]
- Whitehead, K.A.; Langer, R.; Anderson, D.G. Knocking down barriers: Advances in siRNA delivery. Nat. Rev. Drug Discov. 2009, 8, 129–138. [Google Scholar] [CrossRef]
- Garzon, R.; Marcucci, G.; Croce, C.M. Targeting microRNAs in cancer: Rationale, strategies and challenges. Nat. Rev. Drug Discov. 2010, 9, 775–789. [Google Scholar] [CrossRef]
- Cemazar, M.; Golzio, M.; Escoffre, J.M.; Couderc, B.; Sersa, G.; Teissie, J. In vivo imaging of tumor growth after electrochemotherapy with cisplatin. Biochem. Biophys. Res. Commun. 2006, 348, 997–1002. [Google Scholar] [CrossRef]
- Cemazar, M.; Golzio, M.; Sersa, G.; Hojman, P.; Kranjc, S.; Mesojednik, S.; Rols, M.P.; Teissie, J. Control by pulse parameters of DNA electrotransfer into solid tumors in mice. Gene Ther. 2009, 16, 635–644. [Google Scholar] [CrossRef]
- Golzio, M.; Mazzolini, L.; Paganin-Gioanni, A.; Teissie, J. Targeted gene silencing into solid tumors with electrically mediated siRNA delivery. Methods Mol. Biol. 2009, 555, 15–27. [Google Scholar]
- Coster, H.G. A quantitative analysis of the voltage-current relationships of fixed charge membranes and the associated property of “punch-through”. Biophys. J. 1965, 5, 669–686. [Google Scholar] [CrossRef]
- Teissie, J.; Golzio, M.; Rols, M.P. Mechanisms of cell membrane electropermeabilization: A minireview of our present (lack of ?) knowledge. Biochim. Biophys. Acta 2005, 1724, 270–280. [Google Scholar] [CrossRef]
- Hu, W.Y.; Myers, C.P.; Kilzer, J.M.; Pfaff, S.L.; Bushman, F.D. Inhibition of retroviral pathogenesis by RNA interference. Curr. Biol. 2002, 15, 1301–1311. [Google Scholar]
- Calegari, F.; Haubensak, W.; Yang, D.; Huttner, W.B.; Buchholz, F. Tissue-specific RNA interference in postimplantation mouse embryos with endoribonuclease-prepared short interfering RNA. Proc. Natl. Acad. Sci. USA 2002, 22, 14236–14240. [Google Scholar]
- Pekarik, V.; Bourikas, D.; Miglino, N.; Joset, P.; Preiswerk, S.; Stoeckli, E.T. Screening for gene function in chicken embryo using RNAi and electroporation. Nat. Biotechnol. 2003, 21, 93–96. [Google Scholar]
- Golzio, M.; Mazzolini, L.; Ledoux, A.; Paganin, A.; Izard, M.; Hellaudais, L.; Bieth, A.; Pillaire, M.J.; Cazaux, C.; Hoffmann, J.S.; et al. In vivo gene silencing in solid tumors by targeted electrically mediated siRNA delivery. Gene Ther. 2007, 14, 752–759. [Google Scholar] [CrossRef]
- Lewis, D.L.; Hagstrom, J.E.; Loomis, A.G.; Wolff, J.A.; Herweijer, H. Efficient delivery of siRNA for inhibition of gene expression in postnatal mice. Nat. Genet. 2002, 32, 107–108. [Google Scholar]
- Matsuda, T.; Cepko, C.L. Electroporation and RNA interference in the rodent retina in vivo and in vitro. Proc. Natl. Acad. Sci. USA 2004, 101, 16–22. [Google Scholar] [CrossRef]
- Paganin-Gioanni, A.; Bellard, E.; Couderc, B.; Teissie, J.; Golzio, M. Tracking in vitro and in vivo siRNA electrotransfer in tumor cells. J. RNAi Gene Silencing 2008, 4, 281–288. [Google Scholar]
- Wells, D.J. Gene therapy progress and prospects: Electroporation and other physical methods. Gene Ther. 2004, 11, 1363–1369. [Google Scholar] [CrossRef]
- Heller, L.C.; Heller, R. In vivo electroporation for gene therapy. Hum. Gene Ther. 2006, 17, 890–897. [Google Scholar] [CrossRef]
- Mir, L.M.; Glass, L.F.; Sersa, G.; Teissie, J.; Domenge, C.; Miklavcic, D.; Jaroszeski, M.J.; Orlowski, S.; Reintgen, D.S.; Rudolf, Z.; et al. Effective treatment of cutaneous and subcutaneous malignant tumours by electrochemotherapy. Br. J. Cancer 1998, 77, 2336–2342. [Google Scholar] [CrossRef]
- Sersa, G.; Stabuc, B.; Cemazar, M.; Miklavcic, D.; Rudolf, Z. Electrochemotherapy with cisplatin: the systemic antitumour effectiveness of cisplatin can be potentiated locally by the application of electric pulses in the treatment of malignant melanoma skin metastases. Melanoma Res. 2000, 10, 381–385. [Google Scholar] [CrossRef]
- Gothelf, A.; Mir, L.M.; Gehl, J. Electrochemotherapy: results of cancer treatment using enhanced delivery of bleomycin by electroporation. Cancer Treat Rev. 2003, 29, 371–387. [Google Scholar] [CrossRef]
- Sersa, G.; Miklavcic, D.; Cemazar, M.; Rudolf, Z.; Pucihar, G.; Snoj, M. Electrochemotherapy in treatment of tumours. Eur. J. Surg. Oncol. 2008, 34, 232–240. [Google Scholar] [CrossRef]
- Daud, A.I.; DeConti, R.C.; Andrews, S.; Urbas, P.; Riker, A.I.; Sondak, V.K.; Munster, P.N.; Sullivan, D.M.; Ugen, K.E.; Messina, J.L. Phase I trial of interleukin-12 plasmid electroporation in patients with metastatic melanoma. J. Clin. Oncol. 2008, 26, 5896–5903. [Google Scholar]
- Bodles-Brakhop, A.M.; Heller, R.; Draghia-Akli, R. Electroporation for the delivery of DNA-based vaccines and immunotherapeutics: current clinical developments. Mol. Ther. 2009, 17, 585–592. [Google Scholar] [CrossRef]
- Mali, B.; Jarm, T.; Snoj, M.; Sersa, G.; Miklavcic, D. Antitumor effectiveness of electrochemotherapy: A systematic review and meta-analysis. Eur. J. Surg. Oncol. 2013, 39, 4–16. [Google Scholar] [CrossRef]
- Mir, L.M.; Devauchelle, P.; Quintin-Colonna, F.; Delisle, F.; Doliger, S.; Fradelizi, D.; Belehradek, J., Jr.; Orlowski, S. First clinical trial of cat soft-tissue sarcomas treatment by electrochemotherapy. Br. J. Cancer 1997, 76, 1617–1622. [Google Scholar] [CrossRef]
- Tozon, N.; Kodre, V.; Sersa, G.; Cemazar, M. Effective treatment of perianal tumors in dogs with electrochemotherapy. Anticancer Res. 2005, 25, 839–845. [Google Scholar]
- Kodre, V.; Cemazar, M.; Pecar, J.; Sersa, G.; Cor, A.; Tozon, N. Electrochemotherapy compared to surgery for treatment of canine mast cell tumours. In Vivo 2009, 23, 55–62. [Google Scholar]
- Rols, M.P.; Tamzali, Y.; Teissie, J. Electrochemotherapy of horses. A preliminary clinical report. Bioelectrochemistry 2002, 55, 101–105. [Google Scholar] [CrossRef]
- Aihara, H.; Miyazaki, J. Gene transfer into muscle by electroporation in vivo. Nat. Biotechnol. 1998, 16, 867–870. [Google Scholar]
- Mir, L.M.; Bureau, M.F.; Gehl, J.; Rangara, R.; Rouy, D.; Caillaud, J.M.; Delaere, P.; Branellec, D.; Schwartz, B.; Scherman, D. High-efficiency gene transfer into skeletal muscle mediated by electric pulses. Proc. Natl. Acad. Sci. USA 1999, 96, 4262–4267. [Google Scholar] [CrossRef]
- Dean, D.A. Nonviral gene transfer to skeletal, smooth, and cardiac muscle in living animals. Am. J. Physiol. Cell. Physiol. 2005, 289, C233–C245. [Google Scholar] [CrossRef]
- Heller, R.; Jaroszeski, M.; Atkin, A.; Moradpour, D.; Gilbert, R.; Wands, J.; Nicolau, C. In vivo gene electroinjection and expression in rat liver. FEBS Lett. 1996, 389, 225–228. [Google Scholar] [CrossRef]
- Liu, F.; Huang, L. Electric gene transfer to the liver following systemic administration of plasmid DNA. Gene Ther. 2002, 9, 1116–1119. [Google Scholar] [CrossRef]
- Titomirov, A.V.; Sukharev, S.; Kistanova, E. In vivo electroporation and stable transformation of skin cells of newborn mice by plasmid DNA. Biochim. Biophys. Acta 1991, 1088, 131–134. [Google Scholar] [CrossRef]
- Vandermeulen, G.; Staes, E.; Vanderhaeghen, M.L.; Bureau, M.F.; Scherman, D.; Preat, V. Optimisation of intradermal DNA electrotransfer for immunisation. J. Control Release 2007, 124, 81–87. [Google Scholar] [CrossRef]
- Tupin, E.; Poirier, B.; Bureau, M.F.; Khallou-Laschet, J.; Vranckx, R.; Caligiuri, G.; Gaston, A.T.; Duong van Huyen, J.P.; Scherman, D.; Bariety, J.; et al. Non-viral gene transfer of murine spleen cells achieved by in vivo electroporation. Gene Ther. 2003, 10, 569–579. [Google Scholar] [CrossRef]
- Isaka, Y.; Yamada, K.; Takabatake, Y.; Mizui, M.; Miura-Tsujie, M.; Ichimaru, N.; Yazawa, K.; Utsugi, R.; Okuyama, A.; Hori, M.; et al. Electroporation-mediated HGF gene transfection protected the kidney against graft injury. Gene Ther. 2005, 12, 815–820. [Google Scholar] [CrossRef]
- Saito, T.; Nakatsuji, N. Efficient gene transfer into the embryonic mouse brain using in vivo electroporation. Dev. Biol. 2001, 240, 237–246. [Google Scholar] [CrossRef]
- Khoury, M.; Bigey, P.; Louis-Plence, P.; Noel, D.; Rhinn, H.; Scherman, D.; Jorgensen, C.; Apparailly, F. A comparative study on intra-articular versus systemic gene electrotransfer in experimental arthritis. J. Gene Med. 2006, 8, 1027–1036. [Google Scholar] [CrossRef]
- Pavlin, D.; Cemazar, M.; Cör, A.; Sersa, G.; Pogacnik, A.; Tozon, N. Electrogene therapy with interleukin-12 in canine mast cell tumors. Radiol. Oncol. 2011, 45, 31–39. [Google Scholar]
- Bureau, M.F.; Gehl, J.; Deleuze, V.; Mir, L.M.; Scherman, D. Importance of association between permeabilization and electrophoretic forces for intramuscular DNA electrotransfer. Biochim. Biophys. Acta 2000, 1474, 353–359. [Google Scholar] [CrossRef]
- Satkauskas, S.; Bureau, M.F.; Puc, M.; Mahfoudi, A.; Scherman, D.; Miklavcic, D.; Mir, L.M. Mechanisms of in vivo DNA electrotransfer: respective contributions of cell electropermeabilization and DNA electrophoresis. Mol. Ther. 2002, 5, 133–140. [Google Scholar] [CrossRef]
- Sel, D.; Golzio, M.; Pucihar, G.; Tamzali, Y.; Miklavcic, D.; Teissie, J. Non invasive contact electrodes for in vivo localized cutaneous electropulsation and associated drug and nucleic acid delivery. J. Control Release 2009, 134, 125–131. [Google Scholar] [CrossRef]
- Gehl, J.; Sorensen, T.H.; Nielsen, K.; Raskmark, P.; Nielsen, S.L.; Skovsgaard, T.; Mir, L.M. In vivo electroporation of skeletal muscle: Threshold, efficacy and relation to electric field distribution. Biochim. Biophys. Acta 1999, 1428, 233–240. [Google Scholar] [CrossRef]
- Golzio, M.; Mazzolini, L.; Moller, P.; Rols, M.P.; Teissie, J. Inhibition of gene expression in mice muscle by in vivo electrically mediated siRNA delivery. Gene Ther. 2005, 12, 246–251. [Google Scholar] [CrossRef]
- Spugnini, E.P.; Citro, G.; Porrello, A. Rational design of new electrodes for electrochemotherapy. J. Exp. Clin. Cancer Res. 2005, 24, 245–254. [Google Scholar]
- Tjelle, T.E.; Salte, R.; Mathiesen, I.; Kjeken, R. A novel electroporation device for gene delivery in large animals and humans. Vaccine 2006, 24, 4667–4670. [Google Scholar] [CrossRef]
- Cemazar, M.; Golzio, M.; Sersa, G.; Rols, M.P.; Teissié, J. Electrically-assisted nucleic acids delivery to tissues in vivo: where do we stand? Curr. Pharm. Des. 2006, 12, 3817–3825. [Google Scholar] [CrossRef]
- Golzio, M.; Teissie, J.; Rols, M.P. Direct visualization at the single-cell level of electrically mediated gene delivery. Proc. Natl. Acad. Sci. USA 2002, 99, 1292–1297. [Google Scholar] [CrossRef]
- Escoffre, J.M.; Portet, T.; Wasungu, L.; Teissie, J.; Dean, D.; Rols, M.P. What is (still not) known of the mechanism by which electroporation mediates gene transfer and expression in cells and tissues. Mol. Biotechnol. 2009, 41, 286–295. [Google Scholar] [CrossRef]
- Puc, M.; Kotnik, T.; Mir, L.M.; Miklavcic, D. Quantitative model of small molecules uptake after in vitro cell electropermeabilization. Bioelectrochemistry 2003, 60, 1–10. [Google Scholar] [CrossRef]
- Chabot, S.; Orio, J.; Castanier, R.; Bellard, E.; Nielsen, S.J.; Golzio, M.; Teissié, J. LNA-based oligonucleotide electrotransfer for miRNA inhibition. Mol. Ther. 2012, 20, 1590–1598. [Google Scholar] [CrossRef]
- Castanotto, D.; Rossi, J.J. The promises and pitfalls of RNA-interference-based therapeutics. Nature 2009, 457, 426–433. [Google Scholar]
- Sigoillot, F.D.; King, R.W. Vigilance and validation: Keys to success in RNAi screening. ACS Chem. Biol. 2011, 6, 47–60. [Google Scholar] [CrossRef]
- Heidel, J.D.; Hu, S.; Liu, X.F.; Triche, T.J.; Davis, M.E. Lack of interferon response in animals to naked siRNAs. Nat. Biotechnol. 2004, 22, 1579–1582. [Google Scholar]
- Paganin-Gioanni, A.; Bellard, E.; Escoffre, J.M.; Rols, M.P.; Teissie, J.; Golzio, M. Direct visualization at the single-cell level of siRNA electrotransfer into cancer cells. Proc. Natl. Acad. Sci. USA 2011, 108, 10443–10447. [Google Scholar]
- Kishida, T.; Asada, H.; Gojo, S.; Ohashi, S.; Shin-Ya, M.; Yasutomi, K.; Terauchi, R.; Takahashi, K.A.; Kubo, T.; Imanishi, J.; et al. Sequence-specific gene silencing in murine muscle induced by electroporation-mediated transfer of short interfering RNA. J. Gene Med. 2004, 6, 105–110. [Google Scholar] [CrossRef]
- Inoue, A.; Takahashi, K.A.; Mazda, O.; Terauchi, R.; Arai, Y.; Kishida, T.; Shin-Ya, M.; Asada, H.; Morihara, T.; Tonomura, H.; et al. Electro-transfer of small interfering RNA ameliorated arthritis in rats. Biochem. Biophys. Res. Commun. 2005, 336, 903–908. [Google Scholar] [CrossRef]
- Akaneya, Y.; Jiang, B.; Tsumoto, T. RNAi-induced gene silencing by local electroporation in targeting brain region. J. Neurophysiol. 2005, 93, 594–602. [Google Scholar] [CrossRef]
- Takabatake, Y.; Isaka, Y.; Mizui, M.; Kawachi, H.; Shimizu, F.; Ito, T.; Hori, M.; Imai, E. Exploring RNA interference as a therapeutic strategy for renal disease. Gene Ther. 2005, 12, 965–973. [Google Scholar] [CrossRef]
- Broderick, K.E.; Chan, A.; Lin, F.; Shen, X.; Kichaev, G.; Khan, A.S.; Aubin, J.; Zimmermann, T.S.; Sardesai, N.Y. Optimized in vivo transfer of small interfering RNA targeting dermal tissue using in vivo surface electroporation. Mol. Ther. Nucleic Acids 2012, 14, e11. [Google Scholar]
- Nakai, N.; Kishida, T.; Shin-Ya, M.; Imanishi, J.; Ueda, Y.; Kishimoto, S.; Mazda, O. Therapeutic RNA interference of malignant melanoma by electrotransfer of small interfering RNA targeting Mitf. Gene Ther. 2007, 14, 357–365. [Google Scholar] [CrossRef]
- McCaffrey, A.P.; Meuse, L.; Pham, T.T.; Conklin, D.S.; Hannon, G.J.; Kay, M.A. RNA interference in adult mice. Nature 2002, 418, 38–39. [Google Scholar]
- Jackson, A.L.; Linsley, P.S. Recognizing and avoiding siRNA off-target effects for target identification and therapeutic application. Nat. Rev. Drug Discov. 2010, 9, 57–67. [Google Scholar] [CrossRef]
- Ambros, V. MicroRNAs: tiny regulators with great potential. Cell 2001, 107, 823–826. [Google Scholar] [CrossRef]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar]
- Hernando, E. MicroRNAs and cancer: Role in tumorigenesis, patient classification and therapy. Clin. Transl. Oncol. 2007, 9, 155–160. [Google Scholar] [CrossRef]
- Da Costa Martins, P.A.; Salic, K.; Gladka, M.M.; Armand, A.S.; Leptidis, S.; El Azzouzi, H.; Hansen, A.; Coenen-de Roo, C.J.; Bierhuizen, M.F.; van der Nagel, R.; et al. MicroRNA-199b targets the nuclear kinase Dyrk1a in an auto-amplification loop promoting calcineurin/NFAT signalling. Nat. Cell. Biol. 2010, 12, 1220–1227. [Google Scholar] [CrossRef]
- Long, J.; Wang, Y.; Wang, W.; Chang, B.H.; Danesh, F.R. Identification of microRNA-93 as a novel regulator of vascular endothelial growth factor in hyperglycemic conditions. J. Biol. Chem. 2010, 285, 23457–23465. [Google Scholar]
- Brodersen, P.; Voinnet, O. Revisiting the principles of microRNA target recognition and mode of action. Nat. Rev. Mol. Cell. Biol. 2009, 10, 141–148. [Google Scholar] [CrossRef]
- Stenvang, J.; Kauppinen, S. MicroRNAs as targets for antisense-based therapeutics. Expert Opin. Biol. Ther. 2008, 8, 59–81. [Google Scholar] [CrossRef]
- Stenvang, J.; Lindow, M.; Kauppinen, S. Targeting of microRNAs for therapeutics. Biochem. Soc. Trans. 2008, 36, 1197–1200. [Google Scholar] [CrossRef]
- Dalmay, T. MicroRNAs and cancer. J. Intern. Med. 2008, 263, 366–375. [Google Scholar] [CrossRef]
- Care, A.; Catalucci, D.; Felicetti, F.; Bonci, D.; Addario, A.; Gallo, P.; Bang, M.L.; Segnalini, P.; Gu, Y.; Dalton, N.D.; et al. MicroRNA-133 controls cardiac hypertrophy. Nat. Med. 2007, 13, 613–618. [Google Scholar]
- Mercatelli, N.; Coppola, V.; Bonci, D.; Miele, F.; Costantini, A.; Guadagnoli, M.; Bonanno, E.; Muto, G.; Frajese, G.V.; de Maria, R.; et al. The inhibition of the highly expressed miR-221 and miR-222 impairs the growth of prostate carcinoma xenografts in mice. PLoS One 2008, 3, e4029. [Google Scholar]
- Clape, C.; Fritz, V.; Henriquet, C.; Apparailly, F.; Fernandez, P.L.; Iborra, F.; Avances, C.; Villalba, M.; Culine, S.; Fajas, L. miR-143 interferes with ERK5 signaling, and abrogates prostate cancer progression in mice. PLoS One 2009, 4, e7542. [Google Scholar]
- Layzer, J.M.; McCaffrey, A.P.; Tanner, A.K.; Huang, Z.; Kay, M.A.; Sullenger, B.A. In vivo activity of nuclease-resistant siRNAs. RNA 2004, 10, 766–771. [Google Scholar] [CrossRef]
- Raemdonck, K.; Remaut, K.; Lucas, B.; Sanders, N.N.; Demeester, J.; De Smedt, S.C. In situ analysis of single-stranded and duplex siRNA integrity in living cells. Biochemistry 2006, 45, 10614–10623. [Google Scholar] [CrossRef]
- Corey, D.R. Chemical modification: The key to clinical application of RNA interference? J. Clin. Invest. 2007, 117, 3615–3622. [Google Scholar] [CrossRef]
- Braasch, D.A.; Jensen, S.; Liu, Y.; Kaur, K.; Arar, K.; White, M.A.; Corey, D.R. RNA interference in mammalian cells by chemically-modified RNA. Biochemistry 2003, 42, 7967–7975. [Google Scholar]
- Kumar, R.; Singh, S.K.; Koshkin, A.A.; Rajwanshi, V.K.; Meldgaard, M.; Wengel, J. The first analogues of LNA (locked nucleic acids): Phosphorothioate-LNA and 2'-thio-LNA. Bioorg. Med. Chem. Lett. 1998, 8, 2219–2222. [Google Scholar] [CrossRef]
- Petersen, M.; Nielsen, C.B.; Nielsen, K.E.; Jensen, G.A.; Bondensgaard, K.; Singh, S.K.; Rajwanshi, V.K.; Koshkin, A.A.; Dahl, B.M.; Wengel, J.; et al. The conformations of locked nucleic acids (LNA). J. Mol. Recognit. 2000, 13, 44–53. [Google Scholar] [CrossRef]
- Kaur, H.; Babu, B.R.; Mait, S. Perspectives on chemistry and therapeutic applications of Locked Nucleic Acid (LNA). Chem. Rev. 2007, 107, 4672–4697. [Google Scholar] [CrossRef]
- Crinelli, R.; Bianchi, M.; Gentilini, L.; Magnani, M. Design and characterization of decoy oligonucleotides containing locked nucleic acids. Nucleic Acids Res. 2002, 30, 2435–2443. [Google Scholar] [CrossRef]
- Wahlestedt, C.; Salmi, P.; Good, L.; Kela, J.; Johnsson, T.; Hokfelt, T.; Broberger, C.; Porreca, F.; Lai, J.; Ren, K.; et al. Potent and nontoxic antisense oligonucleotides containing locked nucleic acids. Proc. Natl. Acad. Sci. USA 2000, 97, 5633–5638. [Google Scholar] [CrossRef]
- Lanford, R.E.; Hildebrandt-Eriksen, E.S.; Petri, A.; Persson, R.; Lindow, M.; Munk, M.E.; Kauppinen, S.; Orum, H. Therapeutic silencing of microRNA-122 in primates with chronic hepatitis C virus infection. Science 2010, 327, 198–201. [Google Scholar] [CrossRef]
- Braasch, D.A.; Liu, Y.; Corey, D.R. Antisense inhibition of gene expression in cells by oligonucleotides incorporating locked nucleic acids: Effect of mRNA target sequence and chimera design. Nucleic Acids Res. 2002, 30, 5160–5167. [Google Scholar] [CrossRef]
- Corsten, M.F.; Miranda, R.; Kasmieh, R.; Krichevsky, A.M.; Weissleder, R.; Shah, K. MicroRNA-21 knockdown disrupts glioma growth in vivo and displays synergistic cytotoxicity with neural precursor cell delivered S-TRAIL in human gliomas. Cancer Res. 2007, 67, 8994–9000. [Google Scholar] [CrossRef]
- Pelofy, S.; Teissié, J.; Golzio, M.; Chabot, S. Chemically modified oligonucleotide-increased stability negatively correlates with its efficacy despite efficient electrotransfer. J. Membr. Biol. 2012, 245, 565–571. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chabot, S.; Pelofy, S.; Teissié, J.; Golzio, M. Delivery of RNAi-Based Oligonucleotides by Electropermeabilization. Pharmaceuticals 2013, 6, 510-521. https://doi.org/10.3390/ph6040510
Chabot S, Pelofy S, Teissié J, Golzio M. Delivery of RNAi-Based Oligonucleotides by Electropermeabilization. Pharmaceuticals. 2013; 6(4):510-521. https://doi.org/10.3390/ph6040510
Chicago/Turabian StyleChabot, Sophie, Sandrine Pelofy, Justin Teissié, and Muriel Golzio. 2013. "Delivery of RNAi-Based Oligonucleotides by Electropermeabilization" Pharmaceuticals 6, no. 4: 510-521. https://doi.org/10.3390/ph6040510