Advances in Drug Design Based on the Amino Acid Approach: Taurine Analogues for the Treatment of CNS Diseases


Abstract
:1. Introduction
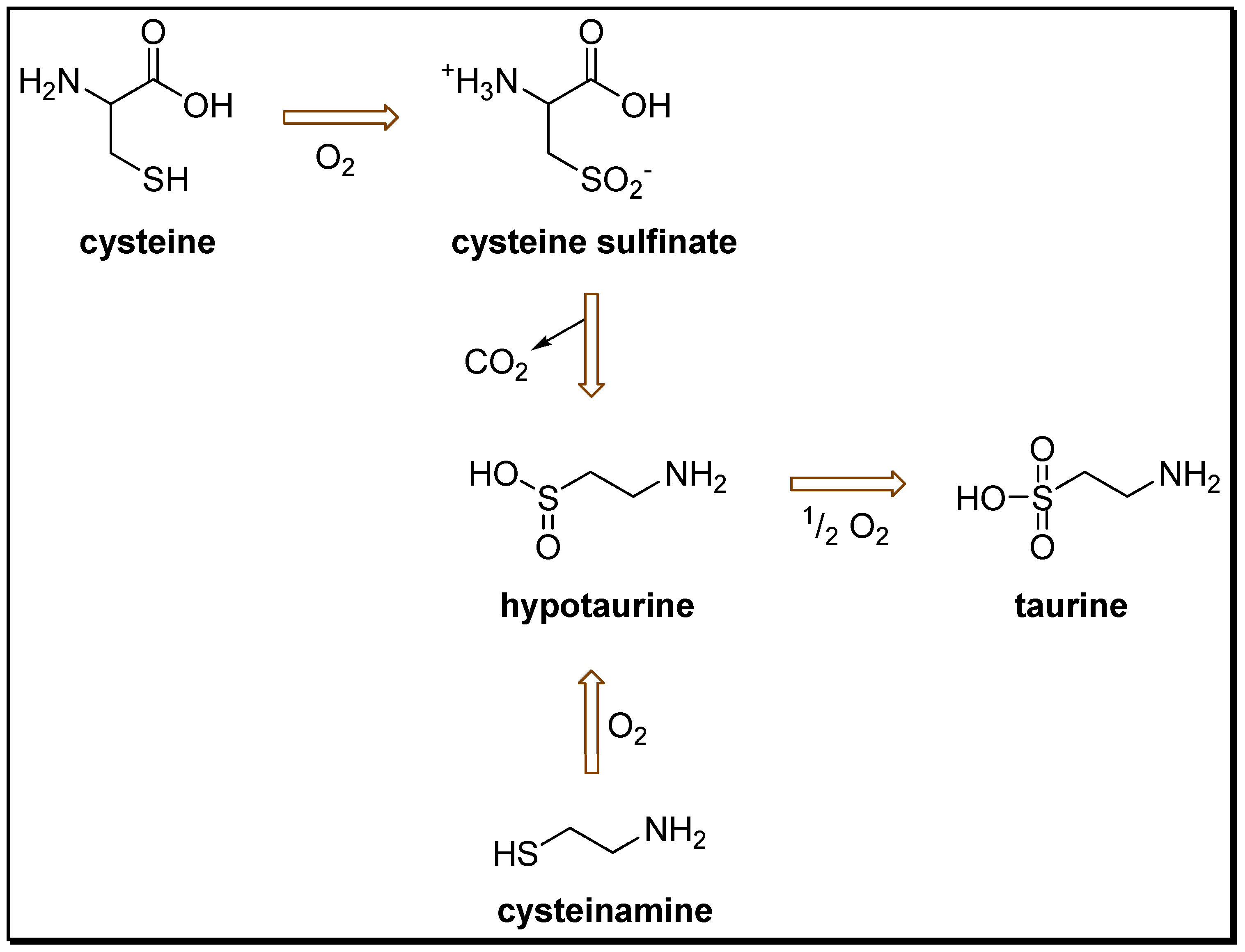
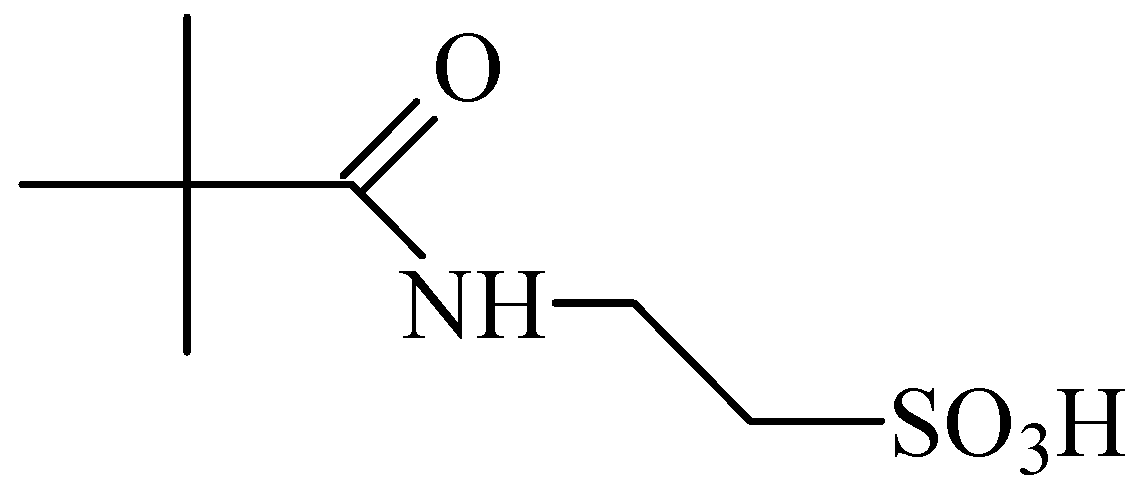
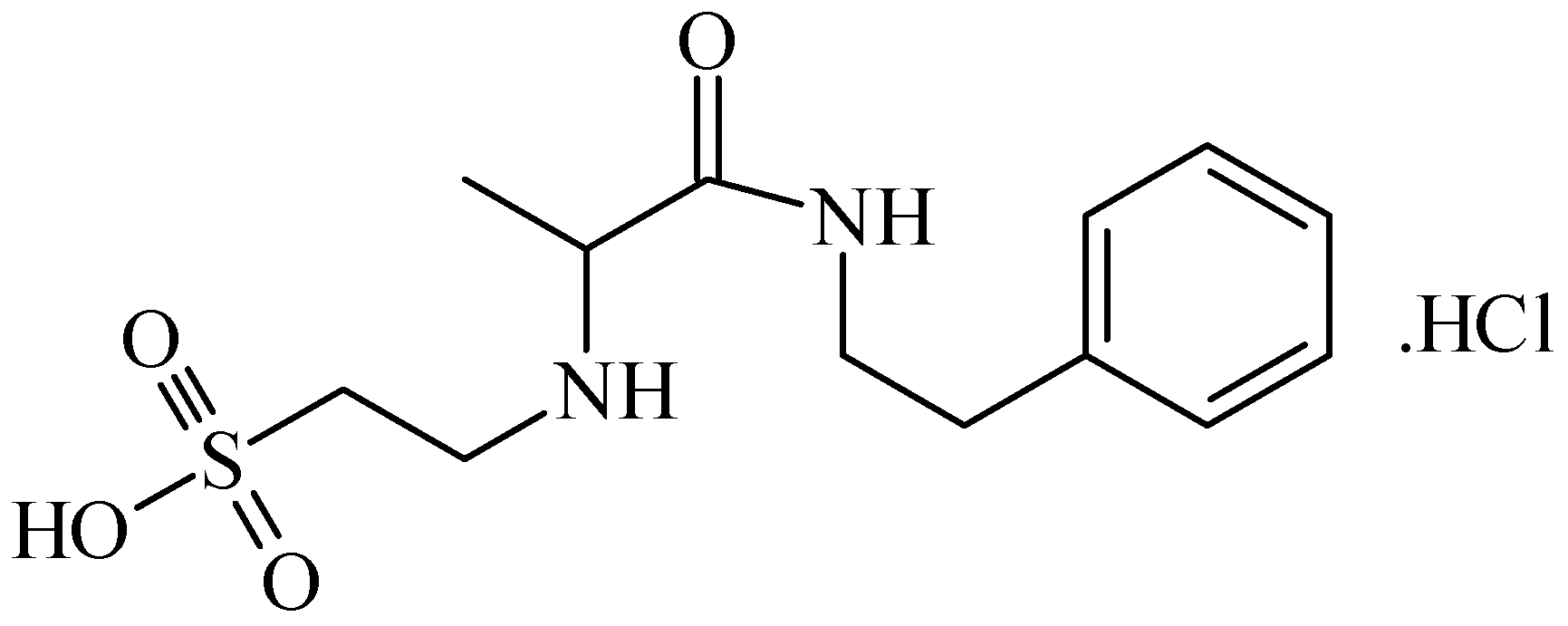
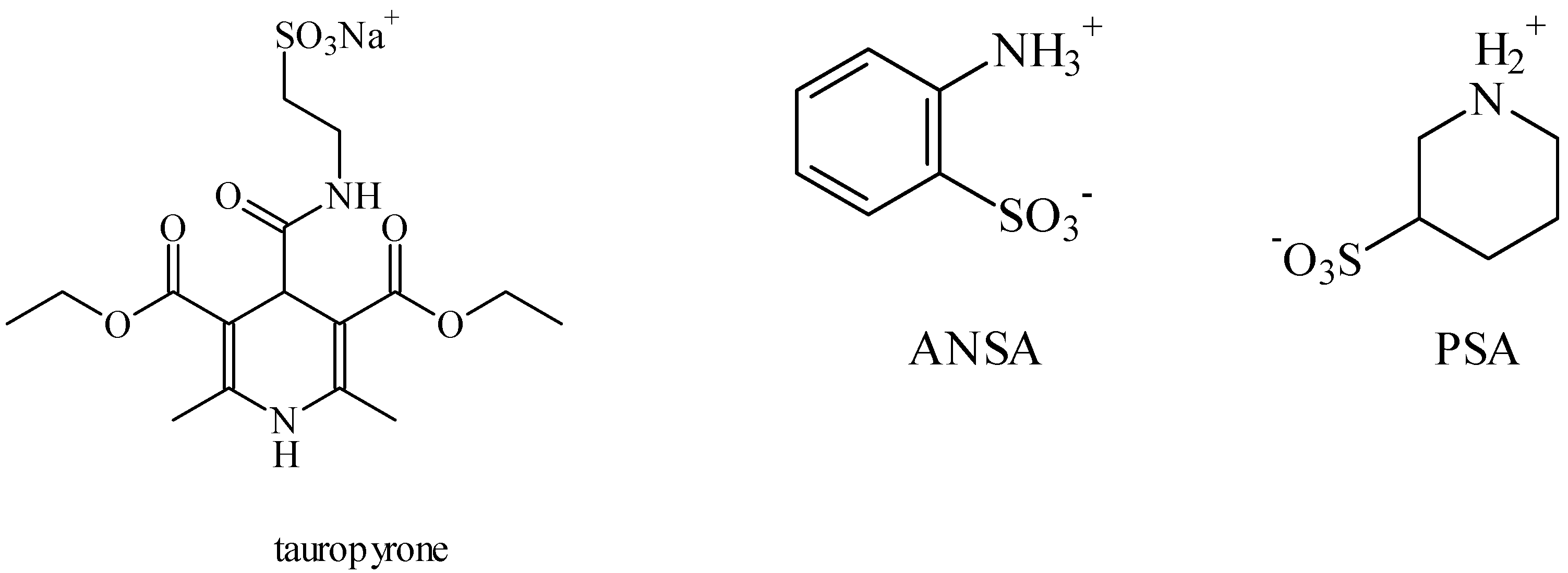
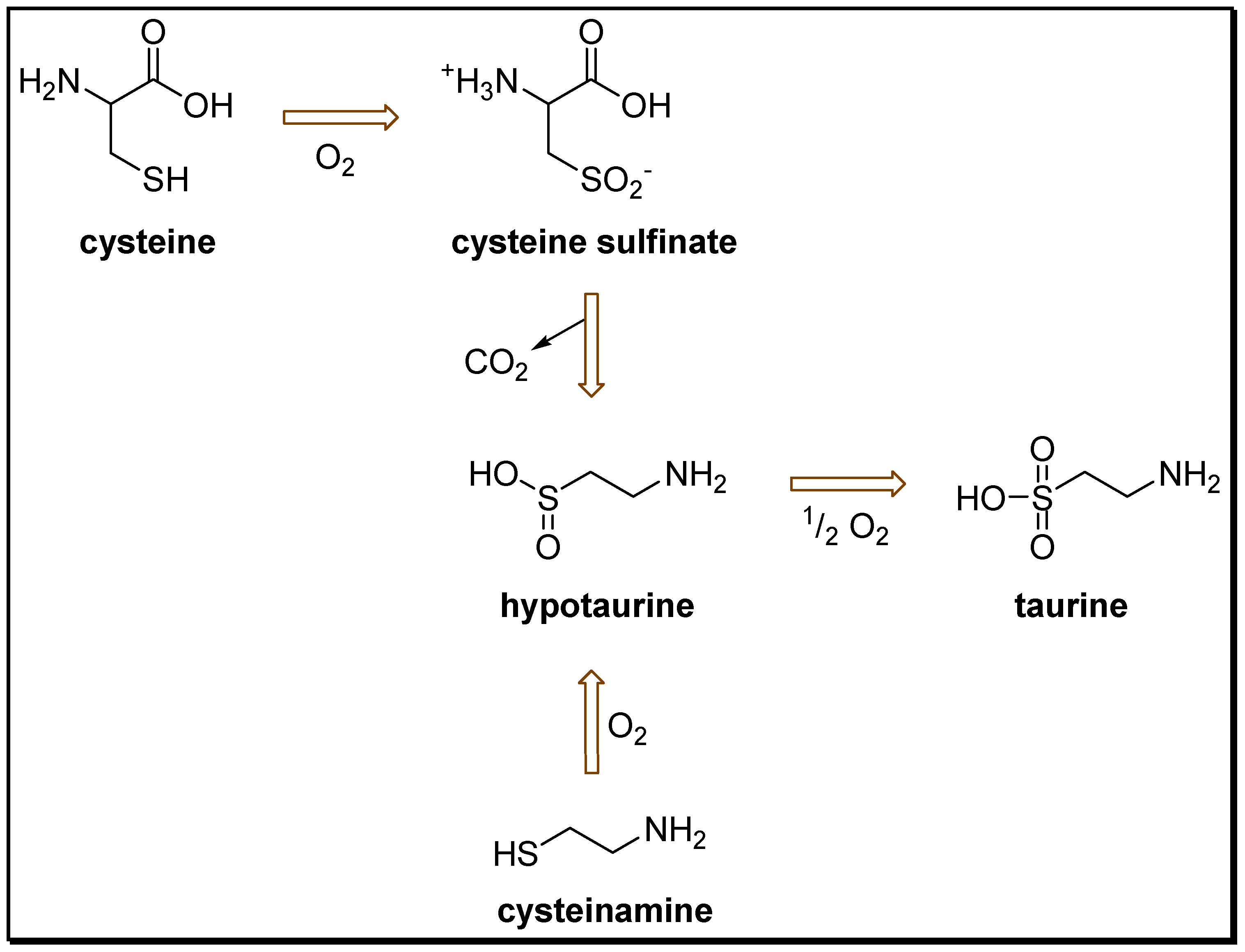
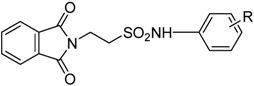
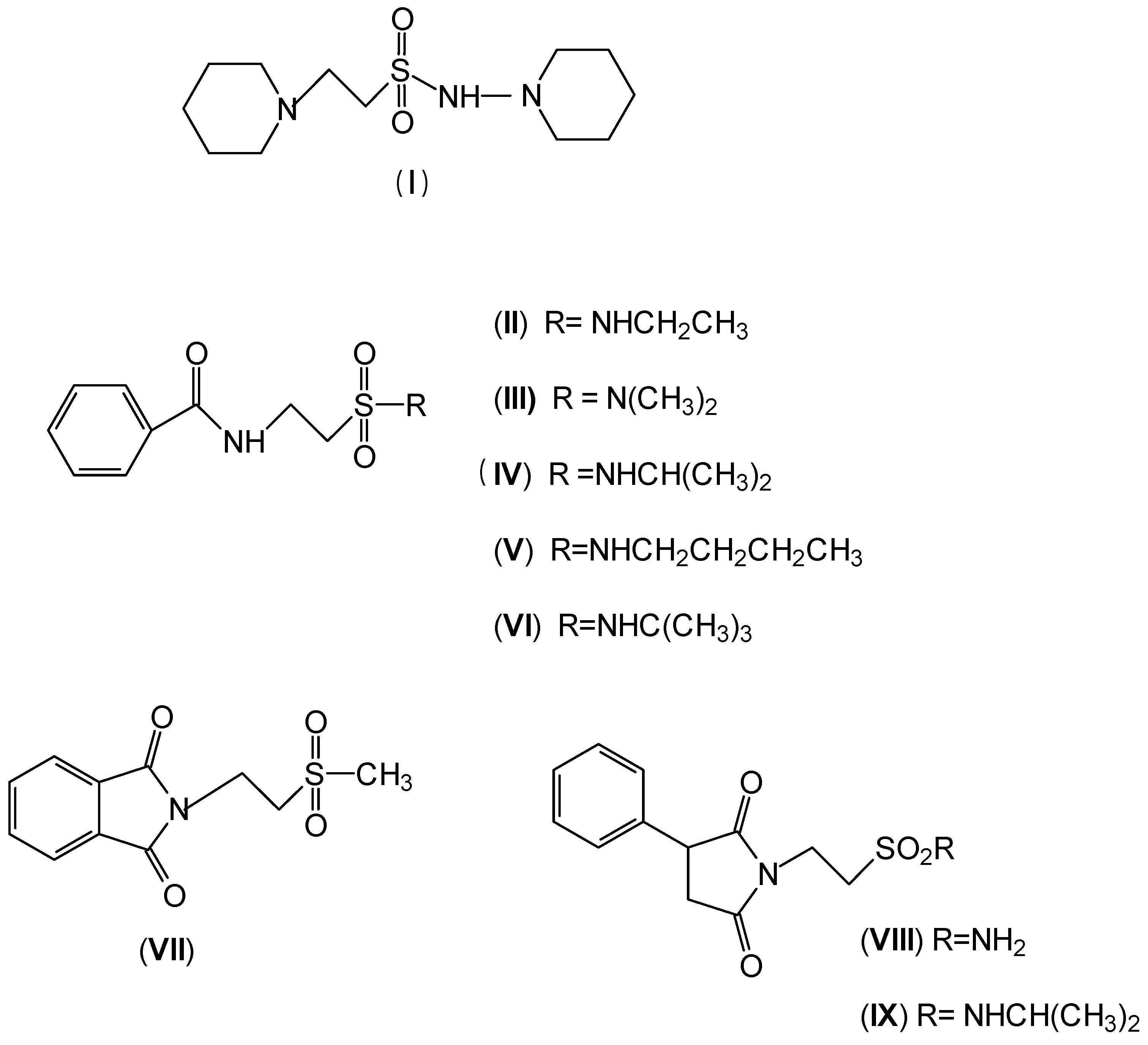
2. Anticonvulsant Taurine Analogues
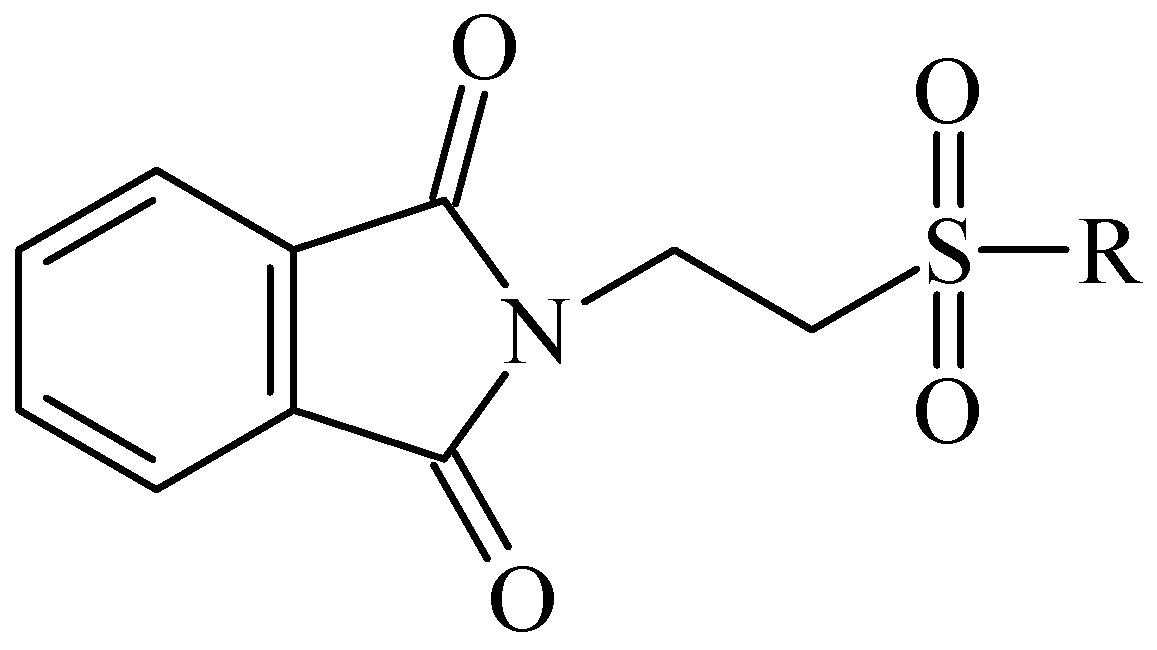
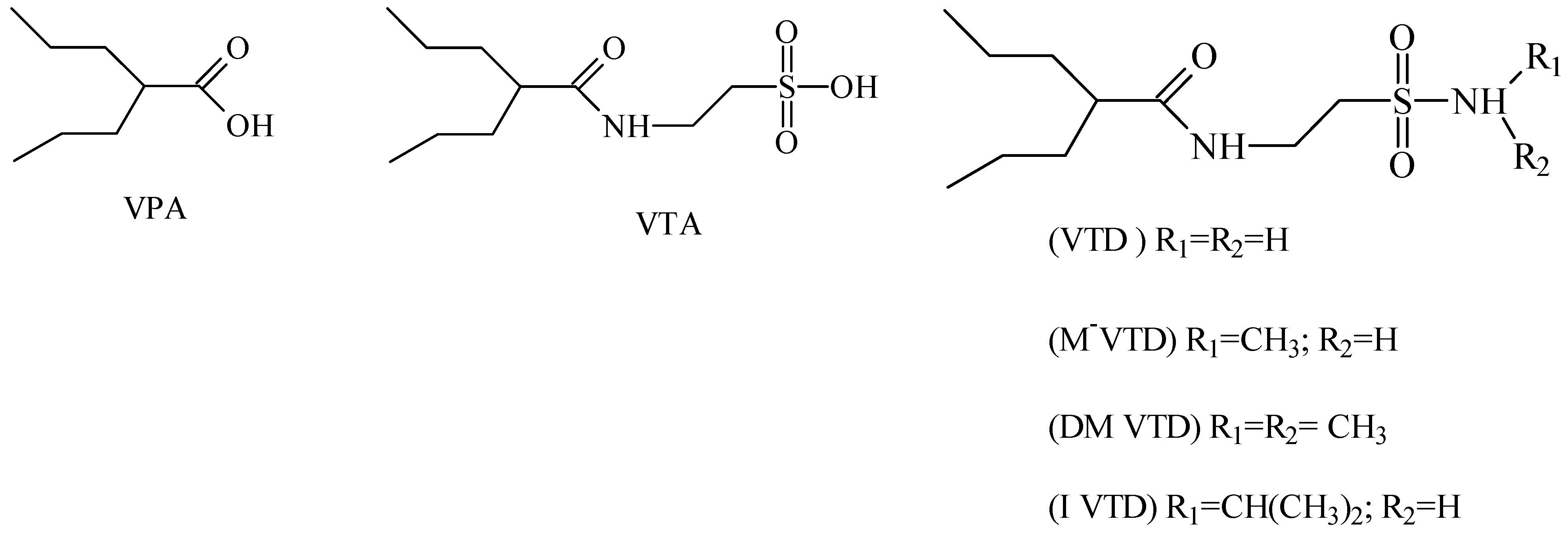

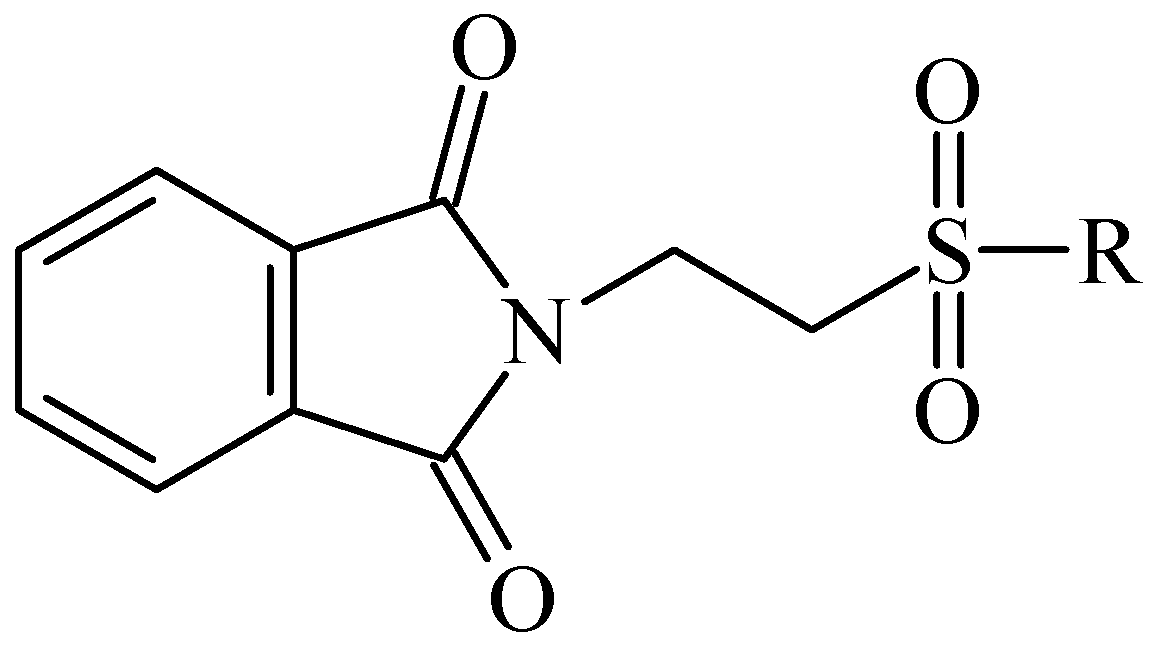{kind=link}
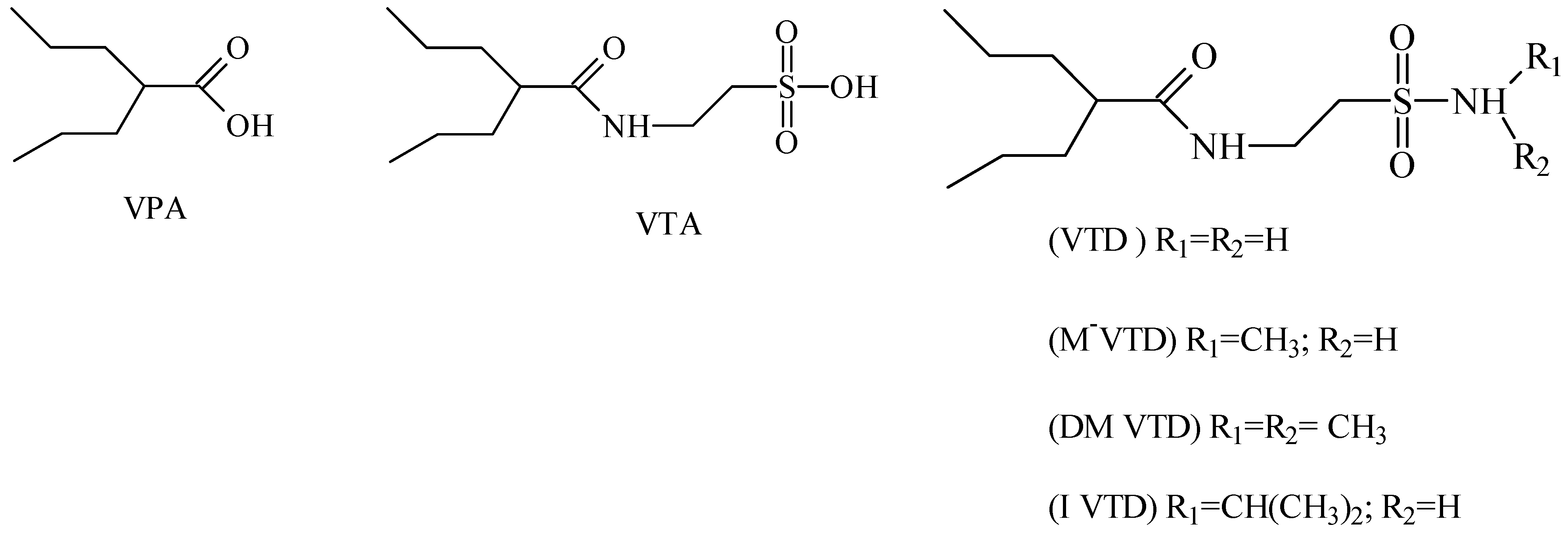{kind=link}
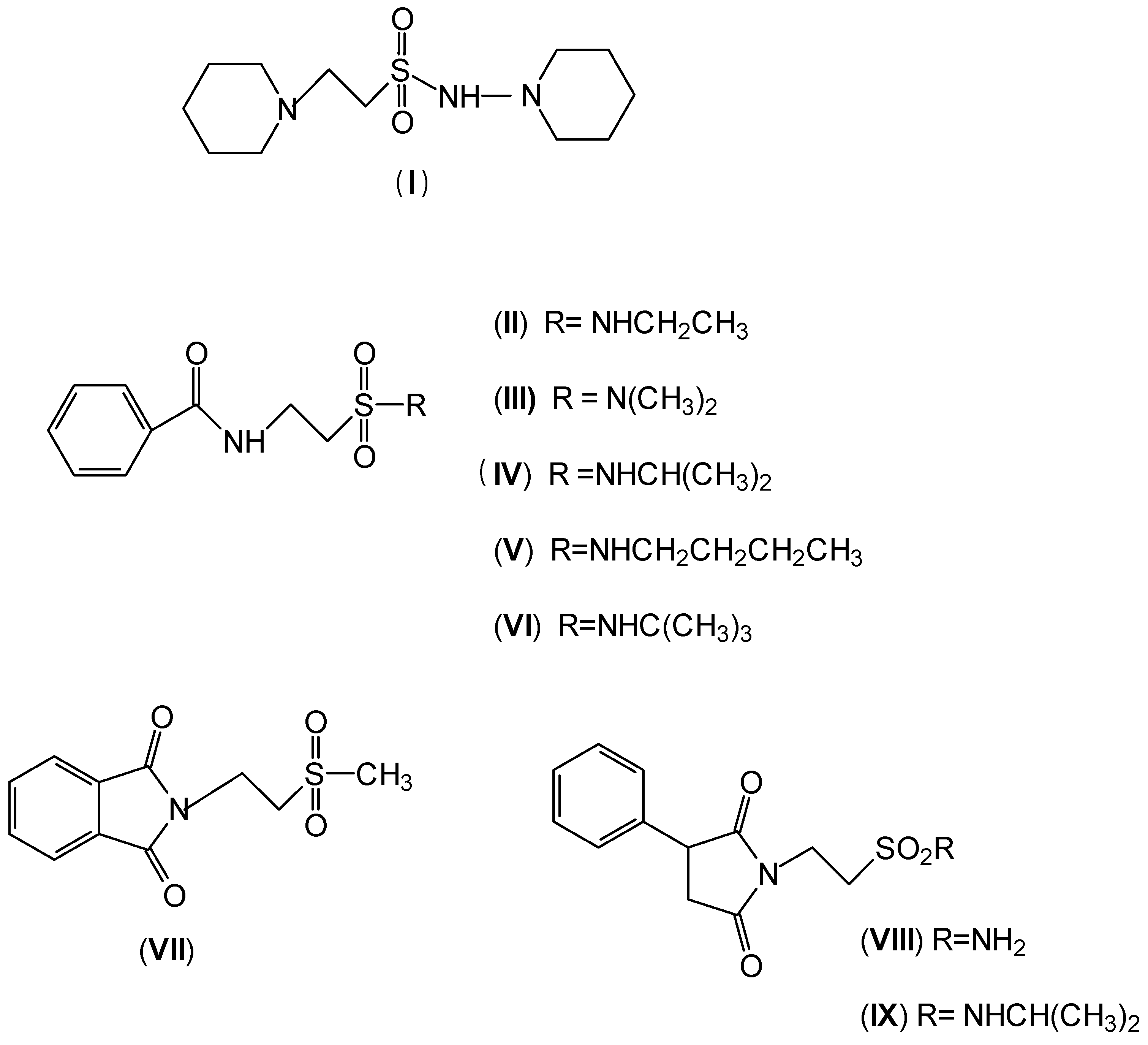{kind=link}
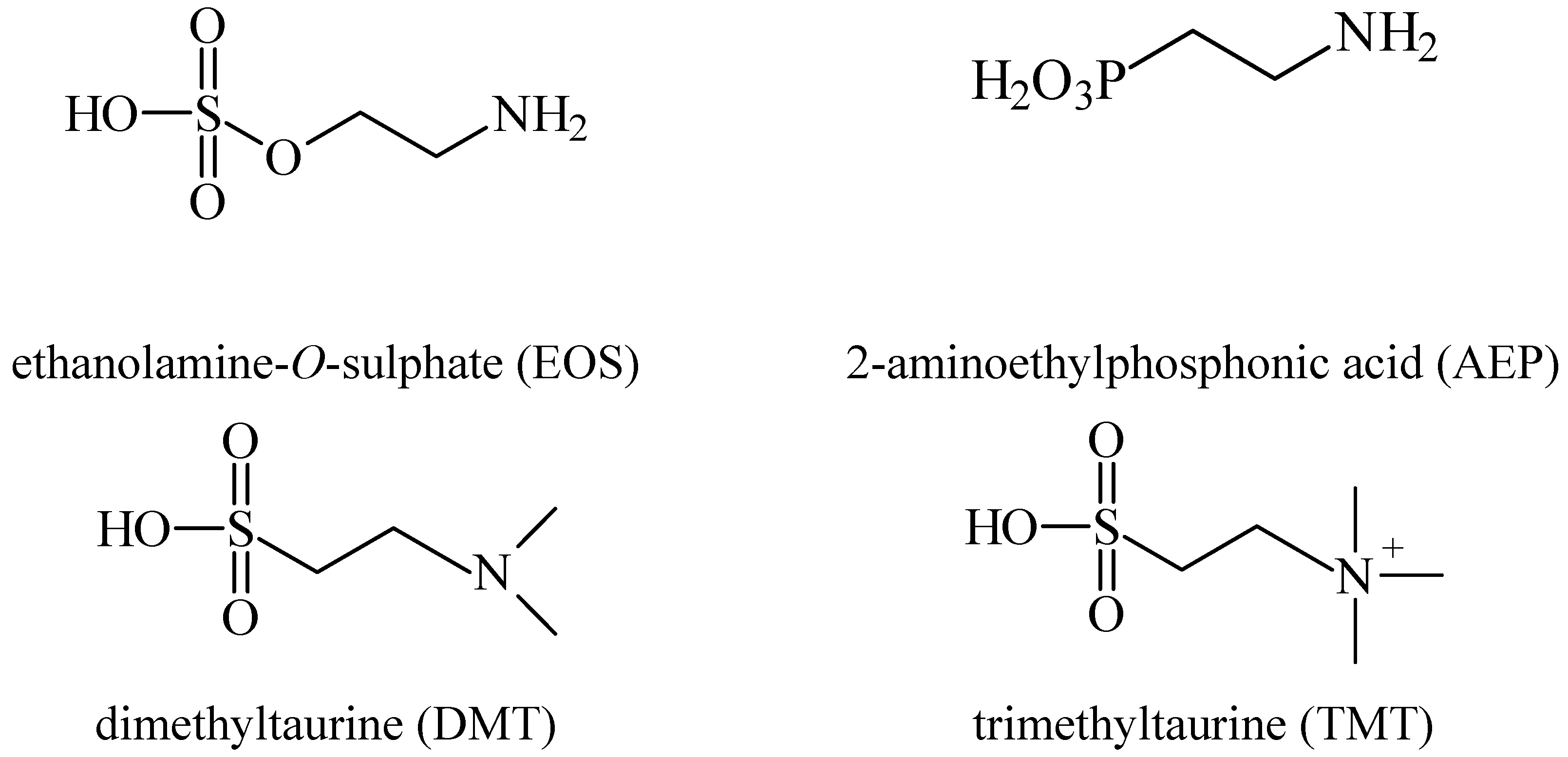{kind=link}
{kind=link}
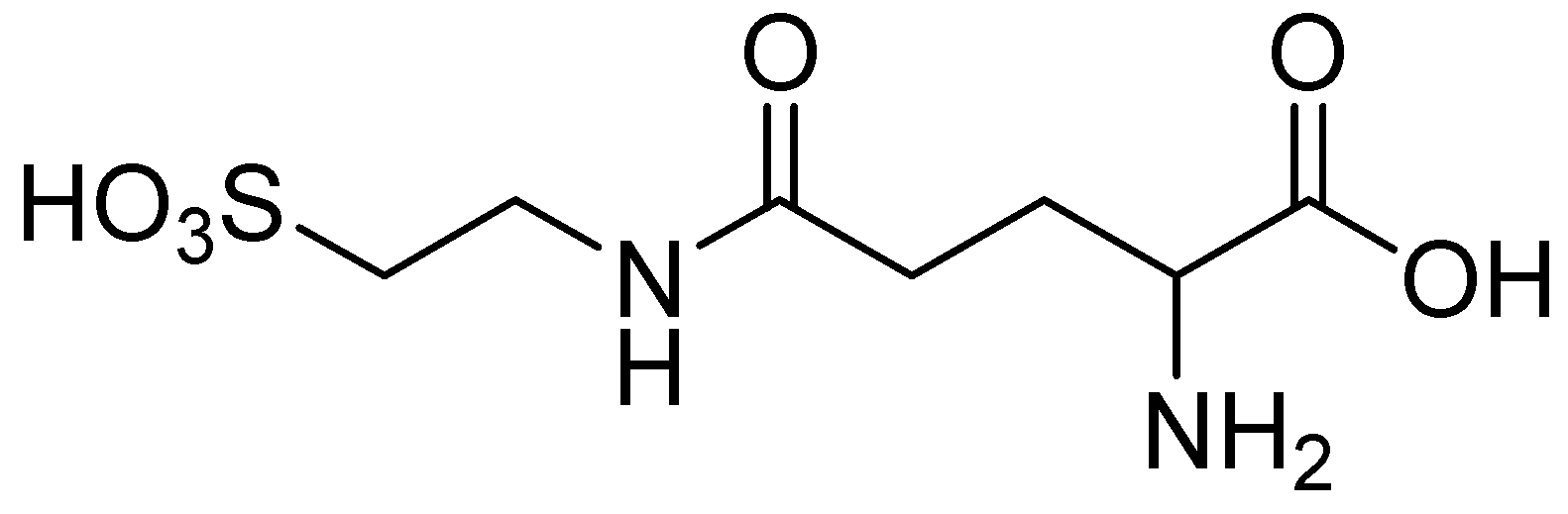{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
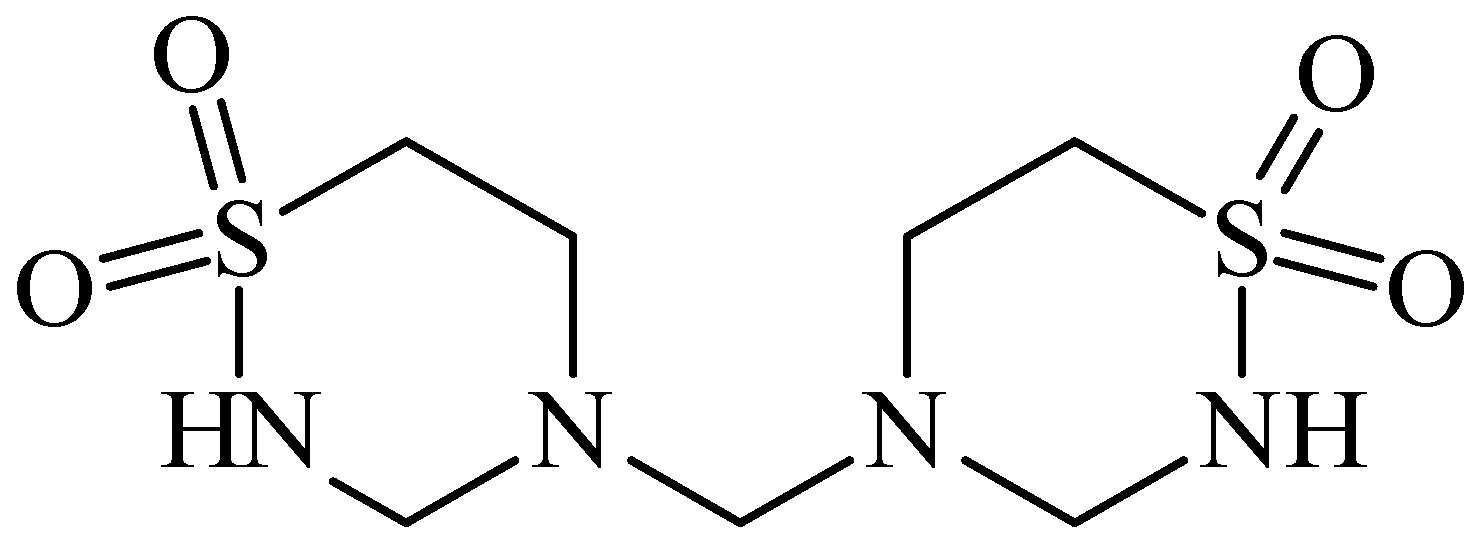{kind=link}
{kind=link}
| Time of the test after administration of the drug | ||
|---|---|---|
| 0.5 h | 4 h | |
| Best activity in MES test | (I) R = 3-NO2 | (III) R = 2-CH3 |
| (II) R = 2-Cl | (IV) R = 2-CH(CH3)2 | |
| (V) R = 4-NO2 | ||
| Higher neurotoxicity | (VI) R = 3-CH3 | (VII) R = 2-CH3 |
3. Alteration in Temperature Activity
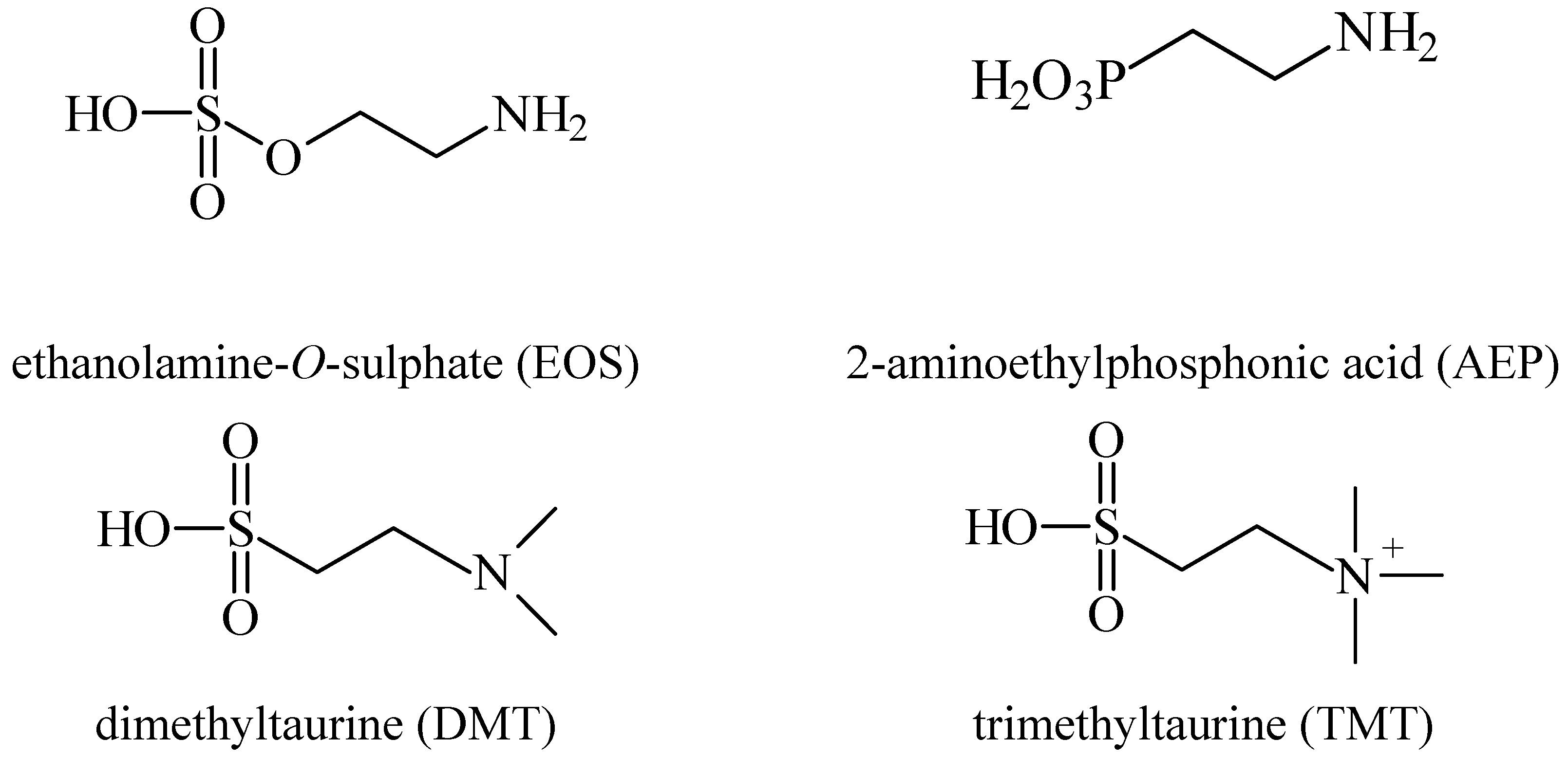

4. Prevention of Alzheimer’s and Neuroprotection
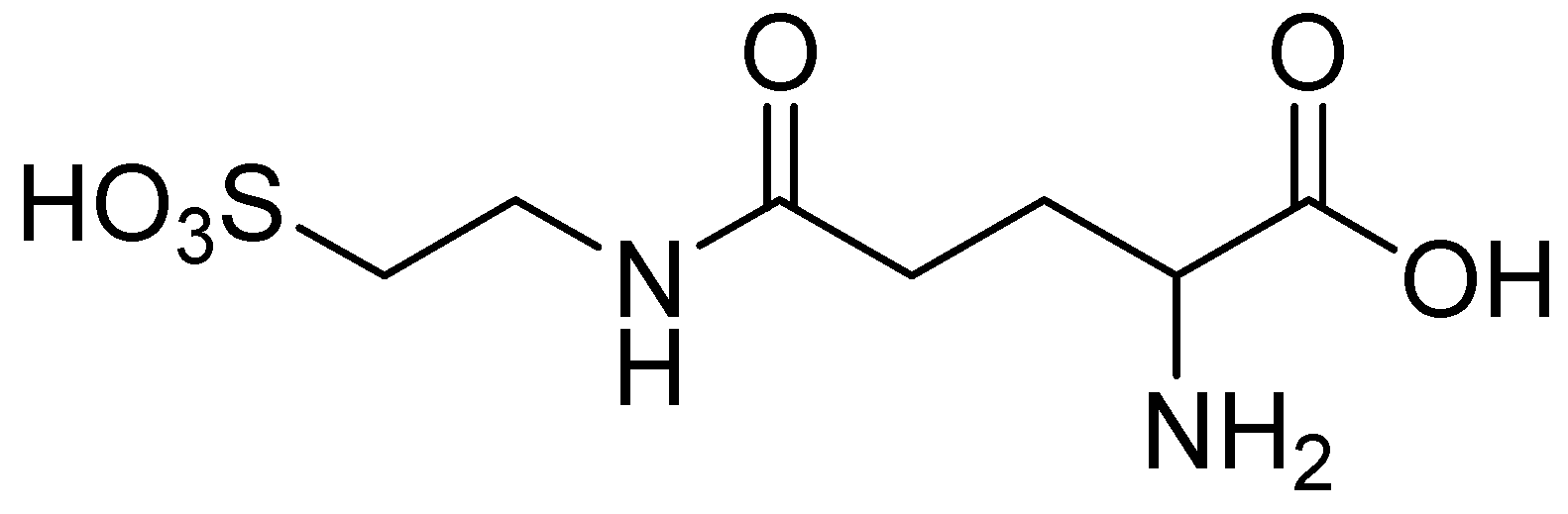

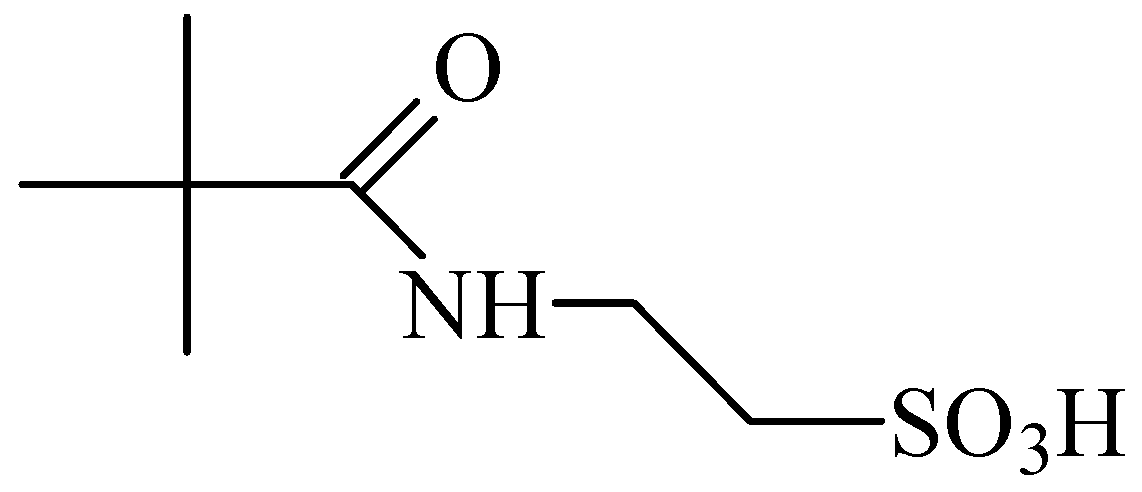
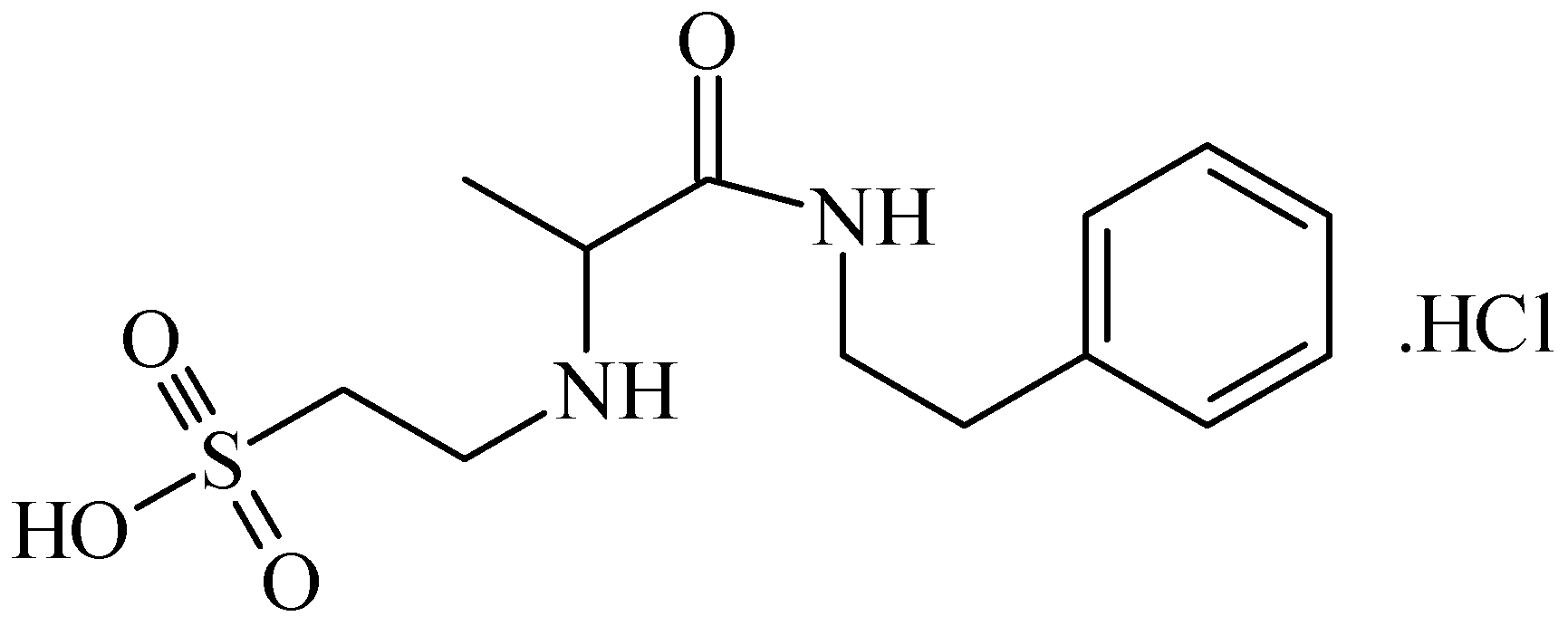
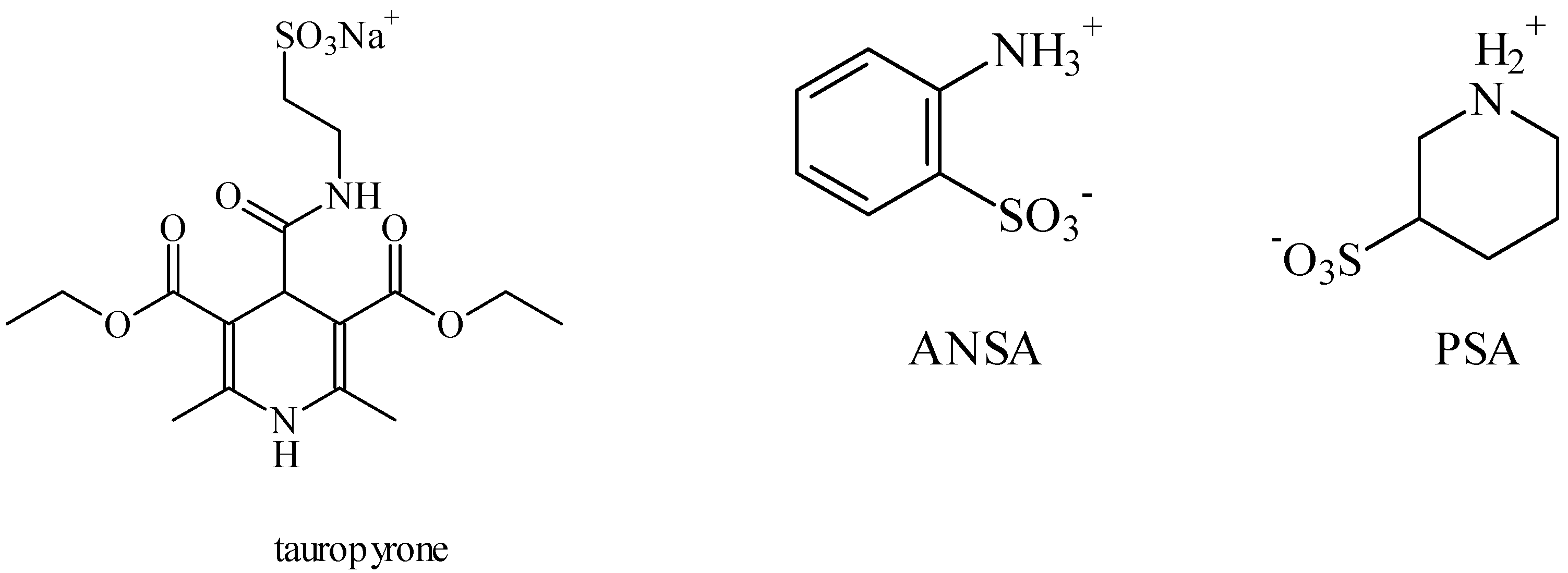
5. Anti-Alcohol Activity

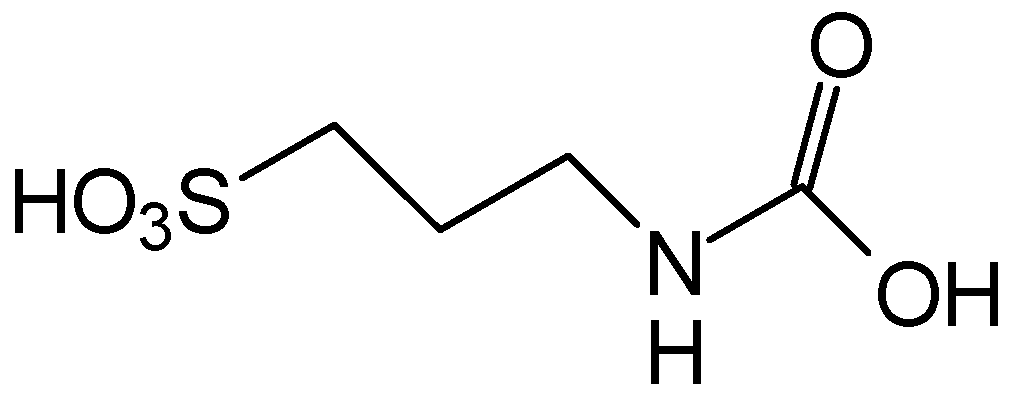
6. Retina Protector
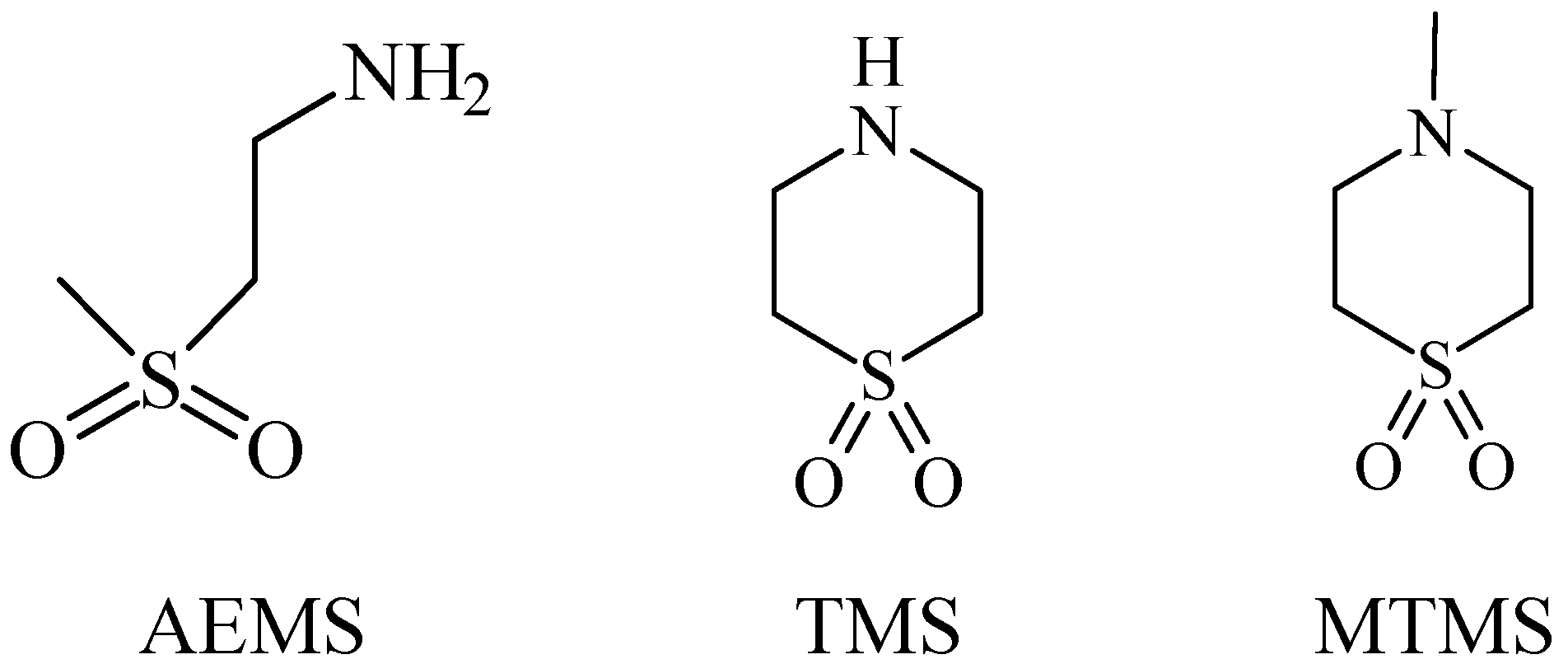
7. Anticancer
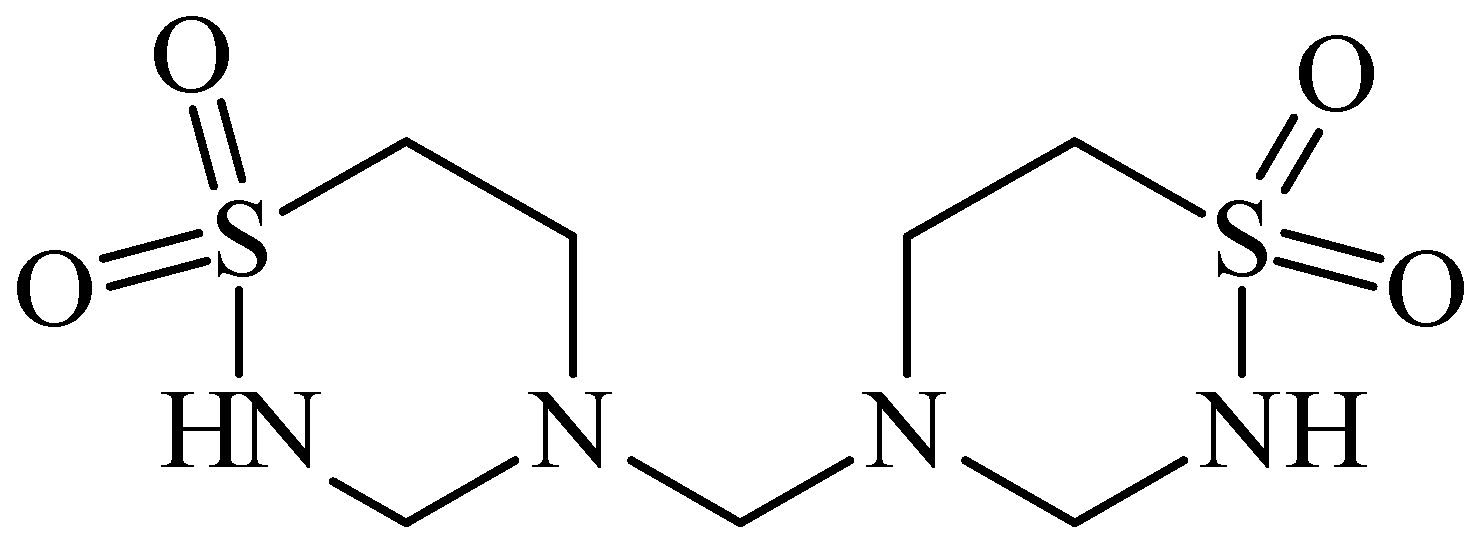
8. CLogP of the Compounds
| Chemical Structures | Clog P | Pharmacological activities | References |
|---|---|---|---|
 | −4.7936 | Physiological aminoacid taurine osmorregulator, neuromodulator, neuroprotector | |
 | 1.0880 | Anticonvulsant | [35] |
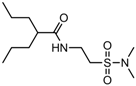 | 2.0546 | Anticonvulsant | [35] |
 | 2.5066 | Anticonvulsant | [35] |
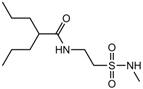 | 1.6686 | Anticonvulsant | [35] |
 | 2.5711 | Anticonvulsant | [33] |
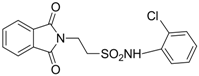 | 2.7571 | Anticonvulsant | [33] |
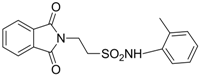 | 2.3471 | Anticonvulsant | [33] |
 | 3.0251 | Anticonvulsant | [33] |
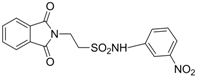 | 2.5711 | Anticonvulsant. | [33] |
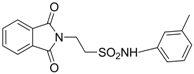 | 2.9071 | Anticonvulsant | [33] |
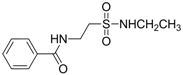 | 0.9486 | Anticonvulsant | [37] |
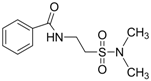 | 0.8056 | Anticonvulsant | [37] |
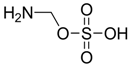 | −3.6202 | Hyperthermia by GABA antogonism | [38] |
 | −3.8958 | Hyperthermia by GABA antogonism | [38] |
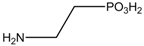 | −3.9796 | Hyperthermia by GABA antogonism | [38] |
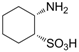 | −3.3916 | Hyperthermia by GABA antogonism | [39] |
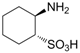 | −3.3916 | Hyperthermia by GABA antogonism | [39] |
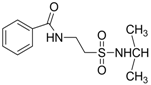 | 1.2576 | Anticonvulsant | [37] |
 | 2,0066 | Anticonvulsant | [37] |
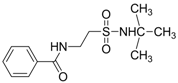 | 1.6566 | Anticonvulsant | [37] |
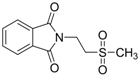 | 0.2834 | Anticonvulsant | [37] |
 | 0.8374 | Anticonvulsant | [37] |
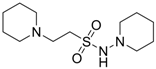 | 2.1490 | Anticonvulsant | [37] |
 | −4.7068 | Alzheimer’s prevention and neuroprotection | [41,42] |
 | −4.4994 | Alzheimer’s prevention and neuroprotection | [42,43,44] |
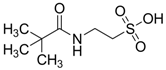 | −1.3964 | Alzheimer’s prevention and neuroprotection | [44,45,46] |
 | −2.8288 | Alzheimer’s prevention and neuroprotection | [47,48,49,50] |
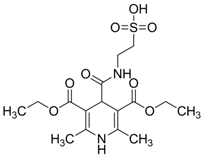 | 0.5202 | Alzheimer’s and Parkinson’s disease prevention and neuroprotection | [51,52] |
 | −1.8720 | Alzheimer’s and Parkinson’s disease prevention and neuroprotection | [51,52] |
 | −3.3915 | Alzheimer’s and Parkinson’s disease prevention and neuroprotection | [51,52] |
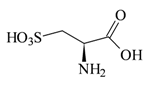 | −4.471 | Anti-alcohol activity | [53,54] |
 | −0.7994 | Anti-alcohol activity. | [53,54] |
 | −3.0638 | Anti-alcohol activity | [55] |
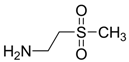 | −1.6418 | Retina protector | [14,56] |
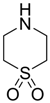 | −1.3740 | Retina protector | [14,56] |
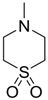 | −0.7990 | Retina protector | [14,56] |
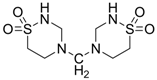 | −0.0866 | Anticancer | [57,58,59,60] |
9. Conclusions
Acknowledgments
Conflict of Interest
References
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Albrecht, J.; Schousboe, A. Taurine interaction with neurotransmitter receptors in the CNS: An update. Neurochem Res. 2005, 30, 1615–1621. [Google Scholar] [CrossRef]
- Wu, J.Y.; Prentice, H. Role of taurine in the central nervous system. J. Biomed. Sci. 2010, 17, S1. [Google Scholar] [CrossRef]
- Bittner, S.; Win, T.; Gupta, R. γ-L-glutamyltaurine. Amino Acids 2005, 28, 343–356. [Google Scholar] [CrossRef]
- Ito, T.; Schaffer, S.W.; Azuma, J. The potential usefulness of taurine on diabetes mellitus and its complications. Amino Acids 2012, 42, 1529–1539. [Google Scholar]
- De la Puerta, C.; Arrieta, F.J.; Balsa, J.A.; Botella-Carretero, J.I.; Zamarrón, I.; Vázquez, C. Taurine and glucose metabolism: A review. Nutr. Hosp. 2010, 25, 910–919. [Google Scholar]
- Moloney, M.A.; Casey, R.G.; O'Donnell, D.H.; Fitzgerald, P.; Thompson, C.; Bouchier-Hayes, D.J. Two weeks taurine supplementation reverses endothelial dysfunction in young male type 1 diabetics. Diab. Vasc. Dis. Res. 2010, 7, 300–310. [Google Scholar] [CrossRef]
- Engelborghs, S.; Marescau, B.; De Deyn, P.P. Amino acids and biogenic amines in cerebrospinal fluid of patients with Parkinson’s disease. Neurochem. Res. 2003, 28, 1145–1150. [Google Scholar] [CrossRef]
- Pomara, N.; Singh, R.; Deptula, D.; Chou, J.C.; Schwartz, M.B.; Le Witt, P.A. Glutamate and other CSF amino acids in Alzheimer's disease. Am. J. Psychiatry 1992, 149, 251–254. [Google Scholar]
- Santa-María, I.; Hernández, F.; Moreno, F.J.; Avila, J. Taurine, an inducer for tau polymerization and a weak inhibitor for amyloid-beta-peptide aggregation. Neurosci. Lett. 2007, 429, 91–94. [Google Scholar] [CrossRef]
- Abebe, W.; Mozaffari, M.S. Role of taurine in the vasculature: An overview of experimental and human studies. Am. J. Cardiovasc. Dis. 2011, 1, 293–311. [Google Scholar]
- Tappia, P.S.; Thliveris, J.; Xu, Y.J.; Aroutiounova, N.; Dhalla, N.S. Effects of amino acid supplementation on myocardial cell damage and cardiac function in diabetes. Exp. Clin. Cardiol. 2011, 16, e17–e22. [Google Scholar]
- Zulli, A. Taurine in cardiovascular disease. Curr. Opin. Clin. Nutr. Metab. Care. 2011, 14, 57–60. [Google Scholar] [CrossRef]
- Gaucher, D.; Arnault, E.; Husson, Z.; Froger, N.; Dubus, E.; Gondouin, P.; Dherbécourt, D.; Degardin, J.; Simonutti, M.; Fouquet, S.; et al. Taurine deficiency damages retinal neurones: Cone photoreceptors and retinal ganglion cells. Amino Acids 2012, 43, 1979–1993. [Google Scholar] [CrossRef]
- Gupta, R.C.; Bittner, S. Taurine analogues: A new class of therapeutics: Retrospect and prospects. Curr. Med. Chem. 2005, 12, 2021–2039. [Google Scholar] [CrossRef]
- Chan-Palay, V.; Lin, C.T.; Palay, S.; Yamamoto, M.; Wu, J.Y. Taurine in the mammalian cerebellum: Demonstration by autoradiography with [3H] taurine and immunocytochemistry with antibodies against the taurine-synthesizing enzyme, cysteine-sulfinic acid decarboxylase. Proc. Natl. Acad. Sci. U.S.A. 1982, 79, 2695–2699. [Google Scholar]
- Magnusson, K.R.; Clements, J.R.; Wu, J.Y.; Beitz, A.J. Colocalization of taurine- and cysteine sulfinic acid decarboxylase-like immunoreactivity in the hippocampus of the rat. Synapse 1989, 4, 55–69. [Google Scholar] [CrossRef]
- Zukin, S.R.; Young, A.B.; Snyder, S.H. Gamma-aminobutyric acid binding to receptor sites in the rat central nervous system. Proc. Natl. Acad. Sci. U.S.A. 1974, 71, 4802–4807. [Google Scholar] [CrossRef]
- Okamoto, K.; Kimura, H.; Sakai, Y. Taurine-induced increase of the Cl-conductance of cerebellar Purkinje cell dendrites in vitro. Brain Res. 1983, 259, 319–323. [Google Scholar] [CrossRef]
- Taber, T.C.; Lin, C.T.; Song, G.X.; Thalman, R.H.; Wu, J.Y. Taurine in the rat hippocampus-localization and postsynaptic action. Brain Res. 1986, 386, 113–121. [Google Scholar] [CrossRef]
- Wu, J.Y.; Tang, X.W.; Tsai, W.H. Taurine receptor: Kinetic analysis and pharmacological studies. Adv. Exp. Med. Biol. 1992, 315, 263–268. [Google Scholar]
- Amoreaux, W.J.; Marsillo, A.; El Idrissi, A. Pharmacological characterization of GABAA receptors in taurine-fed mice. J. Biomed. Sci. 2010, 17, S14. [Google Scholar] [CrossRef]
- El Idrissi, A.; Trenkner, E. Taurine as modulator of excitatory and inhibitory neurotransmission. Neurochem. Res. 2004, 29, 189–197. [Google Scholar] [CrossRef]
- Saransaari, S.; Oja, S.S. Taurine release in the developing and adult mouse hippocampus: Involvement of cyclic guanosine monophosphate. Neurochem. Res. 2002, 27, 15–20. [Google Scholar] [CrossRef]
- Zielińska, M.; Hilgier, W.; Law, R.O.; Goryński, P.; Albrecht, J. Effects of ammonia in vitro on endogenous taurine efflux and cell volume in rat cerebrocortical minislices: Influence of inhibitors of volume-sensitive amino acid transport. Neuroscience 1999, 91, 631–638. [Google Scholar] [CrossRef]
- Louzada, P.R.; Lima, A.C.P.; Mendonca-Silva, D.L.; Noel, F.; Mello, F.G.; Ferreira, S.T. Taurine prevents the neurotoxicity of beta-amyloid and glutamate receptor agonists: Activation of GABA receptors and possible implications for Alzheimer’s disease and other neurological disorders. FASEB J. 2004, 18, 511–518. [Google Scholar] [CrossRef]
- Hilgier, W.; Anderzhanova, E.; Oja, S.S.; Saransaari, P.; Albrecht, J. Taurine reduces ammonia- and N-methyl-Daspartate-induced accumulation of cyclic GMP and hydroxyl radicals in microdialysates of the rat striatum. Eur. J. Pharmacol. 2003, 468, 21–25. [Google Scholar] [CrossRef]
- Albrecht, J.; Wegrzynowicz, M. Endogenous neuro-protectants in ammonia toxicity in the central nervous system: Facts and hypotheses. Metab. Brain Dis. 2005, 20, 253–263. [Google Scholar] [CrossRef]
- Kang, Y.S. The effect of oxidative stress on the transport of taurine in an in vitro model of the blood-brain barrier. Adv. Exp. Med. Biol. 2006, 583, 291–298. [Google Scholar] [CrossRef]
- Kang, Y.S. Taurine transport mechanism through the blood brain barrier in spontaneously hypertensive rats. Adv. Exp. Med. Biol. 2000, 483, 321–324. [Google Scholar] [CrossRef]
- Oja, S.S.; Saransaari, P. Release of endogenous amino acids from the striatum from developing and adult mice inischemia. Neurochem. Res. 2011, 36, 1444–1451. [Google Scholar] [CrossRef]
- Vitreshchak, T.V.; Poleshchuk, V.V.; Piradov, M.A. Plasma levels of mediator amino acids in patients with Parkinson disease. Biomed. Khim. 2004, 50, 92–99. [Google Scholar]
- Lindén, I.B.; Gothóni, G.; Kontro, P.; Oja, S.S. Anticonvulsant activity of 2-phthalimidoethane sulphonamides: New derivatives of taurine. Neurochem. Int. 1983, 5, 319–324. [Google Scholar] [CrossRef]
- Koivisto, K.; Sivenius, J.; Keränen, T.; Partanen, J.; Riekkinen, P.; Gothoni, G.; Tokola, O.; Neuvonen, P.J. Clinical trial with an experimental taurine derivative, taltrimide, in epileptic patients. Epilepsia 1986, 27, 87–90. [Google Scholar] [CrossRef]
- Isoherranen, N.; Yagen, B.; Spiegelstein, O.; Finnell, R.H.; Merriweather, M.; Woodhead, J.H.; Wlodarczyk, B.; White, H.S.; Bialer, M. Anticonvulsant activity, teratogenicity and pharmacokinetics of novel valproyltaurinamide derivatives in mice. Br. J. Pharmacol. 2003, 139, 755–764. [Google Scholar] [CrossRef]
- Akgul, O.; Kilic, F.S.; Erol, K.; Pabuccuoglu, V. Synthesis and anticonvulsant activity of some N-phenyl-2-phtalimidoethanesulfonamide derivatives. Arch. Pharm. (Weinheim) 2007, 340, 656–660. [Google Scholar] [CrossRef]
- Oja, S.S.; Kontro, P.; Lindén, I.B.; Gothóni, G. Anticonvulsant activity of some 2-aminoethanesulfonic acid (taurine) derivatives. Eur. J. Pharmacol. 1983, 87, 191–198. [Google Scholar] [CrossRef]
- Frosini, M.; Ricci, L.; Saponara, S.; Palmi, M.; Valoti, M.; Sgaragli, G. GABA-mediated effects of some taurine derivatives injected i.c.v. on rabbit rectal temperature and gross motor behavior. Amino Acids 2006, 30, 233–242. [Google Scholar] [CrossRef]
- Frosini, M.; Sesti, C.; Saponara, S.; Donati, A.; Palmi, M.; Valoti, M.; Machetti, F.; Sgaragli, G. Effects of taurine and some structurally related analogues on the central mechanism of thermoregulation: A structure-activity relationship study. Adv. Exp. Med. Biol. 2000, 483, 273–282. [Google Scholar]
- Varga, V.; Janáky, R.; Marnela, K.M.; Saransaari, P.; Oja, S. S. Interactions of gamma-L-glutamyltaurine with excitatory aminoacidergic neurotransmission. Neurochem. Res. 1994, 19, 243–248. [Google Scholar] [CrossRef]
- Tóth, S.; Csaba, G. Gamma-L-glutamyl-taurine (Litoralon) prevents the micronucleus formation induced by mitomycin C. Mutat. Res. 1988, 209, 85–89. [Google Scholar] [CrossRef]
- Morris, F.; Cota, D.J. Homotaurine process. U.S. Patent 3,218,352, 16 November 1965. [Google Scholar]
- Aisen, P.S.; Gauthier, S.; Vellas, B.; Briand, R.; Saumier, D.; Laurin, J.; Garceau, D. Alzhemed: A potential treatment for Alzheimer’s disease. Curr Alzheimer Res. 2007, 4, 473–478. [Google Scholar] [CrossRef]
- Aisen, P.S.; Gauthier, S.; Ferris, S.H.; Saumier, D.; Haine, D.; Garceau, D.; Duong, A.; Suhy, J.; Oh, J.; Lau, W.C.; Sampalis, J. Tramiprosate in mild-to-moderate Alzheimer's disease - a randomized, double-blind, placebo-controlled, multi-centre study (the Alphase Study). Arch. Med. Sci. 2011, 7, 102–111. [Google Scholar]
- Ruotsalainen, M.; Majasaari, M.; Salimaki, J.; Ahtee, L. Locally infused taurine, GABA and homotaurine alter differently the striatal extracellular concentrations of dopamine and its metabolites in rats. Amino Acids 1998, 15, 117–134. [Google Scholar] [CrossRef]
- Salimaki, J.; Scriba, G.; Piepponen, T.P.; Rautolahti, N.; Ahtee, L. The effects of systemically administered taurine and N-pivaloyltaurine on striatal extracellular dopamine and taurine in freely moving rats. Naunyn Schmiedebergs Arch. Pharmacol. 2003, 368, 134–141. [Google Scholar] [CrossRef]
- Sapronov, N.S.; Buĺon, V.V.; Krylova, I.B.; Gavrovskaya, L.K.; Selina, E.N.; Evdokimova, N.R. Cerebroprotective effect of a new taurine derivative during cerebral ischemia. Bull. Exp. Biol. Med. 2006, 141, 44–47. [Google Scholar] [CrossRef]
- Krylova, I.B.; Bulion, V.V.; Gavrovskaya, L.K.; Selina, E.N.; Kuznetzova, N.N.; Sapronov, N.S. Neuroprotective effect of a new taurinamide derivative-taurepar. Adv. Exp. Med. Biol. 2006, 583, 543–550. [Google Scholar] [CrossRef]
- Ward, R.; Cirkovic-Vellichovia, T.; Ledeque, F.; Tirizitis, G.; Dubars, G.; Datla, K.; Dexter, D.; Heushling, P.; Crichton, R. Neuroprotection by taurine and taurine analogues. Adv. Exp. Med. Biol. 2006, 583, 299–306. [Google Scholar]
- Klusa, V.; Klimaviciusa, L.; Duburs, G.; Poikans, J.; Zharkovsky, A. Anti-neurotoxic effects of tauropyrone, a taurine analogue. Adv. Exp. Med. Biol. 2006, 583, 499–508. [Google Scholar] [CrossRef]
- Ricci, L.; Frosini, M.; Gaggelli, N.; Valensin, G.; Machetti, F.; Sgaragli, G.; Valoti, M. Inhibition of rabbit brain 4-aminobutyrate transaminase by some taurine analogues: A kinetic analysis. Biochem. Pharmacol. 2007, 71, 1510–1519. [Google Scholar]
- Ricci, L.; Valoti, M.; Sgaragli, G.; Frosini, M. Taurine-like GABA aminotransferase inhibitors prevent rabbit brain slices against oxygen-glucose deprivation-induced damage. Amino Acids 2012, 42, 2139–2147. [Google Scholar] [CrossRef]
- Olive, M.F. Interactions between taurine and ethanol in the central nervous system. Amino Acids 2002, 23, 345–357. [Google Scholar] [CrossRef]
- Messiha, F.S. Taurine, analogues and ethanol elicited responses. Brain Res. Bull. 1979, 4, 603–607. [Google Scholar] [CrossRef]
- Dzirkale, Z.; Pupure, J.; Rumaks, J.; Svirskis, S.; Vanina, M.; Mezhapuke, R.; Sile, V.; Fernandes, M.A.; Duburs, G.; Klusa, V. Comparative study of taurine and tauropyrone: GABA receptor binding, mothocondrial processes and behaviour. J. Pharm. Pharmacol. 2011, 63, 230–237. [Google Scholar]
- Gupta, R.C. Taurine analogues and taurine transport: Therapeutic advantages. Adv. Exp. Med. Biol. 2006, 583, 449–467. [Google Scholar] [CrossRef]
- Caruso, F.; Darnowski, J.W.; Opazo, C.; Goldberg, A.; Kishore, N.; Agoston, E.S.; Rossi, M. Taurolidineantiadhesive properties on interaction with E. coli; its transformation in biological environment and interaction with bacteria cell wall. PLoS ONE 2010, 5, e8927. [Google Scholar]
- Jacobi, C.A.; Menenakos, C.; Braumann, C. Taurolidine-A new drug with anti-tumor and anti-angiogenic effects. Anticancer Drugs 2005, 16, 917–921. [Google Scholar] [CrossRef]
- Harati, K.; Chromik, A.M.; Bulut, D.; Goertz, O.; Hahn, S.; Hirsch, T.; Klein-Hitpass, L.; Lehnhardt, M.; Uhl, W.; Daigeler, A. TRAIL and taurolidine enhance the anticancer activity of doxorubicin, trabectedin and mafosfamide in HT1080 human fibrosarcoma cells. Anticancer Res. 2012, 32, 2967–2984. [Google Scholar]
- Arlt, M.J.; Walters, D.K.; Banke, I.J.; Steinmann, P.; Puskas, G.J.; Bertz, J.; Rentsch, K.M.; Ehrensperger, F.; Born, W.; Fuchs, B. The antineoplastic antibiotic taurolidine promotes lung and liver metastasis in two syngeneicosteosarcoma mouse models and exhibits severe liver toxicity. Int. J. Cancer. 2012, 131, E804–E812. [Google Scholar]
- ChemBioDraw software, version 12.0; A standard industry software used to draw accurate, chemically-aware structures for use in database queries; this software also calculate chemical physical-chemical properties of the molecules such as Log P; CambridgeSoft Coorporation: Cambridge, MA, USA, 2010.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chung, M.C.; Malatesta, P.; Bosquesi, P.L.; Yamasaki, P.R.; Santos, J.L.d.; Vizioli, E.O. Advances in Drug Design Based on the Amino Acid Approach: Taurine Analogues for the Treatment of CNS Diseases. Pharmaceuticals 2012, 5, 1128-1146. https://doi.org/10.3390/ph5101128
Chung MC, Malatesta P, Bosquesi PL, Yamasaki PR, Santos JLd, Vizioli EO. Advances in Drug Design Based on the Amino Acid Approach: Taurine Analogues for the Treatment of CNS Diseases. Pharmaceuticals. 2012; 5(10):1128-1146. https://doi.org/10.3390/ph5101128
Chicago/Turabian StyleChung, Man Chin, Pedro Malatesta, Priscila Longhin Bosquesi, Paulo Renato Yamasaki, Jean Leandro dos Santos, and Ednir Oliveira Vizioli. 2012. "Advances in Drug Design Based on the Amino Acid Approach: Taurine Analogues for the Treatment of CNS Diseases" Pharmaceuticals 5, no. 10: 1128-1146. https://doi.org/10.3390/ph5101128