Applications of Remote Sensing to Alien Invasive Plant Studies

Abstract
:1. Introduction
2. Moderate Spatial/Spectral Resolution Remote Sensing
3. High Spatial Resolution Remote Sensing
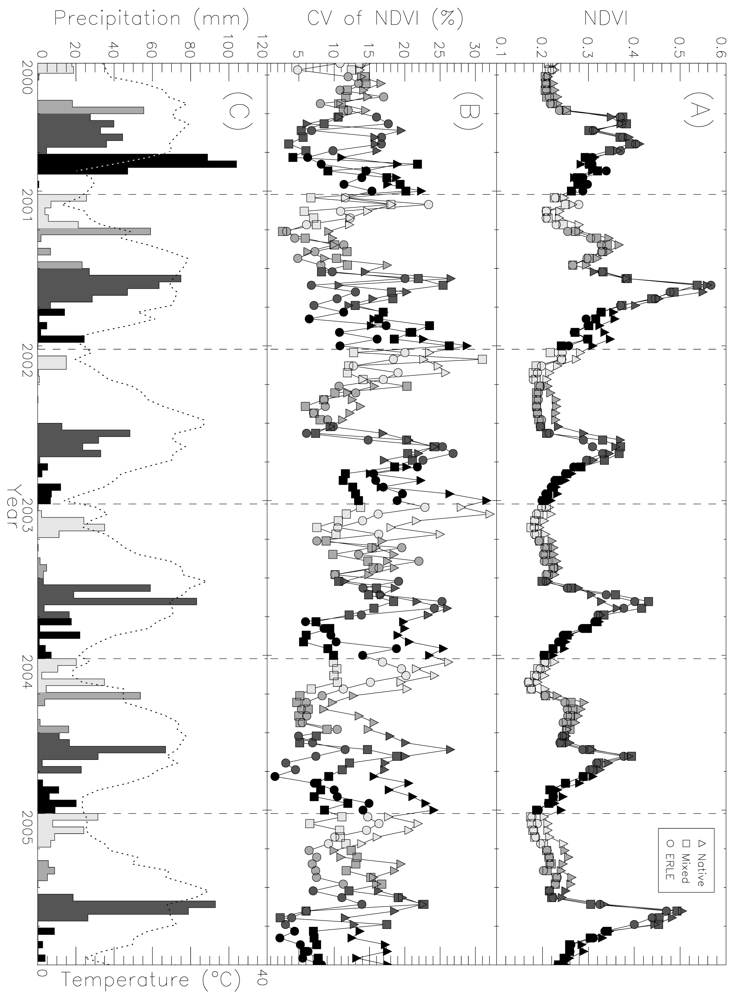
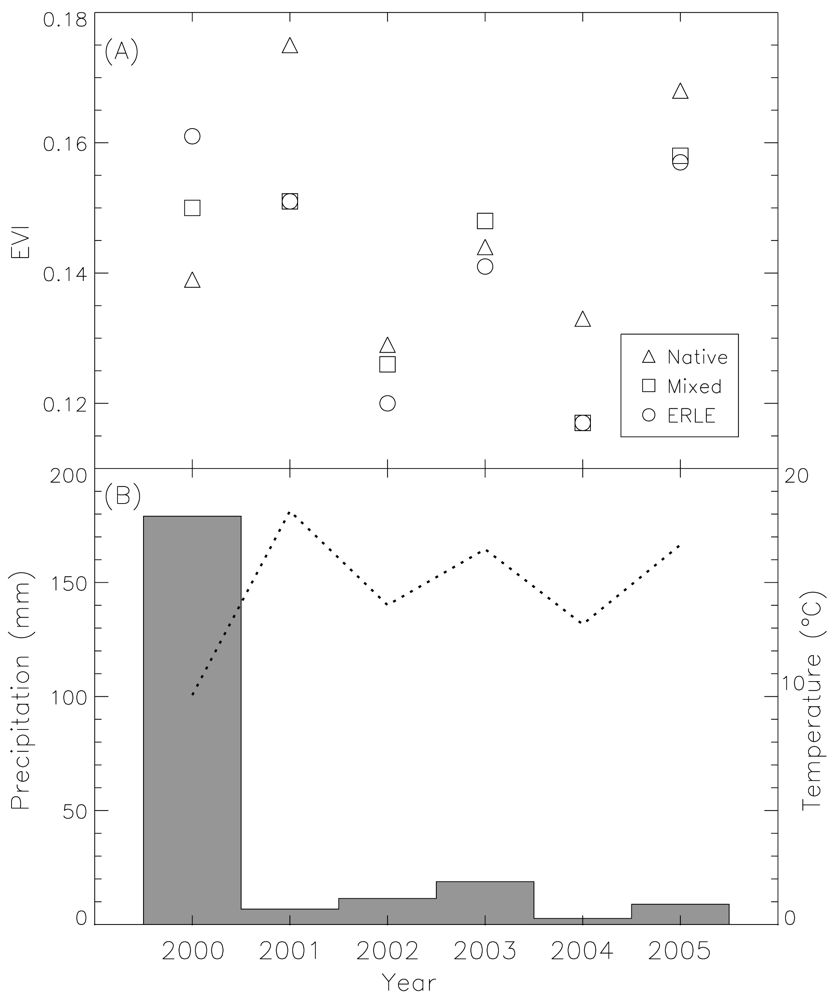
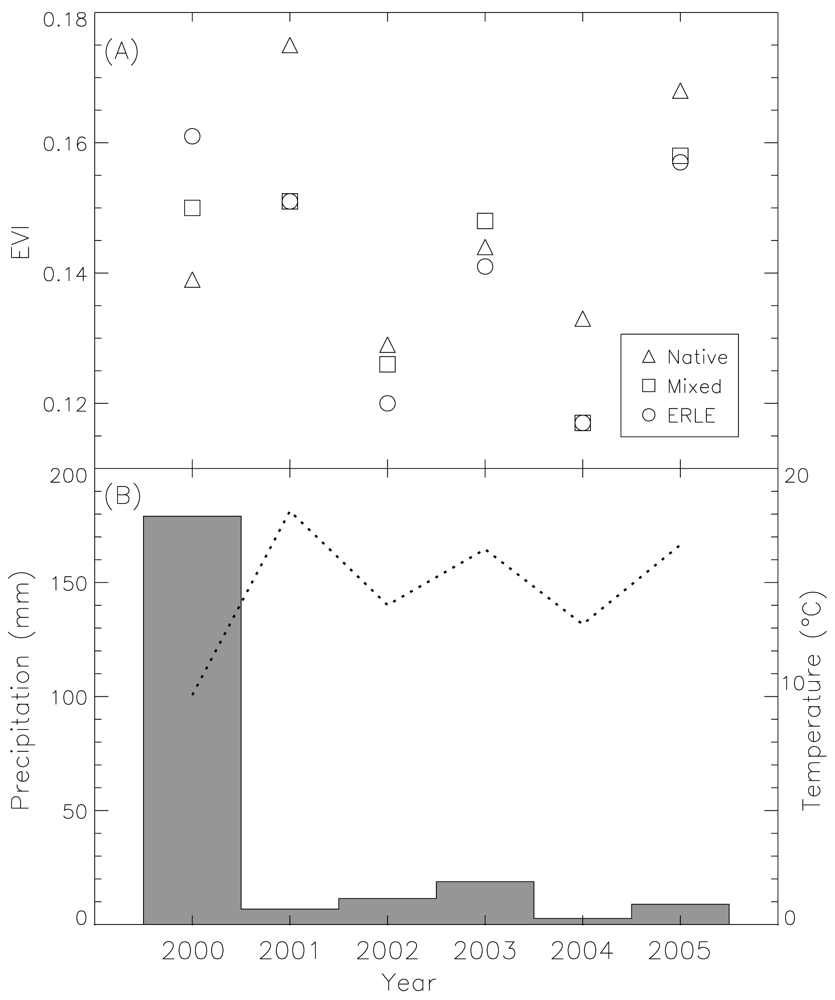
4. High Temporal Resolution Remote Sensing
5. Hyperspectral Remote Sensing
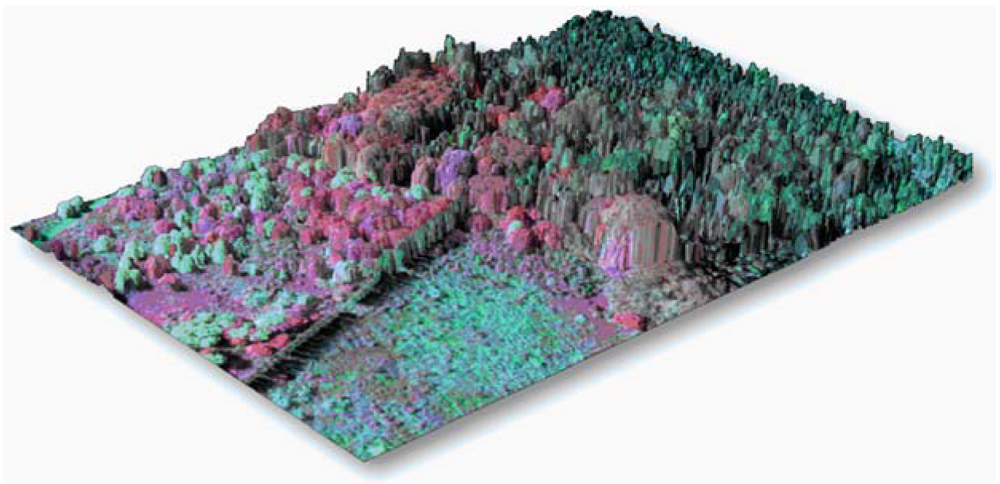
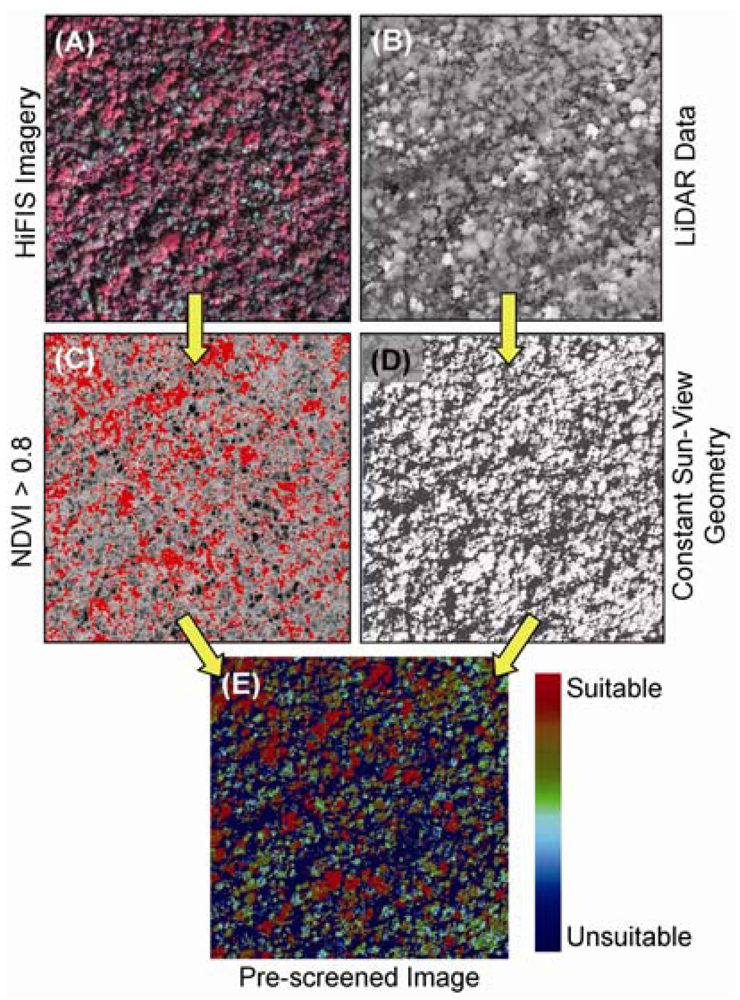
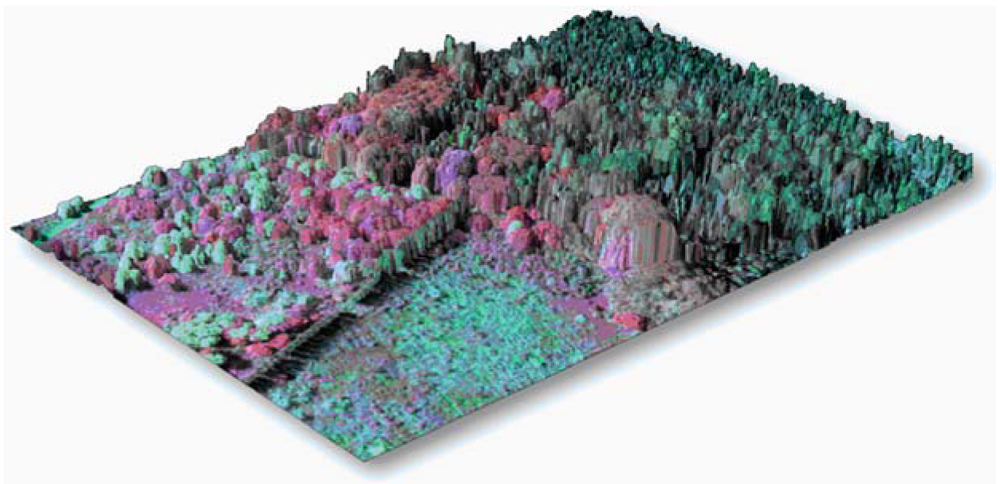
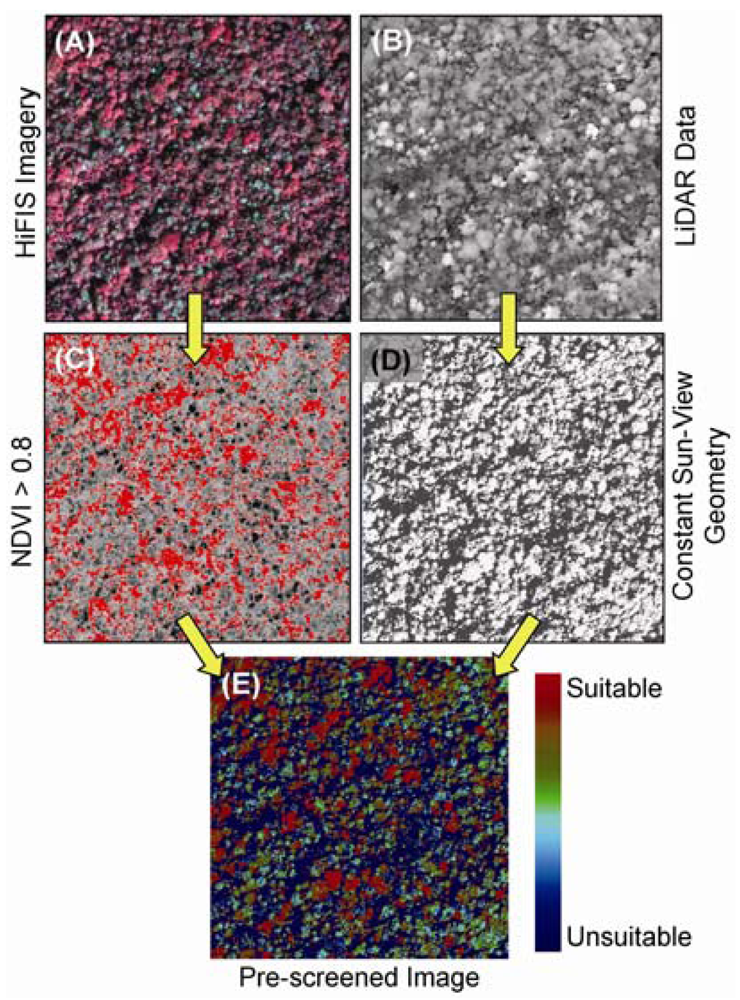
6. LiDAR and Image Fusion
7. The Future of Remote Sensing in Invasive Alien Species Studies
7.1. Limitations of Remote Sensing for Alien Plant Research
7.2. Knowledge Gaps and Research Needs
Acknowledgments
References and Notes
- Parry, M.L.; Canziani, O.F.; Palutikof, J.P.; et al. 2007: Technical Summary. In Climate Change 2007: Impacts, Adaptation and Vulnerability.; Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 23–78. [Google Scholar]
- Beck, K.G.; Zimmerman, K.; Schardt, J.D.; Stone, J.; Lukens, R.R.; Reichard, S.; Randall, J.; Cangelosi, A.A.; Cooper, D.; Thompson, J.P. Invasive species defined in a policy context: Recommendations from the Federal Invasive Species Advisory Committee. Invasive Plant Sci. Manage. 2008, 1, 414–421. [Google Scholar]
- Pimentel, D.; Lach, L.; Zuniga, R.; Morrison, D. Environmental and economic costs of nonindigenous species in the United States. BioScience 2000, 50, 53–65. [Google Scholar]
- Vitousek, P.M. Biological invasions and ecosystem processes: towards an integration of population biology and ecosystem studies. Oikos 1990, 57, 7–13. [Google Scholar]
- D'Antonio, C.M.; Vitousek, P.M. Biological invasions by exotic grasses, the grass/fire cycle, and global change. A. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of earth's ecosystems. Science 1997, 277, 494–499. [Google Scholar]
- Hobbs, R.J. Land-use changes and invasions. In Invasive Species in a Changing World; Mooney, H.A., Hobbs, R.J., Eds.; Island Press: Washington, D.C., USA, 2000; pp. 55–64. [Google Scholar]
- Field, C.B.; Lobell, D.B.; Peters, H.A.; Chiariello, N.R. Feedbacks of terrestrial ecosystems to climate change. A. Rev. Environ. Resour. 2007, 32, 1–29. [Google Scholar]
- Dukes, J.S.; Mooney, H.A. Does global change increase the success of biological invaders? Trend. Ecol. Evol. 1999, 14, 135–139. [Google Scholar]
- Michunas, D.G.; Lauenroth, W.K. Inertia in plant community structure: state changes after cessation of nutrient enrichment stress. Ecol. Appl. 1995, 5, 452–458. [Google Scholar]
- Asner, G.P.; Martin, R.; Carlson, K.; Rascher, U.; Vitousek, P. Vegetation-climate interactions among native and invasive species in Hawaiian rainforest. Ecosystems 2006, 9, 1106–1117. [Google Scholar]
- Vitousek, P.M.; Walker, L.R. Biological invasion by Myrica faya in Hawaii: Plant demography, nitrogen fixation, ecosystem effects. Ecol. Monogr. 1989, 59, 247–265. [Google Scholar]
- Vitousek, P.M.; Walker, L.R.; Whiteaker, L.D.; Mueller-Dombois, D.; Matson, P.A. Biological invasion by Myrica faya alters ecosystem development in Hawaii. Science 1987, 238, 802–804. [Google Scholar]
- Williams, D.G.; Baruch, Z. African grass invasion in the Americas: Ecosystem consequences and the role of ecophysiology. Biol. Invasions. 2000, 2, 123–140. [Google Scholar]
- Cable, D.R. Lehmann lovegrass on the santa rita experimental range, 1937-1968. J. Range Manage. 1971, 24, 17–21. [Google Scholar]
- Cox, J.R.; Martin-R, M.H.; Ibarra-F, F.A.; Fourie, J.H.; Rethman, N.F.G.; Wilcox, D.G. The influence of climate and soils on the distribution of four African grasses. J. Range Manage. 1988, 41, 127–139. [Google Scholar]
- Cox, J.R.; Ruyle, G.B. Influence of climatic and edaphic factors on the distribution of Eragrostis lehmanniana Nees. in Arizona, USA. J. Grassland Soc. S. Afr. 1986, 3, 25–29. [Google Scholar]
- Schussman, H.; Geiger, E.; Mau-Crimmins, T.; Ward, J. Spread and current potential distribution of an alien grass, Eragrostis lehmanniana Nees, in the southwestern USA: comparing historical data and ecological niche models. Divers. Distrib. 2006, 12, 582–592. [Google Scholar]
- Carlson, M.L.; Lapina, I.V.; Shephard, M.; Conn, J.S.; Densmore, R.; Spencer, P.; Heys, J.; Riley, J.; Nielsen, J. Invasiveness Ranking System for Non-native Plants of Alaska; US Forest Service, US Department of Agriculture: Anchorage, AK, USA, 2008. [Google Scholar]
- Schott, J.R. Remote Sensing: The Image Chain Approach; Oxford University Press: New York, NY, USA, 1997. [Google Scholar]
- Asrar, G. Introduction. In Theory and Applications of Optical Remote Sensing; Asrar, G., Ed.; John Wiley & Sons: New York, NY, USA, 1989; p. 1. [Google Scholar]
- Resasco, J.; Hale, A.N.; Henry, M.C.; Gorchov, D.L. Detecting an invasive shrub in a deciduous forest understory using late-fall Landsat sensor imagery. Int. J. Remote Sens. 2007, 28, 3739–3745. [Google Scholar]
- Peterson, E.B. Estimating cover of an invasive grass Bromus tectorum using tobit regression and phenology derived from two dates of Landsat ETM+ data. Int. J. Remote Sens. 2005, 26, 2491–2507. [Google Scholar]
- Groeneveld, D.P.; Watson, R.P. Near-infrared discrimination of leafless saltcedar in wintertime Landsat TM. Int. J. Remote Sens. 2008, 29, 3577–3588. [Google Scholar]
- Asner, G.P. Hyperspectral remote sensing of canopy chemistry, physiology and diversity in tropical rainforests. In Hyperspectral Remote Sensing of Tropical and Subtropical Forests; Kalacska, M., Sanchez-Azofeifa, G.A., Eds.; Taylor and Francis Group: Oxon, UK, 2008; pp. 261–288. [Google Scholar]
- Everitt, J.H.; Anderson, G.L.; Escobar, D.E.; Davis, M.R.; Spencer, N.R.; Andrascik, R.J. Use of remote sensing for detecting and mapping leafy spurge (Euphorbia esula). Weed Technol. 1995, 9, 599–609. [Google Scholar]
- Everitt, J.H.; Escobar, D.E.; Alaniz, M.A.; Davis, M.R.; Richerson, J.V. Using spatial information technologies to map Chinese tamarisk (Tamarix chinensis) infestations. Weed Sci. 1996, 44, 194–201. [Google Scholar]
- Müllerová, J.; Pyšek, P.; Jarošík, V.; Pergl, J. Aerial photographs as a tool for assessing the regional dynamics of the invasive plant species Heracleum mantegazzianum. J. Appl. Ecol. 2005, 42, 1042–1053. [Google Scholar]
- Everitt, J.H.; Escobar, D.E.; Davis, M.R. Reflectance and image characteristics of selected noxious rangeland species. J. Range Manage. 2001, 54, A106–A120. [Google Scholar]
- Fuller, D.O. Remote detection of invasive Melaleuca trees (Melaleuca quinquenervia) in South Florida with multispectral IKONOS imagery. Int. J. Remote Sens. 2005, 26, 1057–1063. [Google Scholar]
- Sanchez-Flores, E.; Rodriguez-Gallegos, H.; Yool, S.R. Plant invasions in dynamic desert landscapes. A field and remote sensing assessment of predictive and change modeling. J. Arid Environ. 2008, 72, 189–206. [Google Scholar]
- Laba, M.; Downs, R.; Smith, S.; Welsh, S.; Neider, C.; White, S.; Richmond, M.; Philpot, W.; Baveye, P. Mapping invasive wetland plants in the Hudson River National Estuarine Research Reserve using QuickBird satellite imagery. Remote Sens. Environ. 2008, 112, 286–300. [Google Scholar]
- Everitt, J.H.; Yang, C.; Deloach, C.J. Remote sensing of giant reed with QuickBird satellite imagery. J. Aquat. Plant Manage. 2005, 43, 81–85. [Google Scholar]
- Bock, C.E.; Bock, J.H.; Jepson, K.L.; Ortega, J.C. Ecological effects of planting African lovegrasses in Arizona. Natl. Geogr. Res. 1986, 2, 456–463. [Google Scholar]
- Fargione, J.; Tilman, D. Niche differences in phenology and rooting depth promote coexistence with a dominant C4 bunchgrass. Oecologia 2005, 143, 598–606. [Google Scholar]
- Justice, C.O.; Townshend, J.R.G.; Holben, B.N.; Tucker, C.J. Analysis of the phenology of global vegetation using meteorological satellite data. Int. J. Remote Sens. 1985, 6, 1271–1318. [Google Scholar]
- Prince, S.D.; Tucker, C.J. Satellite remote sensing of rangelands in Botswana II. NOAA AVHRR and herbaceous vegetation. Int. J. Remote Sens. 1986, 7, 1555–1570. [Google Scholar]
- Rouse, J., Jr.; Haas, R.; Deering, D.; Schell, J.; Harlan, C. Monitoring the Vernal Advancement and Retrogradation (Green Wave Effect) of Natural Vegetation; NASA: Greenbelt, MD, USA, 1974. [Google Scholar]
- Reed, B.C.; Brown, J.F.; VanderZee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar]
- Bradley, B.A.; Mustard, J.F. Identifying land cover variability distinct from land cover change: Cheatgrass in the Great Basin. Remote Sens. Environ. 2005, 94, 204–213. [Google Scholar]
- Morisette, J.T.; Jarnevich, C.S.; Ullah, A.; Cai, W.J.; Pedelty, J.A.; Gentle, J.E.; Stohlgren, T.J.; Schnase, J.L. A tamarisk habitat suitability map for the continental United States. Front. Ecol. Environ. 2006, 4, 11–17. [Google Scholar]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar]
- Friedl, M.A.; McIver, D.K.; Hodges, J.C.F.; Zhang, X.Y.; Muchoney, D.; Strahler, A.H.; Woodcock, C.E.; Gopal, S.; Schneider, A.; Cooper, A.; Baccini, A.; Gao, F.; Schaaf, C. Global land cover mapping from MODIS: algorithms and early results. Remote Sens. Environ. 2002, 83, 287–302. [Google Scholar]
- Anderson, R.P.; Peterson, A.T.; Egbert, S.L. Vegetation-index models predict areas vulnerable to purple loosestrife (lythrum salicaria) invasion in Kansas. Southwest. Nat. 2006, 51, 471–480. [Google Scholar]
- Huang, C.; Geiger, E.; van Leeuwen, W.; Marsh, S. Discrimination of invaded and native species sites in a semi-desert grassland using MODIS multi-temporal data. Int. J. Remote Sens. 2009, 30, 897–917. [Google Scholar]
- Huang, C.; Geiger, E.L. Climate anomalies provide opportunities for large-scale mapping of non-native plant abundance in desert grasslands. Divers. Distrib. 2008, 14, 875–884. [Google Scholar]
- Green, R.O.; Eastwood, M.L.; Sarture, C.M.; Chrien, T.G.; Aronsson, M.; Chippendale, B.J.; Faust, J.A.; Pavri, B.E.; Chovit, C.J.; Solis, M. Imaging spectroscopy and the Airborne Visible/Infrared Imaging Spectrometer (AVIRIS). Remote Sens. Environ. 1998, 65, 227–248. [Google Scholar]
- Guanter, L.; Estellés, V.; Moreno, J. Spectral calibration and atmospheric correction of ultra-fine spectral and spatial resolution remote sensing data. Application to CASI-1500 data. Remote Sens. Environ. 2007, 109, 54–65. [Google Scholar]
- Schlerf, M.; Atzberger, C.; Hill, J. Remote sensing of forest biophysical variables using HyMap imaging spectrometer data. Remote Sens. Environ. 2005, 95, 177–194. [Google Scholar]
- Thenkabail, P.S.; Enclona, E.A.; Ashton, M.S.; Legg, C.; de Dieu, M.J. Hyperion, IKONOS, ALI, and ETM+ sensors in the study of African rainforests. Remote Sens. Environ. 2004, 90, 23–43. [Google Scholar]
- Vane, G.; Goetz, A.F.H. Terrestrial imaging spectroscopy. Remote Sens. Environ. 1988, 24, 1–29. [Google Scholar]
- Underwood, E.; Ustin, S.; DiPietro, D. Mapping nonnative plants using hyperspectral imagery. Remote Sens. Environ. 2003, 86, 150–161. [Google Scholar]
- Parker Williams, A.; Hunt, E.R., Jr. Estimation of leafy spurge cover from hyperspectral imagery using mixture tuned matched filtering. Remote Sens. Environ. 2002, 82, 446–456. [Google Scholar]
- Glenn, N.F.; Mundt, J.T.; Weber, K.T.; Prather, T.S.; Lass, L.W.; Pettingill, J. Hyperspectral data processing for repeat detection of small infestations of leafy spurge. Remote Sens. Environ. 2005, 95, 399–412. [Google Scholar]
- Mundt, J.T.; Glenn, N.F.; Weber, K.T.; Prather, T.S.; Lass, L.W.; Pettingill, J. Discrimination of hoary cress and determination of its detection limits via hyperspectral image processing and accuracy assessment techniques. Remote Sens. Environ. 2005, 96, 509–517. [Google Scholar]
- Andrew, M.E.; Ustin, S.L. The role of environmental context in mapping invasive plants with hyperspectral image data. Remote Sens. Environ. 2008, 112, 4301–4317. [Google Scholar]
- Hestir, E.L.; Khanna, S.; Andrew, M.E.; Santos, M.J.; Viers, J.H.; Greenberg, J.A.; Rajapakse, S.S.; Ustin, S.L. Identification of invasive vegetation using hyperspectral remote sensing in the California Delta ecosystem. Remote Sens. Environ. 2008, 112, 4034–4047. [Google Scholar]
- Miao, X.; Gong, P.; Swope, S.; Pu, R.; Carruthers, R.; Anderson, G.L.; Heaton, J.S.; Tracy, C.R. Estimation of yellow starthistle abundance through CASI-2 hyperspectral imagery using linear spectral mixture models. Remote Sens. Environ. 2006, 101, 329–341. [Google Scholar]
- Asner, G.P.; Vitousek, P.M. Remote analysis of biological invasion and biogeochemical change. Proc. Natl. Acad. Sci. USA 2005, 102, 4383–4386. [Google Scholar]
- Hall, S.J.; Asner, G.P. Biological invasion alters regional nitrogen-oxide emissions from tropical rainforests. Glob. Change Biol. 2007, 13, 2143–2160. [Google Scholar]
- Asner, G.P.; Jones, M.O.; Martin, R.E.; Knapp, D.E.; Hughes, R.F. Remote sensing of native and invasive species in Hawaiian forests. Remote Sens. Environ. 2008, 112, 1912–1926. [Google Scholar]
- Asner, G.P.; Hughes, R.F.; Vitousek, P.M.; Knapp, D.E.; Kennedy-Bowdoin, T.; Boardman, J.; Martin, R.E.; Eastwood, M.; Green, R.O. Invasive plants transform the three-dimensional structure of rain forests. Proc. Natl. Acad. Sci. USA 2008, 105, 4519–4523. [Google Scholar]
- Kruse, F.A. Mineral Mapping with AVIRIS and EO-1 Hyperion. Summaries of the 12th Annual Jet Propulsion Laboratory Airborne Geoscience Workshop, Pasadena, CA, USA; 2003; pp. 149–156. [Google Scholar]
- Lucas, R.; Rowlands, A.; Niemann, O.; Merton, R. Hyperspectral sensors and applications. In Advanced Image Processing Techniques for Remotely Sensed Hyperspectral Data; Varshney, P.K., Arora, M.K., Eds.; Springer: Berlin, Germany, 2004; pp. 11–50. [Google Scholar]
- Pengra, B.W.; Johnston, C.A.; Loveland, T.R. Mapping an invasive plant, Phragmites australis, in coastal wetlands using the EO-1 Hyperion hyperspectral sensor. Remote Sens. Environ. 2007, 108, 74–81. [Google Scholar]
- Tsai, F.; Lin, E.-K.; Yoshino, K. Spectrally segmented Principal Component Analysis of hyperspectral imagery for mapping invasive plant species. Int. J. Remote Sens. 2007, 28, 1023–1039. [Google Scholar]
- Walsh, S.J.; McCleary, A.L.; Mena, C.F.; Shao, Y.; Tuttle, J.P.; Gonzáez, A.; Atkinson, R. QuickBird and Hyperion data analysis of an invasive plant species in the Galapagos Islands of Ecuador: Implications for control and land use management. Remote Sens. Environ. 2008, 112, 1927–1941. [Google Scholar]
- Bachman, C.G. Laser Radar Systems and Techniques; Artech House: Norwood, MA, USA, 1979. [Google Scholar]
- Lefsky, M.A.; Cohen, W.B.; Parker, G.G.; Harding, D.J. Lidar remote sensing for ecosystem studies. BioScience 2002, 52, 19–30. [Google Scholar]
- Rosso, P.H.; Ustin, S.L.; Hastings, A. Use of lidar to study changes associated with Spartina invasion in San Francisco Bay marshes. Remote Sens. Environ. 2006, 100, 295–306. [Google Scholar]
- Boelman, N.T.; Asner, G.P.; Hart, P.J.; Martin, R.E. Multi-trophic invasion resistance in Hawaii: Bioacoustics, field surveys, and airborne remote sensing. Ecol. Appl. 2007, 17, 2137–2144. [Google Scholar]
- Mundt, J.; Streutker, D.; Glenn, N. Mapping sagebrush distribution via fusion of hyperspectral and LiDAR classifications. Photogramm. Eng. Remote Sens. 2006, 72, 47–54. [Google Scholar]
- Andrew, M.E.; Ustin, S.L. Habitat suitability modelling of an invasive plant with advanced remote sensing data. Divers. Distrib. 2009, 15, 627–640. [Google Scholar]
- Asner, G.P.; Knapp, D.E.; Jones, M.O.; Kennedy-Bowdoin, T.; Martin, R.E.; Boardman, J.; Field, C.B. Carnegie Airborne Observatory: in-flight fusion of hyperspectral imaging and waveform light detection and ranging (wLiDAR) for three-dimensional studies of ecosystems. J. Appl. Remote Sens. 2007, 1, 013536. [Google Scholar]
- Blackburn, G.A. Remote sensing of forest pigments using airborne imaging spectrometer and LIDAR imagery. Remote Sens. Environ. 2002, 82, 311–332. [Google Scholar]
- Asner, G.P.; Knapp, D.E.; Kennedy-Bowdoin, T.; Jones, M.O.; Martin, R.E.; Boardman, J.; Hughes, R.F. Invasive species detection in Hawaiian rainforests using airborne imaging spectroscopy and LiDAR. Remote Sens. Environ. 2008, 112, 1942–1955. [Google Scholar]
- Asner, G.P.; Martin, R.E. Spectral and chemical analysis of tropical forests: Scaling from leaf to canopy levels. Remote Sens. Environ. 2008, 112, 3958–3970. [Google Scholar]
- Drake, S.E. Climate-correlative Modeling of Phytogeography at the Watreshed Scale; University of Arizona: Tucson, AZ, USA, 2000. [Google Scholar]
- Anderson, G.L.; Everitt, J.H.; Richardson, A.J.; Escobar, D.E. Using satellite data to map false broomweed (Ericameria austrotexana) infestations on south Texas rangelands. Weed Technol. 1993, 7, 865–871. [Google Scholar]
- Bradley, B.A.; Houghton, R.A.; Mustard, J.F.; Hamburg, S.P. Invasive grass reduces aboveground carbon stocks in shrublands of the Western US. Glob. Change Biol. 2006, 12, 1815–1822. [Google Scholar]
- Jösson, P.; Eklundh, L. TIMESAT - A program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar]
- Okin, G.S.; Roberts, D.A.; Murray, B.; Okin, W.J. Practical limits on hyperspectral vegetation discrimination in arid and semiarid environments. Remote Sens. Environ. 2001, 77, 212–225. [Google Scholar]
- Asner, G.P.; Martin, R.E.; Ford, A.J.; Metcalfe, D.J.; Liddell, M.J. Leaf chemical and spectral diversity in Australian tropical forests. Ecol. Appl. 2009, 19, 236–253. [Google Scholar]
- Asner, G.P.; Haxo, R.S.; Knapp, D.E. Computing for analysis and modeling of hyperspectral imagery. In High Performance Computing for Remote Sensing; Plaza, A., Chang, C.I., Eds.; Chapman and Hall Press: New York, NY, USA, 2008; pp. 109–130. [Google Scholar]
- Braswell, B.H.; Hagen, S.C.; Frolking, S.E.; Salas, W.A. A multivariable approach for mapping sub-pixel land cover distributions using MISR and MODIS: application in the Brazilian Amazon region. Remote Sens. Environ. 2003, 87, 243–256. [Google Scholar]
- Asner, G.P.; Braswell, B.H.; Schimel, D.S.; Wessman, C.A. Ecological research needs from multiangle remote sensing data. Remote Sens. Environ. 1998, 63, 155–165. [Google Scholar]
- Diner, D.J.; Asner, G.P.; Davies, R.; Muller, J.-P.; Pinty, B.; Schaaf, C.B.; Stroeve, J. New directions in Earth observing: Scientific applications of multiple-view-angle remote sensing. Bull. Am. Meteorol. Soc. 1999, 80, 2209–2228. [Google Scholar]
- Cronk, Q.C.B.; Fuller, J.L. Plant Invaders: The Threat to Natural Ecosystems; Earthscan Publications: London, UK, 2001. [Google Scholar]
- Mitich, L.W. Kudzu [Pueraria lobata (Willd.) Ohwi]1. Weed Technol. 2000, 14, 231–235. [Google Scholar]
- Turner, M.G.; Dale, V.H.; Gardner, R.H. Predicting across scales: Theory development and testing. Landscape Ecol. 1989, 3, 245–252. [Google Scholar]
- Levin, S.A. The problem of pattern and scale in ecology: The Robert H. MacArthur award lecture. Ecology 1992, 73, 1943–1967. [Google Scholar]
- Huang, C.; Asner, G.P.; Martin, R.; Barger, N.; Neff, J. Multiscale analysis of tree cover and aboveground carbon stocks in pinyon-juniper woodlands. Ecol. Appl. 2009, 19, 668–681. [Google Scholar]
- Goovaerts, P. Geostatistics for Natural Resources Evaluation; Oxford University Press: New York, NY, USA, 1997. [Google Scholar]
- Hudak, A.T.; Wessman, C.A. Textural analysis of historical aerial photography to characterize woody plant encroachment in South African Savanna. Remote Sens. Environ. 1998, 66, 317–330. [Google Scholar]
- Bradley, B.A. Regional analysis of the impacts of climate change on cheatgrass invasion shows potential risk and opportunity. Glob. Change Biol. 2009, 15, 196–208. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Moderate spatial/ spectral | Sensor specifications and examples:
|
Species traits and remote sensing strategies:
| |
Limitations:
| |
| High spatial | Sensor specifications and examples:
|
Species traits and remote sensing strategies:
| |
Limitations:
| |
| High temporal | Sensor specifications and examples:
|
| High temporal | Species traits and remote sensing strategies:
|
Limitations:
| |
| Hyperspectral | Sensor specifications and examples:
|
Species traits and remote sensing strategies:
| |
Limitations:
| |
| Active remote sensing | Sensor specifications and examples:
|
Species traits and remote sensing strategies:
| |
Limitations:
| |
| Image fusion | Sensor specifications and examples:
|
Species traits and remote sensing strategies:
| |
Limitations:
|
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, C.-y.; Asner, G.P. Applications of Remote Sensing to Alien Invasive Plant Studies. Sensors 2009, 9, 4869-4889. https://doi.org/10.3390/s90604869
Huang C-y, Asner GP. Applications of Remote Sensing to Alien Invasive Plant Studies. Sensors. 2009; 9(6):4869-4889. https://doi.org/10.3390/s90604869
Chicago/Turabian StyleHuang, Cho-ying, and Gregory P. Asner. 2009. "Applications of Remote Sensing to Alien Invasive Plant Studies" Sensors 9, no. 6: 4869-4889. https://doi.org/10.3390/s90604869
APA StyleHuang, C.-y., & Asner, G. P. (2009). Applications of Remote Sensing to Alien Invasive Plant Studies. Sensors, 9(6), 4869-4889. https://doi.org/10.3390/s90604869