Detection of Salmonella by Surface Plasmon Resonance


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
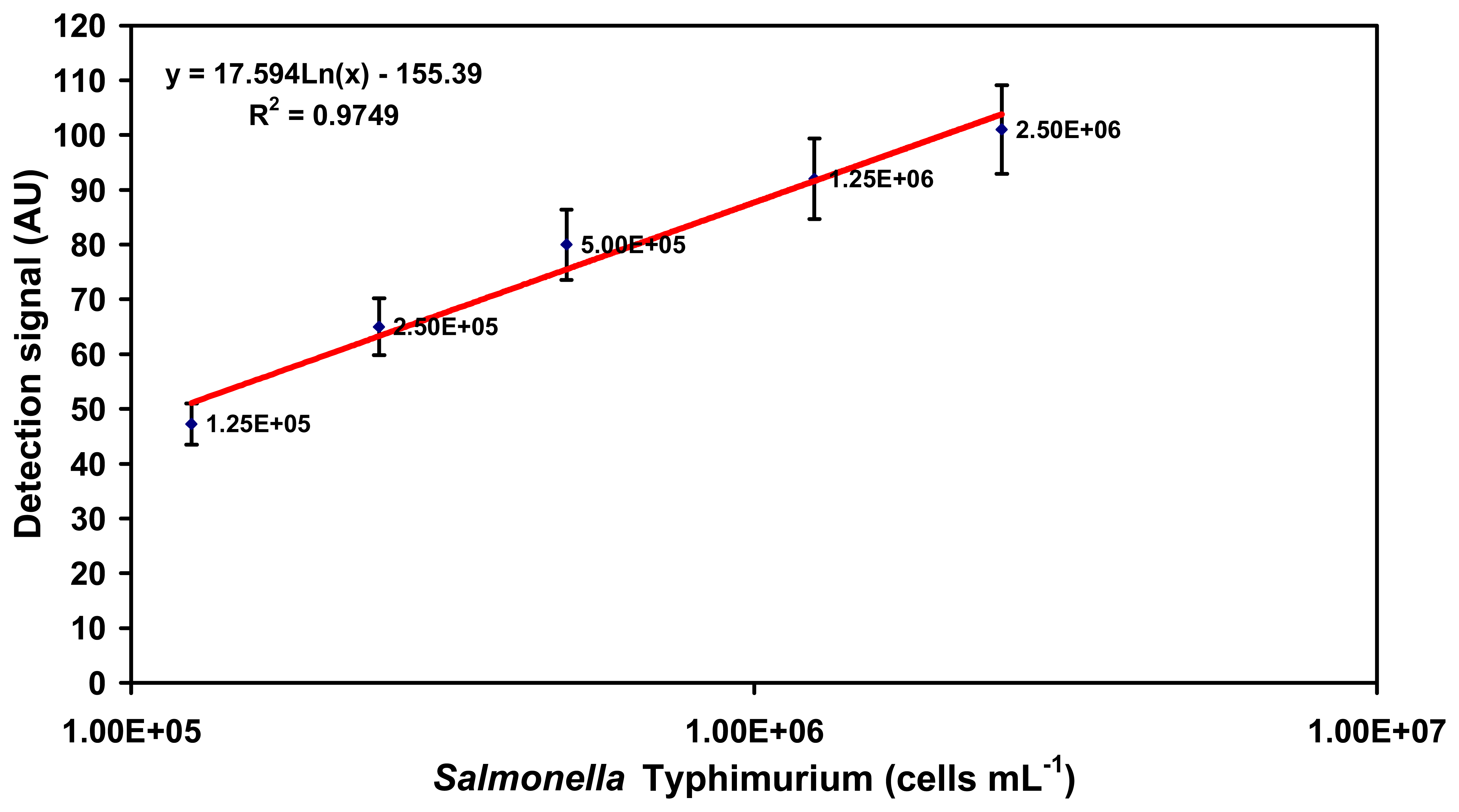
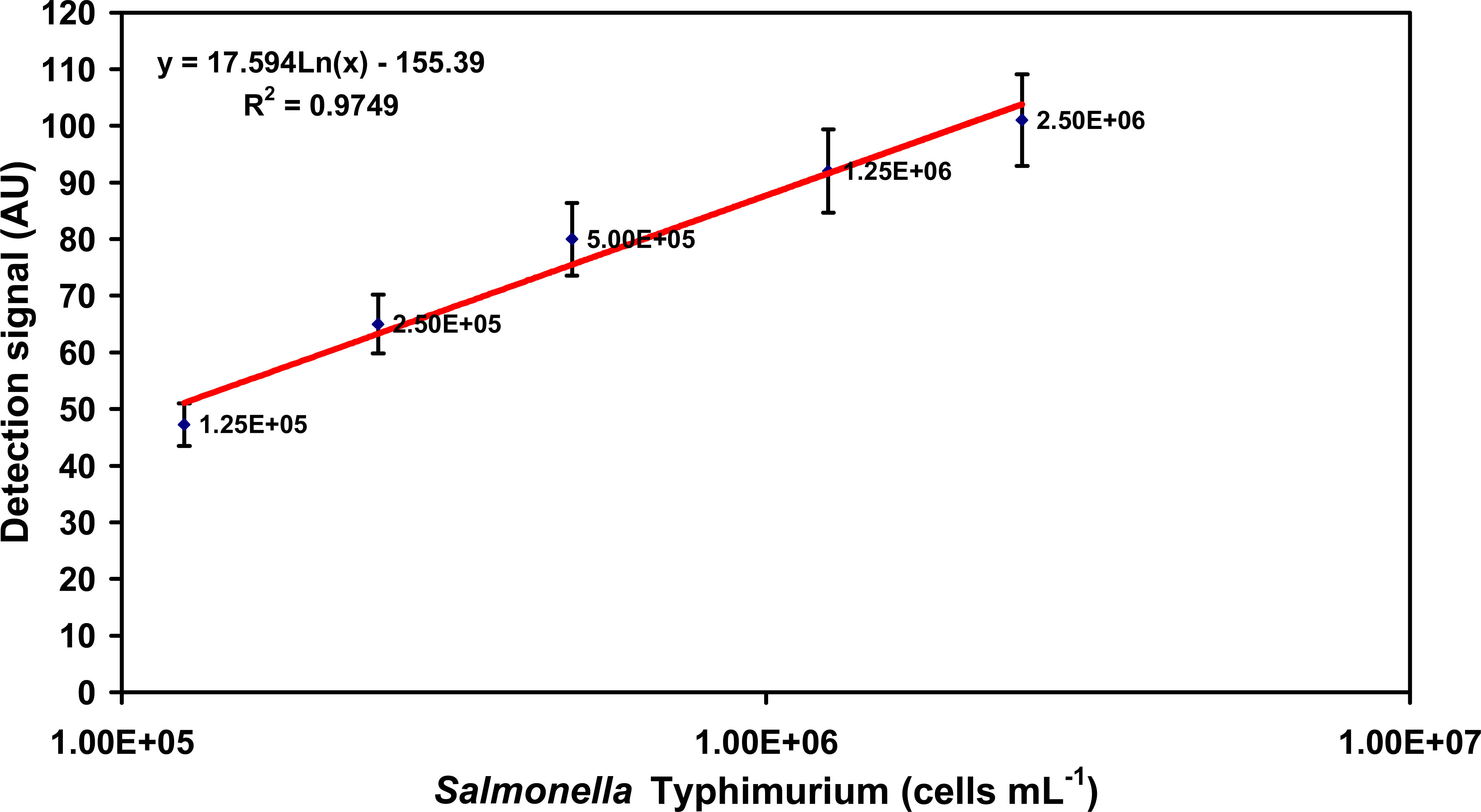
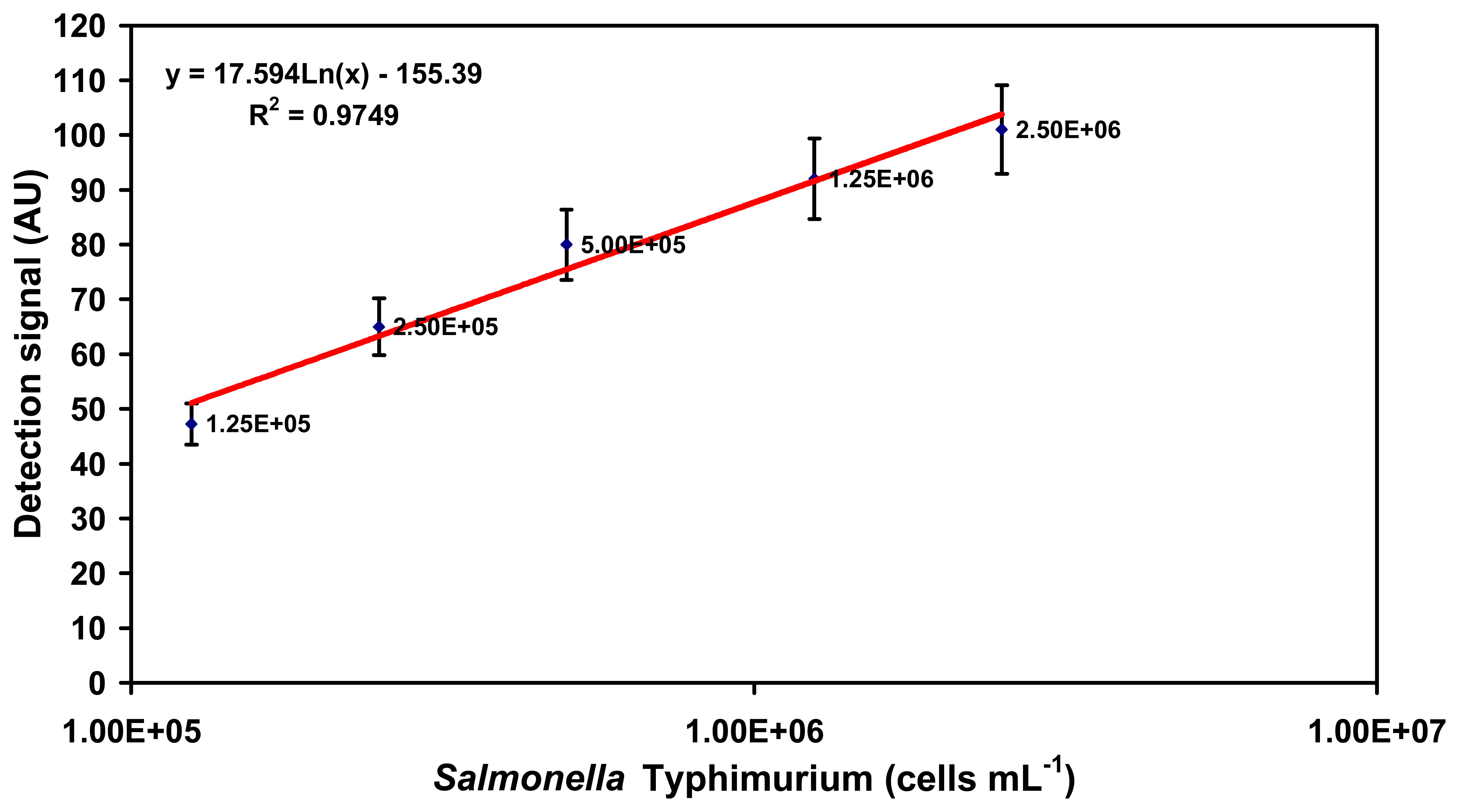
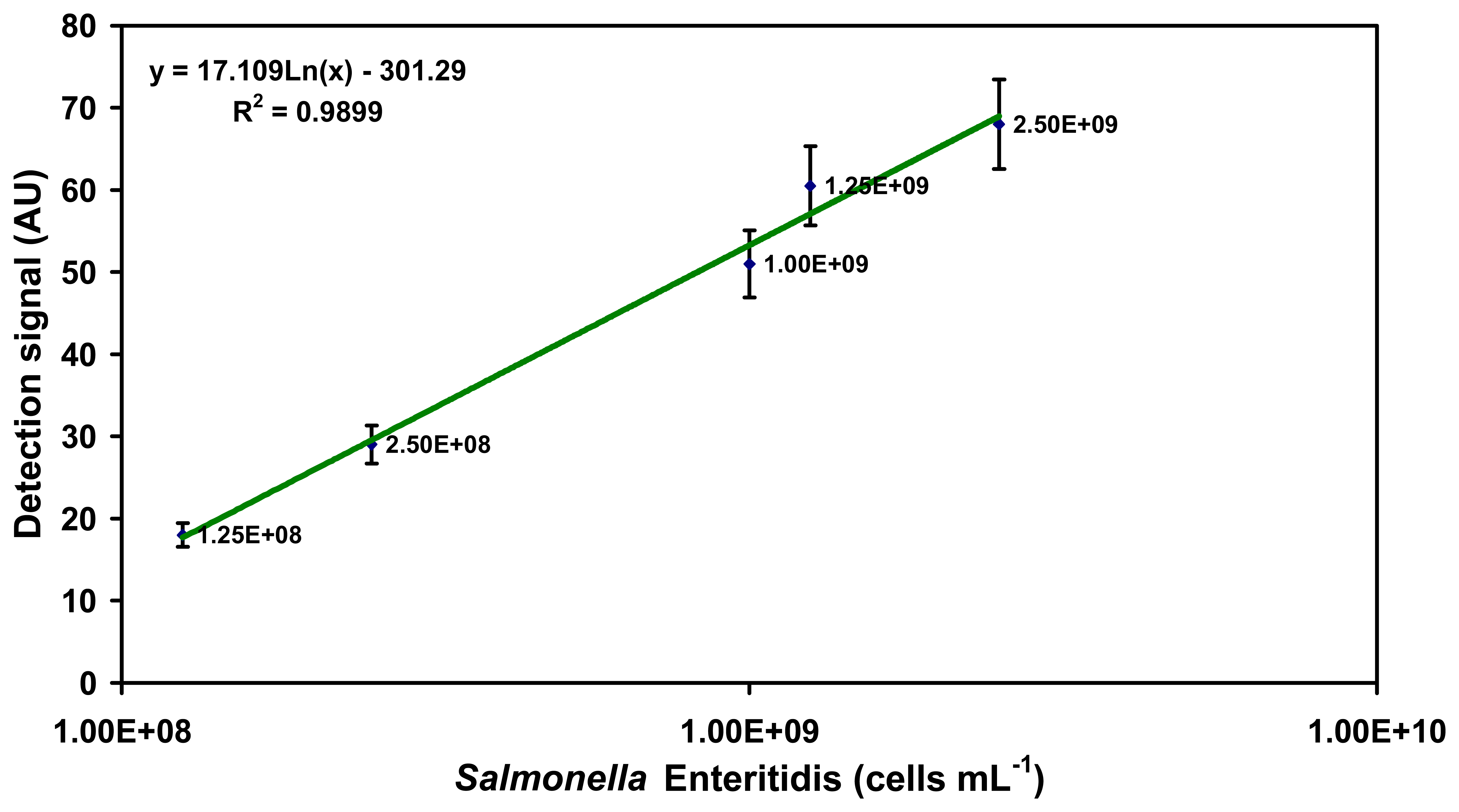
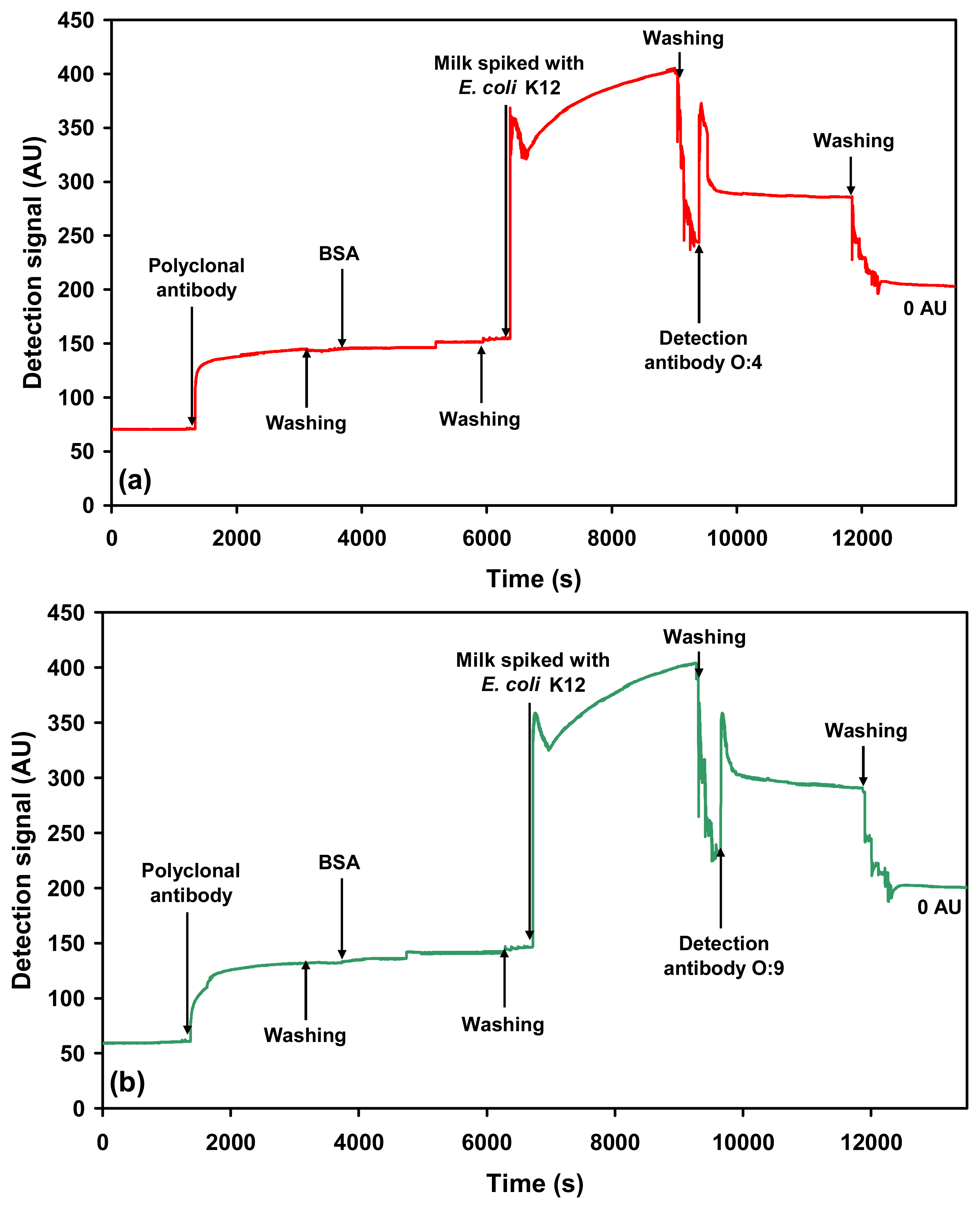
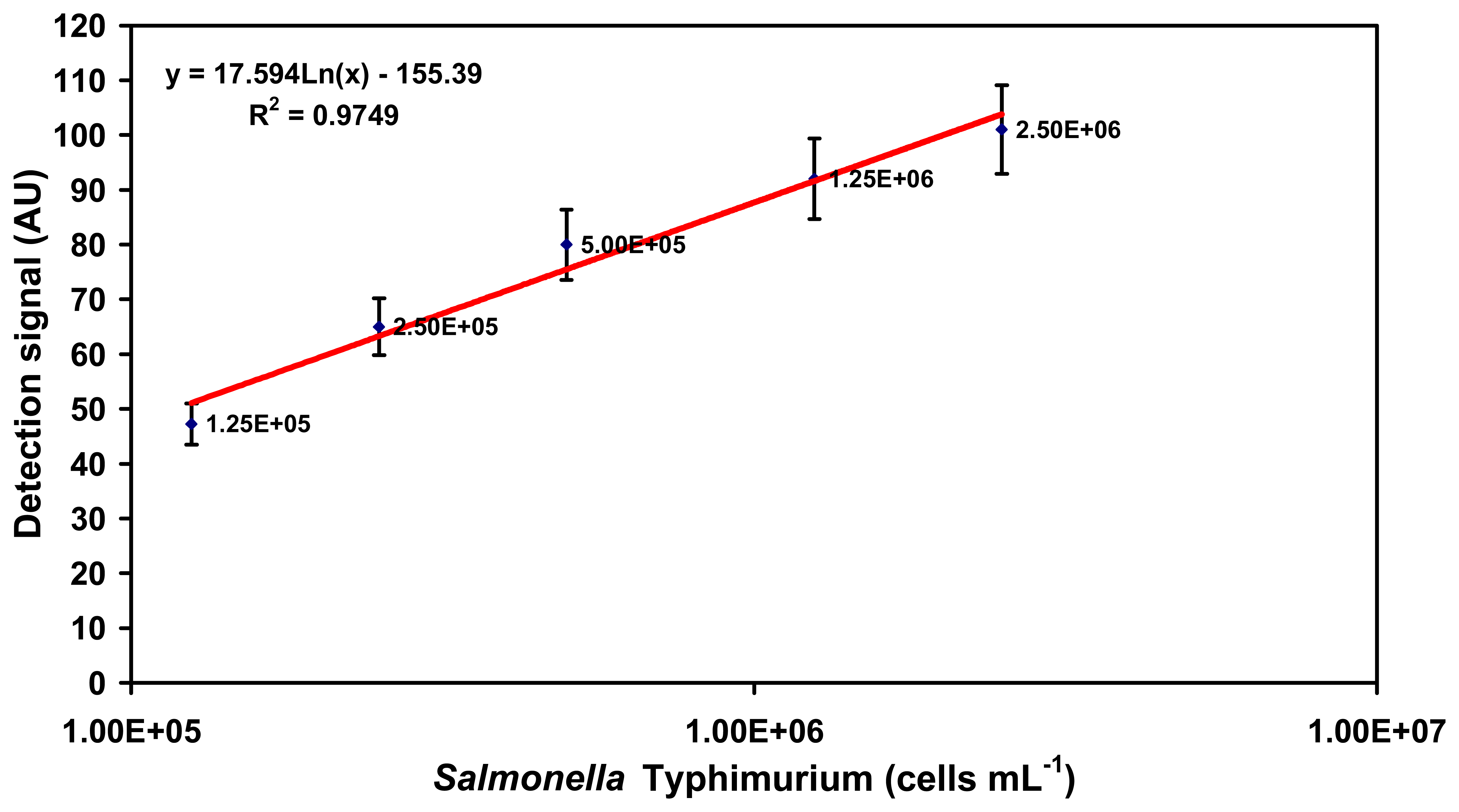
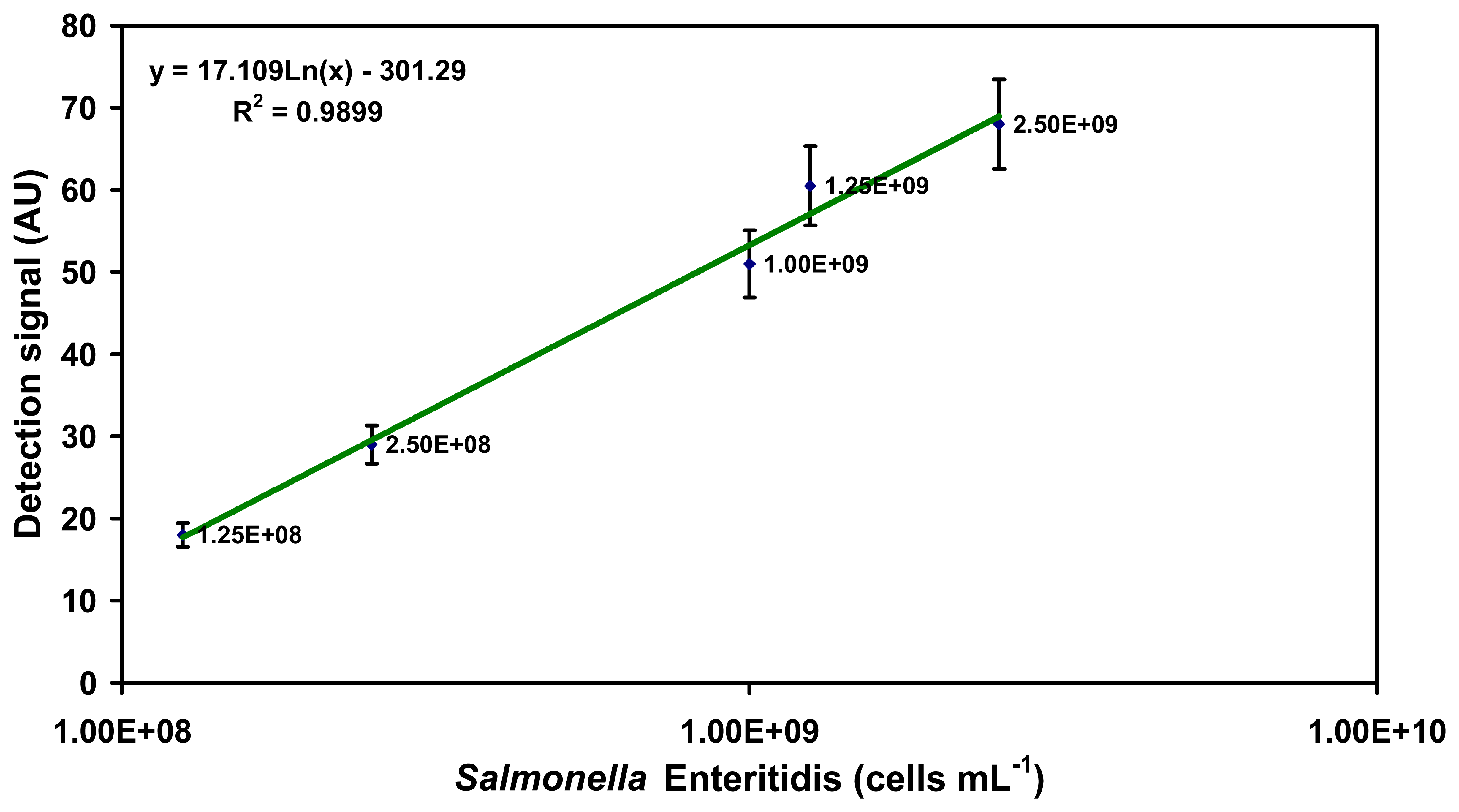
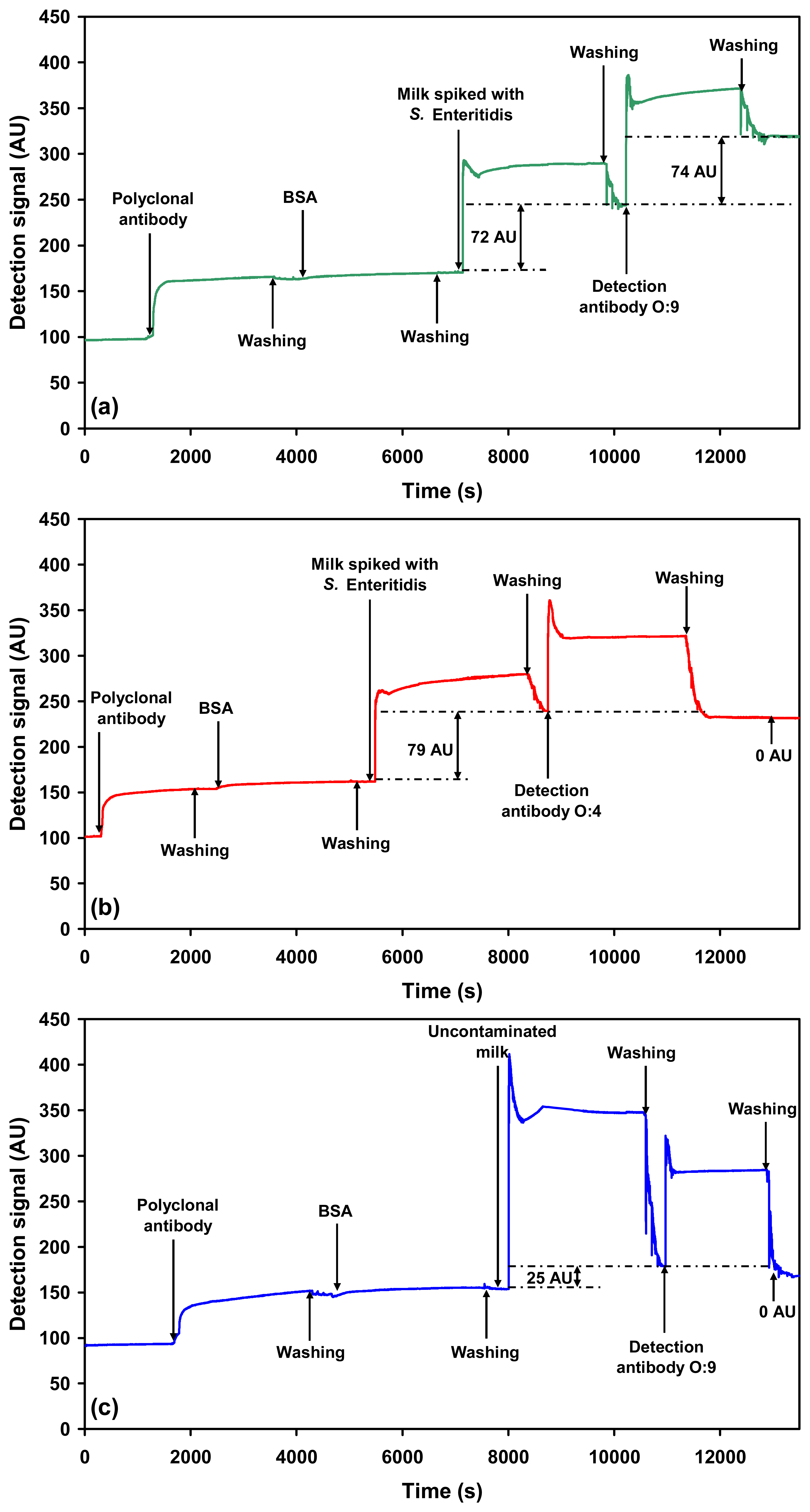
2.1. Specific detection of Salmonella serovars in buffer
2.2. Specific detection of Salmonella serovars in milk
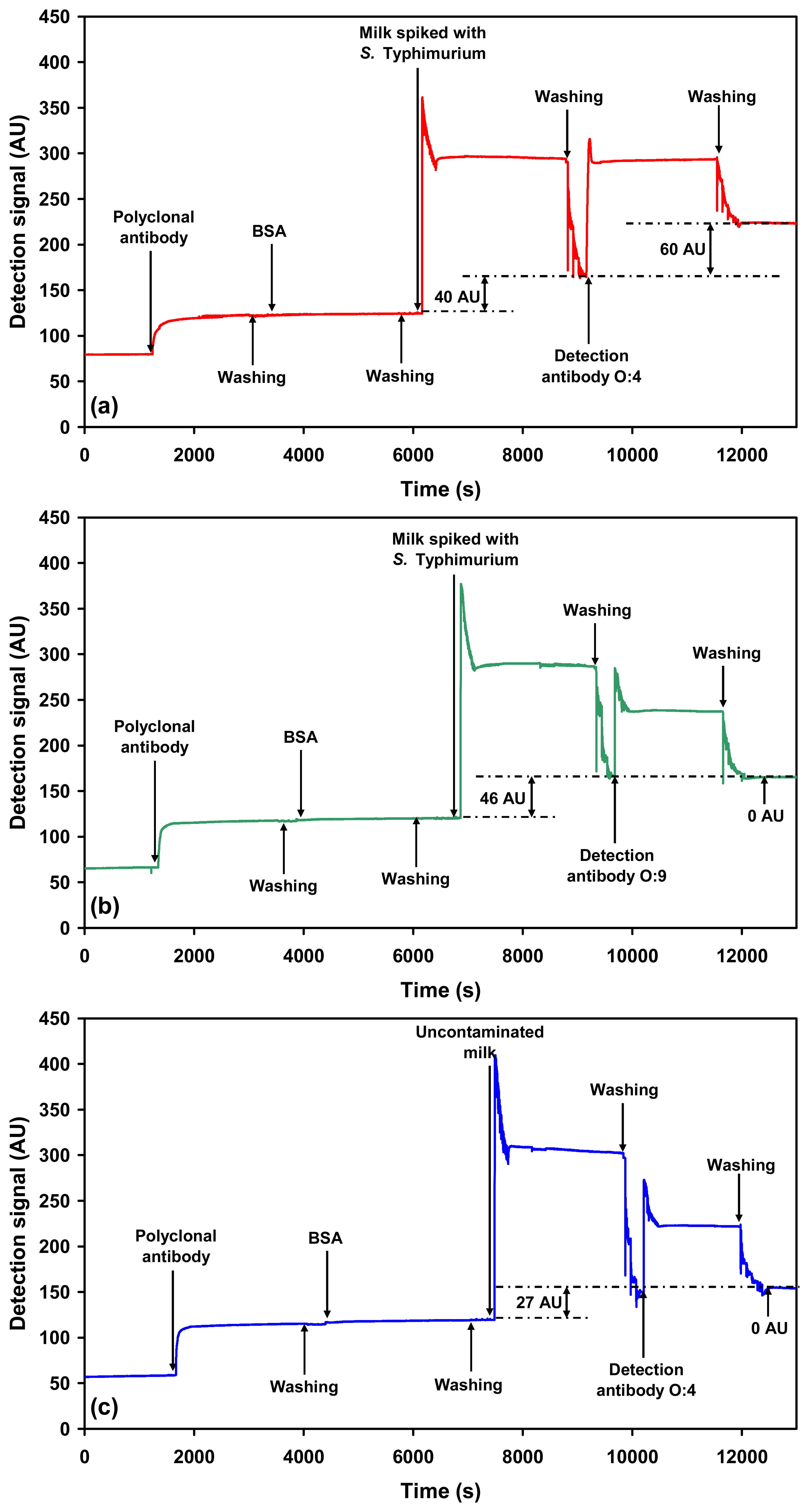
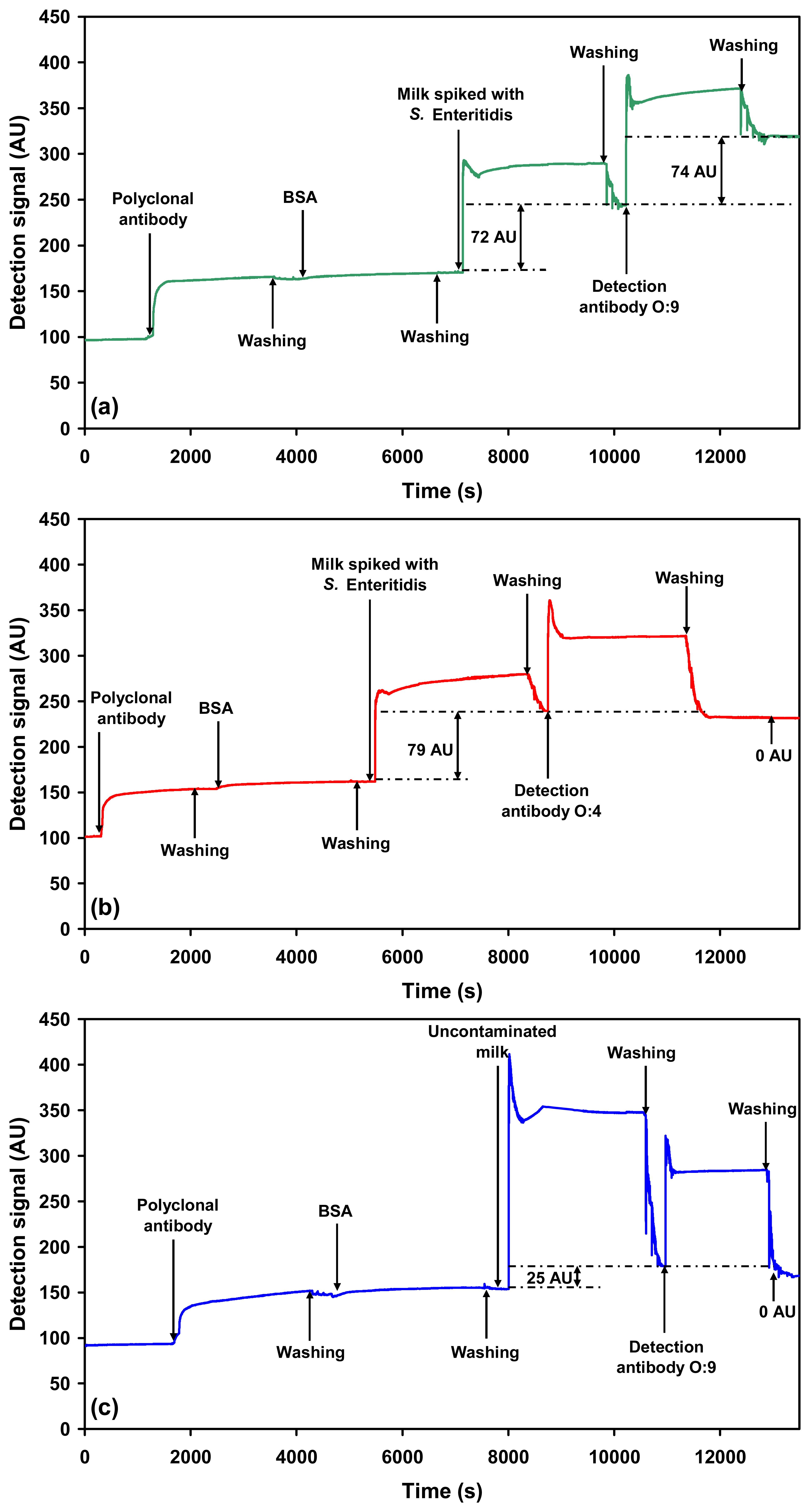
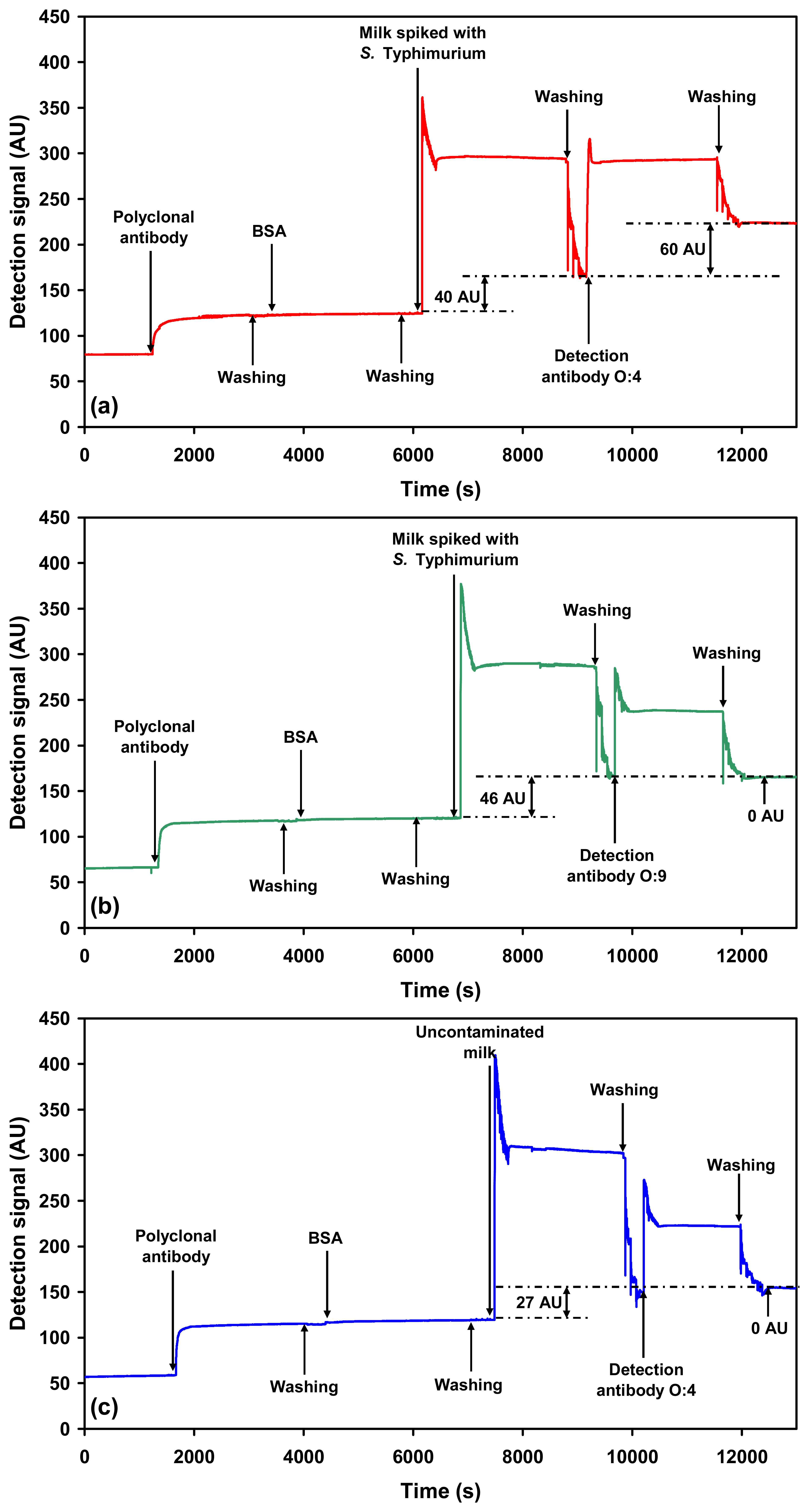
2.2.1. Detection of each serovar in milk using O-specific antibody
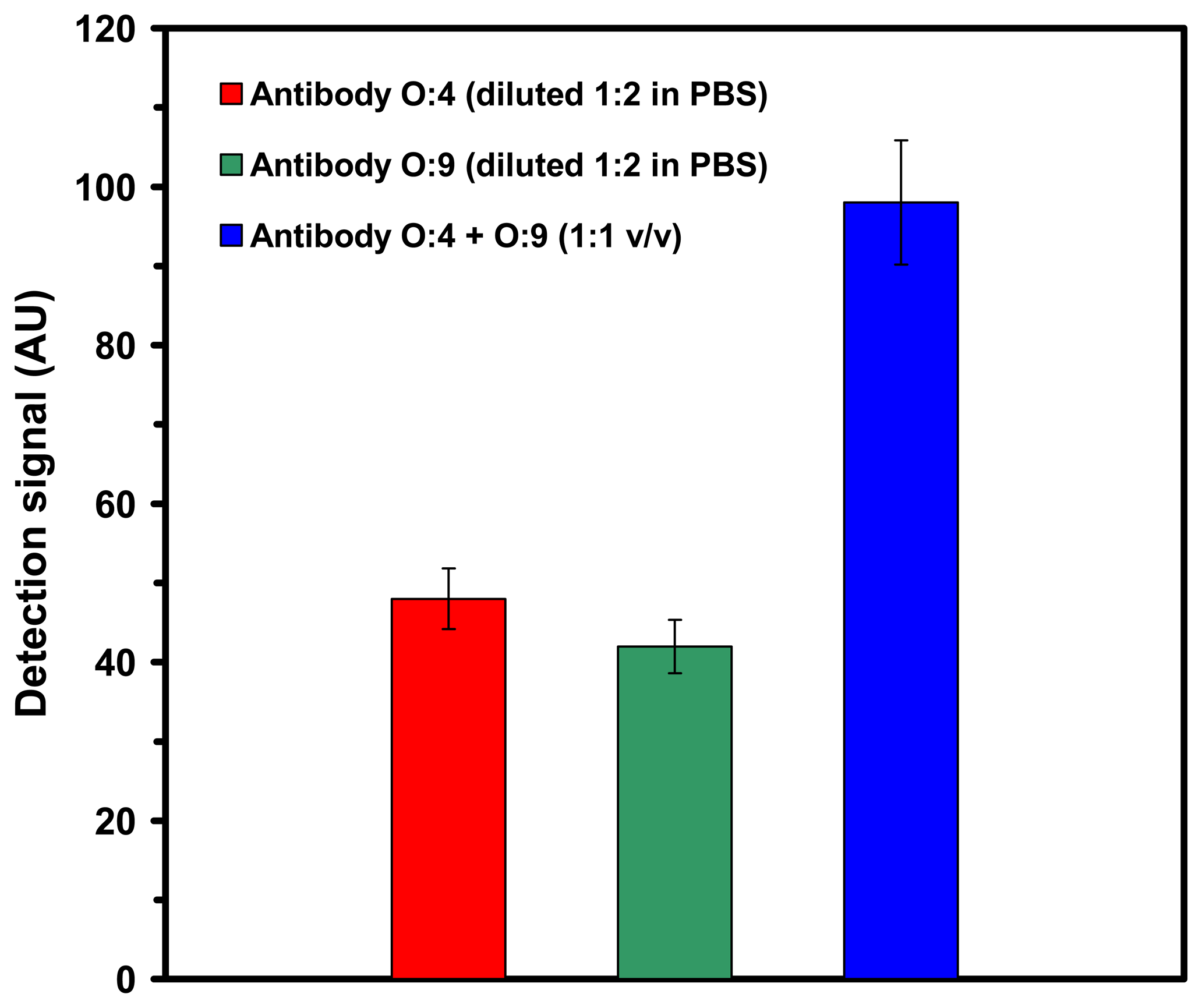
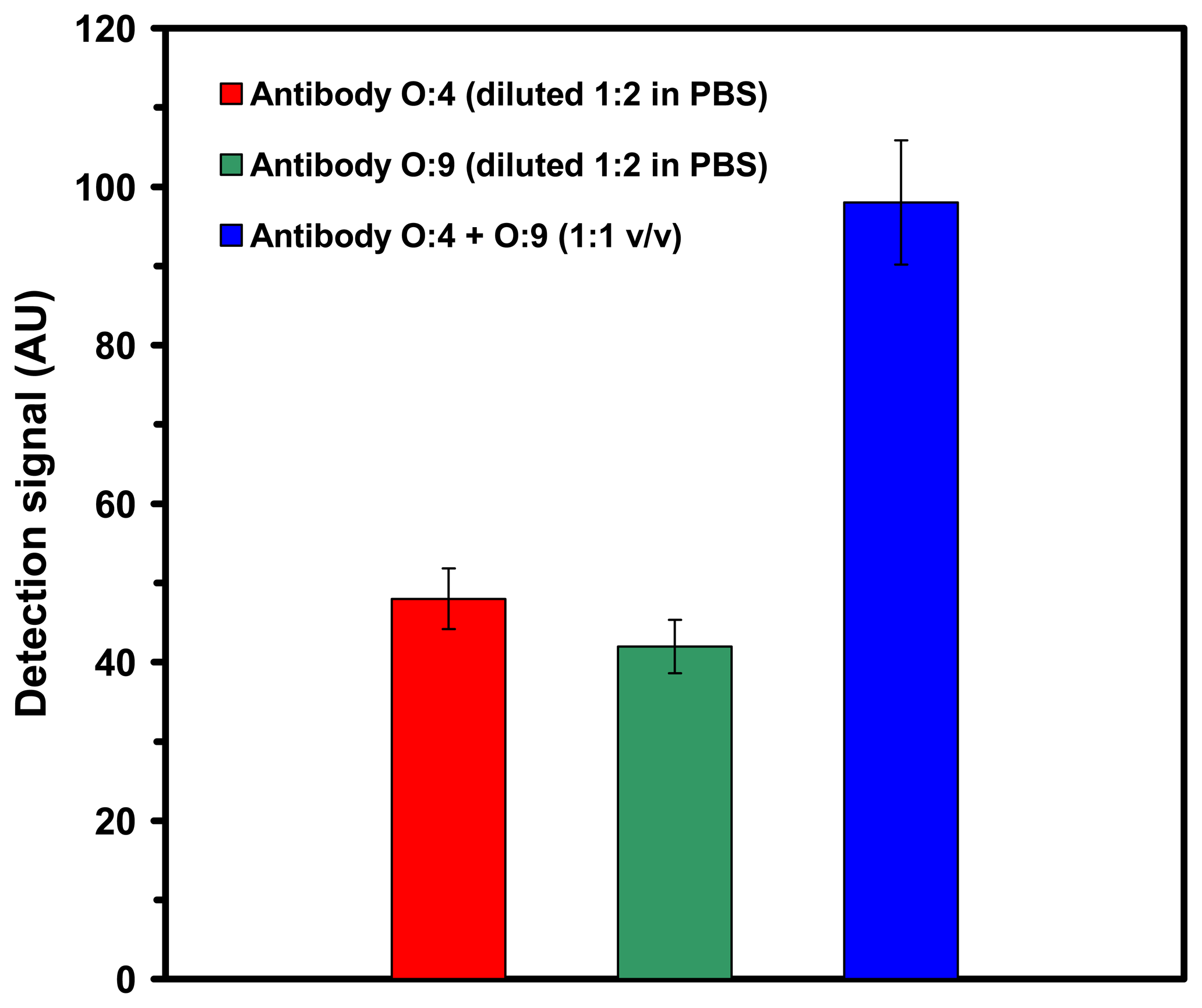
2.2.2. Individual detection of serovars in a mixture using monoclonal O-specific antibodies
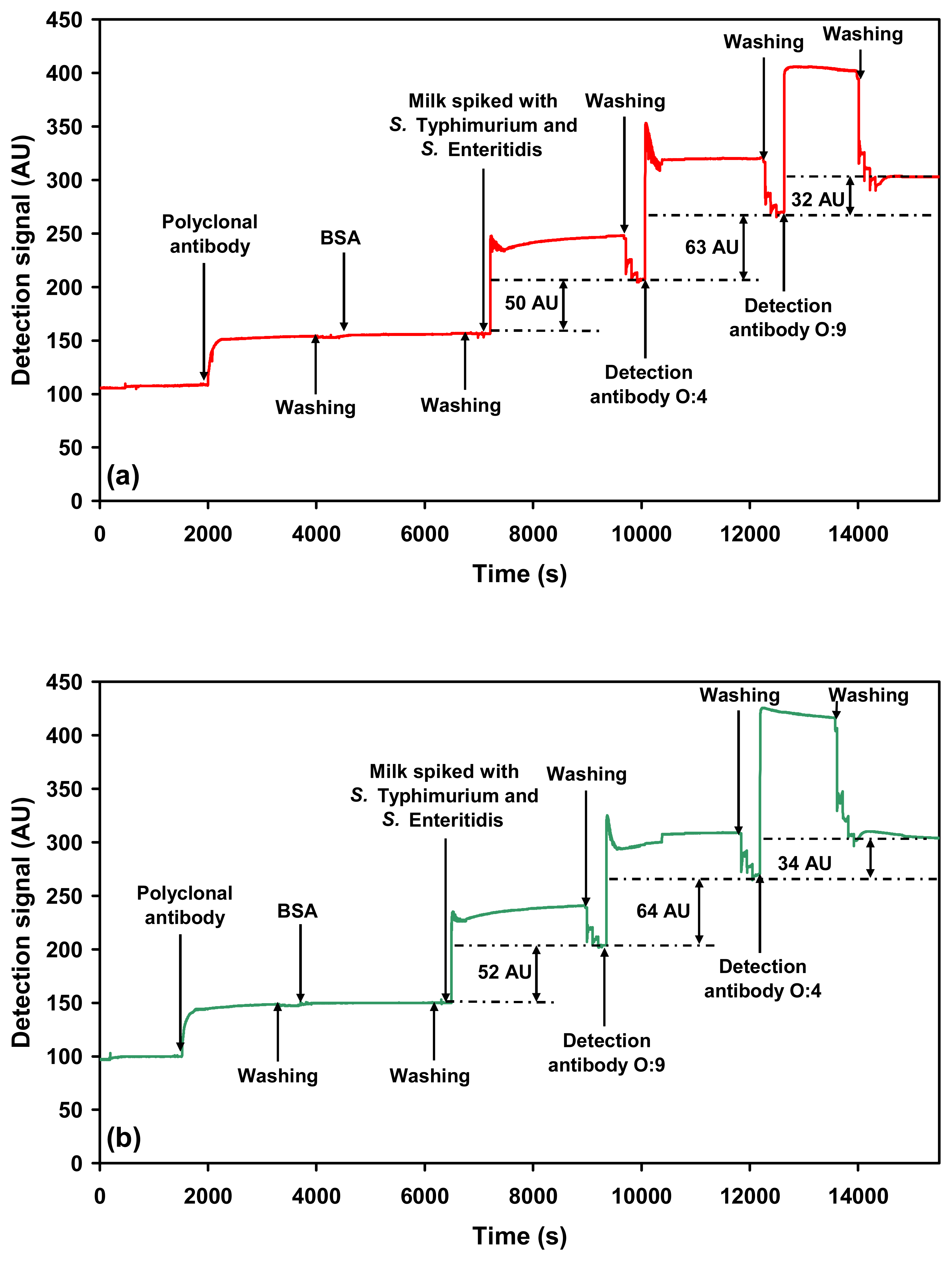
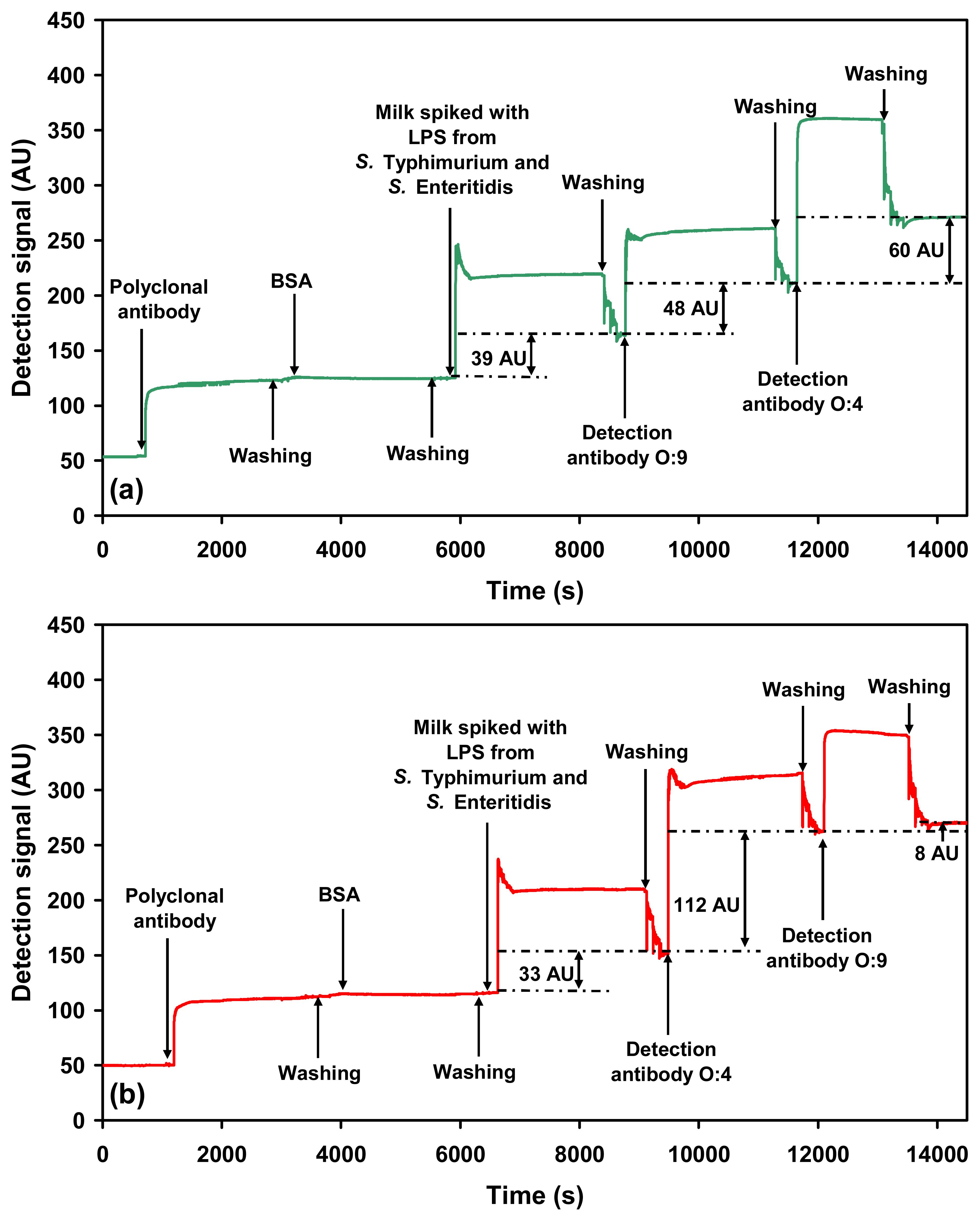
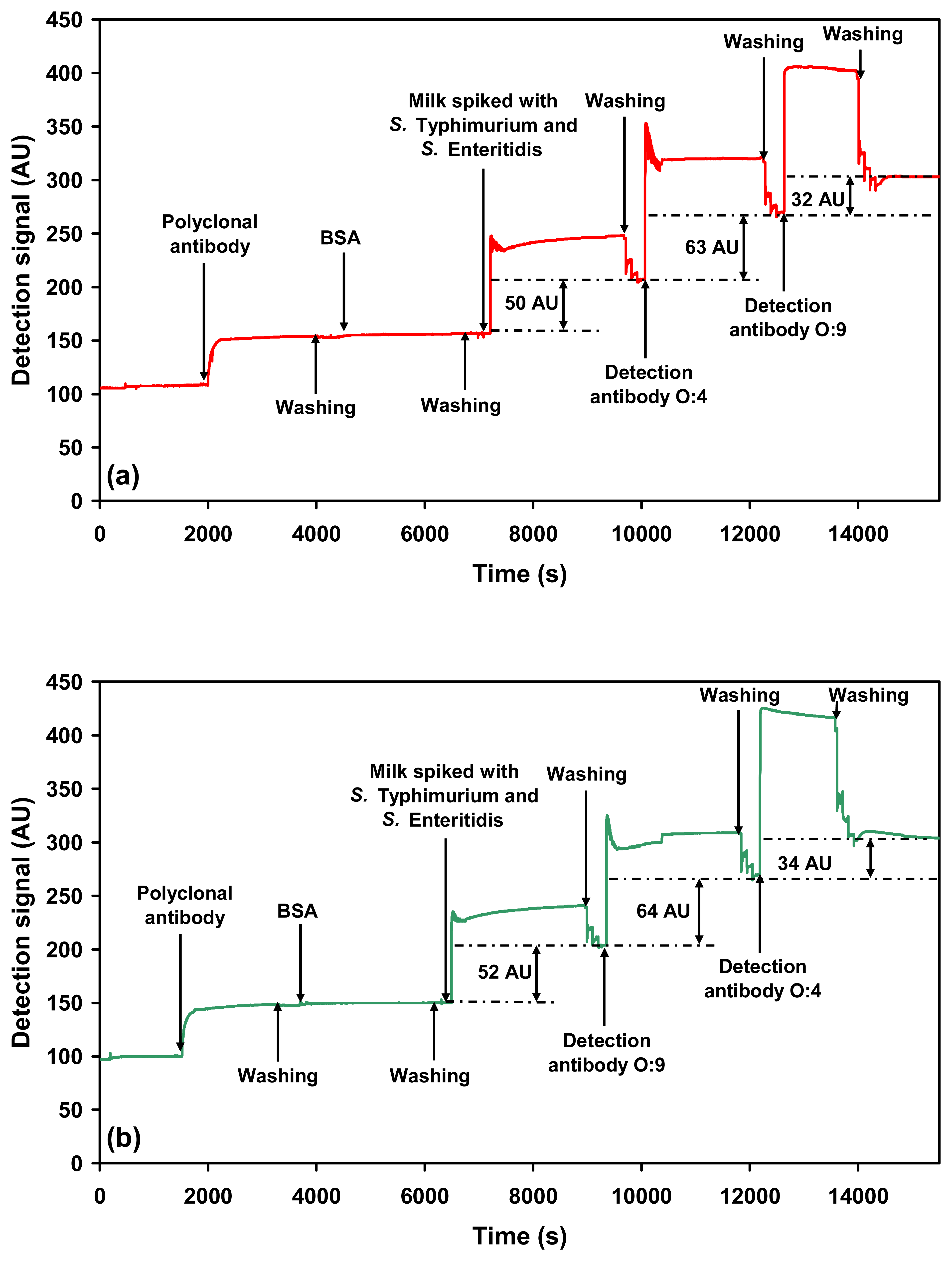
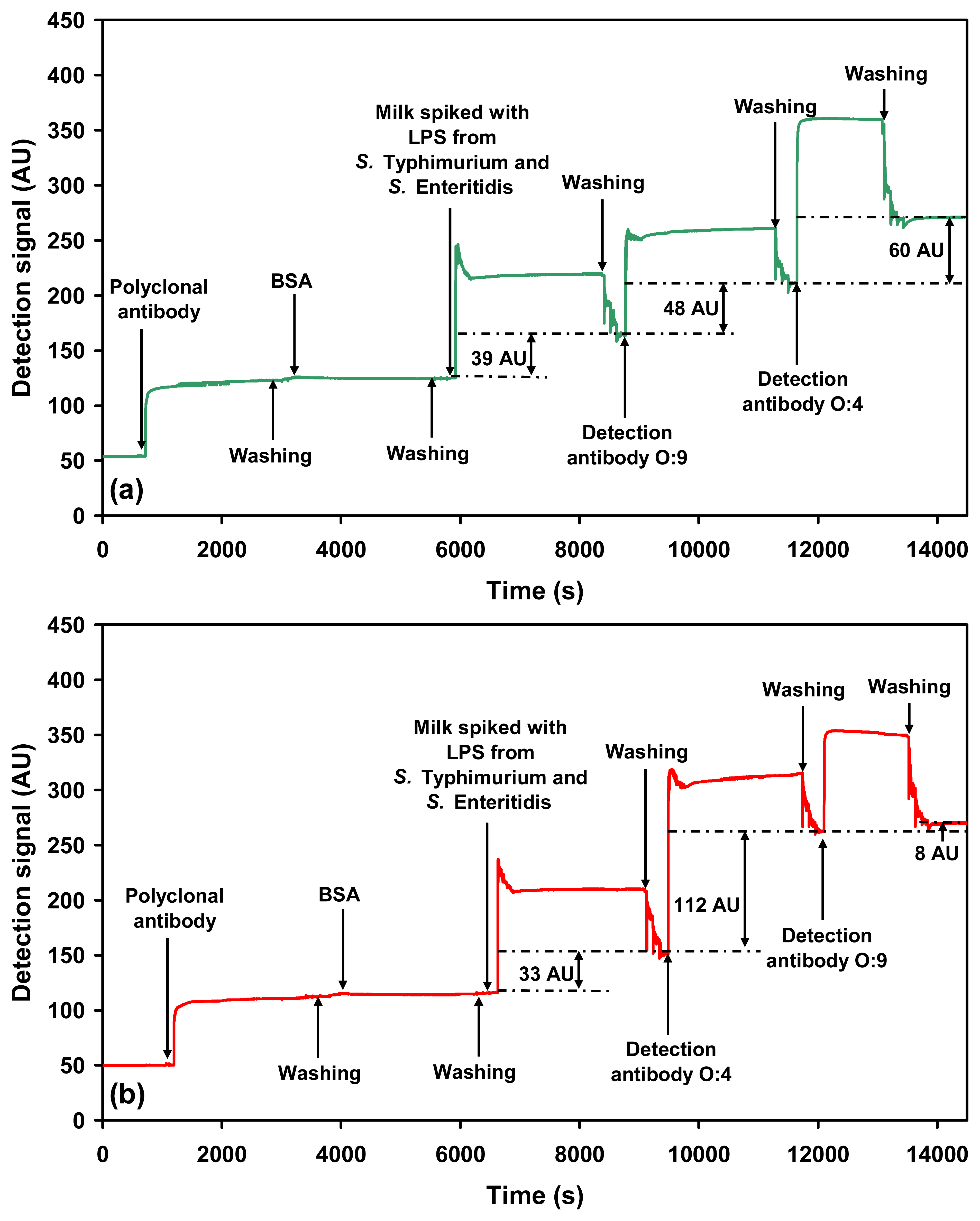
2.2.3. Sequential detection of serovars in a mixture
3. Experimental Section
3.1. Chemicals
3.2. Preparation of Salmonella antigen
3.3. Surface coating
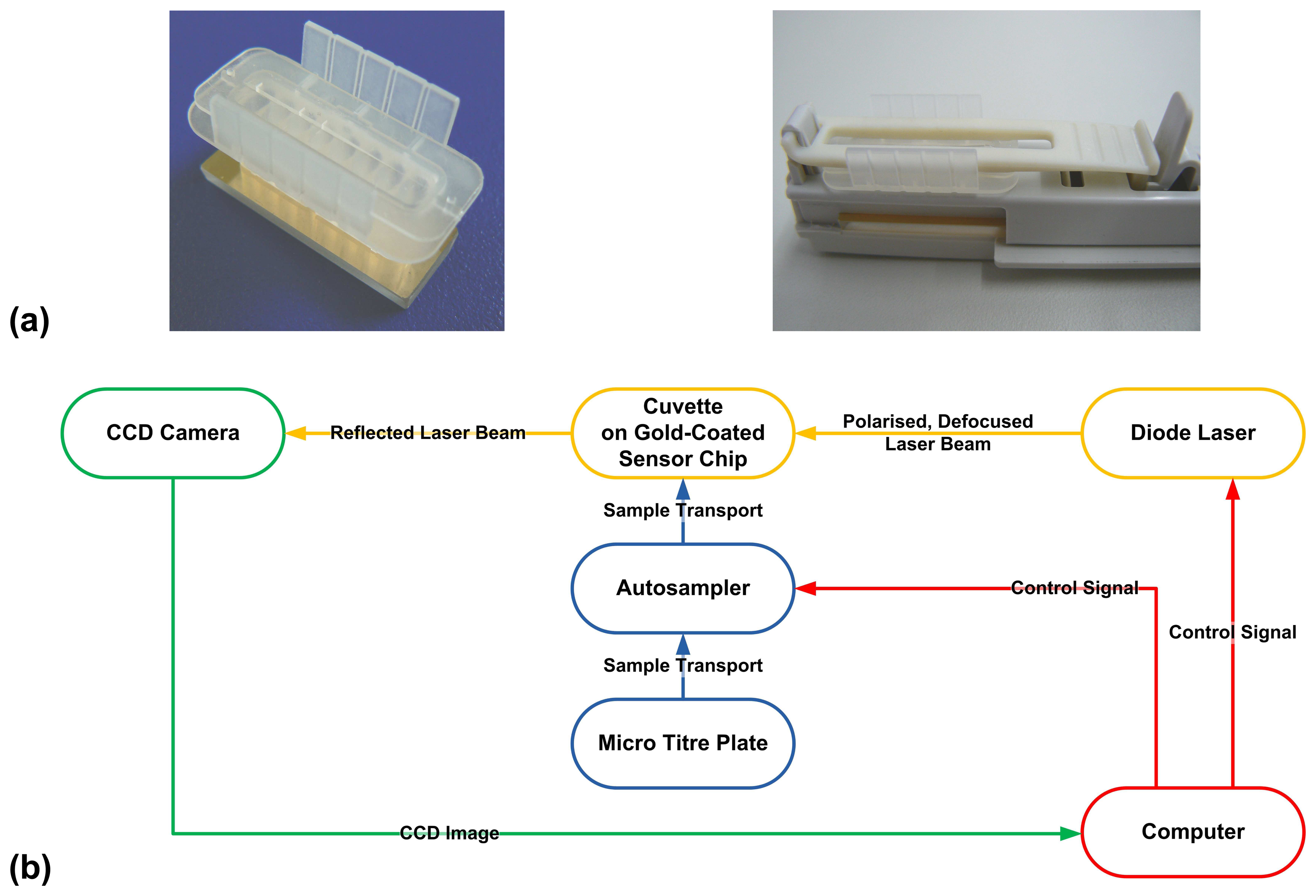
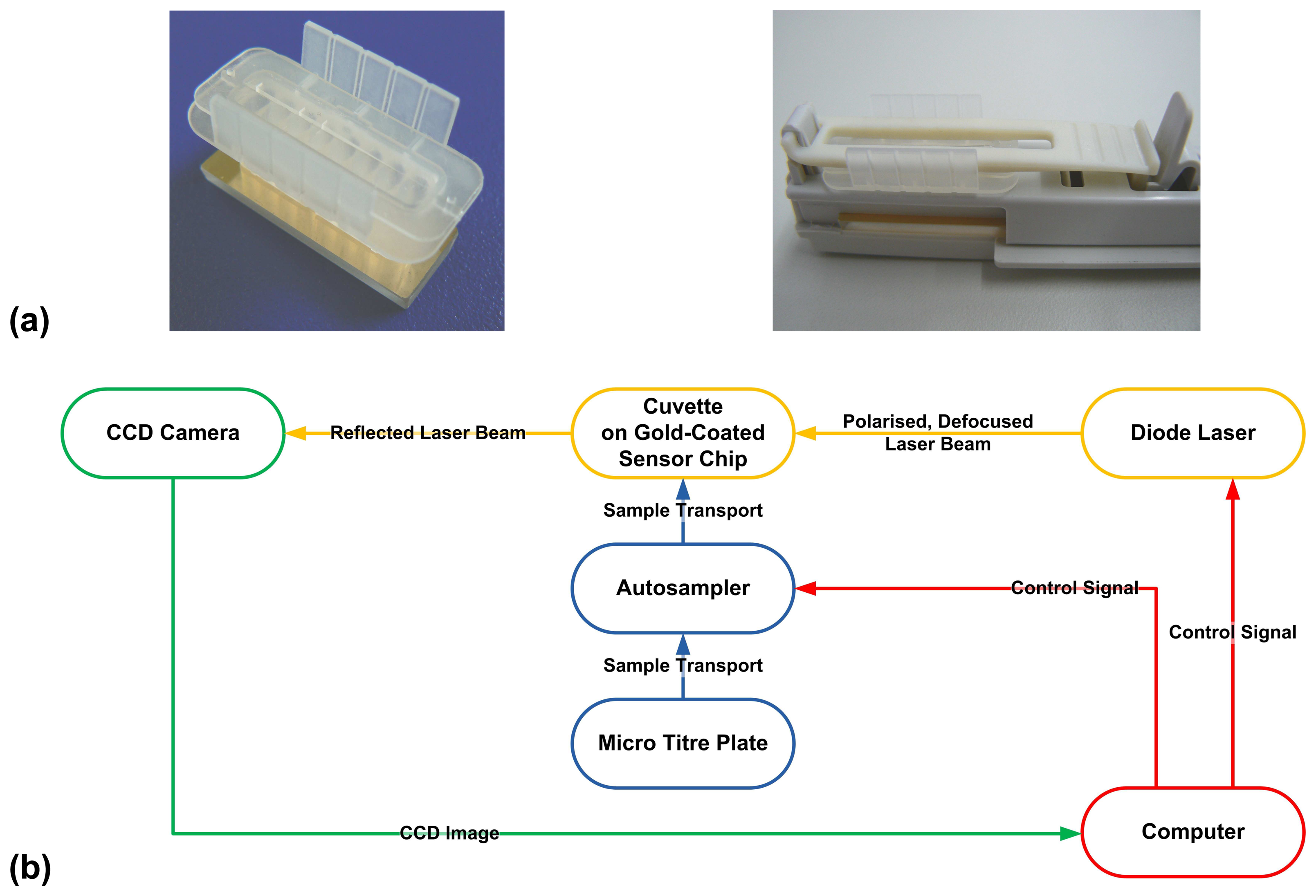
3.4. Surface plasmon resonance device
3.5. Measurement conditions
3.6. Assay setup
Acknowledgments
References and Notes
- Arshad, M.M.; Wilkins, M.J.; Downes, F.P.; Rahbar, M.H.; Erskine, R.J.; Boulton, M.L.; Saeed, A.M. A registry-based study on the association between human salmonellosis and routinely collected parameters in Michigan, 1995-2001. Foodborne Pathog. Dis. 2007, 4(1), 16–25. [Google Scholar]
- Robert-Koch-Institut: Berlin; Aktuelle Statistik meldepflichtiger Infektionskrankheiten. In Epid. Bull.; Volume 21, 2007.
- Thornton, L.; Gray, S.; Bingham, P.; Salmon, R.; Hutchinson, D.; Rowe, B.; Newton, D.; Syed, Q. The problems of tracing a geographically widespread outbreak of salmonellosis from a commonly eaten food: Salmonella typhimurium DT193 in north west England and north Wales in 1991. Epidemiol. Infect. 1993, 111(3), 465–471. [Google Scholar]
- [CDC] Centers for Disease Control and Prevention. Preliminary FoodNet Data on the Incidence of Infection with Pathogens Transmitted Commonly Through Food -10 States, United States, 2006. Morbid. Mortal. Wkly. Rep. 2007, 56(14), 336–339. [Google Scholar]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5(5), 607–625. [Google Scholar]
- June, G.A.; Sherrod, P.S.; Hammack, T.S.; Amaguana, R.M.; Andrews, W.H. Relative effectiveness of selenite cystine broth, tetrathionate broth, and rappaport-vassiliadis medium for recovery of Salmonella spp. from raw flesh, highly contaminated foods, and poultry feed: Collaborative study. J. AOAC Int. 1996, 79(6), 1307–1323. [Google Scholar]
- Rose, B. E. USDA/FSIS Microbiology Laboratory Guidebook, 3rd ed.; Dey, B.P., Lattuada, C.P., Eds.; USDA Food Safety Inspection Service, 1998; Chapter 4; p. 1. [Google Scholar]
- Schneid, A.D.; Rodrigues, K.L.; Chemello, D.; Tondo, E. C.; Ayub, M. A. Z.; Aleixo, J. A. G. Evaluation of an indirect ELISA for the detection of Salmonella in chicken meat. Braz. J. Microbiol. 2006, 37(3), 350–355. [Google Scholar]
- Mozola, M.A. Genetics-based methods for detection of Salmonella spp. in foods. J. AOAC Int. 2006, 89(2), 517–529. [Google Scholar]
- D'Souza, D.H.; Jaykus, L.A. Nucleic acid sequence based amplification for the rapid and sensitive detection of Salmonella enterica from foods. J. Appl. Microbiol. 2003, 95(6), 1343–1350. [Google Scholar]
- Lazcka, O.; Del Campo, F.J.; Munoz, F.X. Pathogen detection: A perspective of traditional methods and biosensors. Biosens. Bioelectron. 2007, 22(7), 1205–1217. [Google Scholar]
- Liedberg, B.; Nylander, C.; Lundstrom, I. Biosensing with Surface-plasmon resonance - how it all started. Biosens. Bioelectron. 1995, 10(8), i–ix. [Google Scholar]
- Lundstrom, I. Real-time biospecific interaction analysis. Biosens. Bioelectron. 1994, 9(9-10), 725–736. [Google Scholar]
- Mazumdar, S.D.; Hartmann, M.; Kämpfer, P.; Keusgen, M. Rapid method for detection of Salmonella in milk by surface plasmon resonance (SPR). Biosens. Bioelectron. 2007, 22(9-10), 2040–2046. [Google Scholar]
- Waswa, J.W.; Irudayaraj, J.; DebRoy, C. Direct detection of E-Coli O157 : H7 in selected food systems by a surface plasmon resonance biosensor. LWT-Food Science and Tech. 2007, 40(2), 187–192. [Google Scholar]
- Waswa, J.W.; Debroy, C.; Irudayaraj, J. Rapid detection of Salmonella enteritidis and Escherichia coli using surface plasmon resonance biosensor. J. Food Process Eng. 2006, 29(4), 373–385. [Google Scholar]
- Jyoung, J.; Hong, S.; Lee, W.; Choi, J. Immunosensor for the detection of Vibrio cholerae O1 using surface plasmon resonance. Biosens. Bioelectron. 2006, 21(12), 2315–2319. [Google Scholar]
- Meeusen, C.; Alocilja, E.; Osburn, W. Detection of E-coli O157 : H7 using a miniaturized surface plasmon resonance biosensor. Trans. ASAE 2005, 48(6), 2409–2416. [Google Scholar]
- Bokken, G.; Corbee, R.J.; van Knapen, F.; Bergwerff, A. A. Immunochemical detection of Salmonella group B, D and E using an optical surface plasmon resonance biosensor. FEMS Microbiol. Lett. 2003, 222(1), 75–82. [Google Scholar]
- Oh, B.K.; Lee, W.; Chun, B. S.; Bae, Y. M.; Lee, W. H.; Choi, J. W. The fabrication of protein chip based on surface plasmon resonance for detection of pathogens. Biosens. Bioelectron. 2005, 20(9), 1847–1850. [Google Scholar]
- Perkins, E.A.; Squirrell, D.J. Development of instrumentation to allow the detection of microorganisms using light scattering in combination with surface plasmon resonance. Biosens. Bioelectron. 2000, 14(10-11), 853–859. [Google Scholar]
- Fratamico, P.; Strobaugh, T.; Medina, M.; Gehring, A. Detection of Escherichia coli O157 : H7 using a surface plasmon resonance biosensor. Biotechnol. Tech. 1998, 12(7), 571–576. [Google Scholar]
- Subramanian, A.S.; Irudayaraj, J.M. Surface plasmon resonance based immunosensing of E. coli O157 : H7 in apple juice. Trans. ASABE 2006, 49(4), 1257–1262. [Google Scholar]
- Subramanian, A.; Irudayaraj, J.; Ryan, T. A mixed self-assembled monolayer-based surface plasmon immunosensor for detection of E-coli O157:H7. Biosens. Bioelectron. 2006, 21(7), 998–1006. [Google Scholar]
- Subramanian, A.; Irudayaraj, J.; Ryan, T. Mono and dithiol surfaces on surface plasmon resonance biosensors for detection of Staphylococcus aureus. Sensor. Actuat. B-Chem. 2006, 114(1), 192–198. [Google Scholar]
- Elkins, K.; Metcalf, E. Monoclonal antibodies demonstrate multiple epitopes on the O antigens of Salmonella typhimurium LPS. J. Immunol. 1984, 133(4), 2255–2260. [Google Scholar]
- Gupta, J.; Reed, C. The direct reaction between Salmonella enteritidis endotoxin and antibody. Measurement of 7S and 19S antibody in normal, tolerant and immune sera. J. Immunol. 1967, 98(5), 1093–1103. [Google Scholar]
- Robbins, J.; Kenny, K.; Suter, E. The isolation and biological activities of rabbit gamma M and gamma G- anti-Salmonella typhimurium antibodies. J. Exp. Med. 1965, 122(2), 385–402. [Google Scholar]
- Parker, C.T.; Liebana, E.; Henzler, D.J.; Guard-Petter, J. Lipopolysaccharide O-chain microheterogeneity of Salmonella serotypes Enteritidis and Typhimurium. Environ. Microbiol. 2001, 3(5), 332–342. [Google Scholar]
- Ekgasit, S.; Thammacharoen, C.; Knoll, W. Surface plasmon resonance spectroscopy based on evanescent field treatment. Anal. Chem. 2004, 76(3), 561–568. [Google Scholar]
- Le Minor, L. Bergey's manual of systematic bacteriology; Krieg, N. R., Holt, J. G., Eds.; The Williams & Wilkins Co.: Baltimore, 1984; Vol 1, Salmonella; pp. 427–458. [Google Scholar]
- Chung, J.W.; Bernhardt, R.; Pyun, J. C. Sequential analysis of multiple analytes using a surface plasmon resonance (SPR) biosensor. J. Immunol. Methods 2006, 311(1-2), 178–188. [Google Scholar]
- Shriver-Lake, L.C.; Turner, S.; Taitt, C. R. Rapid detection of Escherichia coli O157:H7 spiked into food matrices. Anal. Chim. Acta. 2007, 584(1), 66–71. [Google Scholar]
- Kim, N.; Park, I.S.; Kim, W.Y. Salmonella detection with a direct-binding optical grating coupler immunosensor. Sensor. Actuat. B-Chem. 2007, 121(2), 606–615. [Google Scholar]
- Zourob, M.; Mohr, S.; Brown, B.J.T.; Fielden, P.R.; McDonnell, M.B.; Goddard, N.J. Bacteria detection using disposable optical leaky waveguide sensors. Biosens. Bioelectron. 2005, 21(2), 293–302. [Google Scholar]
- Alivisatos, P. The use of nanocrystals in biological detection. Nat. Biotechnol. 2004, 22(1), 47–52. [Google Scholar]
- Vengurlekar, A.S.; Ishihara, T. Surface plasmon enhanced photon drag in metal films. Appl. Phys. Lett. 2005, 87(9), 091118. [Google Scholar]

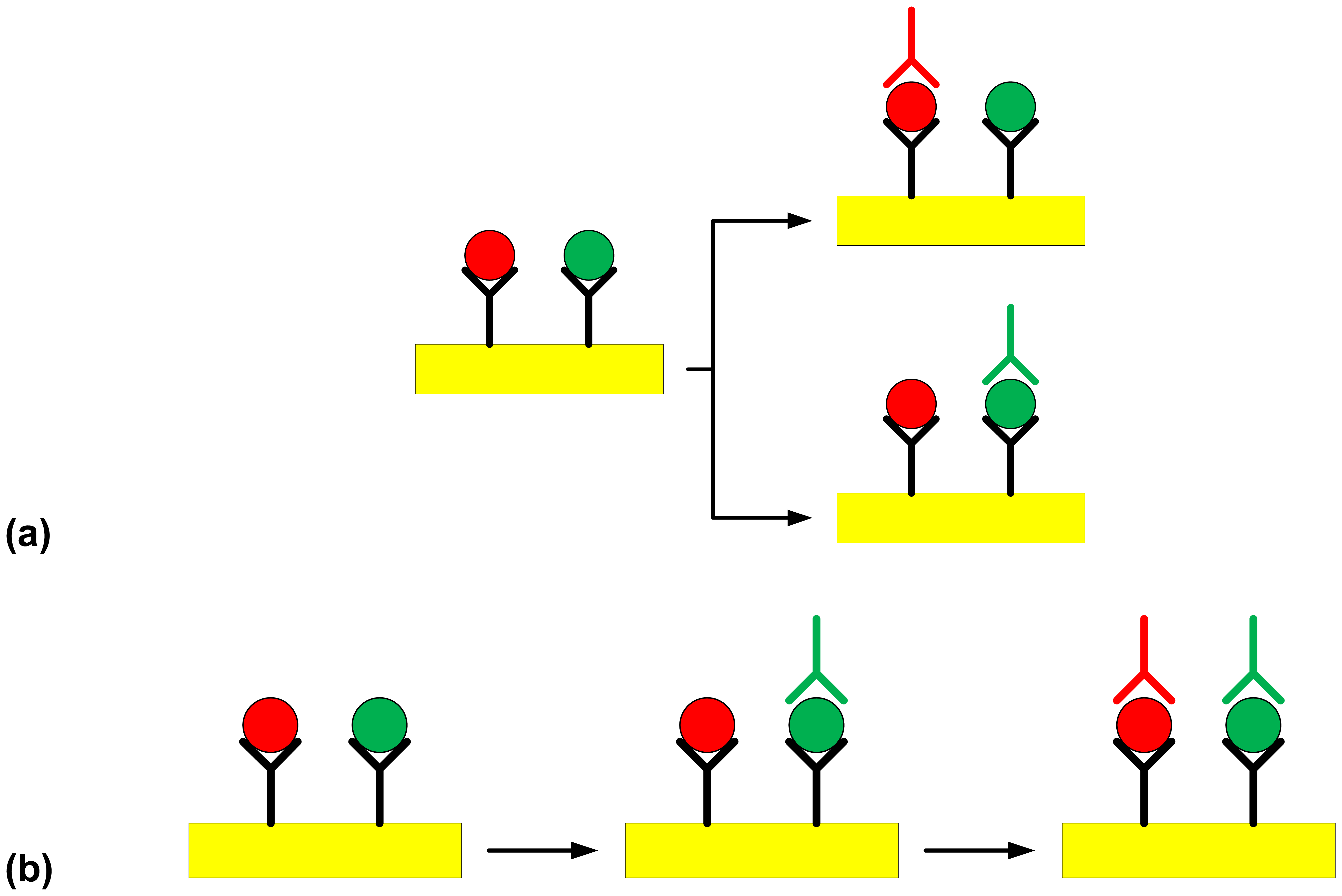
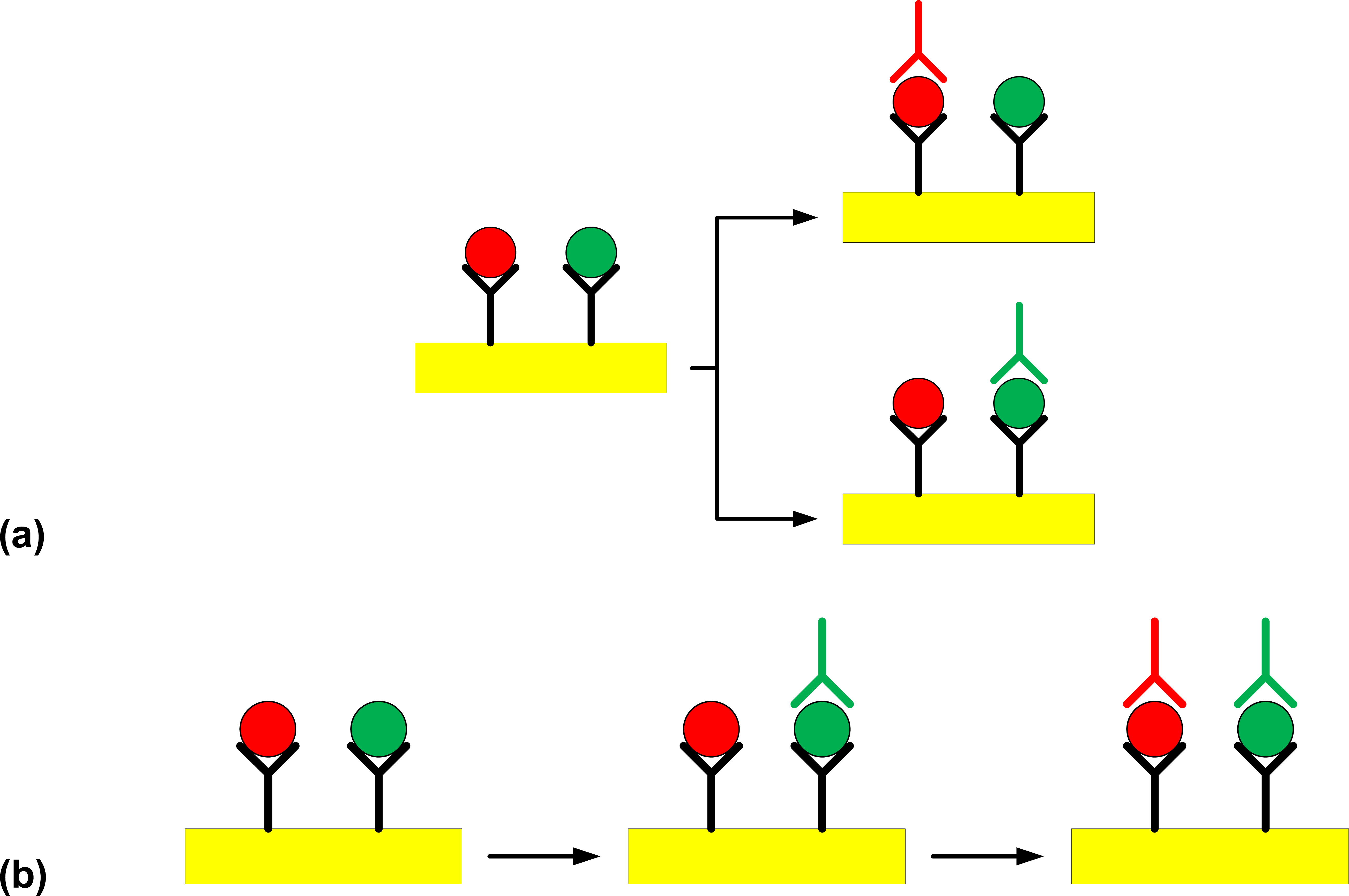
 = S. Typhimurium,
= S. Typhimurium,
 = S. Enteritidis) using the Plasmonic SPR device (
= S. Enteritidis) using the Plasmonic SPR device (
 = C18-modified gold surface, Y = polyclonal capture antibody,
= C18-modified gold surface, Y = polyclonal capture antibody,
 = O:4-specific detection antibody against S. Typhimurium,
= O:4-specific detection antibody against S. Typhimurium,
 = O:9-specific detection antibody against S. Enteritidis). Steps involved in (a) the multi-channel SPR detection of Salmonella serovars present in a sample, (b) the sequential single-channel detection for Salmonella serovars present in a sample.
= S. Typhimurium,
= S. Enteritidis) using the Plasmonic SPR device (
= C18-modified gold surface, Y = polyclonal capture antibody,
= O:4-specific detection antibody against S. Typhimurium,
= O:9-specific detection antibody against S. Enteritidis). Steps involved in (a) the multi-channel SPR detection of Salmonella serovars present in a sample, (b) the sequential single-channel detection for Salmonella serovars present in a sample.
= O:9-specific detection antibody against S. Enteritidis). Steps involved in (a) the multi-channel SPR detection of Salmonella serovars present in a sample, (b) the sequential single-channel detection for Salmonella serovars present in a sample.
= S. Typhimurium,
= S. Enteritidis) using the Plasmonic SPR device (
= C18-modified gold surface, Y = polyclonal capture antibody,
= O:4-specific detection antibody against S. Typhimurium,
= O:9-specific detection antibody against S. Enteritidis). Steps involved in (a) the multi-channel SPR detection of Salmonella serovars present in a sample, (b) the sequential single-channel detection for Salmonella serovars present in a sample.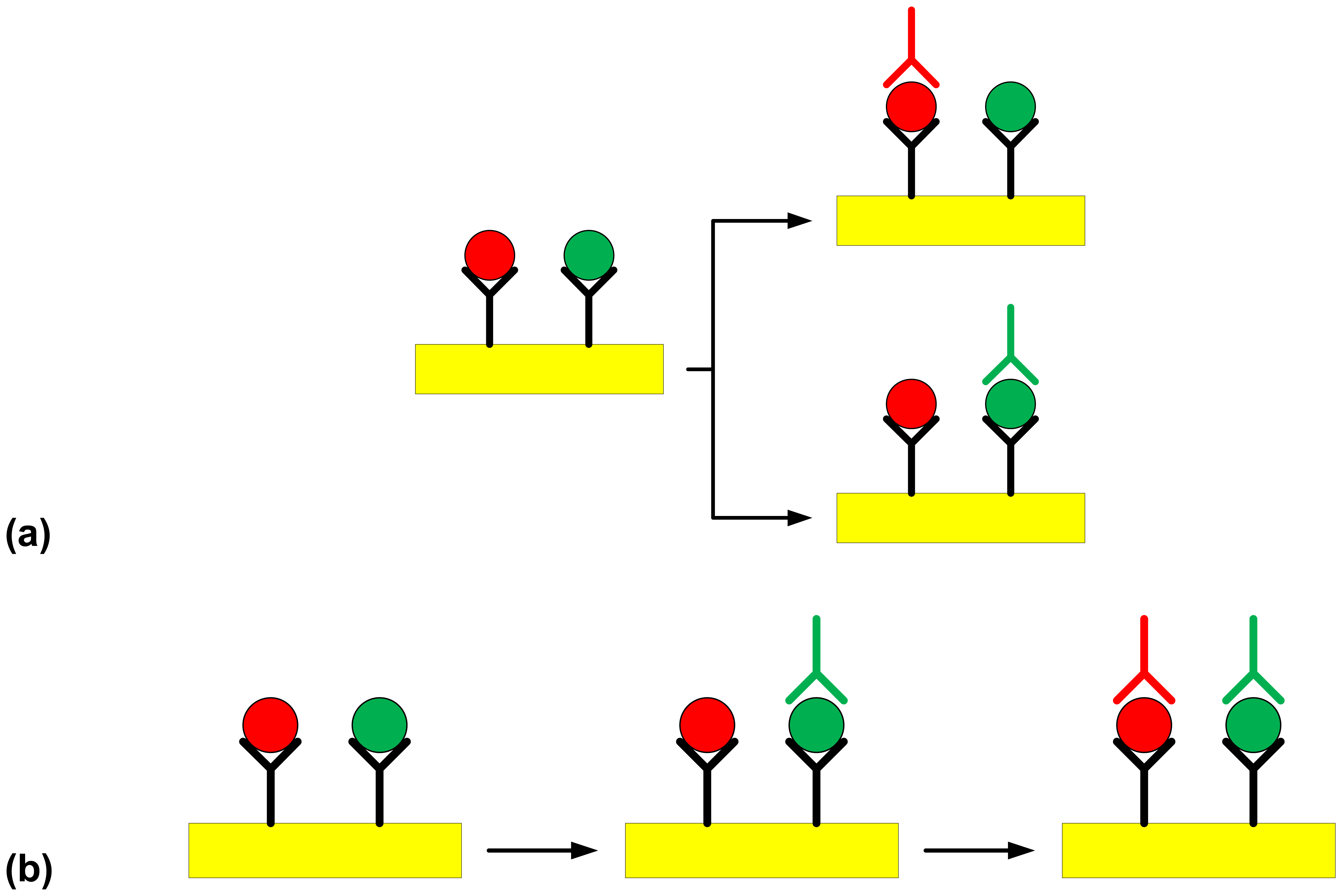
© 2007 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Barlen, B.; Mazumdar, S.D.; Lezrich, O.; Kämpfer, P.; Keusgen, M. Detection of Salmonella by Surface Plasmon Resonance. Sensors 2007, 7, 1427-1446. https://doi.org/10.3390/s7081427
Barlen B, Mazumdar SD, Lezrich O, Kämpfer P, Keusgen M. Detection of Salmonella by Surface Plasmon Resonance. Sensors. 2007; 7(8):1427-1446. https://doi.org/10.3390/s7081427
Chicago/Turabian StyleBarlen, Benjamin, Saikat Datta Mazumdar, Olga Lezrich, Peter Kämpfer, and Michael Keusgen. 2007. "Detection of Salmonella by Surface Plasmon Resonance" Sensors 7, no. 8: 1427-1446. https://doi.org/10.3390/s7081427