Non-invasive Optical Biosensor for Probing Cell Signaling

Biochemical Technologies, Science and Technology Division, Corning Incorporated, Sullivan Park, Corning, NY 14831, USA
Sensors 2007, 7(10), 2316-2329; https://doi.org/10.3390/s7102316
Submission received: 24 September 2007
/
Accepted: 15 October 2007
/
Published: 16 October 2007
(This article belongs to the Special Issue Optical Biosensors)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Cell signaling mediated through a cellular target is encoded by spatial and temporal dynamics of downstream signaling networks. The coupling of temporal dynamics with spatial gradients of signaling activities guides cellular responses upon stimulation. Monitoring the integration of cell signaling in real time, if realized, would provide a new dimension for understanding cell biology and physiology. Optical biosensors including resonant waveguide grating (RWG) biosensor manifest a physiologically relevant and integrated cellular response related to dynamic redistribution of cellular matters, thus providing a non-invasive means for cell signaling study. This paper reviews recent progresses in biosensor instrumentation, and theoretical considerations and potential applications of optical biosensors for whole cell sensing.
1. Introduction
Optical biosensor is a well-established analytical tool for biomolecular interaction analysis [1, 2]. The biosensor typically consists of a biological component, a detector element, and a transducer associated with both components. The biological component is generally a protein target (i.e., bio-receptors) immobilized on the surface of the transducer. Coupled with microfluidics which introduces an analyte as well as separates unbound and bound molecules, the biosensor becomes a powerful biophysical tool for quantitatively characterizing how biomolecular complexes form and dissociate apart over time. However, the ability of optical biosensors to examine living cells was largely unrealized until recent years [3-18]. Advancements in instrumentation, particularly large scale screening platforms [9, 14], enable the discovery of specific types of cellular target-dependent optical signals in cells [9, 15], thus opening a new possibility for whole cell sensing, in particular, cell signaling analysis [18, 19, 20]. This article reviews theoretical, instrumental and experimental progresses of optical biosensor for probing cell signaling.
2. Optical Biosensors
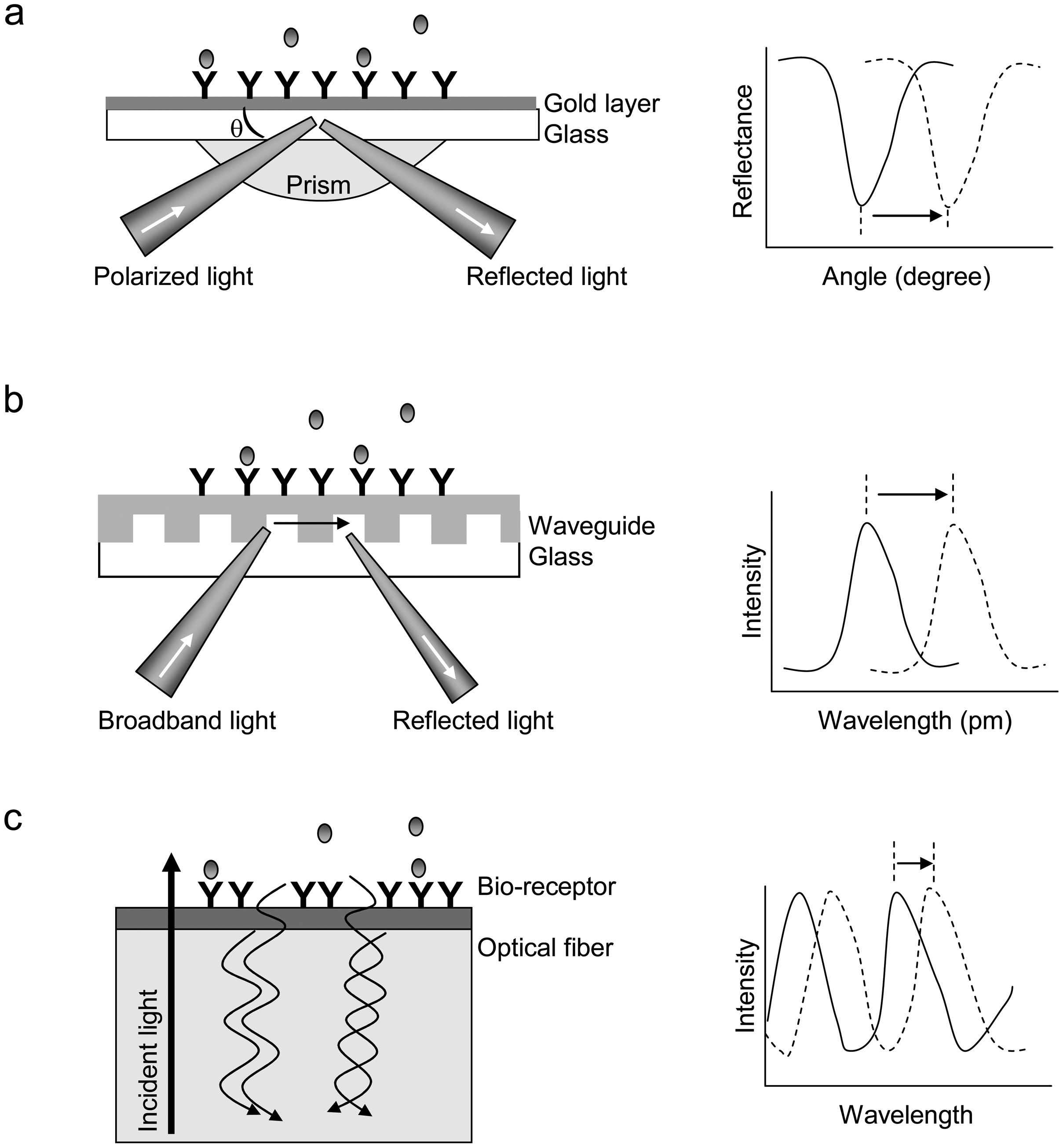
The past decade or so has seen an exponential increase in the number and range of available biosensors based on different optical phenomena. Figure 1 shows operating principles of three types of commercially available optical biosensors: surface plasmon resonance (SPR), and resonant waveguide grating (RWG), and interferometric biosensors.
SPR relies on a prism to direct a wedge of polarized light, covering a range of incident angles, into a planar glass substrate having a gold thin film to excite surface plasmons (Figure 1a). The SPR is a charge-density oscillation or wave that may exist at the interface between two media with dielectric constants of opposite signs (i.e., the gold and a dielectric). The resonant angle at which a minimal in intensity of reflected light occurs is a function of the local refractive index at or near the gold surface [1, 2].
RWG biosensor utilizes the resonant coupling of light into a waveguide by means of a diffraction grating. A polarized light, covering a range of incident wavelengths, is used to illuminate the waveguide; light at specific wavelengths is coupled into and propagate along the waveguide (Figure 1b). The resonant wavelength at which a maximum incoupling efficiency is achieved is a function of the local refractive index at or near the sensor surface [9, 21]. An RWG biosensor typically consists of a thin film (∼75-100nm) of Nb2O5 with a high refractive index (∼2.36) on a glass substrate with an index of ∼1.50 [13].
Interferometry biosensor uses a spectrometer to capture interference patterns in the reflected light from the biosensor interface (Figure 1c). As exampled in this single optical fiber-based interferometric biosensor, a broadband light resource through an optical fiber is directly used to illuminate at the solution-surface interface. The two types of reflected light waves originated from the two reflecting surfaces, the interface with the optical layer and the surface of bio-receptors, can propagate within the fiber and interact each other, leading to an interference pattern. This interference is captured by a spectrometer as a pattern of intensity variation by wavelength with a characteristic profile of peaks and troughs. When there is binding of biological molecules to the biosensor surface, the thickness of bio-receptor layer increases, which in turn leads to a change in the interference pattern at the spectrometer [20].
3. Theories for Living Cell Sensing with Optical Biosensor
Common to all optical biosensors is that they measure changes in local refractive index at or very near the sensor surface. Almost all optical biosensors are applicable for cell sensing in principle – they can employ an evanescent wave to characterize ligand-induced changes in cells. The evanescent-wave is an electromagnetic field, created by the total internal reflection of light at a solution-surface interface, which typically extends a short distance (∼hundreds of nanometers) into the solution with a characteristic depth, termed as penetration depth or sensing volume. This section specifically discusses theoretical considerations for whole cell sensing using RWG biosensor.
Ramsden et al. described a theory to study cell adhesion and spreading on the surface of a RWG biosensor [3]. This theory assumes a 3-layer waveguide system [21], wherein the cells undergo shape changes with well-defined geometrical parameters and steps upon adhesion and spreading on the biosensor surface. Such model allows both the number of cells at the surface and a parameter characterizing their shape to be accurately determined.
Recently, we had developed theoretical and mathematical models that describe the parameters and nature of optical signals measured in living cells in response to stimulation with ligands [13]. These models, based on a 3-layer waveguide system in combination with known cellular biophysics, provide a link from ligand-induced optical signals to several cellular processes mediated through a receptor.
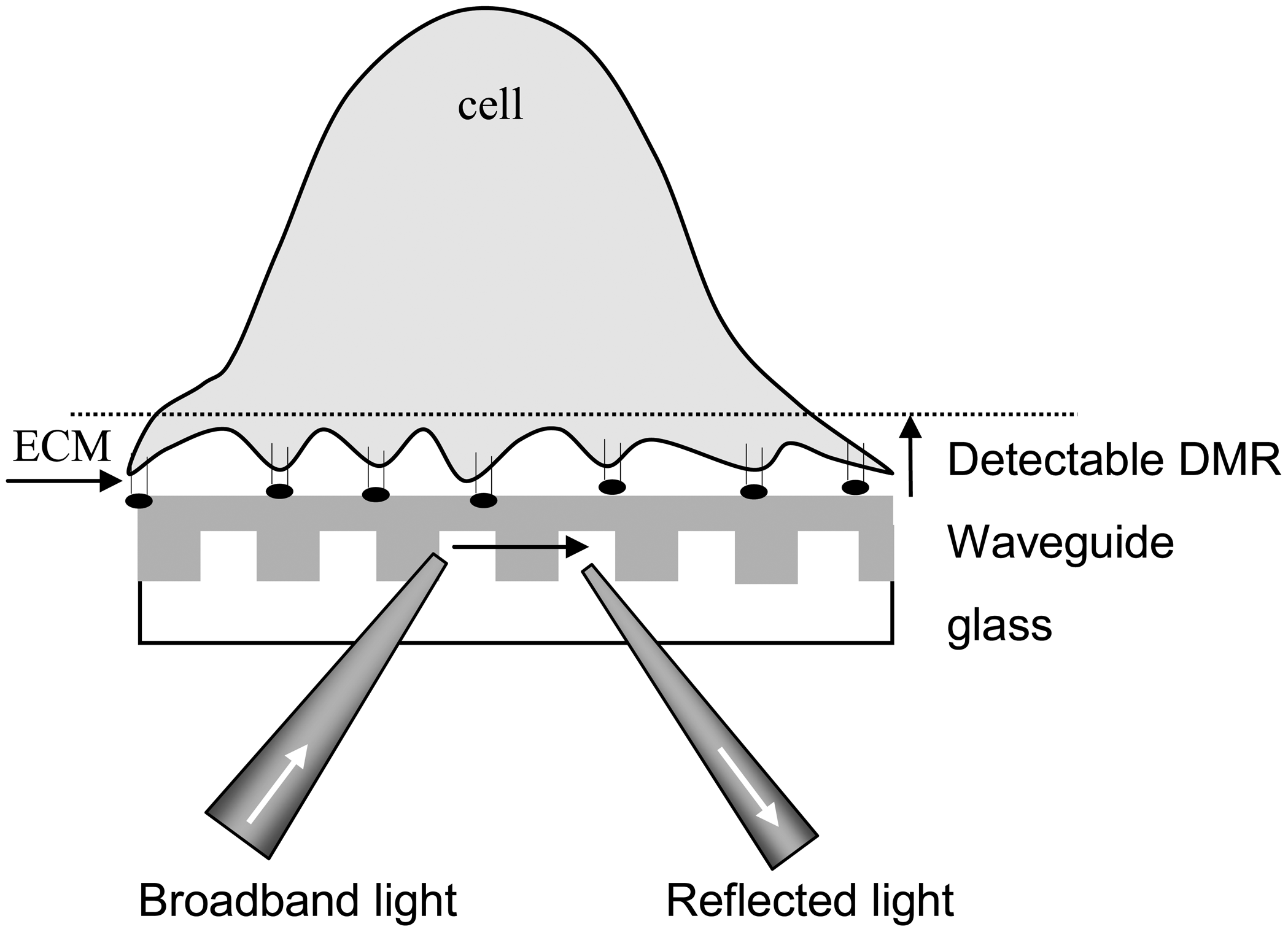
To probe cell signaling, cells are brought to in close contact with the sensor surface, mainly through cell culture. Given that the biosensor measures an averaged response of cells located at the area illuminated by the incident light, a cell layer of highly confluency is used in order to achieve optimal assay results. Because of the large dimension of cells compared to the short penetration depth (∼100-200nm) of a biosensor, the sensor configuration is considered as a non-conventional three-layer system: a substrate, a waveguide film with a grating structure, and a cell layer (Figure 2). Thus, a ligand-induced change in effective refractive index (i.e., the detected signal) is, to first order, directly proportional to the change in refractive index of the bottom portion of cell layer [13]:
where S(C) is the sensitivity to the cell layer, and Δnc the ligand-induced change in local refractive index of the cell layer sensed by the biosensor. Because the refractive index of a given volume within a cell is largely determined by the concentrations of bio-molecules such as proteins [22], Δnc can be assumed to be directly proportional to ligand-induced change in local concentrations of cellular targets or molecular assemblies within the sensing volume. Considering the exponentially decaying nature of the evanescent wave extending away from the sensor surface, the ligand-induced optical signal is governed by [13]:
where ΔZc is the penetration depth into the cell layer, α the specific refraction increment (about 0.18/mL/g for proteins) [23, 24], zi the distance where the mass redistribution occurs, and d an imaginary thickness of a slice within the cell layer. Here the cell layer is divided into an equal-spaced slice in the vertical direction. Eq.2 indicates that the ligand-induced optical signal is a sum of mass redistribution occurring at distinct distances away from the sensor surface, each with an unequal contribution to the overall response. Furthermore, the detected signal, in terms of wavelength or angular shifts, is primarily sensitive to mass redistribution occurring perpendicular to the sensor surface. Because of its dynamic nature, it is also referred to as dynamic mass redistribution (DMR) signal.
4. High Throughput Optical Biosensor Systems
Although there is great difference in operating principle, throughput, sample delivery process and application, most commercially available label free optical systems are configured for bio-molecular interaction analysis at the laboratory scale – the measurements of a small set of samples at one time [20]. For example, Biacore SPR-based systems, the most popular systems, have relative low to medium throughput because limited channels are available for sample analysis, and measurements have to be done in series. However, the applications of optical biosensors in a high throughput screening setting, particularly using living cells, are limited. To address these unmet needs, we have been developing label free optical systems tailoring towards high throughput applications. We are primarily interested in optical systems that are suited for assays using the Society for Biomolecular Sciences (SBS; http://www.sbsonline.org) standard microtiter plates, such as 96well and 384well microplates. The RWG biosensor is chosen, particularly because this type of biosensor with appropriate designs allows lights at nominally normal incident angle to illuminate the waveguide thin film. This is important for illuminating large numbers of biosensors simultaneously.
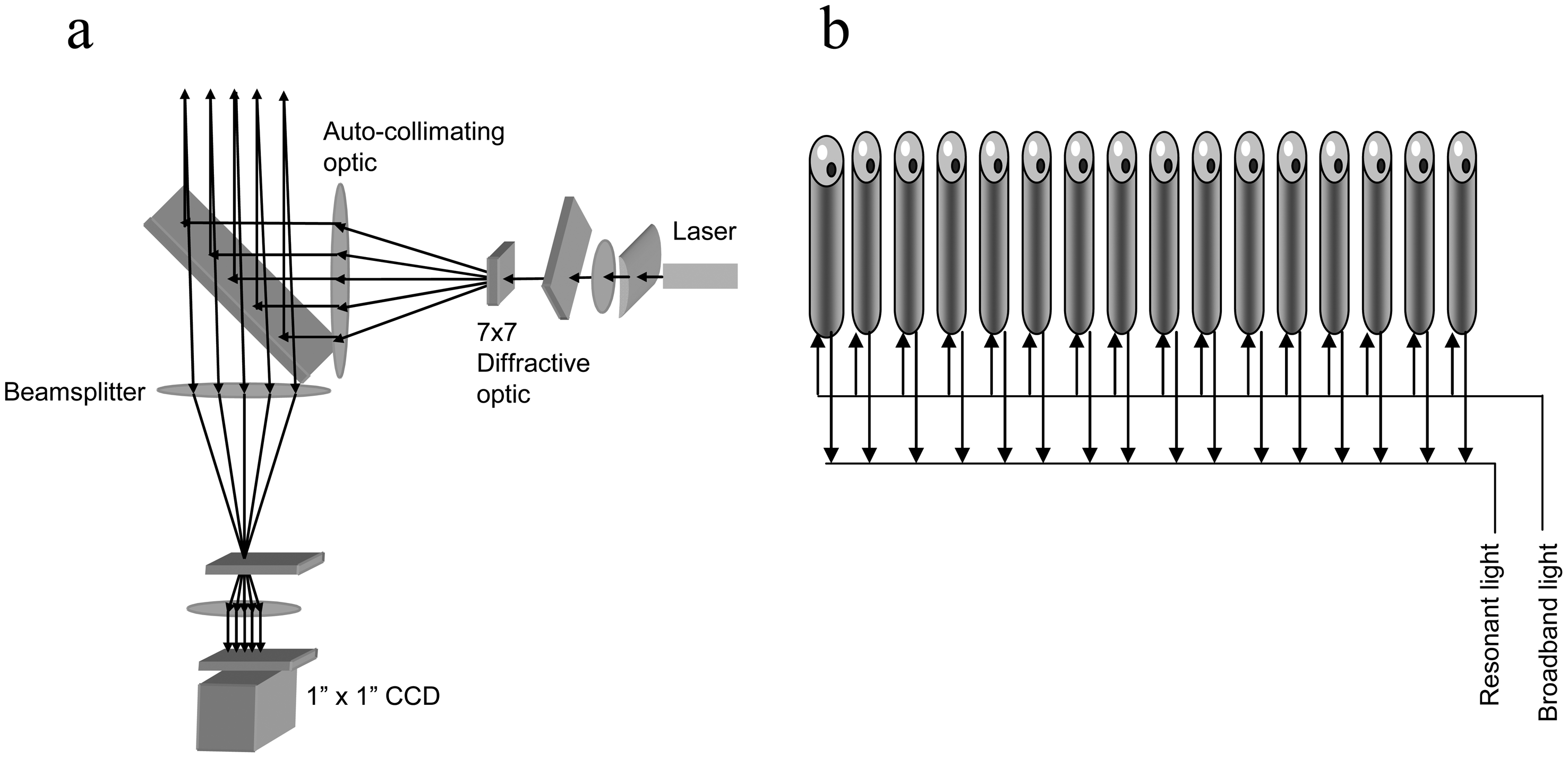
Figure 3 illustrates two types of optical systems developed at our company. In the angular interrogation system (Figure 3a), the total internal reflection is observed at a critical angle of incidence at the interface of the waveguide film and the cell layer. This system consists of a launch system to generate an array of light beams such that each illuminates a RWG biosensor with a dimension of ∼200 μm × 3000 μm, and a CCD camera-based receive system to record changes in resonant angle of the reflected light [9]. The array of light beams is obtained using a beam splitter and diffractive optical lenses. This system allows an array of 7×7 biosensor wells in a 96well microplate to be simultaneously sampled at every 3 seconds. This system does not have any moving parts, except of a liquid handling system for compound addition.
In the wavelength interrogation system (Figure 3b), the total internal reflection is achieved at a resonant wavelength of a broadband light source illuminated at the waveguide. This system, commercialized in 2006, consists of a detector with a temperature-controlled environment, an external liquid handling system, and 384well standard SBS microtiter plates [15]. The detector system is centred on integrated fiber optics to measure any wavelength shift of the reflected lights. A broadband light source, generated through a fiber optic and a collimating lens, is used to illuminate a small region of the biosensor through the bottom of a microplate. A detection fiber for recording the reflected light is bundled with the illumination fiber. A series of illumination/detection heads are arranged in a linear fashion, so that reflection spectra are collected from all 16 wells within the same column of a 384well microplate simultaneously. The whole plate is scanned so that each sensor is addressed multiple times, and each column is addressed in sequence. The temperature-controlling unit is built in to minimize any temperature fluctuation.
5. Probing Cell Signaling with Optical Biosensor
Cells rely on multiple cellular pathways or machineries to process, encode and integrate the information received. Unlike the affinity analysis with optical biosensors that specifically measures the binding of analytes to a protein target, living cells are much more complex and dynamic. This section discusses the cellular mechanisms that provide the basis for cell signaling studies with optical biosensors, as well as the applications of optical biosensor in cell signaling.
5.1 Cell-surface interactions
To study cell signaling, cells are brought to contact with the surface of a biosensor, which can be achieved through cell culture. These cultured cells are attached onto the biosensor surface through three types of contacts: focal contacts, close contacts and extracellular matrix contacts, each with its own characteristic separation distance from the surface [25, 26]. As a result, the basal cell membranes are generally distant away from the surface by ∼10-100nm. Since the penetration depths or sensing volumes of most optical biosensors are in the range of 100-200nm, these biosensors are able to sense the bottom portion of cells.
Cells, in many cases, exhibit surface-dependent adhesion and proliferation. In order to achieve robust cell assays, the biosensor surface may require a coating to enhance cell adhesion and proliferation. On the other hand, the surface properties could have direct impact on cell biology – surface-bound ligands may influence the response of cells [27], while the mechanical compliance of a substrate material, which dictates how it will deform under forces applied by the cell, is influential [28]. Together with the culture conditions (time, serum concentration, confluency, etc.), cellular status obtained may be distinct from one surface to another, and from one condition to another. Thus, special attentions to control cellular status are necessitated for developing biosensor-based cell assays [9, 10, 13].
5.2 Cells under un-stimulated conditions
Cells are dynamic objects with relatively large dimensions – typically tens of microns. Even without stimulation, cells constantly undergo micromotion – a dynamic movement and remodeling of cellular structure, as observed in tissue culture by time lapse microscopy at the sub-cellular resolution, as well as by bio-impedance measurements at the nanometer level [29].
Under un-stimulated conditions the cells generally give rise to an almost net-zero DMR response, as examined with RWG biosensor (Figure 4) [9, 10, 16]. This is partly because of the low spatial resolution of optical biosensors, as determined by the large size of the laser spot and the long propagation length of the coupled light. The size of the laser spot determines the size of the area studied – usually only one analysis point can be tracked at a time. Thus, the biosensor typically measures an averaged response of a large population of cells located at the light incident area. Although cells undergo micromotion at the single cell level, the large populations of cells examined give rise to a net-zero DMR response in average. Furthermore, it is known that intracellular macromolecules are highly organized [11, 30] and spatially restricted to appropriate sites in mammalian cells [31, 32]. The localization of proteins is tightly controlled in order for cells to regulate the specificity and efficiency of proteins interacting with their proper partners, to spatially separate protein activation and deactivation mechanisms, and thus to determine specific cell functions and responses. Thus, under un-stimulated conditions, the local mass density of cells within the sensing volume can reach an equilibrium state, thus leading to a net-zero optical response. It is worthy noting that the cells examined often have been cultured under conventional culture condition for a period of time such that most of the cells have just completed a single cycle of division.
5.3 Cellular responses upon stimulation
Living cells have exquisite abilities to sense and respond to exogenous signals. Cell signaling was originally thought to function via linear routes where an environmental cue would trigger a linear chain of reactions resulting in a single well-defined response. However, amassing evidences show that cellular responses to external stimuli are much more complicated. It has become apparent that the information the cells received is processed and encoded into complex temporal and spatial patterns of phosphorylation and topological relocation of signaling proteins [32, 33]. The spatial and temporal targeting of proteins to appropriate sites is crucial to regulate the specificity and efficiency of protein-protein interactions, thus dictating the timing and intensity of cell signaling and responses. Pivotal cellular decisions, such as cytoskeletal reorganization, cell cycle checkpoints and apoptosis, depend on the precise temporal control and relative spatial distribution of activated signal-transducers [34-36]. Thus, cell signaling mediated through a cellular target such as G protein-coupled receptor (GPCR) typically proceeds in an orderly and regulated manner, and consists of a series of spatial and temporal events, many of which lead to changes in local mass density or redistribution in local cellular matters of cells [18]. These changes or redistribution when occurring within the sensing volume can be followed directly in real time using optical biosensors [13].
5.4 Characteristics of endogenous GPCR DMR signals
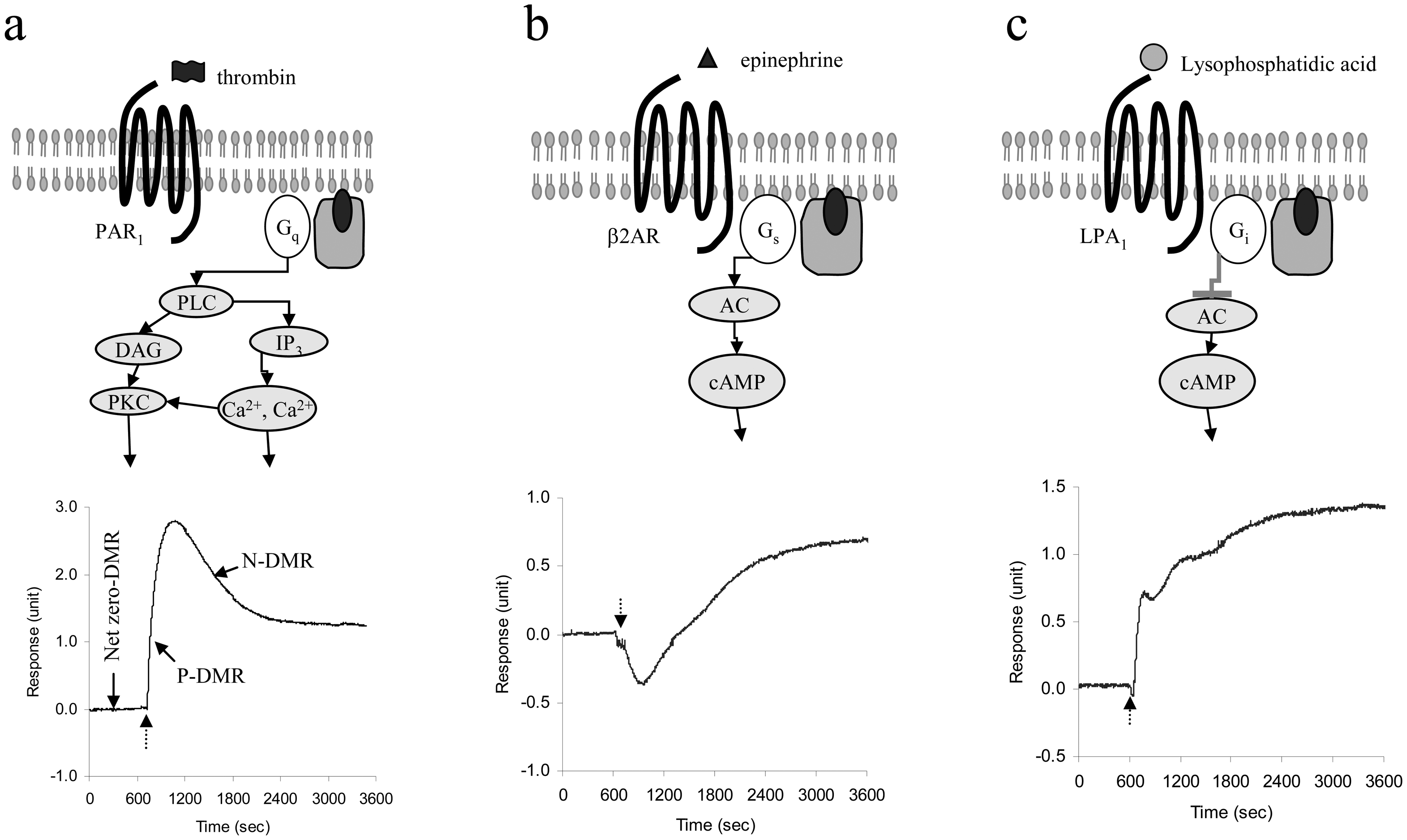
GPCRs represent the largest family of cell membrane receptors [37], and the largest of druggable targets in human genome [38]. Their primary function is to transduce external signals into an array of cellular responses, through both G protein-dependent and independent pathways. Using panels of agonists targeting many GPCRs, three classes of DMR signals in native human epidermoid carcinoma A431 cell were identified [15] (Figure 4). Since each is correlated with the activation of a class of GPCRs depending on G protein with which the receptor is coupled, the DMR signals obtained are named Gq-, Gs- and Gi-DMR signals, respectively. These DMR signals exhibit distinct kinetic and dynamic characteristics, reflecting the unique signaling integration mediated through different classes of GPCRs. Interestingly, the Gq-type DMR signal appears to be rapid, whereas the Gs-type DMR signal is comparatively slow.
To date, we have found that both the Gq and Gs-DMR signals are quite universal across multiple cell lines tested, while the Gi-DMR signals exhibit strong dependence on the cellular context as well as the receptors. However, for the same class of DMR signals mediated through the activation of different receptors in the same cell line or the activation of the same receptor in different cell lines, there are often noticeable differences in fine features such as the amplitudes and kinetics, reflecting the important roles of cellular context in GPCR signaling as well as the uniqueness in signaling of each receptor. The amplitudes of DMR signals for several endogenous Gq-coupled receptors in A431 cells were found to be correlated well with their corresponding Ca2+ mobilization signals, suggesting that a ligand-induced DMR signal is a function of the expression level of its corresponding receptor, as well as the coupling efficiency of the receptor with its G proteins [15, 16].
5.5 Cellular mechanisms of GPCR ligand-induced DMR signals
Historically, optical biosensors have found little use in whole cell sensing. The primary reason is that any optical responses of cells measured with a label free biosensor are often considered as non-specific. In our view, understanding the nature of ligand-induced optical signals is crucial for cell sensing. In the recent years, we have adopted several approaches including chemical biology, cell biology and biophysics approaches to study the cellular mechanisms accounted for ligand-induced DMR signals.
Recent chemical biology and orthogonal fluorescence imaging studies suggest that the Gq-DMR signal is downstream of Ca2+ mobilization [16], whereas the Gs-DMR signal is downstream of cAMP accumulation [17; and unpublished data]. Following classical receptor biology, a numerical analysis suggests that protein translocation and receptor internalization are two primary resources for the Gq-DMR signal [13]. Central to Gq-coupled receptor signaling is the rapid and dynamic translocation of its signaling components, including protein kinase C, GPCR kinases, β-arrestin, phosphatidylinositol phosphate-binding proteins, and diacylglycerol-binding proteins, to name but a few [39].
On the other hand, for the Gs-DMR signal, the increase in cell adhesion is the major contributor to the P-DMR event, as shown in the epinephrine-induced responses in A431 cells [17; unpublished data]. Epinephrine is a natural agonist for β2 adrenergic receptors (β2ARs) endogenously expressed in A431. Here the P-DMR event refers to the DMR phase with an increased signal, while the N-DMR refers to the phase with a decreased signal. The occurrence of the initial N-DMR event reflects the fact that the majority of downstream signaling components involved in the β2AR signaling complexes, with the exception (thus far) of A-kinase anchoring proteins and β-arrestins, are already compartmentalized at or near the cell membrane [40-42]. Thus, for Gs-coupled receptor the recruitment of intracellular targets to the activated receptors is much less pronounced than Gq-coupled receptor, and is overwhelmed by other cellular events leading to an initial decrease in local mass density in cells.
5.6 Applications of optical biosensors in cell signaling
Optical biosensors can follow receptor signaling in real time, and offer a kinetic optical signal in a ligand-dependent manner. A ligand-induced DMR signal is an integrated and quantifiable response of living cells, containing contributions of many downstream signaling events. Chemical biology studies, based on the modulation profiles of various modulators on a ligand-induced DMR signal, suggest that a DMR signal can be treated as a unique readout for systems cell biology of receptor signaling [18]. Such approach has been applied to map the signaling pathways and network interaction of EGF receptor (EGFR) [9, 13], as well as to study the cellular functions of cholesterol in quiescent A431 cells [12]. Results showed that the EGF-induced DMR signal is saturable to EGF concentration, and sensitive to the activity of EGFR tyrosine kinase, MEK, and dynamin [9]. These optical biosensor cell assays link the EGF-induced DMR signal of quiescent A431 cells mainly to the Ras/mitogen-activated protein kinase pathway, which primarily proceeds through MEK and leads to cell detachment [43].
GPCR signaling is sophisticated – a receptor may couple simultaneously to more than one G protein subtype, and may interact with other signaling molecules and scaffolding proteins such as arrestins [36]. The DMR signal imitates the complexity of GPCR signaling, as evidenced by bradykinin B2 receptor signaling in A431. Bradykinin, a vasoactive nonapeptide, is implicated in the regulation of many physiological and pathophysiological responses, including vascular permeability, inflammation, pain and neurotransmitter release [44]. Endogenous bradykinin B2 receptor in A431 cells is mitogenic or anti-mitogenic, dependent on the cellular states as well as the doses of bradykinin [10, 45]. In proliferating states, low doses (<100nM) of bradykinin trigger Gs-mediated signaling, while high doses (>100nM) of bradykinin favors Gq-signaling. On the other hand, bradykinin between 0.5nM and about 100nM mediated both Gs and Gq-pathways in quiescent A431 cells obtained with 0.1% fetal bovine serum for about 20hours [10]. As summarized in Figure 5, chemical biology study showed that in those quiescent cells, blockage of protein kinase C activity only partially attenuates the initial P-DMR, but potentiates the retarded event, as indicated by the broken arrows in Figure 5. Conversely, blockage of protein kinase A activity increases the initial P-DMR event, but suppresses the retarded event. These results suggest that the activation of B2 receptors leads to both Gq and Gs signaling, and both pathways negatively cross-regulated each other [10, 45].
DMR signal is a novel physiological response of living cells, and can be used to study ligand pharmacology. When a ligand is specific to a receptor expressed in a cell system, the ligand-induced DMR signal is also receptor-specific, dose-dependent and saturable [9, 10, 15, 16]. Furthermore, the DMR signal exhibits expected desensitization patterns [16]. Desensitization and re-sensitization is common to almost all GPCRs [46]. Figure 6 compares the desensitization patterns of A431 cells in response to repeated stimulation with distinct combinations of two agonists obtained using the biosensor with those measured with Ca2+ mobilization assays. Results showed that both technologies lead to almost identical desensitization patterns. Furthermore, the DMR signal also maintains the fidelity of GPCR ligands, similar to those obtained using conventional technologies. The biosensor can distinguish full agonists, partial agonists, inverse agonists, antagonists, and allosteric modulators [10, 16, 17; unpublished data].
6. Conclusion
The ability of examining living cells is vital to understand the biological functions of cellular targets, and to the success of drug discovery and development. Although more complex and less specific than biomolecular interaction analysis, cell-based assays show superior ability to facilitate the measurements of mode of action, pathway activation, toxicity, and phenotypic responses of cells induced by exogenous stimuli. Together with the commercial availability of high throughput instruments, the newly discovered capability for studying cell signaling will accelerate wide acceptance of optical biosensors in both fundamental researches and drug discovery processes, beyond biomolecular interaction analysis.
References
- Leatherbarrow, R.J.; Edwards, P.R. Analysis of molecular recognition using optical biosensors. Curr. Opin. Chem. Biol. 1999, 3, 544–547. [Google Scholar]
- Cooper, M.A. Optical biosensors in drug discovery. Nature Rev. Drug Discov. 2002, 1, 515–528. [Google Scholar]
- Ramsden, J.J.; Li, S.-Y.; Prenosil, J.E.; Heinzle, E. Kinetics of adhesion and spreading of animan cells. Biotechnol. Bioeng. 1994, 43, 939–945. [Google Scholar]
- Giebel, K.-F.; Bechinger, C.; Herminghaus, S.; Riedel, M.; Leiderer, P.; Weiland, U.; Bastmeyer, M. Imaging of cell/substrate contacts of living cells with surface plasmon resonance microscopy. Biophys. J. 1999, 76, 509–516. [Google Scholar]
- Horath, R.; Pedersen, H.C.; Skivesen, N.; Selmeczi, D.; Larsen, N.B. Monitoring of living cell attachment and spreading using reverse symmetry waveguide sensing. Appl. Phys. Lett. 2005, 86, e071101. [Google Scholar]
- Quinn, J.G.; O'Neill, S.; Doyle, A.; McAtamney, C.; Diamond, D.; MacCraith, B.D.; O'Kennedy, R. Development and application of surface plasmon resonance-based biosensors for the detection of cell-ligand interactions. Anal. Biochem. 2000, 281, 135–143. [Google Scholar]
- Hide, M.; Tsutsui, T.; Sato, H.; Nishimura, T.; Morimoto, K.; Yamamoto, S.; Yoshizato, K. Real-time analysis of ligand-induced cell surface and intracellular reactions of living mast cells using a surface plasmon resonance-based biosensor. Anal. Biochem. 2002, 302, 28–37. [Google Scholar]
- Voros, J.; Graf, R.; Kenausis, G.L.; Bruinink, A.; Mayer, J.; Textor, M.; Wintermantel, E.; Spencer, N.D. Feasibility study of an online toxicological sensor based on the optical waveguide technique. Biosensors & Bioelectronics 2000, 15, 423–429. [Google Scholar]
- Fang, Y.; Ferrie, A.M.; Fontaine, N.H.; Yuen, P.K. Characteristics of dynamic mass redistribution of EGF receptor signaling in living cells measured with label free optical biosensors. Anal. Chem. 2005, 77, 5720–5725. [Google Scholar]
- Fang, Y.; Li, G.; Peng, J. Optical biosensor provides insights for bradykinin B2 receptor signaling in A431 cells. FEBS Lett. 2005, 579, 6365–6374. [Google Scholar]
- Fang, Y.; Ferrie, A.M.; Li, G. Probing cytoskeleton modulation by optical biosensors. FEBS Lett. 2005, 579, 4175–4180. [Google Scholar]
- Fang, Y.; Ferrie, A.M.; Li, G.; Fontaine, N.H. Cellular functions of cholesterol probed with optical biosensors”. Biochim. Biophys. Acta 2006, 1763, 254–261. [Google Scholar]
- Fang, Y.; Ferrie, A.M.; Fontaine, N.H.; Mauro, J.; Balakrishnan, J. Resonant waveguide grating biosensor for living cell sensing. Biophys. J. 2006, 91, 1925–1940. [Google Scholar]
- Li, G.; Ferrie, A.M.; Fang, Y. Label-free profiling of endogenous G protein-coupled receptors using a cell-based high throughput screening technology. J. Assoc. Lab. Automat. 2006, 11, 181–187. [Google Scholar]
- Fang, Y.; Li, G.; Ferrie, A.M. Non-invasive optical biosensor for assaying endogenous G protein-coupled receptors in adherent cells. J. Pharmacol. Toxicol. Methods 2007, 55, 314–322. [Google Scholar]
- Fang, Y.; Ferrie, A.M. Optical biosensor differentiates signaling of endogenous PAR1 and PAR2 in A431 cells. BMC Cell Biol. 2007, 8, 24:1–24:12. [Google Scholar]
- Fang, Y.; Ferrie, A.M.; Lahiri, J. Systems biology and pharmacology of β2 adrenergic receptors in A431. In Trends in Signal Transduction Research; Meyers, J.N., Ed.; Nova Science Publishers Inc.: New York, 2007; pp. 145–171. [Google Scholar]
- Fang, Y. Label-free cell-based assays with optical biosensors in drug discovery. Assays and Drug Development Technologies 2006, 4, 583–595. [Google Scholar]
- Cooper, M.A. Optical biosensors: where next and how soon. Drug Discov. Today 2006, 11, 1061–1067. [Google Scholar]
- Rich, R.L.; Myszka, D.G. Higher-throughput, label-free, real-time molecular interaction analysis. Anal. Biochem. 2007, 361, 1–6. [Google Scholar]
- Tiefenthaler, K.; Lukosz, W. Sensitivity of grating couplers as integrated-optical chemical sensors. J. Opt. Soc. Am. B 1989, 6, 209–220. [Google Scholar]
- Barer, R.; Joseph, S. Refractometry of living cells. Part I. basic principles. Quart. J. Microsc. Science 1954, 95, 399–423. [Google Scholar]
- De Feijter, J.A.; Benjamins, J.; Veer, F.A. Ellipsometry as a tool to study the adsorption behavior of synthetic and biopolymers at the air-water interface. Biopolymers 1978, 17, 1759–1772. [Google Scholar]
- Brandenburg, A.; Krauter, R.; Kunzel, C.; Stefan, M.; Schulte, H. Interferometric sensor for detection of surface-bound bioreactions. Appl. Optics 2000, 39, 6396–6405. [Google Scholar]
- Burridge, K.; Fath, K.; Kelly, T.; Nuckolls, G.; Turner, C. Focal adhesions: transmembrane junctions between the extracellular matrix and the cytoskeleton. Annu. Rev. Cell Biol. 1988, 4, 487–525. [Google Scholar]
- Chen, W.T.; Singer, S.J. Immunoelectron microscopic studies of the sites of cell-substratum and cell-cell contacts in cultured fibroblasts. J. Cell Boil. 1982, 95, 205–222. [Google Scholar]
- Discher, D.E.; Janmey, P.; Wang, Y. Tissue cells feel and respond to the stiffness of their substrate. Science 2005, 310, 1139–1143. [Google Scholar]
- Kato, M.; Mrksich, M. Rewiring cell adhesion. J. Am. Chem. Soc. 2006, 126, 6504–6505. [Google Scholar]
- Giaever, I.; Keese, C.R. Micromotion of mammalian cells measured electrically. Proc. Natl. Acad. Sci. USA 1991, 88, 7896–7900. [Google Scholar]
- Hudder, A.; Nathanson, L.; Deutscher, M.P. Organization of mammalian cytoplasm. Mol. Cell Biol. 2003, 23, 9318–9326. [Google Scholar]
- Sato, T.K.; Overduin, M.; Emr, S.D. Location, location, location: membrane targeting directed by PX domains. Science 2001, 294, 1881–1885. [Google Scholar]
- Kholodenko, B.N. Four-dimensional organization of protein kinase signaling cascades: the roles of diffusion, endocytosis and molecular motors. J. Exper. Biol. 2003, 206, 2073–2082. [Google Scholar]
- Aldridge, B.B.; Burke, J.M.; Lauffenburger, D.A.; Sorger, P.K. Physicochemical modelling of cell signalling pathways. Nat. Cell Biol. 2006, 8, 1195–1203. [Google Scholar]
- Wiley, H.S. Trafficking of the ErbB receptors and its influence on signaling. Exper. Cell Res. 2003, 284, 78–88. [Google Scholar]
- Schnapp, B.J. Trafficking of signaling modules by kinesin motors. J. Cell Sci. 2003, 116, 2125–2135. [Google Scholar]
- Lefkowitz, R.J.; Whalen, E.J. β-arrestins: traffic cops of cell signaling. Curr. Opinion Cell Biol. 2004, 16, 162–168. [Google Scholar]
- Fredriksson, R.; Schioth, H.B. The repertoire of G-protein-coupled receptors in fully sequenced genomes. Mol. Pharmacol. 2005, 67, 1414–1425. [Google Scholar]
- Fang, Y.; Lahiri, J.; Picard, L. G protein-coupled receptor microarrays for drug discovery. Drug Discov. Today 2003, 8, 755–761. [Google Scholar]
- van Baal, J.; de Widt, J.; Divecha, N.; van Blitterswijk, W.J. Translocation of diacylglycerol kinase θ from cytosol to plasma membrane in response to activation of G protein-coupled receptors and protein kinase C. J. Biol. Chem. 2005, 280, 9870–9878. [Google Scholar]
- Degerman, E.; Belfrage, P.; Manganiello, V.C. Structure, localization, and regulation of cGMP-inhibited phosphodiesterase (PDE3). J. Biol. Chem. 1997, 272, 6823–6826. [Google Scholar]
- Fan, G.; Shumay, E.; Wang, H.; Malbon, C.C. The scaffold protein gravin (cAMP-dependent protein kinase-anchoring protein 250) binds the β2-adrenergic receptor via the receptor cytoplasmic Arg-329 to Leu-413 domain and provides a mobile scaffold during desensitization. J. Biol. Chem. 2001, 276, 24005–24014. [Google Scholar]
- Lin, F.; Wang, H.-Y.; Malbon, C.C. Gravin-mediated formation of signaling complexes in β2-adrenergic receptor desensitization and resensitization. J. Biol. Chem. 2000, 275, 19025–19034. [Google Scholar]
- Lu, Z.; Jiang, G.; Blume-Jensen, P.; Hunter, T. Epidermal growth factor-induced tumor cell invasion and metastasis initiated by dephosphorylation and downregulation of focal adhesion kinase. Mol. Cell Biol. 2001, 21, 4016–4031. [Google Scholar]
- Marceau, F.; Regoli, D. Bradykinin receptor ligands: therapeutic perspectives. Nat. Rev. Drug Discov. 2004, 3, 845–852. [Google Scholar]
- Liebmann, C.; Graness, A.; Ludwig, B.; Adomeit, A.; Boehmer, A.; Boehmer, F.-D.; Nürnberg, B.; Wetzker, R. Dual bradykinin B2 receptor signalling in A431 human epidermoid carcinoma cells: activation of protein kinase C Is counteracted by a Gs-mediated stimulation of the cyclic AMP pathway. Biochem. J. 1996, 313, 109–118. [Google Scholar]
- Ferguson, S.S.G. Evolving concepts in G protein-coupled receptor endocytosis: the role in receptor desensitization and signaling. Pharmacol. Rev. 2001, 53, 1–24. [Google Scholar]
Figure 1.
Principles of three types of optical biosensors for biomolecular interaction analysis. (a) SPR. The receptor (Y) is covalently coupled to the derivatized surface of a thin layer of gold, which is deposited onto a glass substrate. The light incident on the gold layer is directed by a prism. The reflectance of the gold surface is a function of incident angle. The resonance angle shifts when target molecules (
![Sensors 07 02316i1]() ) in solution bind to the immobilized receptors. (b) RWG biosensor. The coupling of incident light into the waveguide is achieved by the diffraction grating. The intensity of reflected light is a function of the resonant wavelength. The binding of target molecules in a sample to the immobilized receptors results in a shift in the resonant wavelength. (c) Interferometric biosensor. A broadband light resource is directly incident at the solution-surface interface. Two types of reflected light waves originated from the two reflecting surfaces, the interface with the optical layer and the surface of bio-receptors, interact each other, leading to interference patterns. The binding of biomolecules to the bio-receptors alters the inference pattern.
) in solution bind to the immobilized receptors. (b) RWG biosensor. The coupling of incident light into the waveguide is achieved by the diffraction grating. The intensity of reflected light is a function of the resonant wavelength. The binding of target molecules in a sample to the immobilized receptors results in a shift in the resonant wavelength. (c) Interferometric biosensor. A broadband light resource is directly incident at the solution-surface interface. Two types of reflected light waves originated from the two reflecting surfaces, the interface with the optical layer and the surface of bio-receptors, interact each other, leading to interference patterns. The binding of biomolecules to the bio-receptors alters the inference pattern.
 ) in solution bind to the immobilized receptors. (b) RWG biosensor. The coupling of incident light into the waveguide is achieved by the diffraction grating. The intensity of reflected light is a function of the resonant wavelength. The binding of target molecules in a sample to the immobilized receptors results in a shift in the resonant wavelength. (c) Interferometric biosensor. A broadband light resource is directly incident at the solution-surface interface. Two types of reflected light waves originated from the two reflecting surfaces, the interface with the optical layer and the surface of bio-receptors, interact each other, leading to interference patterns. The binding of biomolecules to the bio-receptors alters the inference pattern.
) in solution bind to the immobilized receptors. (b) RWG biosensor. The coupling of incident light into the waveguide is achieved by the diffraction grating. The intensity of reflected light is a function of the resonant wavelength. The binding of target molecules in a sample to the immobilized receptors results in a shift in the resonant wavelength. (c) Interferometric biosensor. A broadband light resource is directly incident at the solution-surface interface. Two types of reflected light waves originated from the two reflecting surfaces, the interface with the optical layer and the surface of bio-receptors, interact each other, leading to interference patterns. The binding of biomolecules to the bio-receptors alters the inference pattern.
Figure 1.
Principles of three types of optical biosensors for biomolecular interaction analysis. (a) SPR. The receptor (Y) is covalently coupled to the derivatized surface of a thin layer of gold, which is deposited onto a glass substrate. The light incident on the gold layer is directed by a prism. The reflectance of the gold surface is a function of incident angle. The resonance angle shifts when target molecules (
![Sensors 07 02316i1]() ) in solution bind to the immobilized receptors. (b) RWG biosensor. The coupling of incident light into the waveguide is achieved by the diffraction grating. The intensity of reflected light is a function of the resonant wavelength. The binding of target molecules in a sample to the immobilized receptors results in a shift in the resonant wavelength. (c) Interferometric biosensor. A broadband light resource is directly incident at the solution-surface interface. Two types of reflected light waves originated from the two reflecting surfaces, the interface with the optical layer and the surface of bio-receptors, interact each other, leading to interference patterns. The binding of biomolecules to the bio-receptors alters the inference pattern.
) in solution bind to the immobilized receptors. (b) RWG biosensor. The coupling of incident light into the waveguide is achieved by the diffraction grating. The intensity of reflected light is a function of the resonant wavelength. The binding of target molecules in a sample to the immobilized receptors results in a shift in the resonant wavelength. (c) Interferometric biosensor. A broadband light resource is directly incident at the solution-surface interface. Two types of reflected light waves originated from the two reflecting surfaces, the interface with the optical layer and the surface of bio-receptors, interact each other, leading to interference patterns. The binding of biomolecules to the bio-receptors alters the inference pattern.
) in solution bind to the immobilized receptors. (b) RWG biosensor. The coupling of incident light into the waveguide is achieved by the diffraction grating. The intensity of reflected light is a function of the resonant wavelength. The binding of target molecules in a sample to the immobilized receptors results in a shift in the resonant wavelength. (c) Interferometric biosensor. A broadband light resource is directly incident at the solution-surface interface. Two types of reflected light waves originated from the two reflecting surfaces, the interface with the optical layer and the surface of bio-receptors, interact each other, leading to interference patterns. The binding of biomolecules to the bio-receptors alters the inference pattern.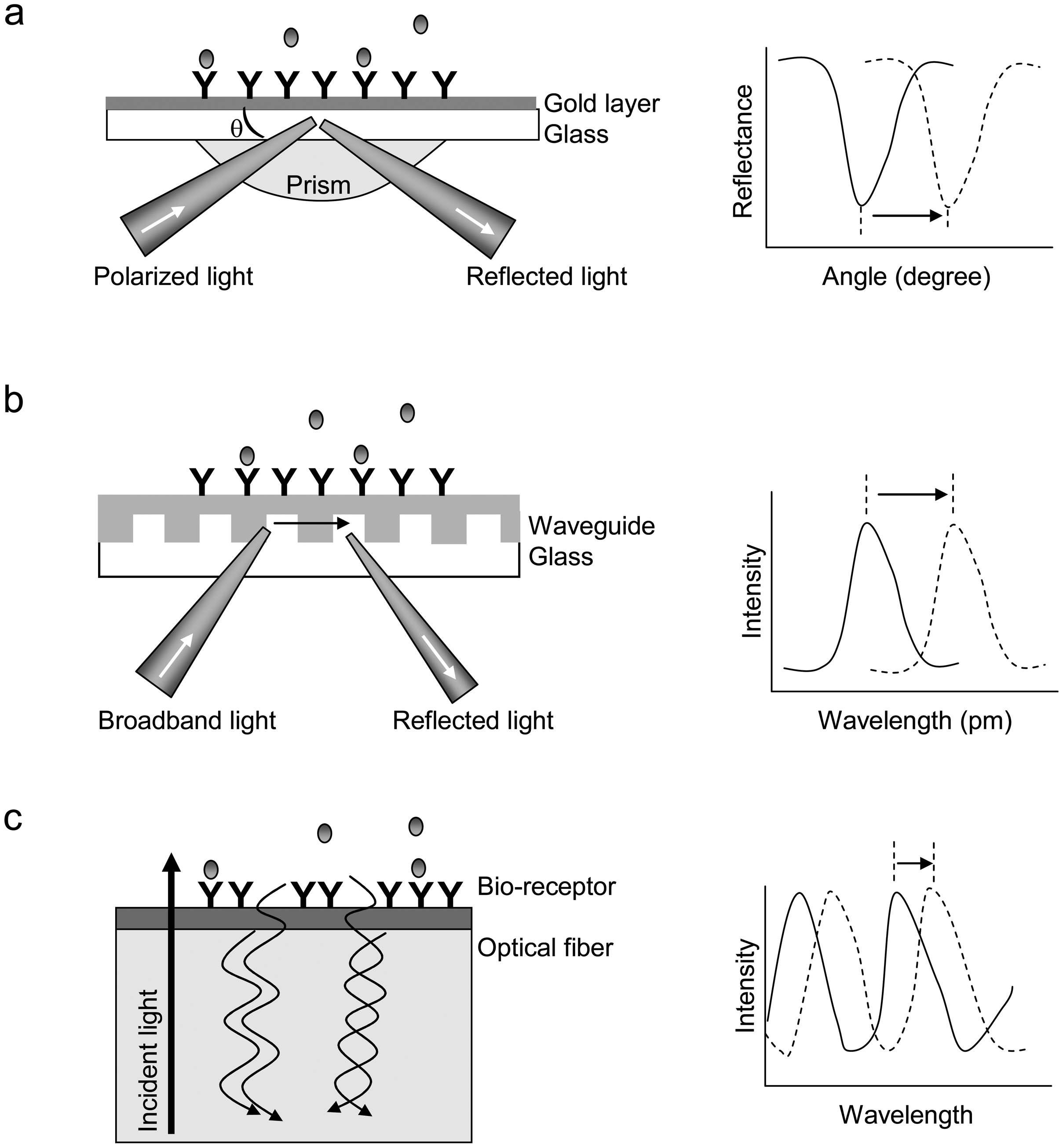
Figure 2.
A three-layer waveguide system for detecting ligand-induced dynamic mass redistribution signals in living cells using RWG biosensor. Cells are anchored to the sensor surface through interaction with the extracellular matrix (ECM).
Figure 2.
A three-layer waveguide system for detecting ligand-induced dynamic mass redistribution signals in living cells using RWG biosensor. Cells are anchored to the sensor surface through interaction with the extracellular matrix (ECM).
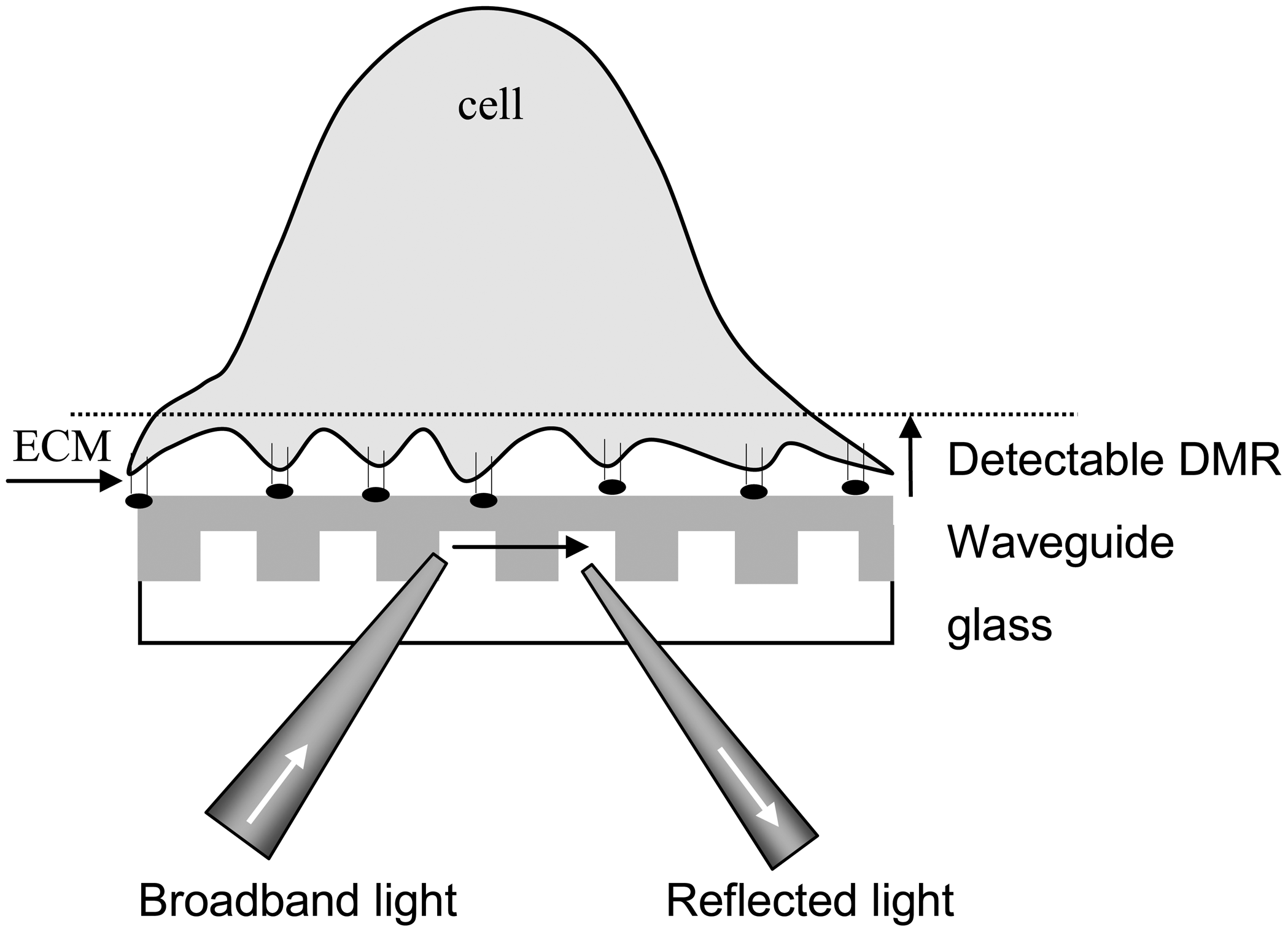
Figure 3.
Two detection schemes of RWG biosensors for high throughput applications. (a) Angular interrogation system. A laser light is split into an array of 7x7 light beams, such that each illuminates a biosensor located within one well of 96well SBS standard microplate. The reflected lights are received with a CCD camera as an image of 7x7 resonant bands. (b) Wavelength interrogation system. A linear array of 16 illumination/detection heads is used to scan a 384well microplate, one column at a time. In each bundled head, there is an optical fiber to send a laser light to illuminate a biosensor located in each well, and a second optical fiber to receive the reflected light.
Figure 3.
Two detection schemes of RWG biosensors for high throughput applications. (a) Angular interrogation system. A laser light is split into an array of 7x7 light beams, such that each illuminates a biosensor located within one well of 96well SBS standard microplate. The reflected lights are received with a CCD camera as an image of 7x7 resonant bands. (b) Wavelength interrogation system. A linear array of 16 illumination/detection heads is used to scan a 384well microplate, one column at a time. In each bundled head, there is an optical fiber to send a laser light to illuminate a biosensor located in each well, and a second optical fiber to receive the reflected light.
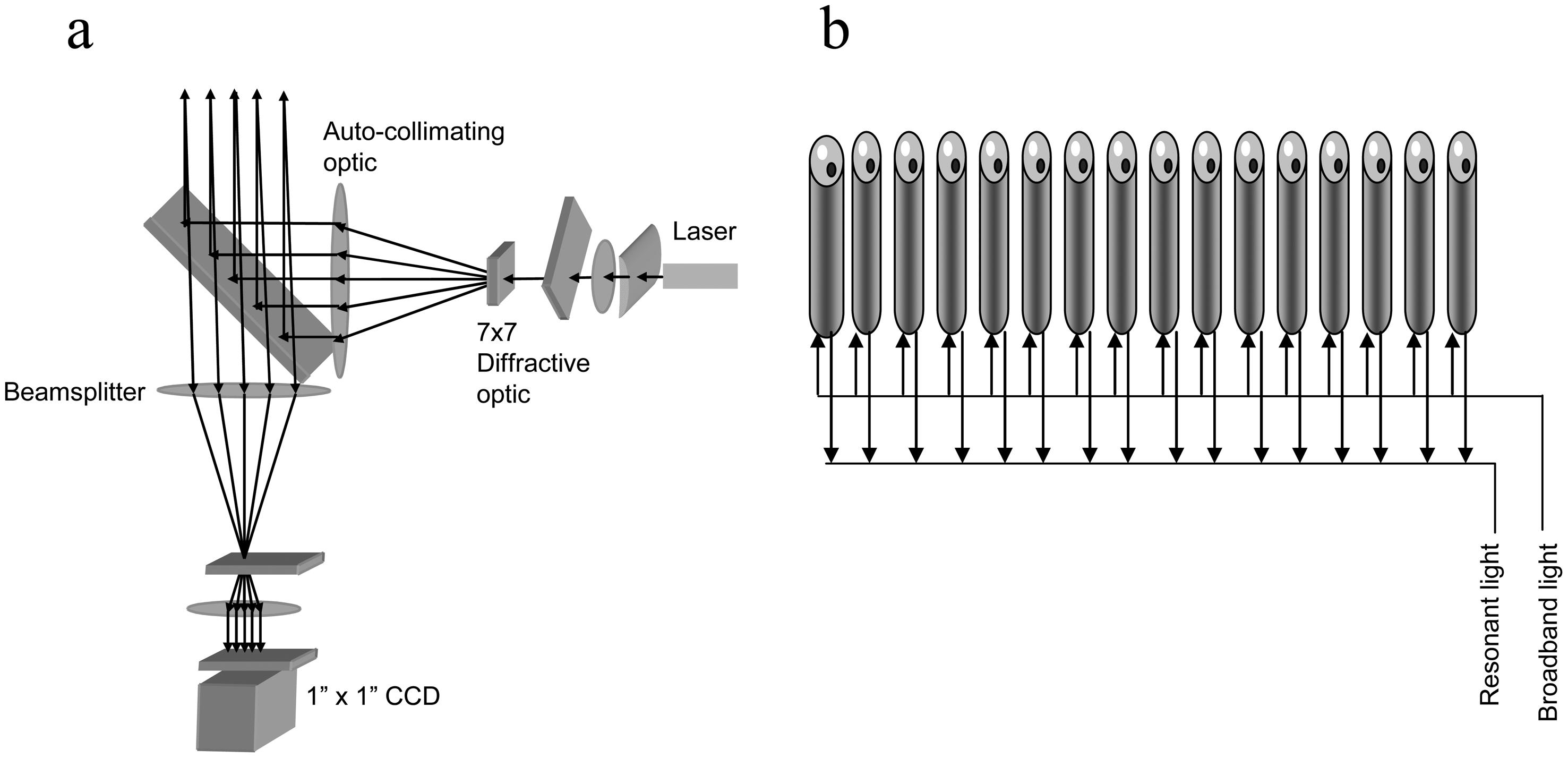
Figure 4.
GPCR signaling and three major classes of DMR signals in living cells using RWG biosensor. (a) Gq-signaling and its DMR signal (protease activated receptor subtype 1 in Chinese hamster ovary cells, 10 unit/ml thrombin). (b) Gs-signaling and its DMR signal (β2-adrenergic receptors in A431 cells, 2nM epinephrine). (c) Gi-signaling and its DMR signal (lysophophatidic acid (LPA) receptor subtype 1 in A431, 100nM LPA). The broken arrows indicate the time when an agonist solution is introduced. Data was generated using the angular interrogation system. 1 unit equals to 100 pm shift in resonant wavelength. Figure was reproduced with permission from Ref. 15.
Figure 4.
GPCR signaling and three major classes of DMR signals in living cells using RWG biosensor. (a) Gq-signaling and its DMR signal (protease activated receptor subtype 1 in Chinese hamster ovary cells, 10 unit/ml thrombin). (b) Gs-signaling and its DMR signal (β2-adrenergic receptors in A431 cells, 2nM epinephrine). (c) Gi-signaling and its DMR signal (lysophophatidic acid (LPA) receptor subtype 1 in A431, 100nM LPA). The broken arrows indicate the time when an agonist solution is introduced. Data was generated using the angular interrogation system. 1 unit equals to 100 pm shift in resonant wavelength. Figure was reproduced with permission from Ref. 15.
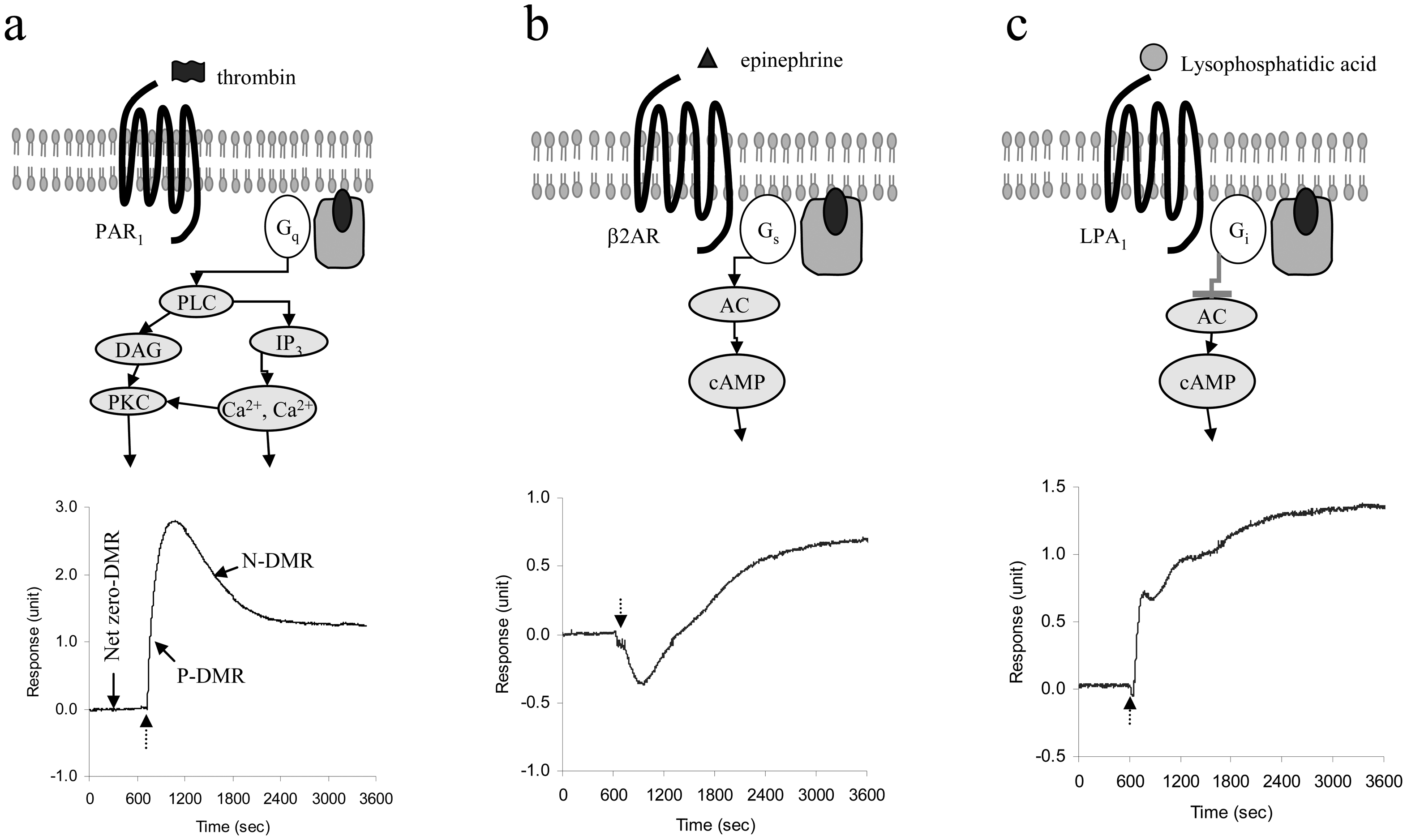
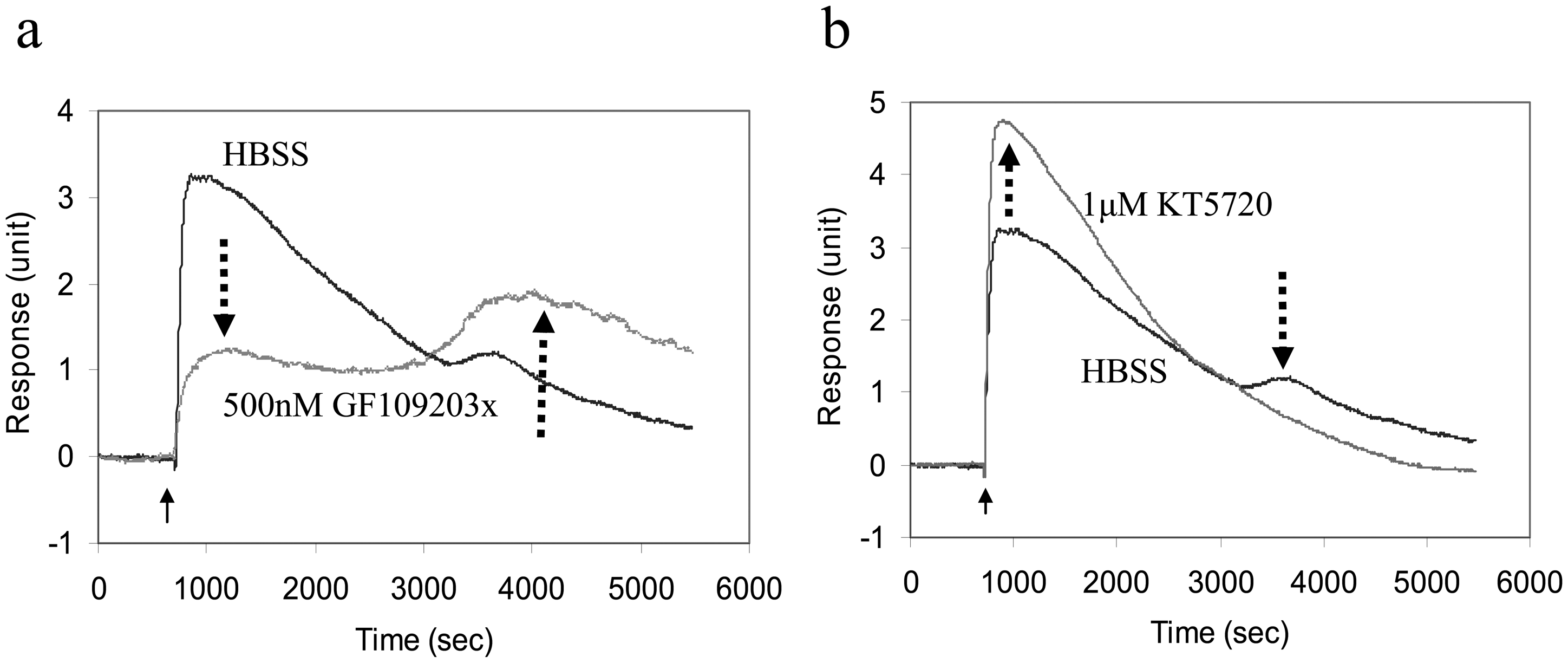
Figure 5.
Chemical intervention reveals dual signaling of bradykinin B2 receptor in A431 cells. The bradykinin-induced DMR responses of cells pre-treated with the buffer only (HBSS) were in comparison with those pretreated with 500nM GF109203x (a protein kinase C inhibitor) (a), or 1 μM KT5720 (a protein kinase B inhibitor) (b). The bradykinin concentration was 16nM, and the cells were starved with 0.1% fetal bovine serum medium for 18hrs before assays. The solid arrows indicate the time when the agonist solution is introduced. Data was reproduced with permission from ref. 10.
Figure 5.
Chemical intervention reveals dual signaling of bradykinin B2 receptor in A431 cells. The bradykinin-induced DMR responses of cells pre-treated with the buffer only (HBSS) were in comparison with those pretreated with 500nM GF109203x (a protein kinase C inhibitor) (a), or 1 μM KT5720 (a protein kinase B inhibitor) (b). The bradykinin concentration was 16nM, and the cells were starved with 0.1% fetal bovine serum medium for 18hrs before assays. The solid arrows indicate the time when the agonist solution is introduced. Data was reproduced with permission from ref. 10.
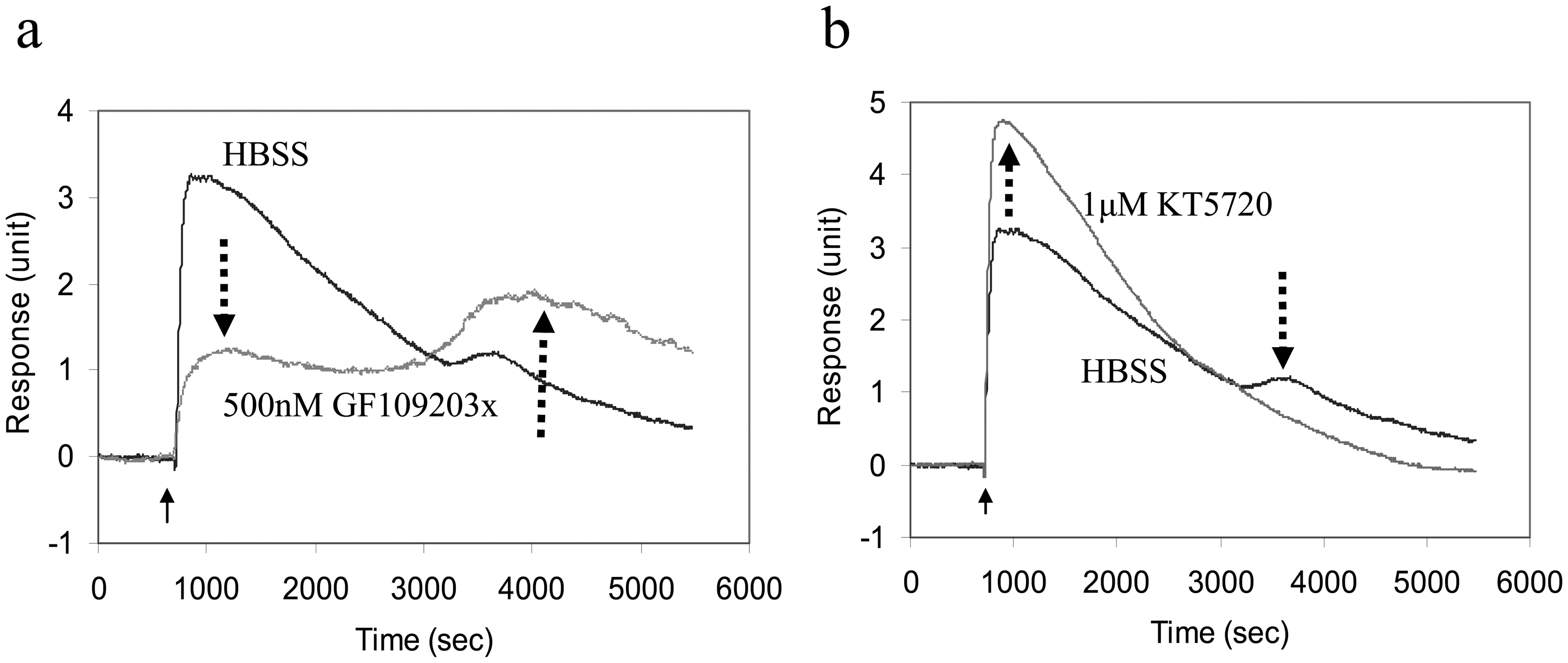
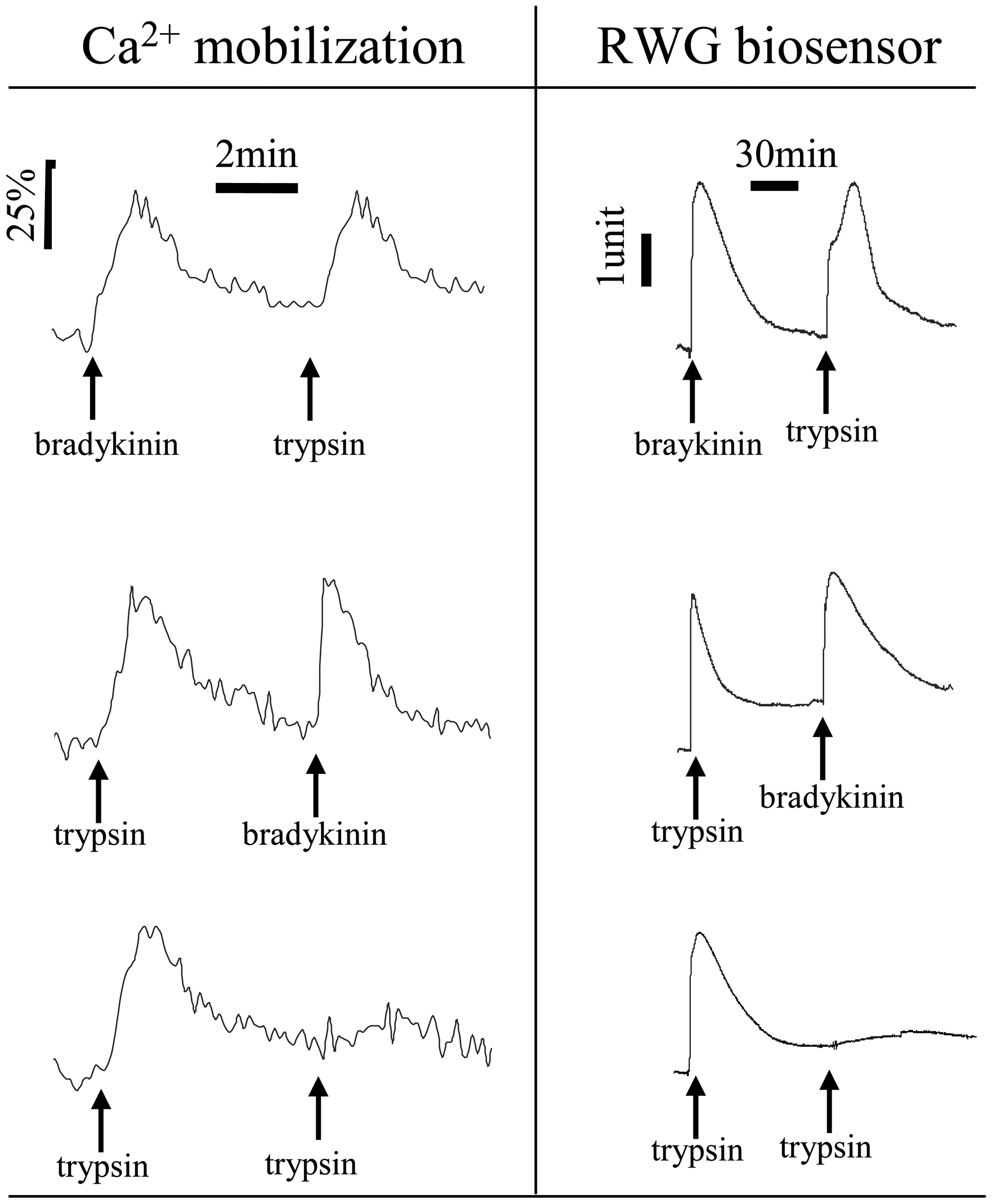
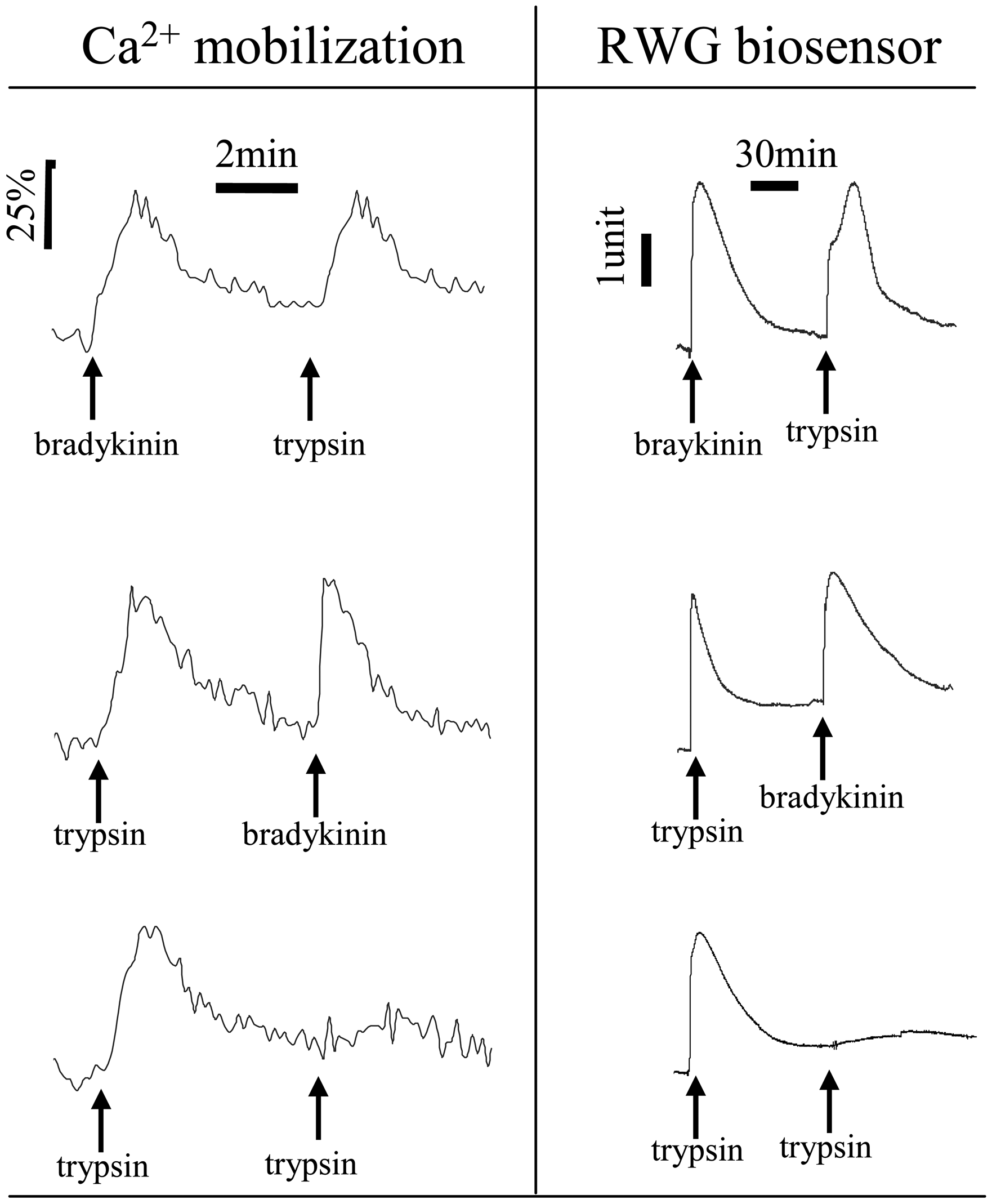
Figure 6.
GPCR agonist-induced DMR signals follow classical desensitization patterns, similar to those measured with Ca2+ mobilization assays. A431 cells responded to repeated stimulation with distinct combinations of two agonists: bradykinin and trypsin, for endogenous B2 receptor and protease activated receptor subtype 2 (PAR2), respectively. Data was reproduced with permission from ref. 16.
Figure 6.
GPCR agonist-induced DMR signals follow classical desensitization patterns, similar to those measured with Ca2+ mobilization assays. A431 cells responded to repeated stimulation with distinct combinations of two agonists: bradykinin and trypsin, for endogenous B2 receptor and protease activated receptor subtype 2 (PAR2), respectively. Data was reproduced with permission from ref. 16.
© 2007 by MDPI ( http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
MDPI and ACS Style
Fang, Y. Non-invasive Optical Biosensor for Probing Cell Signaling. Sensors 2007, 7, 2316-2329. https://doi.org/10.3390/s7102316
AMA Style
Fang Y. Non-invasive Optical Biosensor for Probing Cell Signaling. Sensors. 2007; 7(10):2316-2329. https://doi.org/10.3390/s7102316
Chicago/Turabian StyleFang, Ye. 2007. "Non-invasive Optical Biosensor for Probing Cell Signaling" Sensors 7, no. 10: 2316-2329. https://doi.org/10.3390/s7102316