
Involvement of Acylated Homoserine Lactones (AHLs) of Aeromonas sobria in Spoilage of Refrigerated Turbot (Scophthalmus maximus L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Bacterial Strains
2.2. Isolation and Identification of Bacterial Strains
2.3. AHL Detection of the Isolated Strain
2.4. Extraction of AHLs
2.5. AHL-Production Kinetics
2.6. Effect of Several Conditions on AHL Production in AS7
2.7. AHL Identification via Gas Chromatography-Mass Spectrometry (GC-MS)
2.8. Siderophore Assay
2.9. Fish Spoilage Assays
3. Results and Discussion
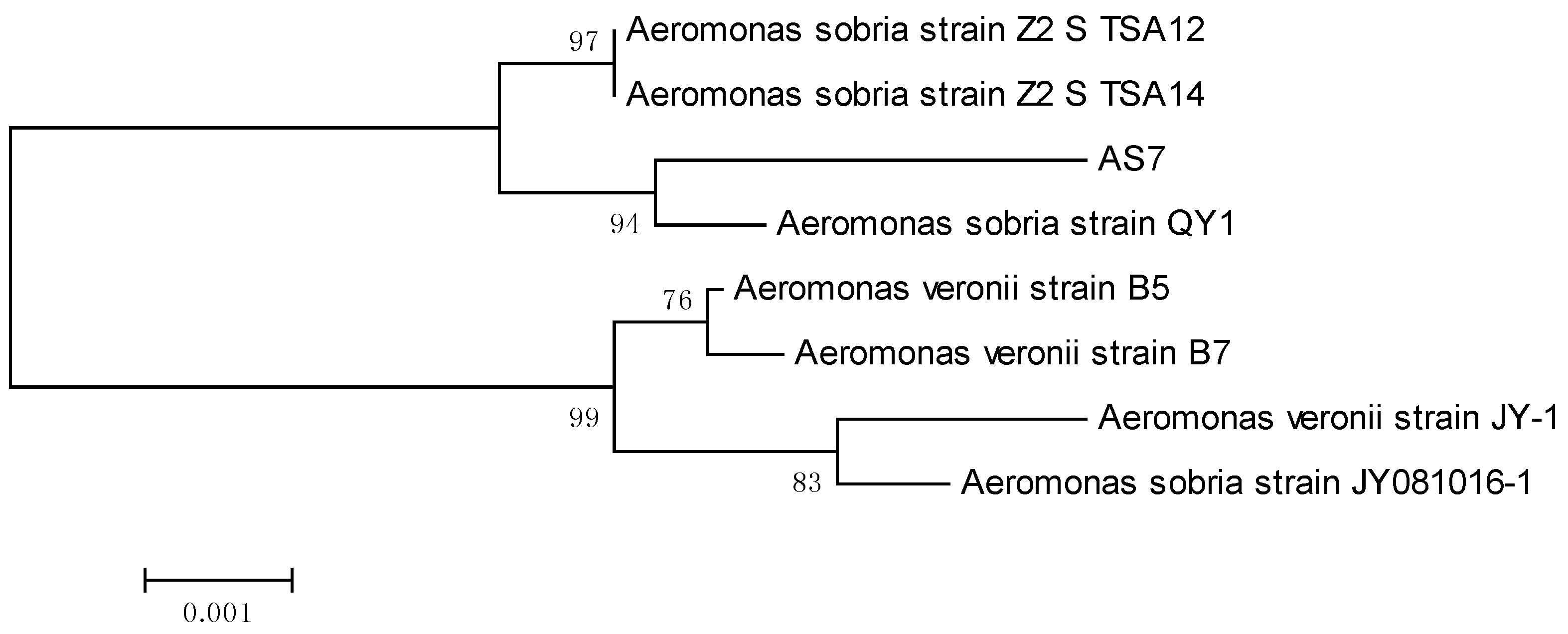
3.1. Identification of Bacteria Isolates
3.2. Detection of AHLs
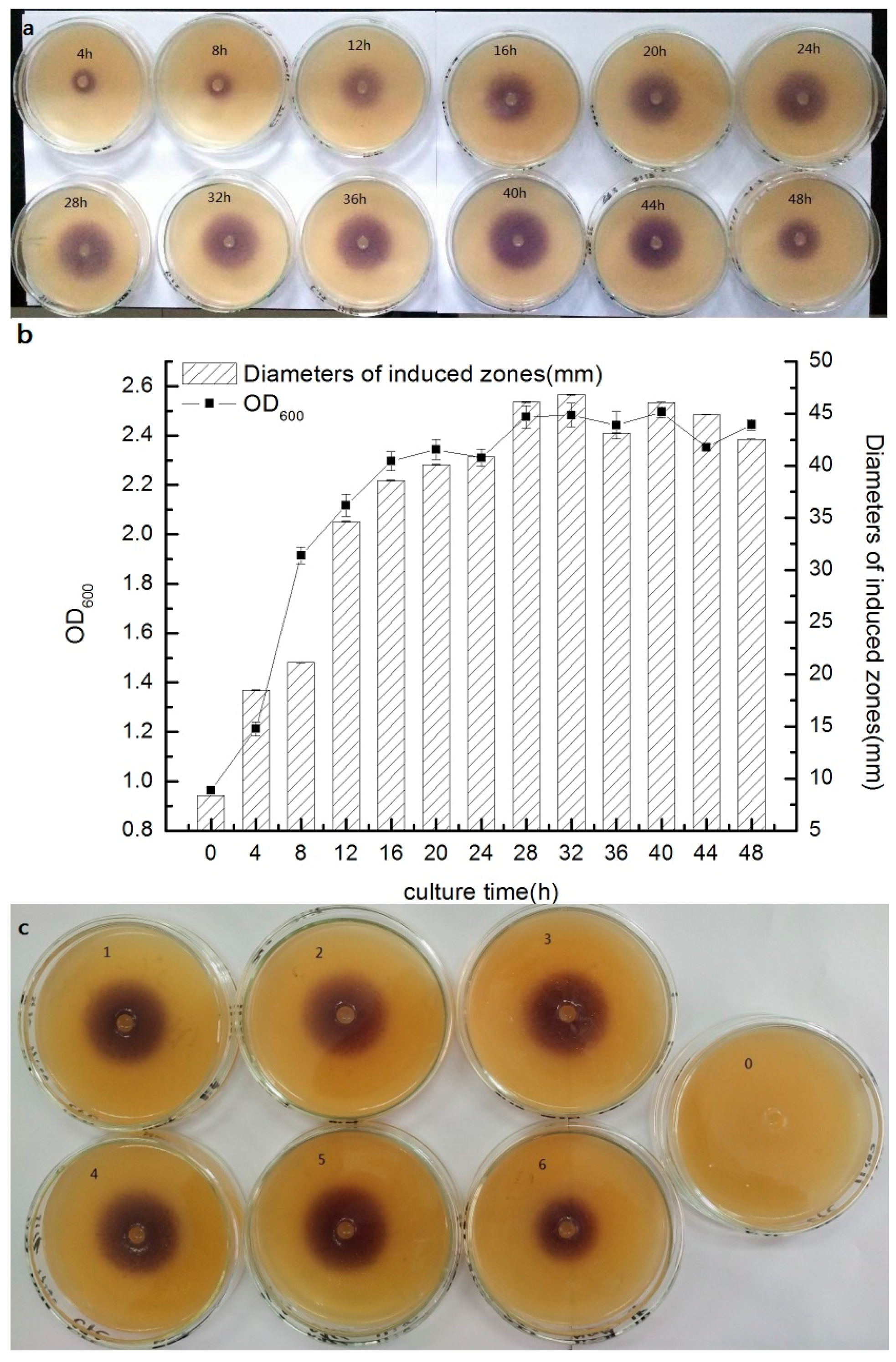
3.3. Short Chain AHL Production Kinetics
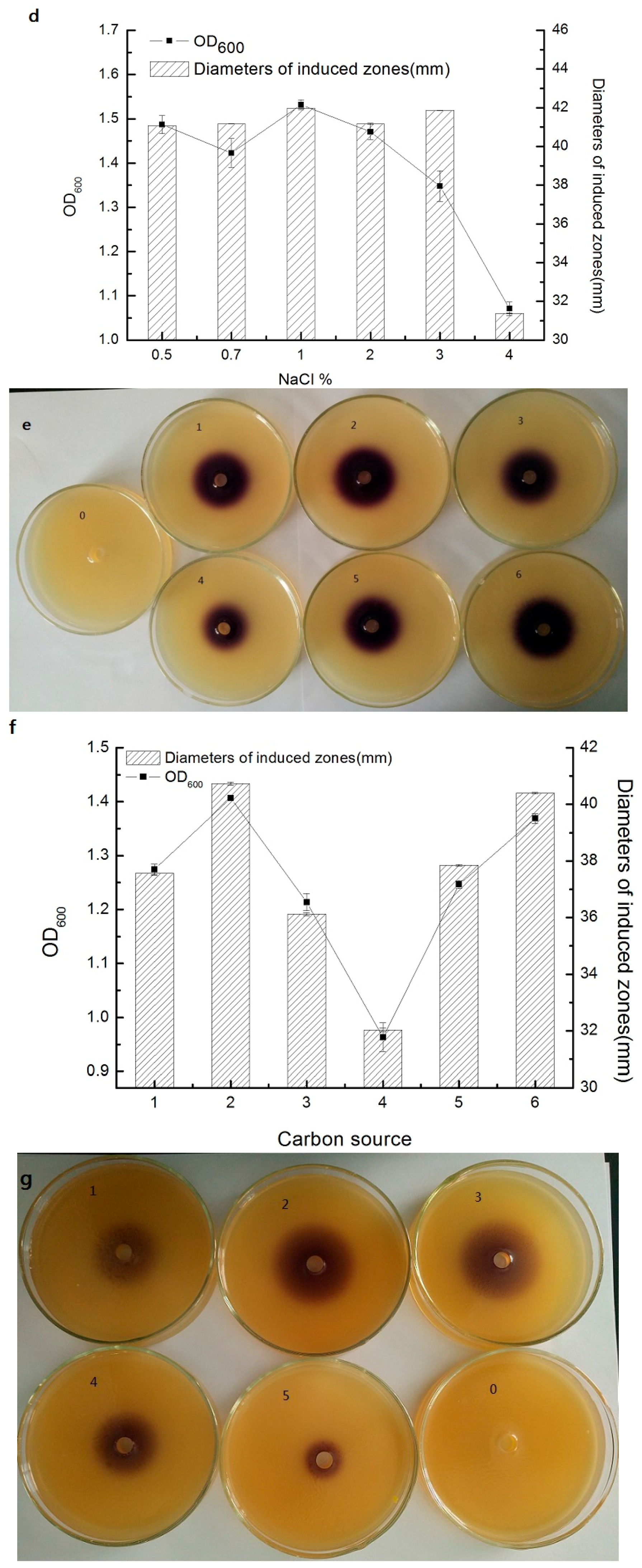
3.4. Effect of Different Conditions on Short Chain AHL Production by Strain AS7
3.5. GC-MS Analysis of AHLs in Extracts of Bacterial Cultures
3.6. The Regulation of Siderophore Secretion by QS in AS7
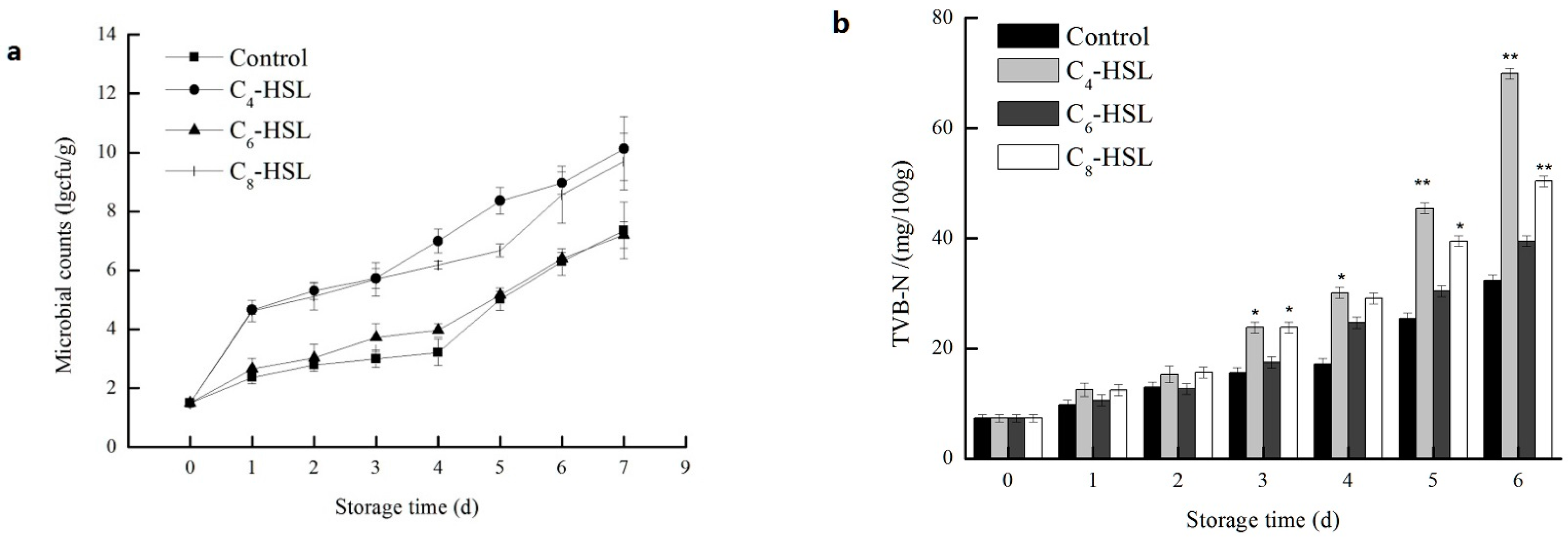
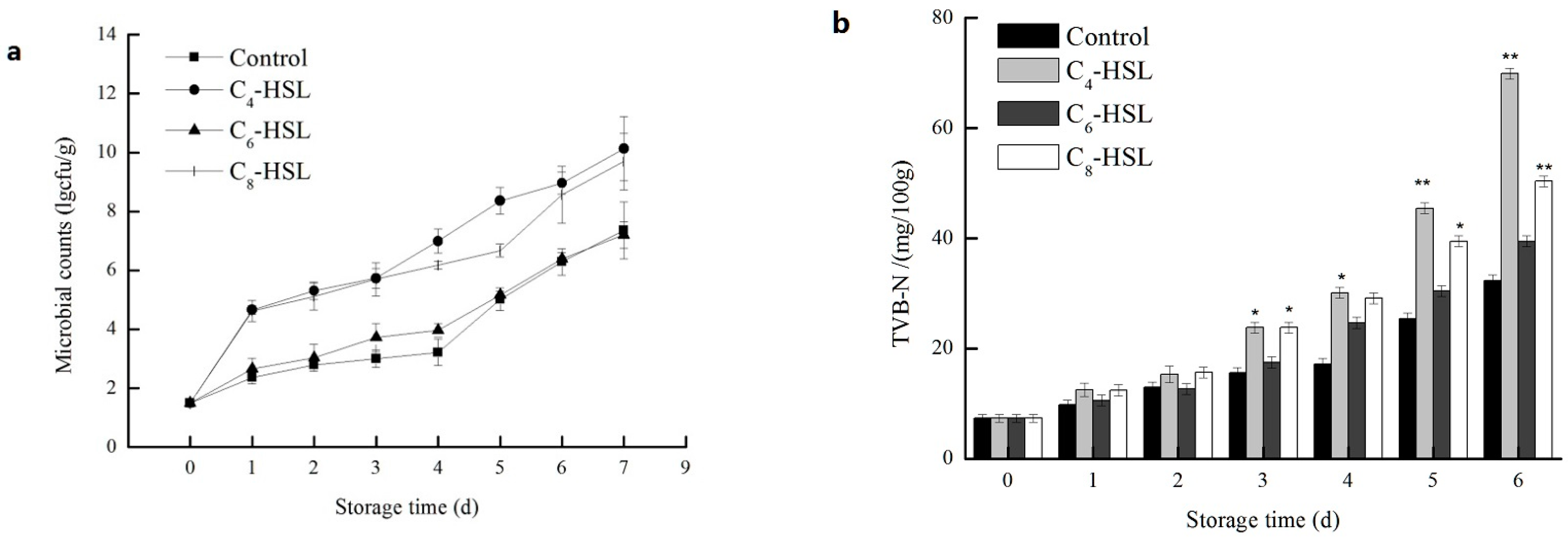
3.7. The Effect of QS on the Spoilage Process
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| APPA | Ala-Phe-Pro-ARYLAMIDASE |
| H2S | H2S PRODUCTION |
| BGLU | BETA-GLUCOSIDASE |
| ProA | l-Proline ARYLAMIDASE |
| SAC | SACCHAROSE/SUCROSE |
| ILATk | l-LACTATE alkalinisation |
| GlyA | Glycine ARYLAMIDASE |
| O129R | O/129 RESISTANCE (comp.vibrio.) |
| ADO | ADONITOL |
| BNAG | BETA-N-ACETYL-GLUCOSAMINIDASE |
| dMAL | d-MALTOSE |
| LIP | LIPASE |
| dTAG | d-TAGATOSE |
| AGLU | ALPHA-GLUCOSIDASE |
| ODC | ORNITHINE DECARBOXYLASE |
| GGAA | Glu-Gly-Arg-ARYLAMIDASE |
| PyrA | l-Pyrrolydonyl-ARYLAMIDASE |
| AGLTp | Glutamyl Arylamidase pNA |
| dMAN | d-MANNITOL |
| PLE | PALATINOSE |
| dTRE | d-TREHALOSE |
| SUCT | SUCCINATE alkalinisation |
| LDC | LYSINE DECARBOXYLASE |
| IMLTa | l-MALATE assimilation |
| IARL | l-ARABITOL |
| dGLU | d-GLUCOSE |
| dMNE | d-MANNOSE |
| TyrA | Tyrosine ARYLAMIDASE |
| CIT | CITRATE (SODIUM) |
| NAGA | Beta-N-ACETYL-GALACTOSAMINIDASE |
| IHISa | l-HISTIDINE assimilation |
| ELLM | ELLMAN |
| dCEL | d-CELLOBIOSE |
| GGT | GAMMA-GLUTAMYL-TRANSFERASE |
| BXYL | BETA-XYLOSIDASE |
| URE | UREASE |
| MNT | MALONATE |
| AGAL | ALPHA-GALACTOSIDASE |
| CMT | COURMARATE |
| ILATa | l-LACTATE assimilation |
| BGAL | BETA-GALACTOSIDASE |
| OFF | FERMENTATION/GLUCOSE |
| BAIap | BETA-Alanine arylamidase pNA |
| dSOR | d-SORBITOL |
| 5KG | 5-KETO-d-GLUCONATE |
| PHOS | PHOSPHATASE |
| BGUR | BETA-GLUCORONIDASE |
References
- Yunos, N.Y.M.; Tan, W.S.; Koh, C.L.; Sam, C.K.; Mohamad, N.I.; Tan, P.W.; Adrian, T.G.S.; Yin, W.F.; Chan, K.G. Pseudomonas cremoricolorata strain ND07 produces N-acyl homoserine lactones as quorum sensing molecules. Sensors 2014, 14, 11595–11604. [Google Scholar] [CrossRef] [PubMed]
- Schuster, M.; Joseph Sexton, D.; Diggle, S.P.; Peter Greenberg, E. Acyl-homoserine lactone quorum sensing: From evolution to application. Ann. Rev. Microbiol. 2013, 67, 43–63. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 2013, 31, 224–245. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Ye, K.P.; Zhang, Q.Q.; Dong, Y.; Xu, X.L.; Zhou, G.H. Biofilm formation of meat-borne Salmonella enterica and inhibition by the cell-free supernatant from Pseudomonas aeruginosa. Food Control 2013, 32, 650–658. [Google Scholar] [CrossRef]
- Hense, B.A.; Kuttler, C.; Müller, J.; Rothballer, M.; Hartmann, A.; Kreft, J.U. Does efficiency sensing unify diffusion and quorum sensing? Nat. Rev. Microbiol. 2007, 5, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.; Rothballer, M.; Hense, B.A.; Schröder, P. Bacterial quorum sensing compounds are important modulators of microbe-plant interactions. In The Plant Microbiome and Its Importance for Plant and Human Health; Frontiers E-books: Lausanne, Switzerland, 2015; p. 41. [Google Scholar]
- De Kievit, T.R.; Iglewski, B.H. Bacterial quorum sensing in pathogenic relationships. Infect. Immun. 2000, 68, 4839–4849. [Google Scholar] [CrossRef] [PubMed]
- Jahid, I.K.; Mizan, M.F.R.; Ha, A.J.; Ha, S.D. Effect of salinity and incubation time of planktonic cells on biofilm formation, motility, exoprotease production, and quorum sensing of Aeromonas hydrophila. Food Microbiol. 2015, 49, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Rani, S.; Kumar, A.; Malik, A.K.; Schmitt-Kopplin, P. Occurrence of N-acyl homoserine lactones in extracts of bacterial strain of Pseudomonas aeruginosa and in sputum sample evaluated by gas chromatography-mass spectrometry. Am. J. Anal. Chem. 2011, 2, 294. [Google Scholar] [CrossRef]
- Swift, S.; Winson, M.K.; Chan, P.F.; Bainton, N.J.; Birdsall, M.; Reeves, P.J.; Rees, C.E.D.; Chhabra, S.R.; Hill, P.J.; Throup, J.P.; et al. A novel strategy for the isolation of luxl homologues: Evidence for the widespread distribution of a LuxR: Luxl superfamily in enteric bacteria. Mol. Microbiol. 1993, 10, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.; Christensen, A.B.; Ravn, L.; Molin, S.; Givskov, M. Production of acylated homoserine lactones by psychrotrophic members of the Enterobacteriaceae isolated from foods. Appl. Environ. Microbiol. 1999, 65, 3458–3463. [Google Scholar] [PubMed]
- Ravn, L.; Christensen, A.B.; Molin, S.; Givskov, M.; Gram, L. Methods for detecting acylated homoserine lactones produced by Gram-negative bacteria and their application in studies of AHL-production kinetics. J. Microbiol. Methods 2001, 44, 239–251. [Google Scholar] [CrossRef]
- Shaw, P.D.; Ping, G.; Daly, S.L.; Cha, C.; Cronan, J.E.; Rinehart, K.L.; Farrand, S.K. Detecting and characterizing N-acyl-homoserine lactone signal molecules by thin-layer chromatography. Proc. Natl. Acad. Sci. USA 1997, 94, 6036–6041. [Google Scholar] [CrossRef] [PubMed]
- Ortori, C.A.; Dubern, J.F.; Chhabra, S.R.; Cámara, M.; Hardie, K.; Williams, P.; Barrett, D.A. Simultaneous quantitative profiling of N-acyl-l-homoserine lactone and 2-alkyl-4 (1H)-quinolone families of quorum-sensing signaling molecules using LC-MS/MS. Anal. Bioanal. Chem. 2011, 399, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Wu, H.; Zeng, M.; Liu, Z.; Wang, Y. The involvement of bacterial quorum sensing in the spoilage of refrigerated Litopenaeus vannamei. Int. J. Food Microbiol. 2015, 192, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhu, N.; Kong, L.; Bei, Y.; Zheng, T.; Ding, X.; He, Z. First case of soft shell disease in Chinese soft-shelled turtle (Trionyx sinens) associated with Aeromonas sobria–A. veronii complex. Aquaculture 2013, 406, 62–67. [Google Scholar] [CrossRef]
- Majtán, J.; Černy, J.; Ofúkaná, A.; Takáč, P.; Kozánek, M. Mortality of therapeutic fish Garra rufa caused by Aeromonas sobria. Asian Pac. J. Trop. Biomed. 2012, 2, 85–87. [Google Scholar] [CrossRef]
- Stano, F.; Brindicci, G.; Monno, R.; Rizzo, C.; Ghezzani, F.; Carbonara, S.; Guaglianone, E.; Donelli, G.; Monno, L. Aeromonas sobria sepsis complicated by rhabdomyolysis in an HIV-positive patient: Case report and evaluation of traits associated with bacterial virulence. Int. J. Infect. Dis. 2009, 13, e113–e118. [Google Scholar] [CrossRef] [PubMed]
- Namdari, H.; Bottone, E.J. Aeromonas species: Pathogens of aquatic inhabitants with a human host range. Clin. Microbiol. Newsl. 1991, 13, 113–116. [Google Scholar] [CrossRef]
- Jahid, I.K.; Lee, N.Y.; Kim, A.; Ha, S.D. Influence of glucose concentrations on biofilm formation, motility, exoprotease production, and quorum sensing in Aeromonas hydrophila. J. Food Prot. 2013, 76, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; Liu, Y.; Jiang, Y.; Zhu, W.; Zhuang, X. Production of N-acyl homoserine lactones and virulence factors of waterborne Aeromonas hydrophila. Indian J. Microbiol. 2013, 53, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Khajanchi, B.K.; Sha, J.; Kozlova, E.V.; Erova, T.E.; Suarez, G.; Sierra, J.C.; Popov, V.L.; Horneman, A.J.; Chopra, A.K. N-acylhomoserine lactones involved in quorum sensing control the type VI secretion system, biofilm formation, protease production, andin vivo virulence in a clinical isolate of Aeromonas hydrophila. Microbiology 2009, 155, 3518–3531. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.A.; Khattab, M.A.; Rohman, R.Z.A. Performance of Vitek-2 System for Detection of Inducible Clindamycin Resistance among Clinical Isolates of Staphylococci in Comparison to the d-Test. Egypt. J. Med. Microbiol. (EJMM) 2015, 24, 99–104. [Google Scholar]
- Chen, J.W.; Koh, C.L.; Sam, C.K.; Yin, W.F.; Chan, K.G. Short chain N-acyl homoserine lactone production by soil isolate Burkholderia sp. strain A9. Sensors 2013, 13, 13217–13227. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Gray, J.M.; Griffiths, M.W. Occurrence of proteolytic activity and N-acyl-homoserine lactone signals in the spoilage of aerobically chill-stored proteinaceous raw foods. J. Food Prot. 2006, 69, 2729–2737. [Google Scholar] [PubMed]
- Zimmer, B.L.; May, A.L.; Bhedi, C.D.; Dearth, S.P.; Prevatte, C.W.; Pratte, Z.; Campagna, S.R.; Richardson, L.L. Quorum sensing signal production and microbial interactions in a polymicrobial disease of corals and the coral surface mucopolysaccharide layer. PLoS ONE 2014, 9, e108541. [Google Scholar] [CrossRef] [PubMed]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Herbert, R.A.; Hendrie, M.S.; Gibson, D.M.; Shewan, J.M. Bacteria active in the spoilage of certain sea foods. J. Appl. Bacteriol. 1971, 34, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed]
- Cha, C.; Gao, P.; Chen, Y.C.; Shaw, P.D.; Farrand, S.K. Production of acyl-homoserine lactone quorum-sensing signals by gram-negative plant-associated bacteria. Mol. Plant-Microbe Interact. 1998, 11, 1119–1129. [Google Scholar] [CrossRef] [PubMed]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [PubMed]
- Medina-Martinez, M.S.; Uyttendaele, M.; Demolder, V.; Debevere, J. Influence of food system conditions on N-acyl-l-homoserine lactones production by Aeromonas spp. Int. J. Food Microbiol. 2006, 112, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Flodgaard, L.R.; Christensen, A.B.; Molin, S.; Givskov, M.; Gram, L. Influence of food preservation parameters and associated microbiota on production rate, profile and stability of acylated homoserine lactones from food-derived Enterobacteriaceae. Int. J. Food Microbiol. 2003, 84, 145–156. [Google Scholar] [CrossRef]
- Fuqua, C.; Parsek, M.R.; Greenberg, E.P. Regulation of gene expression by cell-to-cell communication: Acyl-homoserine lactone quorum sensing. Ann. Rev. Genet. 2001, 35, 439–468. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, T.R.; Bianco, G.; Palazzo, L.; Quaranta, V. Occurrence of N-acyl-l-homoserine lactones in extracts of some Gram-negative bacteria evaluated by gas chromatography–mass spectrometry. Anal. Biochem. 2007, 361, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.G.; Puthucheary, S.D.; Chan, X.Y.; Yin, W.F.; Wong, C.S.; Too, W.S.S.; Chua, K.H. Quorum sensing in Aeromonas species isolated from patients in Malaysia. Curr. Microbiol. 2011, 62, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.; Ravn, L.; Rasch, M.; Bruhn, J.B.; Christensen, A.B.; Givskov, M. Food spoilage—Interactions between food spoilage bacteria. Int. J. Food Microbiol. 2002, 78, 79–97. [Google Scholar] [CrossRef]
- Gram, L. The influence of substrate on siderophore production by fish spoilage bacteria. J. Microbiol. Methods 1996, 25, 199–205. [Google Scholar] [CrossRef]
- Bruhn, J.B.; Christensen, A.B.; Flodgaard, L.R.; Nielsen, K.F.; Larsen, T.O.; Givskov, M.; Gram, L. Presence of acylated homoserine lactones (AHLs) and AHL-producing bacteria in meat and potential role of AHL in spoilage of meat. Appl. Environ. Microbiol. 2004, 70, 4293–4302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhu, S.; Jatt, A.N.; Zeng, M. Characterization of N-acyl homoserine lactones (AHLs) producing bacteria isolated from vacuum-packaged refrigerated turbot (Scophthalmus maximus) and possible influence of exogenous AHLs on bacterial phenotype. J. Gen. Appl. Microbiol. 2016, 62, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.B.; Riedel, K.; Eberl, L.; Flodgaard, L.R.; Molin, S.; Gram, L.; Givskov, M. Quorum-sensing-directed protein expression in Serratia proteamaculans B5a. Microbiology 2003, 149, 471–483. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Phenotype | Item | Phenotype | Item | Phenotype |
|---|---|---|---|---|---|
| APPA | + | PyrA | − | dCEL | − |
| H2S | − | AGLTp | − | GGT | − |
| BGLU | − | dMAN | + | BXYL | − |
| ProA | − | PLE | − | URE | − |
| SAC | + | dTRE | + | MNT | − |
| ILATk | − | SUCT | + | AGAL | − |
| GlyA | − | LDC | − | CMT | + |
| O129R | + | IMLTa | + | ILATa | − |
| ADO | − | IARL | − | BGAL | + |
| BNAG | + | dGLU | + | OFF | + |
| dMAL | + | dMNE | + | BAIap | − |
| LIP | − | TyrA | + | dSOR | − |
| dTAG | − | CIT | − | 5KG | − |
| AGLU | − | NAGA | − | PHOS | − |
| ODC | − | IHISa | − | BGUR | − |
| GGAA | + | ELLM | + |
| AHLs | Fragment | Retention Times/min |
|---|---|---|
| C4–HSL | 32,43.1,57.1,71.1,83,102.1,125.1,143,153.1,171.1 | 4.172 |
| C6–HSL | 32,43.1,56.1,71.1,83,99.1,102.1,125,143.1,156.1 | 5.956 |
| C8–HSL | 32,43.1,57.1,69.1,83.1,102.1,125.1,143.1,156.1,207 | 7.942 |
| C10–HSL | 32,43.1,57.1,69,83.1,102.1,125,143,156.1,207 | 10.239 |
| C12–HSL | 32,43.1,57.1,71.1,83,102.1,125.1,143,156.1,207 | 12.836 |
| C14–HSL | 32,43.1,57.1,69,83.1,102.1,125.1,143,157.1 | 16.044 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Cui, F.; Bai, F.; Zhao, G.; Li, J. Involvement of Acylated Homoserine Lactones (AHLs) of Aeromonas sobria in Spoilage of Refrigerated Turbot (Scophthalmus maximus L.). Sensors 2016, 16, 1083. https://doi.org/10.3390/s16071083
Li T, Cui F, Bai F, Zhao G, Li J. Involvement of Acylated Homoserine Lactones (AHLs) of Aeromonas sobria in Spoilage of Refrigerated Turbot (Scophthalmus maximus L.). Sensors. 2016; 16(7):1083. https://doi.org/10.3390/s16071083
Chicago/Turabian StyleLi, Tingting, Fangchao Cui, Fengling Bai, Guohua Zhao, and Jianrong Li. 2016. "Involvement of Acylated Homoserine Lactones (AHLs) of Aeromonas sobria in Spoilage of Refrigerated Turbot (Scophthalmus maximus L.)" Sensors 16, no. 7: 1083. https://doi.org/10.3390/s16071083