1. Introduction
As organisms move through landscapes, they respond to biotic and abiotic factors to maximize access to resources and mates, while minimizing fitness costs. The structure of the landscape will interact with the movement response of organisms to affect connectivity [
1,
2]. The connectivity of populations is critical, both for maintaining regional populations and for species to shift their geographic range in response to climate change [
3]. Increasing recognition of the importance of connectivity to population persistence, combined with the development of new algorithms, fast computers and user-friendly software [
4], has led to a proliferation of research on population connectivity [
5] and conservation actions applying this knowledge across broad landscapes [
6,
7].
The Greek word,
συνοπτικός (
synopticos), means seeing everything together. A synoptic view is one that simultaneously integrates all elements. Most past applications of population connectivity modeling have been based on assessments of movement cost or corridor routes between a few select locations [
6,
7]. For example, least cost path modeling identifies the single, lowest cost route through a landscape between two points [
8]. However, effective conservation often depends on understanding the connectivity of every location simultaneously to all other locations, in a synoptic view.
There is obvious advantage in adopting synoptic perspectives on population connectivity, but how can they be achieved? Factorial least cost path analysis [
9] is a spatially synoptic form of least cost path analysis in which least cost paths are calculated for thousands or millions of combinations of locations across the landscape. These paths are then summed to show the density of least cost crossing any point in the study area. A second synoptic approach to landscape connectivity is cumulative resistant kernel modeling [
10], which calculates the expected density of dispersing individuals in each pixel in the landscape.
These synoptic connectivity modeling approaches have several advantages in assessing population connectivity. First, they provide prediction and mapping of expected functional connectivity for every pixel in the study area, rather than only for a few selected ‘linkage zones’ or source locations [
11]. Second, factorial least cost path and resistant kernel analysis can evaluate how different population sizes, dispersal abilities and vagilities will affect connectivity through the specification of dispersal thresholds [
12]. However, despite their theoretical advantages, synoptic approaches have not been widely used, in part because these new methods are substantially more computationally intensive than traditional approaches. In addition, little is known about how well synoptic methods perform in predicting organism movement.
Most assessments of population connectivity are based on applying connectivity models, synoptic or otherwise, to resistance surfaces [
5,
6,
7,
13]. Resistance surfaces depict the unit cost of traversing each location on the map. The spatial pattern of resistance determines the location of least cost paths, corridors and the extent of resistant kernels. Therefore, an essential part of assessing the performance of different connectivity modeling approaches would include evaluation of how well alternative resistance surfaces reflect the functional cost of movement for an organism of interest. The vast majority of published resistance surfaces used in connectivity assessments have been derived from unvalidated expert opinion [
5], and there have been few assessments of the performance of resistance surfaces in predicting organism movement with data independent of that used in parameterizing them (see [
14,
15,
16]).
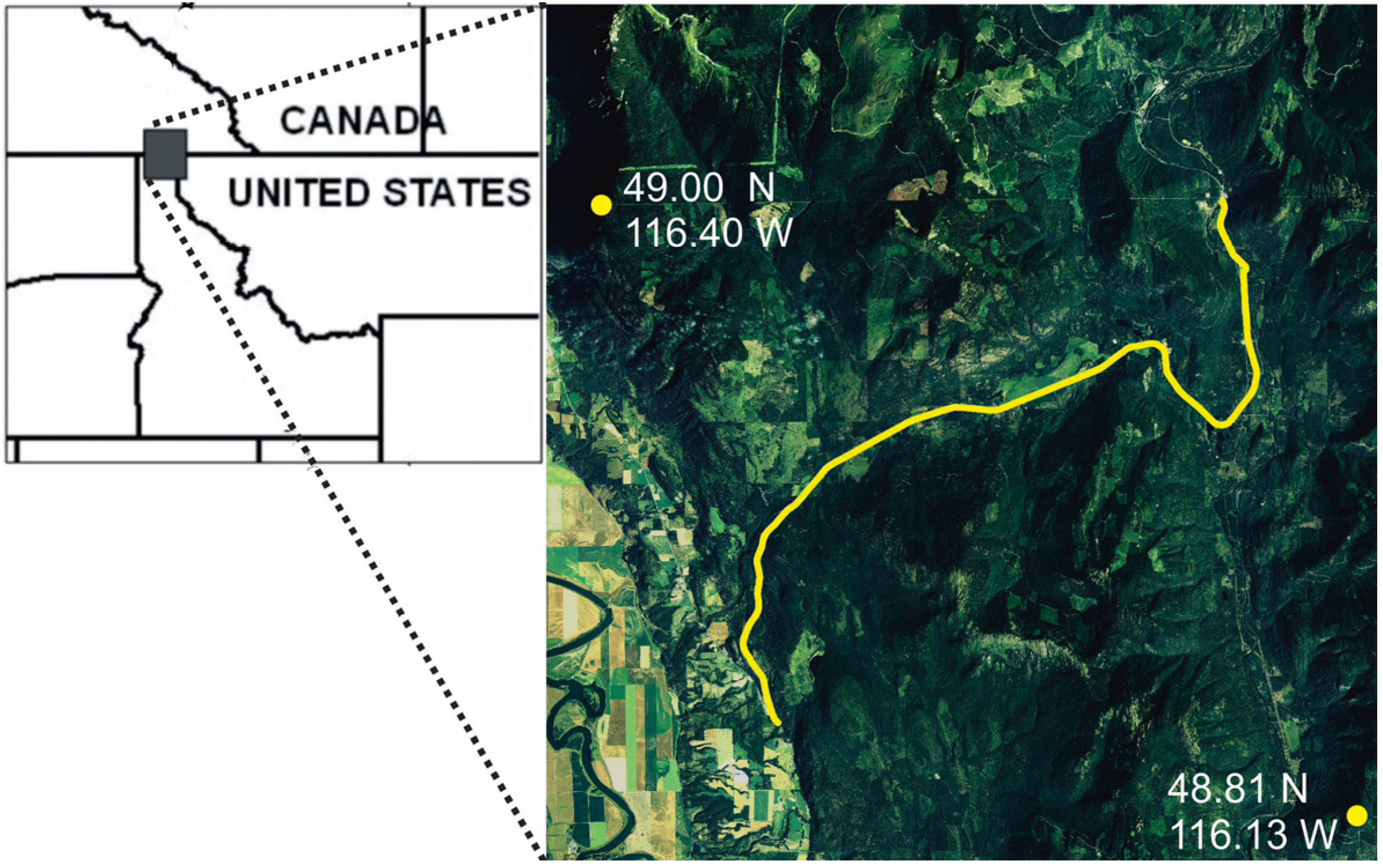
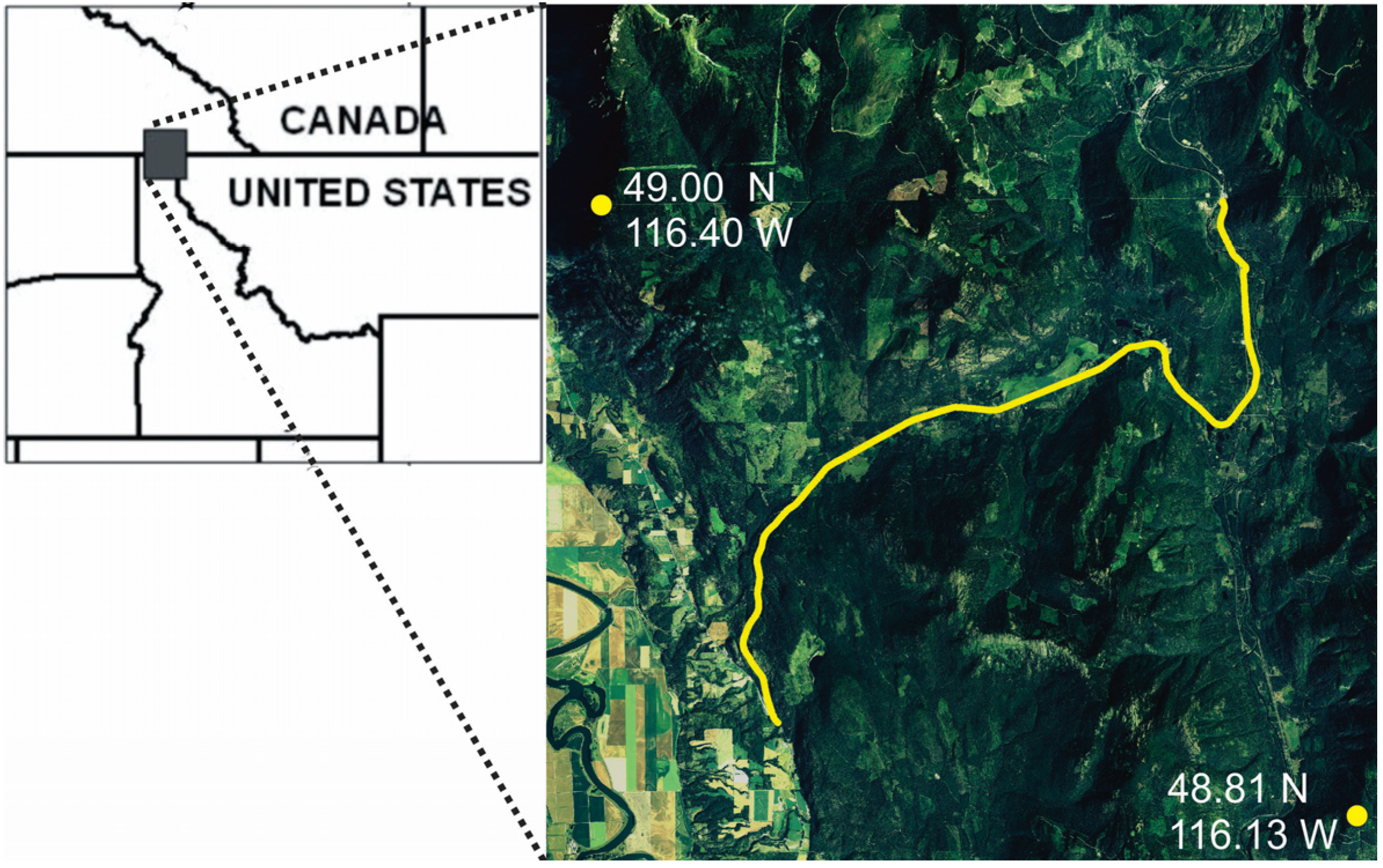
In this paper, we evaluate the performance of synoptic connectivity modeling approaches and alternative resistance models in predicting highway crossing locations of American black bear (Ursus americanus) in northern Idaho, USA. Our specific goals are to: (1) evaluate the relative performance of resistance surfaces derived from (a) individual-based landscape genetic modeling and (b) path-level modeling of landscape resistance based on GPS movement data; and (2) evaluate the relative performance of one local (neighborhood average landscape resistance) and two synoptic (factorial least cost path, resistant kernel) approaches to predicting connectivity across these two resistance surfaces. We hypothesize that (H1) the resistance surface derived from movement data will outperform the surface derived from gene flow in predicting bear highway crossings, that (H2) synoptic measures of connectivity would perform better than local landscape resistance and (H3) that the resistant kernel would be less sensitive to differences between resistance surfaces than factorial least cost paths.
3. Results
Consistent with our first hypothesis, we found that the resistance map produced from the movement data of black bears in this study area [
15] greatly outperformed the resistance map produced from the analysis of genetic differentiation [
19] across all methods of connectivity modeling (
Table 1). Specifically, for each of the connectivity modeling methods, predicted connectivity based on the Cushman and Lewis [
15] resistance model was higher at the locations of actual bear highway crossings than at randomly selected locations along the highway. In contrast, connectivity predictions based on landscape genetic analysis [
19] were not significantly associated with bear crossing locations in any of the methods.
Table 1.
Proportion of instances, of 1 × 10−7 random samples, where a random draw of 56 locations along U.S. Highway 95 in the study area produced a median connectivity value higher than the median connectivity value of the 56 actual black bear crossing locations for the two resistance maps across the three connectivity modeling approaches. The connectivity maps are a combination of the type of resistance surface (genetic based or movement based) and the type of connectivity modeling approach (average neighborhood resistance, factorial least cost path and resistant kernel).
Table 1.
Proportion of instances, of 1 × 10−7 random samples, where a random draw of 56 locations along U.S. Highway 95 in the study area produced a median connectivity value higher than the median connectivity value of the 56 actual black bear crossing locations for the two resistance maps across the three connectivity modeling approaches. The connectivity maps are a combination of the type of resistance surface (genetic based or movement based) and the type of connectivity modeling approach (average neighborhood resistance, factorial least cost path and resistant kernel).
| Connectivity Approach | Genetic | Movement |
|---|
| Average Neighborhood Resistance | 0.215 | 5.63 × 10−5 |
| Factorial Least Cost Paths | 0.999 | 3 × 10−7 |
| Resistant Kernel | 0.373 | 2.5 × 10−6 |
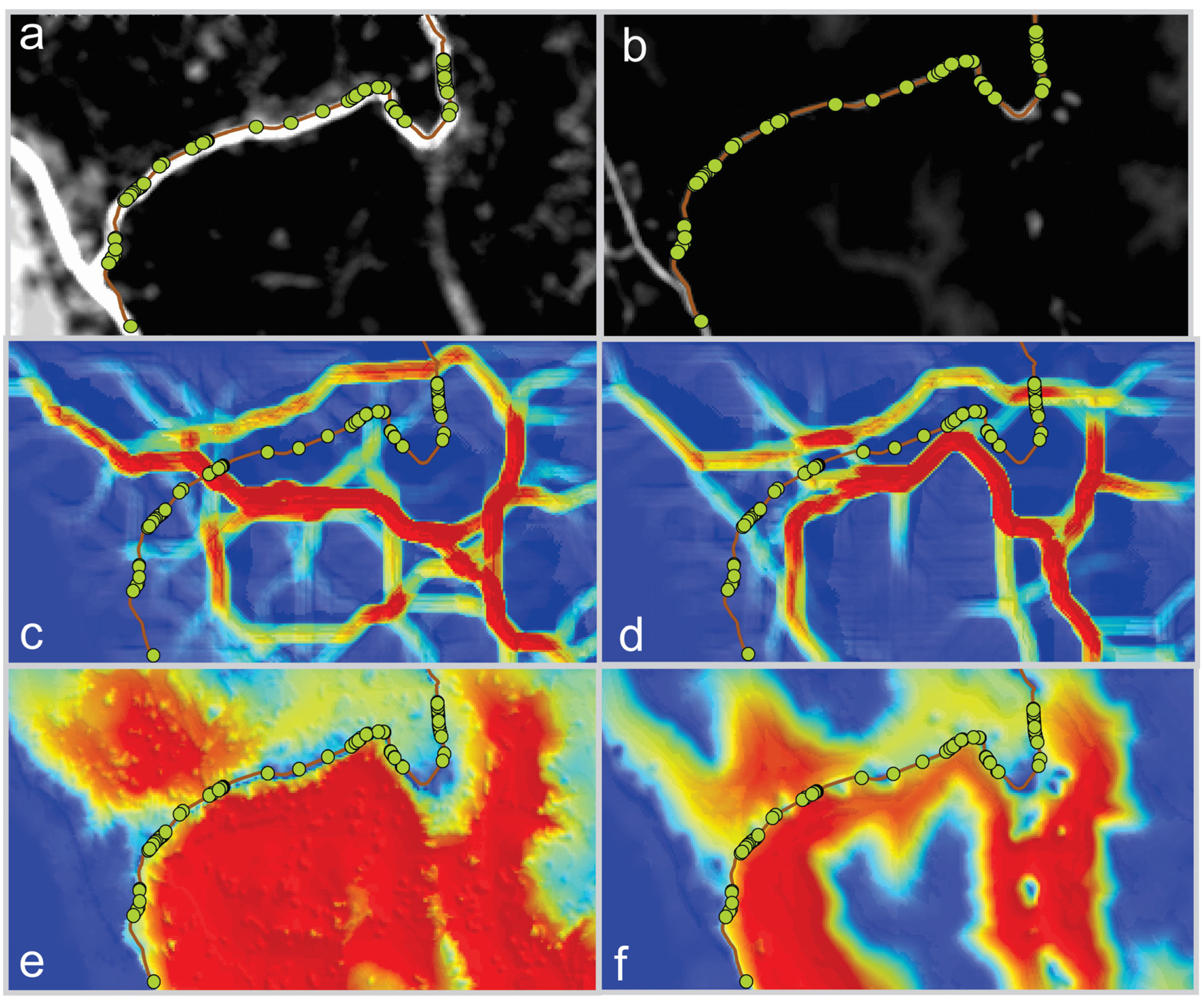
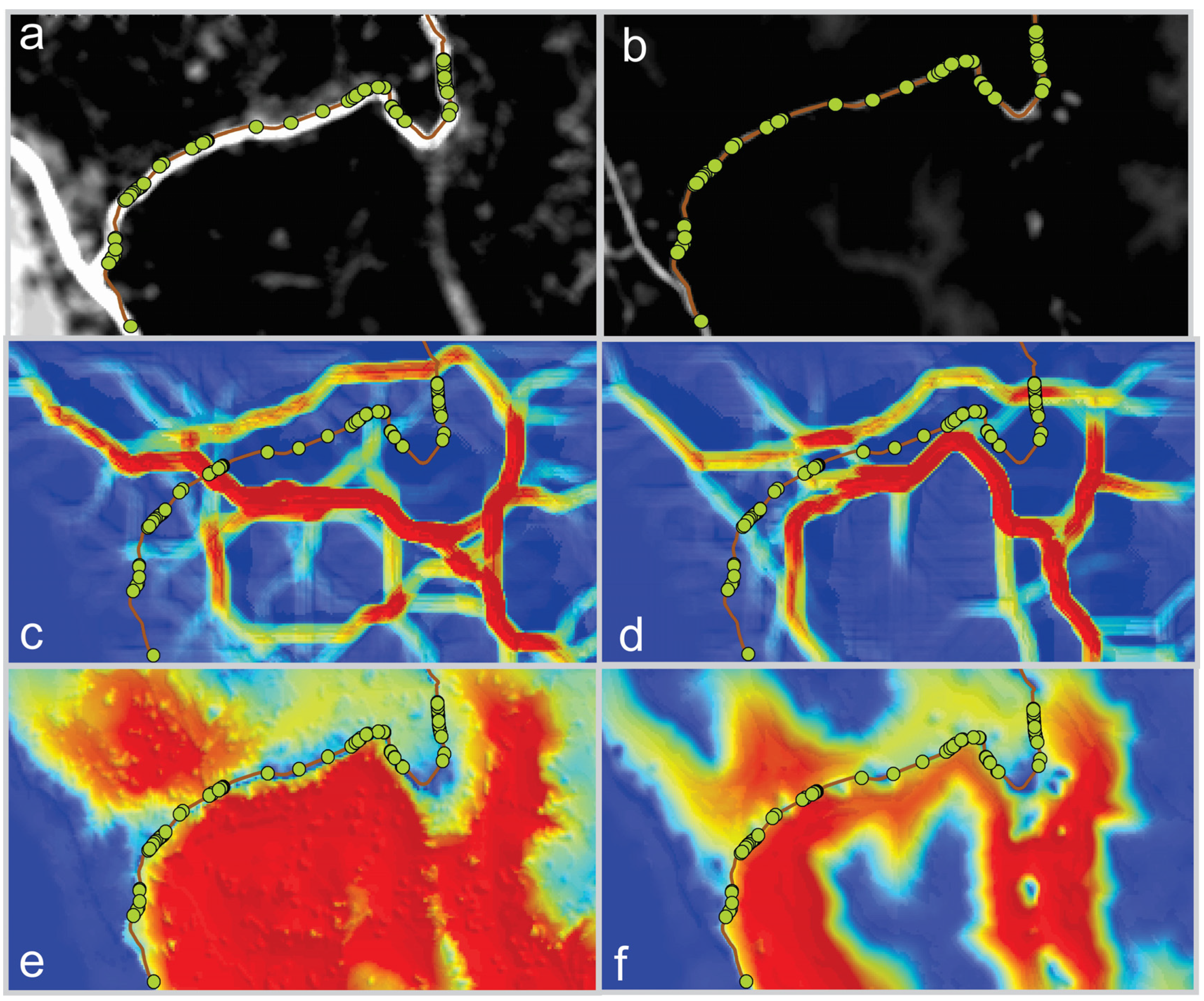
Consistent with our second hypothesis, the synoptic methods, factorial least cost paths and cumulative resistant kernels performed best at predicting the location of actual highway crossing events (
Table 1). For the movement-derived resistance surface, 3, 25 and 563 of the 1 × 10
7 random samples produced a median connectivity value as high as observed for the actual crossing locations in the factorial least cost path, resistant kernel and local landscape resistance connectivity methods, respectively. This indicates that the factorial least cost path approach had 8.3-times and 187.7-times fewer permutations with a median value less than the observed median in the kernel and local resistance and circuit approaches, respectively, for the movement-derived resistance surface. Conversely, for the genetics-derived resistance surface, 99.9%, 37.3% and 21.5% of the random samples had a median connectivity value higher than the actual crossing locations in the least cost path, kernel and focal resistance methods, respectively.
We produced a measure of sensitivity to the differences between resistance surfaces by calculating the proportional change in the number of permutations producing a median value greater than that observed in the actual crossing locations between the two resistance surfaces for each method. Based on this, and consistent with Hypothesis 3, we found that the factorial least cost path was very highly sensitive to differences between resistance surfaces (change from 3 × 10−7 to 99% of permutations less than the median resistance of actual crossings) and that local landscape resistance and resistant kernels were relatively insensitive to differences between the two resistance surfaces.
4. Discussion
Our analysis is among the first to simultaneously evaluate the performance of multiple resistance surfaces and connectivity modeling approaches in predicting independent animal movement data. Our first hypothesis was that a resistance surface derived directly from movement data would outperform a resistance surface produced from the analysis of genetic differentiation. We based this on the expectation that the landscape factors that govern movement may differ to some degree from those that govern gene flow. Gene flow in animals, such as the black bear, is effected through mating and dispersal events, which are related to movement, but are particular and rare kinds of movement. Thus, a resistance model optimizing the factors related to gene flow [
19] may not ideally reflect the factors that drive the behavioral decisions of individuals in selecting crossing locations. Conversely, resistance surfaces derived directly from movement data [
15] may be expected to more effectively predict the specific movement decisions made by individual bears. Our results dramatically demonstrate this, with extremely high performance of all four connectivity modeling methods when applied to the movement data-derived resistance surface and universally poor performance when applied to the landscape genetic-derived surface.
Cushman and Lewis [
15] compared these two resistance surfaces and noted that they are heuristically similar, containing the same factors with the same qualitative relationships between landscape features and resistance. They further noted a high correlation between the pixel values of the two surfaces and argued that movement data and genetic differentiation both supported the conclusion that landscape resistance for American black bears in the Rocky Mountains is facilitated by middle elevation forest and resisted by roads and human land uses. Our analysis, in contrast, suggested that these surfaces in fact differ substantially in terms of their predictions of functional connectivity across the study area in each of three connectivity modeling approaches. This suggests that even when two resistance surfaces include the same variables and have a high spatial correlation of resistance values, they may perform very differently in predicting animal movement and population connectivity.
Our second hypothesis proposed that synoptic connectivity modeling approaches, such as factorial least cost paths and resistant kernels, would outperform a local measure of population connectivity based on the average landscape resistance within a local neighborhood. We based this expectation on the idea that the movement path choices of individual bears, including where they cross a highway, would be influenced by the synoptic pattern of connectivity across a broad landscape and not just by the local landscape resistance at a crossing location. Consistent with this expectation, for the movement-derived resistance surface, two of the synoptic methods (cumulative factorial least cost path and cumulative resistant kernel) outperformed local landscape resistance, with factorial least cost paths apparently performing best. This is an important finding, as most applications of connectivity modeling in conservation biology have used local information or non-synoptic connectivity methods to predict movement corridors [
6,
11]. Our results suggest that synoptic methods (such as factorial paths and kernels) are best able to predict actual organism movement, since population connectivity is an emergent phenomenon driven by the cumulative influences of landscape structure throughout the population. Furthermore, factorial least cost path modeling best predicted crossing locations, likely because this method emphasizes optimal movement routes, and animals choosing locations to cross a potentially dangerous obstacle, such as a highway, are likely to select the routes that minimize risk. Consistent with this conclusion, Lewis
et al. [
17] evaluated movement data from GPS-collared black bears and found that bears selected for areas of forest on the landscape and away from human development along the road when crossing Highway 95.
We interpret the high performance of all connectivity modeling methods when applied to the movement-derived resistance map and low performance when applied to the genetics-derived map to indicate that the movement-derived resistance map is a close match to the factors that the bears were responding to, and the genetics-based map was a relatively poor match. Given this, we evaluated the sensitivity of the methods to the misspecification of the resistance surface. Ideally, one should use a method that is moderately sensitive to differences between resistance surfaces, such that the implications of landscape change in a single landscape or differences in habitat extent and fragmentation in different landscapes for connectivity can be quantified, but not so sensitive that the method fails to predict connectivity well when the resistance surface is approximately, but not ideally, parameterized. Our third hypothesis was intended to evaluate the performance of these methods based on these criteria. As expected, we found that the factorial least cost path method was extremely sensitive to the difference between the two resistance surfaces (more than 22-times more sensitive than the resistant kernel method). We expected this method to be most sensitive given that least cost path locations are spatially constrained to optimal narrow routes, and small changes in overall landscape resistance across the landscape may completely shift those paths. Cumulative resistant kernel methods had moderate sensitivity to differences in resistance between the two surfaces, which might provide sufficient sensitivity to evaluate different landscapes, while enough stability to obtain reliable predictions of the patterns and relative strength of connectivity across the landscape. Our results suggest that cumulative resistant appeared to offer the best combination of high predictive performance and sensitivity to differences in resistance surface parameterization. Specifically, resistant kernels appear to be the preferred choice when the goal is quantifying differences in connectivity between different landscapes or overtime, while factorial least cost paths would be best suited for the analysis of resistance maps where there is high certainty in resistance values and where researchers wish to localize predictions to prioritize specific locations for protection rather than evaluating landscape-wide patterns of connectivity. It would be interesting to compare the methods tested here with other spatial methods to identify where animals cross highways (e.g., [
22,
23,
24,
25])


{kind=link}
{kind=link}