Can Youthful Mesenchymal Stem Cells from Wharton’s Jelly Bring a Breath of Fresh Air for COPD?


Abstract
:1. Introduction
2. Inflammation and Airway Remodeling in COPD
3. Mesenchymal Stem Cells
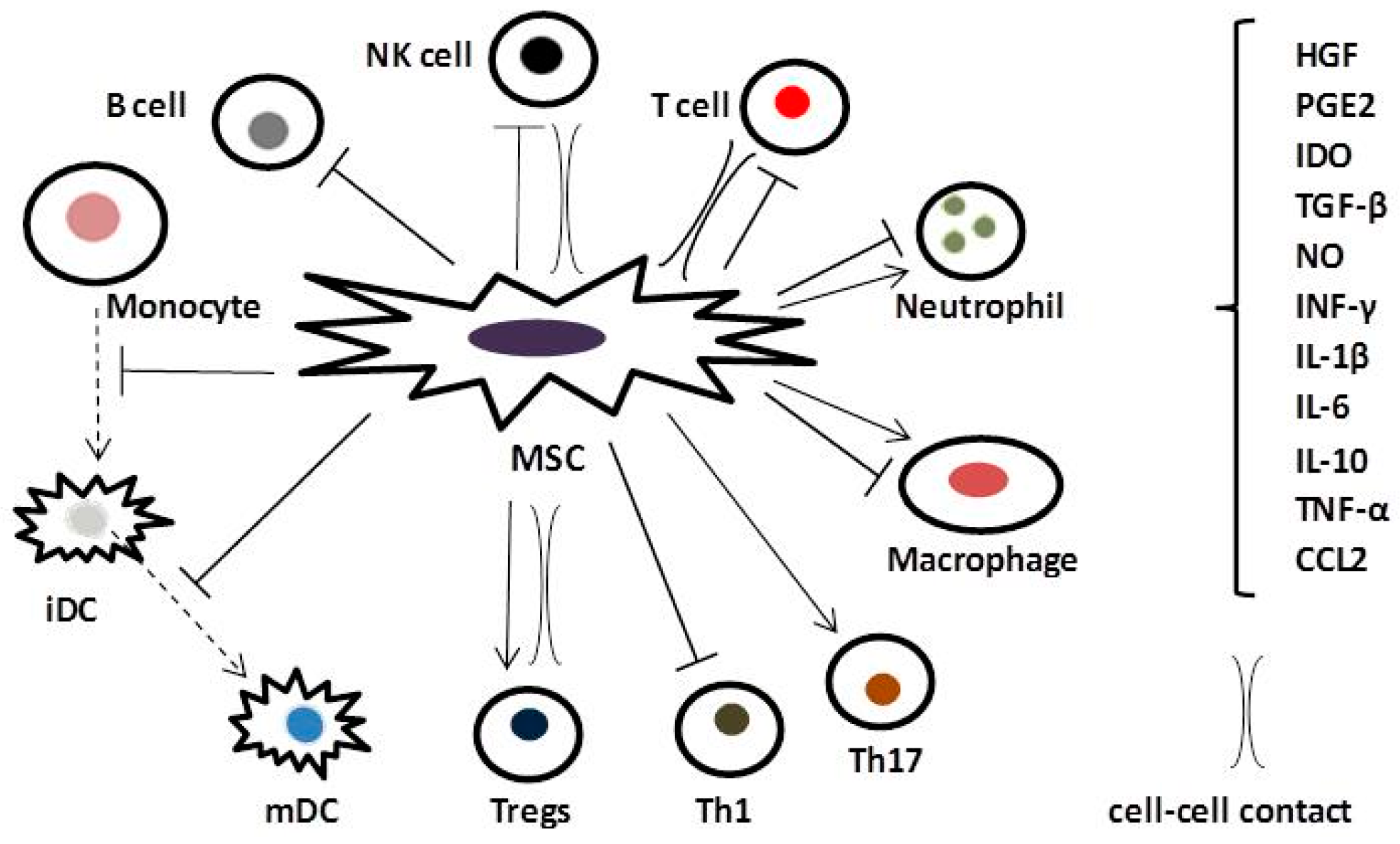
4. Anti-Inflammatory and Immunomodulatory Effects of MSCs in COPD
5. Current Status of MSC Therapy for the Treatment of COPD
6. WJ-MSCs: A Promising Youthful Contender in Stem Cell Therapy for COPD
7. Finding Optimal MSC Treatment Methodologies for COPD
8. Alternative Cell-Based Therapies for COPD
9. Conclusions
Acknowledgments
Conflicts of Interest
References
- Strategy for the Diagnosis, Management and Prevention of COPD. Global Initiative for Chronic Obstructive Lung Disease (GOLD). 2017. Available online: http://goldcopd.org (accessed on 23 August 2017).
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Chung, K.F.; Adcock, I.M. Multifaceted mechanisms in COPD: Inflammation, immunity, and tissue repair and destruction. Eur. Respir. J. 2008, 31, 1334–1356. [Google Scholar] [CrossRef] [PubMed]
- Tuder, R.M.; Petrache, I. Pathogenesis of chronic obstructive pulmonary disease. J. Clin. Investig. 2012, 122, 2749–2755. [Google Scholar] [CrossRef] [PubMed]
- Bagdonas, E.; Raudoniute, J.; Bruzauskaite, I.; Aldonyte, R. Novel aspects of pathogenesis and regeneration mechanisms in COPD. Int. J. Chronic Obstr. Pulm. Dis. 2015, 10, 995–1013. [Google Scholar] [PubMed]
- Chen, Z.-H.; Kim, H.P.; Ryter, S.W.; Cho, A.M.K. Identifying targets for COPD treatment through gene expression analyses. Int. J. Chronic Obstr. Pulm. Dis. 2008, 3, 359–370. [Google Scholar] [PubMed]
- Molfino, N.A.; Coyle, A.J. Gene-environment interactions in chronic obstructive pulmonary disease. Int. J. Chronic Obstr. Pulm. Dis. 2008, 3, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Fischer, B.M.; Pavlisko, E.; Voynow, J.A. Pathogenic triad in COPD: Oxidative stress, protease-antiprotease imbalance, and inflammation. Int. J. Chronic Obstr. Pulm. Dis. 2011, 6, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, S.D. End-stage chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2001, 164, 339–340. [Google Scholar] [CrossRef] [PubMed]
- Ind, P.W. COPD disease progression and airway inflammation: Uncoupled by smoking cessation. Eur. Respir. J. 2005, 26, 764–766. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, G.C.; Seemungal, T.A.R.; Patel, I.S.; Bhowmik, A.; Wilkinson, T.M.; Hurst, J.R.; Maccallum, P.K.; Wedzicha, J.A. Airway and systemic inflammation and decline in lung function in patients with COPD. Chest 2005, 128, 1995–2004. [Google Scholar] [CrossRef] [PubMed]
- Rovina, N.; Koutsoukou, A.; Koulouris, N.G. Inflammation and Immune Response in COPD: Where do we stand? Mediat. Inflamm. 2013. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. New concepts in chronic obstructive pulmonary disease. Annu. Rev. Med. 2003, 54, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Boer, W.; Alagappan, V.; Sharma, H. Molecular mechanisms in chronic obstructive pulmonary disease. Cell Biochem. Biophys. 2007, 47, 131–147. [Google Scholar] [CrossRef]
- Hogg, J.C.; Timens, W. The pathology of chronic obstructive pulmonary disease. Annu. Rev. Pathol. 2009, 4, 435–459. [Google Scholar] [CrossRef] [PubMed]
- Wecht, S.; Rojas, M. Mesenchymal stem cells in the treatment of chronic lung disease. Respirology 2016, 21, 1366–1375. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.; Son, Y. Crosstalk between mesenchymal stem cells and macrophages in tissue repair. Tissue Eng. Regen. Med. 2014, 11, 431–438. [Google Scholar] [CrossRef]
- Williams, T.J.; Jose, P.J. Neutrophils in chronic obstructive pulmonary disease. Novartis Found. Symp. 2001, 234, 136–141. [Google Scholar] [PubMed]
- Barnes, P.J.; Cosio, M.G. Characterization of T lymphocytes in chronic obstructive pulmonary disease. PLoS Med. 2004, 1, e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharafkhaneh, A.; Hanania, N.A.; Kim, V. Pathogenesis of emphysema: From the bench to the bedside. Proc. Am. Thorac. Soc. 2008, 5, 475–477. [Google Scholar] [CrossRef] [PubMed]
- Hogg, J.C.; Paré, P.D.; Hackett, T.L. The contribution of small airway obstruction to the pathogenesis of chronic obstructive pulmonary disease. Physiol. Rev. 2017, 97, 529–552. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, G.; Fox, J.; Ashton, B.; Middleton, J. Concise review: Mesenchymal stem cells: Their phenotype, differentiation capacity, immunological features, and potential for homing. Stem Cells 2007, 25, 2739–2749. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.J.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Ostanin, A.A.; Petrovskii, Y.L.; Sheleva, E.Y.; Chernykh, E.R. Multiplex analysis of cytokines, chemokines, growth factors, MMP-9 and TIMP-1 produced by human bone marrow, adipose tissue, and placental mesenchymal stromal cells. Bull. Exp. Biol. Med. 2011, 151, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.M.; Barry, F.P.; Murphy, J.M.; Mahon, B.P. Mesenchymal stem cells avoid allogeneic rejection. J. Inflamm. 2005, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- English, K. Mechanisms of mesenchymal stromal cell immunomodulation. Immunol. Cell Biol. 2013, 91, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Spees, J.L.; Lee, R.H.; Gregory, C.A. Mechanisms of mesenchymal stem/stromal cell function. Stem Cell Res. Ther. 2016, 7, 125. [Google Scholar] [CrossRef] [PubMed]
- Moodley, Y.; Manuelpillai, U.; Weiss, D. Cellular therapies for lung disease: A distant Horizon. Respirology 2011, 16, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Inamdar, A.C.; Inamdar, A.A. Mesenchymal stem cell therapy in lung disorders: Pathogenesis of lung diseases and mechanism of action of mesenchymal stem cell. Exp. Lung Res. 2013, 39, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Conese, M.; Carbone, A.; Castellani, S.; di Gioia, S. Paracrine effects and heterogeneity of marrow-derived stem/progenitor cells: Relevance for the treatment of respiratory diseases. Cells Tissues Organs 2013, 197, 445–473. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.J. Current status of stem cells and regenerative medicine in lung biology and diseases. Stem Cells 2014, 32, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Chiu, M.; Motan, D.A.L.; Zhang, Z.; Chen, L.; Ji, H.-L.; Tse, H.-F.; Fu, Q.-L.; Lian, Q. Mesenchymal stem cells and immunomodulation: Current status and future prospects. Cell Death Dis. 2016, 7, e2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Blanc, K.; Ringden, O. Immunomodulation by mesenchymal stem cells and clinical experience. J. Intern. Med. 2007, 262, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-T.; Ting, C.-H.; Yen, M.-L.; Liu, K.J.; Sytwu, H.K.; Wu, K.K.; Yen, B.L. Human mesenchymal stem cells (MSCs) for treatment towards immune- and inflammation-mediated diseases: Review of current clinical trials. J. Biomed. Sci. 2016, 23, 76. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.-L.; Lin, C.-H.; Yao, C.-L. Mesenchymal stem cell administration in patients with chronic obstructive pulmonary disease: State of the science. Stem Cells Int. 2017, 2017, 8916570. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.K.; Shin, I.S.; Ko, M.S.; Jo, J.Y.; Ra, J.C. Journey of mesenchymal stem cells for homing: Strategies to enhance efficacy and safety of stem cell therapy. Stem Cells Int. 2012, 2012, 342968. [Google Scholar] [CrossRef] [PubMed]
- Karp, J.M.; Leng Teo, G.S. Mesenchymal stem cell homing: The devil is in the details. Cell Stem Cell 2009, 4, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Schrepfer, S.; Deuse, T.; Reichenspurner, H.; Fischbein, M.P.; Robbins, R.C.; Pelletier, M.P. Stem cell transplantation: The lung barrier. Transplant. Proc. 2007, 39, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Barbash, I.M.; Chouraqui, P.; Baron, J.; Feinberg, M.S.; Etzion, S.; Tessone, A.; Miller, L.; Guetta, E.; Zipori, D.; Kedes, L.H.; et al. Systemic delivery of bone marrow-derived mesenchymal stem cells to the infarcted myocardium: Feasibility, cell migration, and body distribution. Circulation 2003, 108, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Loi, R.; Beckett, T.; Goncz, K.K.; Suratt, B.T.; Weiss, D.J. Limited restoration of cystic fibrosis lung epithelium in vivo with adult bone marrow-derived cells. Am. J. Respir. Crit. Care Med. 2006, 173, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Sueblinvong, V.; Loi, R.; Eisenhauer, P.L.; Bernstein, I.M.; Suratt, B.T.; Spees, J.L.; Weiss, D.J. Derivation of lung epithelium from human cord blood-derived mesenchymal stem cells. Am. J. Respir. Crit. Care Med. 2008, 177, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Moodley, Y.; Atienza, D.; Manuelpillai, U.; Samuel, C.S.; Tchongue, J.; Ilancheran, S.; Boyd, R.; Trounson, A. Human umbilical cord mesenchymal stem cells reduce fibrosis of bleomycin-induced lung injury. Am. J. Pathol. 2009, 175, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.R.; Liu, L.; Chen, S.; Yang, Y.; Zhao, H.J.; Liu, L.; Guo, F.M.; Lu, X.M.; Qiu, H.B. Activation of canonical wnt pathway promotes differentiation of mouse bone marrow-derived MSCs into type II alveolar epithelial cells, confers resistance to oxidative stress, and promotes their migration to injured lung tissue in vitro. J. Cell. Physiol. 2013, 228, 1270–1283. [Google Scholar] [CrossRef] [PubMed]
- Yagi, H.; Soto-Gutierrez, A.; Parekkadan, B.; Kitagawa, Y.; Tompkins, R.G.; Kobayashi, N.; Yarmush, M.L. Mesenchymal stem cells: Mechanisms of immunomodulation and homing. Cell Transplant. 2010, 19, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Hass, R.; Kasper, C.; Bohm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemeth, K.; Leelahavanichkul, A.; Yuen, P.S.; Mayer, B.; Parmelee, A.; Doi, K.; Robey, P.G.; Leelahavanichkul, K.; Koller, B.H.; Brown, J.M.; et al. Bone marrow stromal cells attenuate sepsis via prostaglandin E2-dependent reprogramming of host macrophages to increase their interleukin-10 production. Nat. Med. 2009, 15, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Su, X.; Popov, B.; Lee, J.W.; Serikov, V.; Matthay, M.A. Intrapulmonary delivery of bone marrow-derived mesenchymal stem cells improves survival and attenuates endotoxin-induced acute lung injury in mice. J. Immunol. 2007, 179, 1855–1863. [Google Scholar] [CrossRef] [PubMed]
- Zhen, G.; Xue, Z.; Zhao, J.; Gu, N.; Tang, Z.; Xu, Y.; Zhang, Z. Mesenchymal stem cell transplantation increases expression of vascular endothelial growth factor in papain-induced emphysematous lungs and inhibits apoptosis of lung cells. Cytotherapy 2010, 12, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.W.; Kim, S.Y.; Lee, J.H.; Lee, J.S.; Van Ta, Q.; Kim, M.; Oh, Y.M.; Lee, Y.S.; Lee, S.D. Bone marrow cells repair cigarette smoke-induced emphysema in rats. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 301, L255–L266. [Google Scholar] [CrossRef] [PubMed]
- Raffaghello, L.; Bianchi, G.; Bertolotto, M.; Montecucco, F.; Busca, A.; Dallegri, F.; Ottonello, L.; Pistoia, V. Human mesenchymal stem cells inhibit neutrophil apoptosis: A model for neutrophil preservation in the bone marrow niche. Stem Cells 2008, 26, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Munir, H.; Luu, N.T.; Clarke, L.S.; Nash, G.B.; McGettrick, H.M. Comparative ability of mesenchymal stromal cells from different tissues to limit neutrophil recruitment to inflamed endothelium. PLoS ONE 2016, 11, e0155161. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Ro, H.; Hong, J.H.; Lee, E.M.; Cho, B.; Yeom, H.J.; Kim, M.G.; Oh, K.H.; Ahn, C.; Yang, J. Immunosuppressive mechanisms of embryonic stem cells and mesenchymal stem cells in alloimmune response. Transplant. Immunol. 2011, 25, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Krampera, M.; Glennie, S.; Dyson, J.; Scott, D.; Laylor, R.; Simpson, E.; Dazzi, F. Bone marrow mesenchymal stem cells inhibit the response of naive and memory antigen-specific T cells to their cognate peptide. Blood 2003, 101, 3722–3729. [Google Scholar] [CrossRef] [PubMed]
- Sotiropoulou, P.A.; Perez, S.A.; Gritzapis, A.D.; Baxevanis, C.N.; Papamichail, M. Interactions between human mesenchymal stem cells and natural killer cells. Stem Cells 2006, 24, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Wang, Y.; Jin, Y.; Zhang, Q.; Zhang, Y.; Wang, L.; Shen, B.; Yin, S.; Liu, W.; Cui, L.; et al. A critical role of IFN γ in priming MSC-mediated suppression of T cell proliferation through up-regulation of B7-H1. Cell Res. 2008, 18, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Zhao, X.; Zhang, L.; Zhang, J.; L’Huillier, A.; Ling, W.; Roberts, A.I.; Le, A.D.; Shi, S.; Shao, C.; et al. Inflammatory cytokine-induced intercellular adhesion molecule-1 and vascular cell adhesion molecule-1 in mesenchymal stem cells are critical for immunosuppression. J. Immunol. 2010, 184, 2321–2328. [Google Scholar] [CrossRef] [PubMed]
- Yuhgetsu, H.; Ohno, Y.; Funaguchi, N.; Asai, T.; Sawada, M.; Takemura, G.; Minatoguchi, S.; Fujiwara, H.; Fujiwara, T. Beneficial effects of autologous bone marrow mononuclear cell transplantation against elastase-induced emphysema in rabbit. Exp. Lung Res. 2006, 32, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.P.; Dutly, A.E.; Sacher, A.; Lee, H.; Hwang, D.M.; Liu, M.; Keshavjee, S.; Hu, J.; Waddell, T.K. Targeted cell replacement with bone marrow cells for airway epithelial regeneration. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, L740–L752. [Google Scholar] [CrossRef] [PubMed]
- Zhen, G.; Liu, H.; Gu, N.; Zhang, H.; Xu, Y.; Zhang, Z. Mesenchymal stem cells transplantation protects against rat pulmonary emphysema. Front. Biosci. 2008, 13, 3415–3422. [Google Scholar] [CrossRef] [PubMed]
- Katsha, A.M.; Ohkouchi, S.; Xin, H.; Kanehira, M.; Sun, R.; Nukiwa, T.; Saijo, Y. Paracrine factors of multipotent stromal cells ameliorate lung injury in an elastase-induced emphysema model. Mol. Ther. 2010, 19, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, K.S.; Johnstone, B.H.; Garrison, J.; Rush, N.I.; Cooper, S.; Traktuev, D.O.; Feng, D.; Adamowicz, J.J.; van Demark, M.; Fisher, A.J.; et al. Adipose stem cell treatment in mice attenuates lung and systemic injury induced by cigarette smoking. Am. J. Respir. Crit. Care Med. 2011, 183, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Furuya, N.; Takenaga, M.; Ohta, Y.; Tokura, Y.; Hamaguchi, A.; Sakamaki, A.; Kida, H.; Handa, H.; Nishine, H.; Mineshita, M.; et al. Cell therapy with adipose tissue-derived stem/stromal cells for elastase-induced pulmonary emphysema in rats. Regen. Med. 2012, 7, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Cruz, F.F.; Antunes, M.A.; Abreu, S.C.; Fujisaki, L.C.; Silva, J.D.; Xisto, D.G.; Maron-Gutierrez, T.; Ornellas, D.S.; Sá, V.K.; Rocha, N.N.; et al. Protective effects of bone marrow mononuclear cell therapy on lung and heart in an elastase-induced emphysema model. Respir. Physiol. Neurobiol. 2012, 182, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.J.; Song, L.; Han, F.F.; Cui, Z.L.; Chen, X.; Guo, X.J.; Xu, W.G. Mesenchymal stem cells protect cigarette smoke-damaged lung and pulmonary function partly via VEGF-VEGF receptors. J. Cell. Biochem. 2013, 114, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xu, A.; Xu, Q.; Zhao, W.; Li, D.; Fang, X.; Ren, Y. Bone marrow mesenchymal stem cell transplantation for treatment of emphysemic rats. Int. J. Clin. Exp. Med. 2014, 7, 968–972. [Google Scholar] [PubMed]
- Antunes, M.A.; Abreu, S.C.; Cruz, F.F.; Teixeira, A.C.; Lopes-Pacheco, M.; Bandeira, E.; Olsen, P.C.; Diaz, B.L.; Takyia, C.M.; Freitas, I.P.; et al. Effects of different mesenchymal stromal cell sources and delivery routes in experimental emphysema. Respir. Res. 2014, 15, 118. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Y.; Yeung, S.C.; Liang, Y.; Liang, X.; Ding, Y.; Ip, M.S.; Tse, H.F.; Mak, J.C.; Lian, Q. Mitochondrial transfer of induced pluripotent stem cell-derived mesenchymal stem cells to airway epithelial cells attenuates cigarette smoke-induced damage. Am. J. Respir. Cell Mol. Biol. 2014, 51, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, A.; Feizpour, A.; Hashemzahi, M.; Gholami, L.; Hosseini, M.; Soukhtanloo, M.; Bagheri, F.V.; Khodaei, E.; Roshan, N.M.; Boskabady, M.H. The effect of adipose derived stromal cells on oxidative stress level, lung emphysema and white blood cells of guinea pigs model of chronic obstructive pulmonary disease. Daru 2014, 22, 26. [Google Scholar] [CrossRef] [PubMed]
- Tibboel, J.; Keijzer, R.; Irwin Reiss, I.; de Jongste, J.C.; Post, M. Intravenous and intratracheal mesenchymal stromal cell injection in a mouse model of pulmonary emphysema. Int. J. Chronic Obstr. Pulm. Dis. 2014, 11, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Zarogoulidis, P.; Hohenforst-Schmidt, W.; Huang, H.; Sahpatzidou, D.; Freitag, L.; Sakkas, L.; Rapti, A.; Kioumis, I.; Pitsiou, G.; Kouzi-Koliakos, K.; et al. A gene therapy induced emphysema model and the protective role of stem cells. Diagn. Pathol. 2014, 9, 195. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gu, C.; Xu, W.; Yan, J.; Xia, Y.; Ma, Y.; Chen, C.; He, X.; Tao, H. Therapeutic effects of amniotic fluid-derived mesenchymal stromal cells on lung injury in rats with emphysema. Respir. Res. 2014, 15, 120. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Song, L.; Li, X.M.; Di, W.; Wang, X.J.; Xu, W.G. Mesenchymal stem cells alleviate airway inflammation and emphysema in COPD through down-regulation of cyclooxygenase-2 (COX-2) via p38 and ERK MAPK pathways. Sci. Rep. 2015, 5, 8733. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.B.; Lan, Y.W.; Chen, L.G.; Huang, T.T.; Choo, K.B.; Cheng, W.T.; Lee, H.S.; Chong, K.Y. Mesenchymal stem cell-based HSP70 promoter-driven VEGFA induction by resveratrol alleviates elastase-induced emphysema in a mouse model. Cell Stress Chaperones 2015, 20, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, J.Y.; Huh, J.W.; Lee, S.W.; Choi, S.J.; Oh, Y.M. The therapeutic effects of optimal dose of mesenchymal stem cells in a murine model of an elastase induced-emphysema. Tuberc. Respir. Dis. 2015, 78, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Peron, J.P.; de Brito, A.A.; Pelatti, M.; Brandão, W.N.; Vitoretti, L.B.; Greiffo, F.R.; da Silveira, E.C.; Oliveira-Junior, M.C.; Maluf, M.; Evangelista, L.; et al. Human tubal-derived mesenchymal stromal cells associated with low level laser therapy significantly reduces cigarette smoke-induced COPD in C57BL/6 mice. PLoS ONE 2015, 10, e0136942. [Google Scholar]
- Shigemura, N.; Okumura, M.; Mizuno, S.; Imanishi, Y.; Nakamura, T.; Sawa, Y. Autologous transplantation of adipose tissue-derived stromal cells ameliorates pulmonary emphysema. Am. J. Transplant. 2006, 6, 2592–2600. [Google Scholar] [CrossRef] [PubMed]
- Kennelly, H.; Mahon, B.P.; English, K. Human mesenchymal stromal cells exert HGF dependent cytoprotective effects in a human relevant pre-clinical model of COPD. Sci. Rep. 2016, 6, 38207. [Google Scholar] [CrossRef] [PubMed]
- Antunes, M.A.; Laffey, J.G.; Pelosi, P.; Rocco, P.R.M. Mesenchymal stem cell trials for pulmonary diseases. J. Cell. Biochem. 2014, 115, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.Gov. Available online: https://clinicaltrials.gov (accessed on 17 October 2017).
- Weiss, D.J.; Casaburi, R.; Flannery, R.; LeRoux-Williams, M.; Tashkin, D.P. A placebo-controlled, randomized trial of mesenchymal stem cells in COPD. Chest 2013, 143, 1590–1598. [Google Scholar] [CrossRef] [PubMed]
- Stessuk, T.; Ruiz, M.A.; Greco, O.T.; Bilaqui, A.; de Oliveira Ribeiro-Paes, M.J.; Ribeiro-Paes, J.T. Phase I clinical trial of cell therapy in patients with advanced chronic obstructive pulmonary disease: Follow-up for up to 3 years. Rev. Bras. Hematol. Hemoter. 2013, 35, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.S.; Hung, S.C.; Peng, S.T.; Huang, C.C.; Wei, H.M.; Guo, Y.J.; Fu, Y.S.; Lai, M.C.; Chen, C.C. Mesenchymal stem cells in the Wharton’s jelly of the human umbilical cord. Stem Cells 2004, 22, 1330–1337. [Google Scholar] [CrossRef] [PubMed]
- Pappa, K.I.; Anagnou, N.P. Novel sources of fetal stem cells: Where do they fit on the developmental continuum? Regen. Med. 2009, 4, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Kalaszczynska, I.; Ferdyn, K. Wharton’s jelly derived mesenchymal stem cells: Future of regenerative medicine? Recent findings and clinical significance. BioMed Res. Int. 2015, 2015, 430847. [Google Scholar] [CrossRef] [PubMed]
- Stolzing, A.; Jones, E.; McGonagle, D.; Scutt, A. Age-related changes in human bone marrow-derived mesenchymal stem cells: Consequences for cell therapies. Mech. Ageing Dev. 2008, 129, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Van Zglinicki, T.; Saretzki, G.; Docke, W.; Lotze, C. Mild hyperoxia shortens telomeres and inhibits proliferation: A model for senescence? Exp. Cell Res. 1995, 220, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhou, Z.W.; Ju, Z.; Wang, Z.Q. DNA damage response in hematopoietic stem cell ageing. Genom. Proteom. Bioinform. 2016, 14, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Oñate, B.; Vilahur, G.; Camino-López, S.; Díez-Caballero, A.; Ballesta-López, C.; Ybarra, J.; Moscatiello, F.; Herrero, J.; Badimon, L. Stem cells isolated from adipose tissue of obese patients show changes in their transcriptomic profile that indicate loss in stemcellness and increased commitment to an adipocyte-like phenotype. BMC Genom. 2013, 14, 625. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Raabe, O.; Addicks, K.; Wenisch, S.; Arnhold, S. Effects of non-steroidal anti-inflammatory drugs on proliferation, differentiation and migration in equine mesenchymal stem cells. Cell Biol. Int. 2011, 35, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Baksh, D.; Yao, R.; Tuan, R.S. Comparison of proliferative and multilineage differentiation potential of human mesenchymal stem cells derived from umbilical cord and bone marrow. Stem Cells 2007, 25, 1384–1392. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.F.; Wang, N.N.; Liu, Y.S.; Wei, X. Differentiation of Wharton’s jelly primitive stromal cells into insulin-producing cells in comparison with bone marrow mesenchymal stem cells. Tissue Eng. Part A 2009, 15, 2865–2873. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, S.J.; Jahnavi, V.S. Wharton’s jelly mesenchymal stem cells as off-the-shelf cellular therapeutics: A closer look into their regenerative and immunomodulatory properties. Open Tissue Eng. Regen. Med. J. 2011, 4, 28–38. [Google Scholar] [CrossRef]
- Riezzo, I.; Pascale, N.; la Russa, R.; Liso, A.; Salerno, M.; Turillazzi, E. Donor selection for allogenic hemopoietic stem cell transplantation: Clinical and ethical considerations. Stem Cells Int. 2017, 2017, 5250790. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.R.; Pfeifer, K.; Petry, F.; Powell, N.; Delzeit, J.; Weiss, M. Standardizing umbilical cord mesenchymal stromal cells for translation to clinical use: Selection of GMP Compliant medium and a simplified isolation method. Stem Cells Int. 2016, 14. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.R.; Cromer, A.; Weiss, M.L. Human umbilical cord mesenchymal stromal cell isolation, expansion, cryopreservation, and characterization. Curr. Protoc. Stem Cell Biol. 2017, 41. [Google Scholar] [CrossRef]
- Rao, C.V.; Li, X.; Toth, P.; Lei, Z.M. Expression of epidermal growth factor, transforming growth factor-α, and their common receptor genes in human umbilical cords. J. Clin. Endocrinol. Metab. 1995, 80, 1012–1020. [Google Scholar] [PubMed]
- Palka, J.; Bankowski, E.; Jaworski, S. An accumulation of IGF-I and IGF-binding proteins in human umbilical cord. Mol. Cell. Biochem. 2000, 206, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, R.; lo Iacono, M.; Corrao, S.; Magno, F.; Loria, T.; Cappello, F.; Zummo, G.; Farina, F.; la Rocca, G. New emerging potentials for human Wharton’s jelly mesenchymal stem cells: Immunological features and hepatocyte-like differentiative capacity. Stem Cells Dev. 2010, 19, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S.; Chang, Y.S.; Choi, S.J.; Kim, J.K.; Yoo, H.S.; Ahn, S.Y.; Sung, D.K.; Kim, S.Y.; Park, Y.R.; Park, W.S. Intratracheal transplantation of human umbilical cord blood-derived mesenchymal stem cells attenuates Escherichia coli-induced acute lung injury in mice. Respir. Res. 2011, 12, 108. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, D.; Liu, X.; Tang, S.; Wei, F. Human umbilical cord mesenchymal stem cells reduce systemic inflammation and attenuate LPS-induced acute lung injury in rats. J. Inflamm. 2012, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Yu, X.; Wang, Z.; Wang, F.; Wang, L.; Gao, H.; Chen, Y.; Zhao, W.; Jia, Z.; Yan, S.; et al. Long term effects of the implantation of Wharton’s jelly-derived mesenchymal stem cells from the umbilical cord for newly-onset type 1 diabetes mellitus. Endocr. J. 2013, 60, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zheng, P.; Wang, X.; Dai, G.; Cheng, H.; Zhang, Z.; Hua, R.; Niu, X.; Shi, J.; An, Y. A preliminary evaluation of efficacy and safety of Wharton’s jelly mesenchymal stem cell transplantation in patients with type 2 diabetes mellitus. Stem Cell Res. Ther. 2014, 5, 57. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, J.; Zhang, Y.; Zhang, M.; Chen, J.; Li, X.; Hu, X.; Jiang, S.; Shi, S.; Sun, L. Umbilical cord mesenchymal stem cell transplantation in active and refractory systemic lupus erythematosus: A multicenter clinical study. Arthritis Res. Ther. 2014, 16, R7. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, F.; Gu, B.; Chen, G.; Chang, H.; Wu, D. Mesenchymal stromal cells as an adjuvant treatment for severe late-onset hemorrhagic cystitis after allogeneic hematopoietic stem cell transplantation. Acta Haematol. 2015, 133, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Kean, T.J.; Lin, P.; Caplan, A.I.; Dennis, J.E. MSCs: Delivery routes and engraftment, cell-targeting strategies, and immune modulation. Stem Cells Int. 2013, 2013, 732742. [Google Scholar] [CrossRef] [PubMed]
- Curley, G.F.; Ansari, B.; Hayes, M.; Devaney, J.; Masterson, C.; Ryan, A.; Barry, F.; O’Brien, T.; Toole, D.O.; Laffey, J.G. Effects of intratracheal mesenchymal stromal cell therapy during recovery and resolution after ventilator-induced lung injury. Anesthesiology 2013, 118, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Montecino-Rodriguez, E.; Berent-Maoz, B.; Dorshkind, K. Causes, consequences, and reversal of immune system aging. J. Clin. Investig. 2013, 123, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Pawitan, J.A. Prospect of stem cell conditioned medium in regenerative medicine. BioMed Res. Int. 2014, 2014, 965849. [Google Scholar] [CrossRef] [PubMed]
- Akram, K.M.; Patel, N.; Spiteri, M.A.; Forsyth, N.R. Lung Regeneration: Endogenous and Exogenous Stem Cell Mediated Therapeutic Approaches. Int. J. Mol. Sci. 2016, 17, 128. [Google Scholar] [CrossRef] [PubMed]
- Garcia, O.; Carraro, G.; Navarro, S.; Bertoncello, I.; McQualter, J.; Driscoll, B.; Jesudason, E.; Warburton, D. Cell-based therapies for lung disease. Br. Med. Bull. 2012, 101, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Foronjy, R.F.; Majka, S.M. The potential for resident lung mesenchymal stem cells to promote functional tissue regeneration: Understanding microenvironmental cues. Cells 2012, 1, 874–885. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, C.; Gilbert, J.L.; O’Dea, S.; O’Brien, F.J.; Cryan, S.A. Respiratory Tissue Engineering: Current Status and Opportunities for the Future. Tissue Eng. Part B Rev. 2015, 21, 323–344. [Google Scholar] [CrossRef] [PubMed]
- Schilders, K.A.A.; Eenjes, E.; van Riet, S.; Poot, A.A.; Stamatialis, D.; Truckenmüller, R.; Hiemstra, P.S.; Rottier, R.J. Regeneration of the lung: Lung stem cells and the development of lung mimicking devices. Respir. Res. 2016, 17, 44. [Google Scholar] [CrossRef] [PubMed]
- Summer, R.; Fitzsimmons, K.; Dwyer, D.; Murphy, J.; Fine, A. Isolation of an adult mouse lung mesenchymal progenitor cell population. Am. J. Respir. Cell Mol. Biol. 2007, 37, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Ampollini, L.; Madeddu, D.; Falco, A.; Frati, C.; Lorusso, B.; Graiani, G.; Saccani, F.; Gervasi, A.; Rossetti, P.; Bonomini, S.; et al. Lung mesenchymal cells function as an inductive microenvironment for human lung cancer propagating cells dagger. Eur. J. Cardiothorac. Surg. 2014, 46, e103–e112. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Animal/Model | MSC Type/Mode of Administration | Outcome/Potential Mechanism of Action | Reference |
|---|---|---|---|
| Rabbit elastase-induced | BM-MC/IT | ↓ cell count in bronchoalveolar lavage fluid ↓ apoptotic cells and MMP-2 ↑ number of proliferative (Ki-67-positive) alveolar cells | Yuhgetsu et al., 2006 [57] |
| Mouse naphthalene-induced | BM-MSCs/IV | ↑ regeneration of airway epithelial cells ↑ GFP transgene expression targeted delivery | Wong et al., 2007 [58] |
| Rat papain-induced | BM-MSCs/IT | ↑ Bcl-2 and Bax differentiation of MSCs into type II alveolar epithelial cells ↓ alveolar cell apoptosis | Zhen et al., 2008 [59] |
| Mouse bleomycin-induced | hMSCs/IN | ↓ TGFβ-1, MMIF, TNF-α ↓ collagen concentration in the lung ↓ Smad2 phosphorylation (transforming growth factor-beta activity) ↑ MMP2 ↓ fibrosis | Moodley et al., 2009 [42] |
| Rat papain-induced | BM-MSC/IV | ↓ alveolar enlargement ↓ apoptosis ↑ VEGF-A | Zhen et al. 2010 [48] |
| Mouse elastase-induced | BM-MSC/IT | ↑ HGF ↑ EGF ↑ SLPI | Katsha et al., 2010 [60] |
| Rat cigarette smoke-induced | BM-MC/IV | ↓ apoptosis ↑ number of small pulmonary vessels ↓ pulmonary arterial pressure | Huh et al., 2011 [49] |
| Mouse cigarette smoke-induced | hAD-SC/IV mAD-SC/IV | ↑ number of macrophages and polymorphonuclear leukocytes in the BAL, caspase-3 ↓ alveolar space size ↑ MAPK signal transduction pathways involved in inflammation and apoptosis ↓ VEGF | Schweitzer et al., 2011 [61] |
| Rat elastase-induced | AD-SC/SCI | ↓ alveolar airspaces ↑ HGF, CINC-1 ↑ angiogenesis ↑ IL-1β | Furuya et al., 2012 [62] |
| Mouse elastase-induced | BM-MC/IV | ↓ neutrophil infiltration, elastolysis, collagen fiber deposition ↓ lung cell apoptosis ↑ HGF, IGF ↓ PDGF, TGFβ-1, caspase-3 | Cruz et al., 2012 [63] |
| Rat cigarette smoke-induced | BM-MSCs/IT hBM-MSCs in vitro | ↓ TNF-α, IL-1β, MCP-1, IL-6 ↓ MMP9, MMP12 ↑ VEGF, VEGF receptor 2, TGFβ-1 ↓ pulmonary cell apoptosis | Guan et al., 2013 [64] |
| Rat LPS-induced | BM-MSCs/IV | ↑ alveoli epithelial cells number | Zhao et al., 2014 [65] |
| Mouse elastase-induced | BM-MSCs, AD-MSC or lung tissue (LMSC)/IV/IT | ↓ neutrophil infiltration, cell apoptosis ↑ elastic fiber content ↓ alveolar epithelial and endothelial cell damage ↓ keratinocyte-derived chemokine (KC, a mouse analog of interleukin-8), TGFβ-1 ↓ alveolar hyperinflation (BM-MSCs), collagen fiber content (BM-MSCs and L-MSC) ↓ M1 macrophages and pulmonary hypertension ↑ VEGF | Antunes at al., 2014 [66] |
| Rat cigarette smoke-induced | iPSC-MSCs/IV BM-MSCs/IV | ↑ adenosine triphosphate | Li et al., 2014 [67] |
| Guinea pig cigarette smoke-induced | ADSCs/IV/IT | ↑ antioxidant effects ↓ apoptotic cells ↓ oxidative damage weight restoration | Ghorbani et al., 2014 [68] |
| Mouse elastase-induced | BM-MSCs/IV/IT | ↑ IL-6, keratinocyte-derived-chemokine (KC) ↓ MCP levels at day 2 after elastase injection | Tibboel et al., 2014 [69] |
| Mouse DDMC, non-viral vector | hMSCs/IT | ↓ emphysema ↑ cells within the lung parenchyma | Zarogoulidis et al., 2014 [70] |
| Rat cigarette smoke-induced | AFMSCs/IT | ↑ SPA, ↑ SPC ↑ TTF-1 ↓ AECII apoptosis ↓ lung injury | Li et al., 2014 [71] |
| Rat cigarette smoke-induced | BM-MSCs/IT | ↓ COX-2, COX-2-mediated prostaglandin E2 (PGE2) in macrophages through inhibition of phosphorylation of p38 MAPK and ERK-activation | Gu et al., 2015 [72] |
| Mouse elastase-induced | BD-MSCs/IV | ↓ VEGF-A ↓ HO-1 | Chen et al., 2015 [73] |
| Mouse elastase-induced | hMSCs/IV | ↓ MMP-9 ↑ VEGF ↑ IL-1β, INF-γ, IL-2 | Kim et al., 2015 [74] |
| Mouse cigarette smoke-induced | htMSCs/IV | ↓ lung inflammation ↓ IL-1β, IL-6, TNF-α, KC - (C-X-C motif) ligand 1 (CXCL1) ↓ mucus production, collagen accumulation, tissue damage ↓ NF-κB ↑ IL-10 | Peron et al., 2015 [75] |
| Rat elastase-induced | ADSCs/IV | ↑ HGF expression in lung tissues ↑ alveolar and vascular regeneration ↓ alveolar cell apoptosis ↑ VEGF, HGF, bFGF | Shigemura et al., 2016 [76] |
| Mouse elastase-induced | hBM-MSCs/IV | ↑ HGF | Kennelly et al., 2016 [77] |
| Nct Number | Time Frame | Msctype | Msc Source | Delivery Route | MSC Dose | Application Schedule | No. of Patients | Follow-up Period | Trial Status | Studyphase | Study Location |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NCT00683722 | 2008–2010 | BM-MSC | autologous | IV | 1 × 108 | Four monthly | 62 | 2 years | Completed | II | USA |
| NCT01110252 | 2010–2011 | BM-MC | autologous | IV | 1 × 108 | Single dose | 4 | 1 year | Completed | * | Brazil |
| NCT01306513 | 2010–2012 | BM-MSC | autologous | IV | * | Twice weekly | 10 | 8 weeks | Completed | I | Netherlands |
| NCT01849159 | 2014–2017 | BM-MSC | autologous | IV | 2 × 108 | Every 2 mo for 1 year | 30 | 2 years | Unknown | I/II | Russia |
| NCT01872624 | 2013–2015 | BM-MSC | autologous | EB | * | Single dose | 10 | 4 months | Completed | * | Brazil |
| NCT01758055 | 2012–2014 | BM-MC | autologous | EB | 60 × 106 | Single dose | 12 | 1 year | Unknown | I | Iran |
| NCT02041000 | 2014 | ADSC | autologous | * | * | * | 0 | 6 months | Withdrawn | * | USA |
| NCT01559051 | 2014–2017 | AD-SVF | autologous | IV/IN | * | Single dose | 100 | 6 months | Recruiting | I/II | USA |
| NCT02161744 | 2014–2017 | AD-SVF | autologous | IV | * | Single dose | 60 | 1 year | Recruiting | I | USA |
| NCT02216630 | 2014–2017 | AD-SVF | autologous | IV | * | Single dose | 26 | 1 year | Completed | I/II | USA |
| NCT02135380 | 2014–2015 | AD-SVF | autologous | IV | * | Single dose | 60 | 1 year | Unknown | I/II | India |
| AD-MSC | autologous | IV | 2 × 106/kg b.m. | Three doses | |||||||
| NCT02645305 | 2015–2016 | ADSC & PRP | autologous | IV | * | Single dose | 20 | 1 year | Recruiting | I/II | Vietnam |
| NCT02348060 | 2015–2018 | AD-SVF | autologous | IV | * | Single dose | 75 | 1 year | Recruiting | * | USA |
| NCT03044431 | 2016–2018 | BM-MC & PRP | autologous | IV | * | Single dose | 214 | 6 months | Active, not recruiting | * | USA |
| NCT02412332 | 2015–2017 | BM-MC | autologous | IV | 1 × 108 | Single dose | 20 | 1 year | Enrolling | I/II | Brazil |
| ADSC | 1 × 108 | ||||||||||
| BM-MC +ADSC | 5 × 107 + 5 × 107 | ||||||||||
| NCT03228121 | 2017–2018 | PBSC & PRP | autologous | IV | * | Three doses | 100 | 1 year | Enrolling | * | USA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janczewski, A.M.; Wojtkiewicz, J.; Malinowska, E.; Doboszyńska, A. Can Youthful Mesenchymal Stem Cells from Wharton’s Jelly Bring a Breath of Fresh Air for COPD? Int. J. Mol. Sci. 2017, 18, 2449. https://doi.org/10.3390/ijms18112449
Janczewski AM, Wojtkiewicz J, Malinowska E, Doboszyńska A. Can Youthful Mesenchymal Stem Cells from Wharton’s Jelly Bring a Breath of Fresh Air for COPD? International Journal of Molecular Sciences. 2017; 18(11):2449. https://doi.org/10.3390/ijms18112449
Chicago/Turabian StyleJanczewski, Andrzej M., Joanna Wojtkiewicz, Ewa Malinowska, and Anna Doboszyńska. 2017. "Can Youthful Mesenchymal Stem Cells from Wharton’s Jelly Bring a Breath of Fresh Air for COPD?" International Journal of Molecular Sciences 18, no. 11: 2449. https://doi.org/10.3390/ijms18112449
APA StyleJanczewski, A. M., Wojtkiewicz, J., Malinowska, E., & Doboszyńska, A. (2017). Can Youthful Mesenchymal Stem Cells from Wharton’s Jelly Bring a Breath of Fresh Air for COPD? International Journal of Molecular Sciences, 18(11), 2449. https://doi.org/10.3390/ijms18112449