
Stem Cells and Labeling for Spinal Cord Injury

 , ,
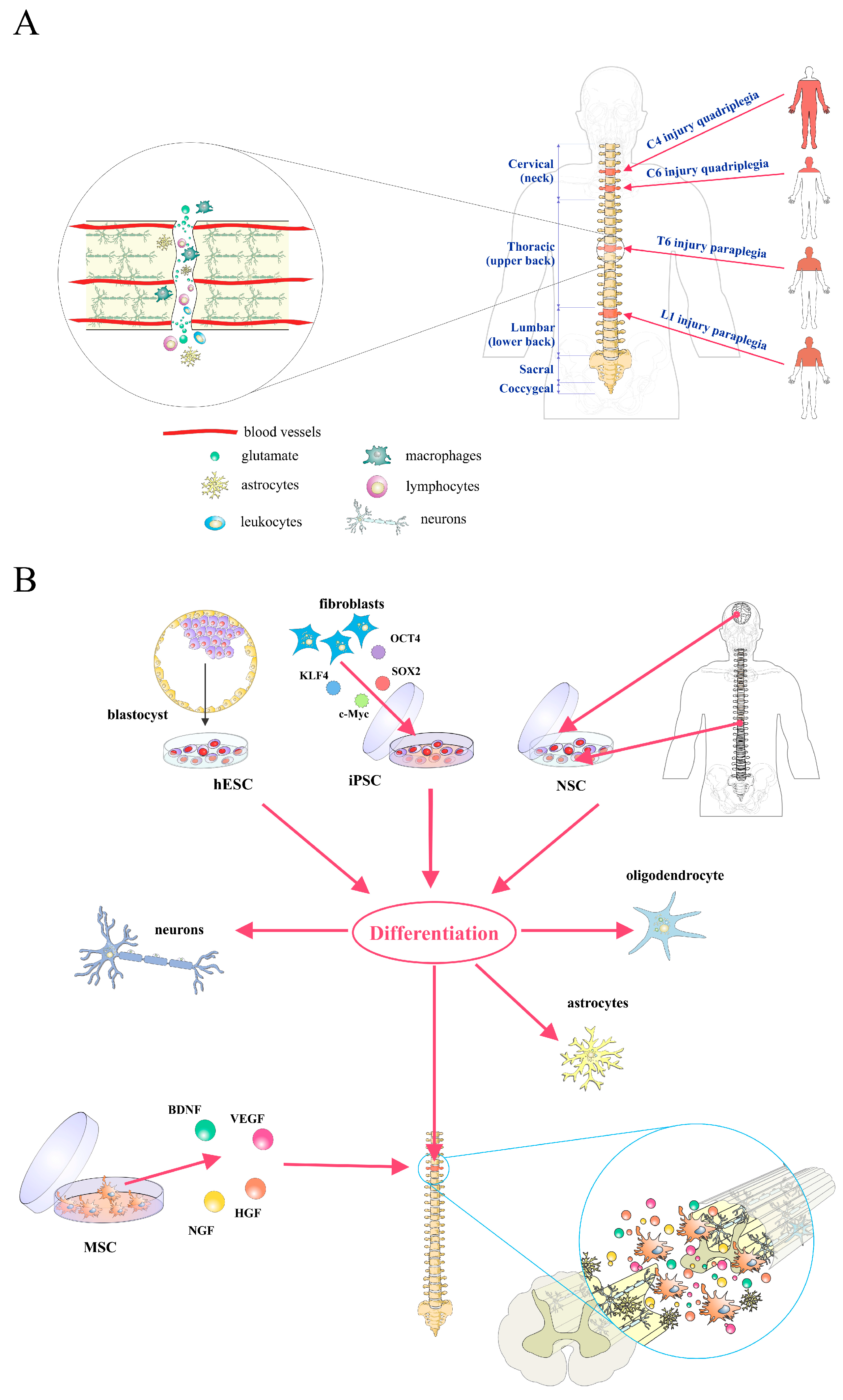
, , 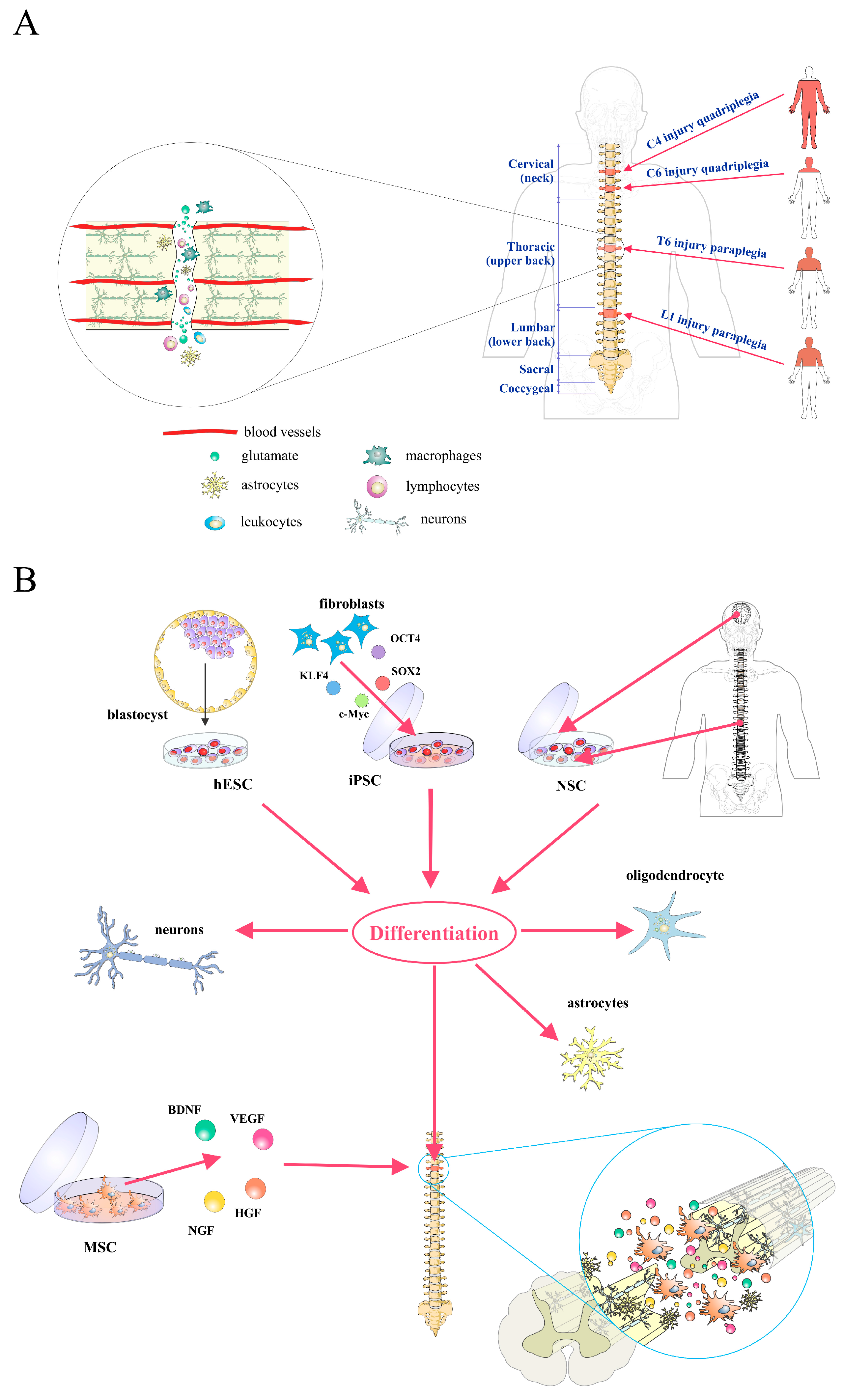{kind=link}
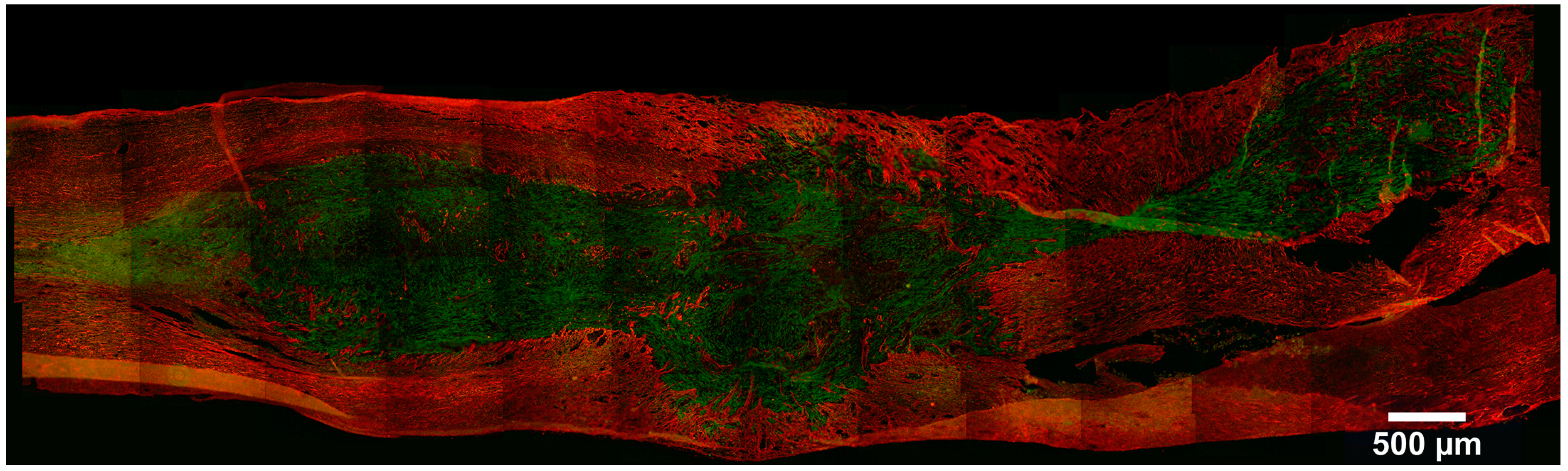
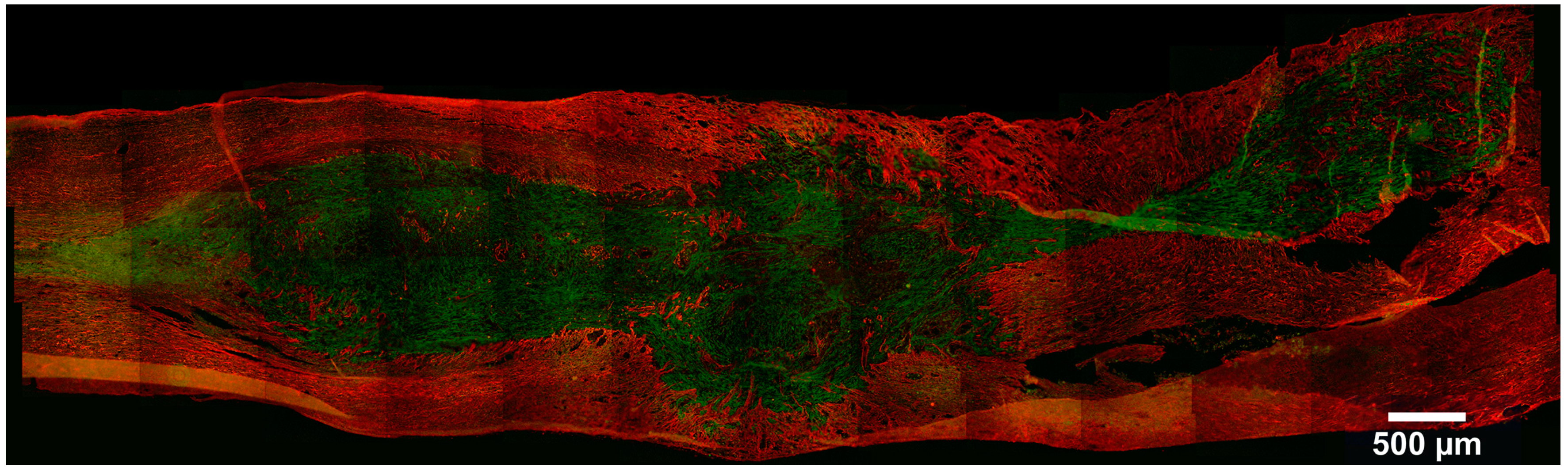{kind=link}
Abstract
:1. Introduction
2. Pathophysiology of Spinal Cord Injury
3. Stem/Progenitor Cell Therapy for Spinal Cord Injury
4. Stem Cell Labeling and Tracking
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ronaghi, M.; Erceg, S.; Moreno-Manzano, V.; Stojkovic, M. Challenges of stem cell therapy for spinal cord injury: Human embryonic stem cells, endogenous neural stem cells, or induced pluripotent stem cells? Stem Cells 2010, 28, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Tetreault, L.; Kalsi-Ryan, S.; Nouri, A.; Fehlings, M.G. Global prevalence and incidence of traumatic spinal cord injury. Clin. Epidemiol. 2014, 6, 309–331. [Google Scholar] [PubMed]
- National Spinal Cord Injury Statistical Center. Spinal cord injury (SCI) 2016 facts and figures at a glance. J. Spinal Cord Med. 2016, 39, 493–494. [Google Scholar]
- Goel, A. Stem cell therapy in spinal cord injury: Hollow promise or promising science? J. Craniovertebr. Junction Spine 2016, 7, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Beattie, M.S.; Li, Q.; Bresnahan, J.C. Cell death and plasticity after experimental spinal cord injury. Prog. Brain Res. 2000, 128, 9–21. [Google Scholar] [PubMed]
- Blight, A.R. Spinal cord injury models: Neurophysiology. J. Neurotrauma 1992, 9, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Grossman, S.D.; Rosenberg, L.J.; Wrathall, J.R. Relationship of altered glutamate receptor subunit mRNA expression to acute cell loss after spinal cord contusion. Exp. Neurol. 2001, 168, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Oyinbo, C.A. Secondary injury mechanisms in traumatic spinal cord injury: A nugget of this multiply cascade. Acta Neurobiol. Exp. (Wars) 2011, 71, 281–299. [Google Scholar] [PubMed]
- Pineau, I.; Lacroix, S. Proinflammatory cytokine synthesis in the injured mouse spinal cord: Multiphasic expression pattern and identification of the cell types involved. J. Comp. Neurol. 2007, 500, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Lukovic, D.; Stojkovic, M.; Moreno-Manzano, V.; Jendelova, P.; Sykova, E.; Bhattacharya, S.S.; Erceg, S. Concise review: Reactive astrocytes and stem cells in spinal cord injury: Good guys or bad guys? Stem Cells 2015, 33, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Pekny, M.; Nilsson, M. Astrocyte activation and reactive gliosis. Glia 2005, 50, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Silver, J.; Miller, J.H. Regeneration beyond the glial scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Lukovic, D.; Valdés-Sanchez, L.; Sanchez-Vera, I.; Moreno-Manzano, V.; Stojkovic, M.; Bhattacharya, S.S.; Erceg, S. Brief report: Astrogliosis promotes functional recovery of completely transected spinal cord following transplantation of hESC-derived oligodendrocyte and motoneuron progenitors. Stem Cells 2014, 32, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Erceg, S.; Laínez, S.; Ronaghi, M.; Stojkovic, P.; Pérez-Aragó, M.A.; Moreno-Manzano, V.; Moreno-Palanques, R.; Planells-Cases, R.; Stojkovic, M. Differentiation of human embryonic stem cells to regional specific neural precursors in chemically defined medium conditions. PLoS ONE 2008, 3, e2122. [Google Scholar] [CrossRef] [PubMed]
- Volarevic, V.; Erceg, S.; Bhattacharya, S.S.; Stojkovic, P.; Horner, P.; Stojkovic, M. Stem cell-based therapy for spinal cord injury. Cell Transplant. 2013, 22, 1309–1323. [Google Scholar] [CrossRef] [PubMed]
- Lukovic, D.; Moreno-Manzano, V.; Lopez-Mocholi, E.; Rodriguez-Jiménez, F.J.; Jendelova, P.; Sykova, E.; Oria, M.; Stojkovic, M.; Erceg, S. Complete rat spinal cord transection as a faithful model of spinal cord injury for translational cell transplantation. Sci. Rep. 2015, 5, 9640. [Google Scholar] [CrossRef] [PubMed]
- Erceg, S.; Ronaghi, M.; Oria, M.; Roselló, M.G.; Aragó, M.A.; Lopez, M.G.; Radojevic, I.; Moreno-Manzano, V.; Rodríguez-Jiménez, F.J.; Bhattacharya, S.S.; et al. Transplanted oligodendrocytes and motoneuron progenitors generated from human embryonic stem cells promote locomotor recovery after spinal cord transection. Stem Cells 2010, 28, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Manzano, V.; Rodríguez-Jiménez, F.J.; García-Roselló, M.; Laínez, S.; Erceg, S.; Calvo, M.T.; Ronaghi, M.; Lloret, M.; Planells-Cases, R.; Sánchez-Puelles, J.M.; et al. Activated spinal cord ependymal stem cells rescue neurological function. Stem Cells 2009, 27, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Lako, M.; Armstrong, L.; Stojkovic, M. Induced pluripotent stem cells: It looks simple but can looks deceive? Stem Cells 2010, 28, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Angelos, M.G.; Kaufman, D.S. Pluripotent stem cell applications forregenerative medicine. Curr. Opin. Organ Transplant. 2015, 20, 663–670. [Google Scholar] [PubMed]
- Erceg, S.; Lukovic, D.; Moreno-Manzano, V.; Stojkovic, M.; Bhattacharya, S.S. Derivation of cerebellar neurons from human pluripotent stem cells. Curr. Protoc. Stem Cell Biol. 2012. [Google Scholar] [CrossRef]
- Hodgetts, S.I.; Edel, M.; Harvey, A.R. The state of play with iPSCs and spinal cord injury models. J. Clin. Med. 2015, 4, 193–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Lin, T.; Xu, Y. Stem cell therapy and immunological rejection in animal models. Curr. Mol. Pharmacol. 2015. [Google Scholar] [CrossRef]
- Lee-Kubli, C.A.; Lu, P. Induced pluripotent stem cell-derived neural stem cell therapies for spinal cord injury. Neural Regen. Res. 2015, 10, 10–16. [Google Scholar] [PubMed]
- Amemori, T.; Ruzicka, J.; Romanyuk, N.; Jhanwar-Uniyal, M.; Sykova, E.; Jendelova, P. Comparison of intraspinal and intrathecal implantation of induced pluripotent stem cell-derived neural precursors for the treatment of spinal cord injury in rats. Stem Cell Res. Ther. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, S.; Takano, M.; Numasawa-Kuroiwa, Y.; Itakura, G.; Kobayashi, Y.; Nishiyama, Y.; Sugai, K.; Nishimura, S.; Iwai, H.; Isoda, M.; et al. Grafted human iPScell-derived oligodendrocyte precursor cells contribute to robust remyelination of demyelinated axons after spinal cord injury. Stem Cell Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pomeshchik, Y.; Puttonen, K.A.; Kidin, I.; Ruponen, M.; Lehtonen, S.; Malm, T.; Åkesson, E.; Hovatta, O.; Koistinaho, J. Transplanted human induced pluripotent stem cell-derived neural progenitor cells do not promote functional recovery of pharmacologically immunosuppressed mice with contusion spinal cord injury. Cell Transplant. 2015, 24, 1799–1812. [Google Scholar] [CrossRef] [PubMed]
- Karimi-Abdolrezaee, S.; Eftekharpour, E.; Wang, J.; Morshead, C.M.; Fehlings, M.G. Delayed transplantation of adult neural precursor cells promotes remyelination and functional neurological recovery after spinal cordinjury. J. Neurosci. 2006, 26, 3377–3389. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Villafuertes, R.; Rodríguez-Jiménez, F.J.; Alastrue-Agudo, A.; Stojkovic, M.; Miras-Portugal, M.T.; Moreno-Manzano, V. Purinergic receptors in spinal cord-derived ependymal stem/progenitor cells and their potential role in cell-based therapy for spinal cord injury. Cell Transplant. 2015, 24, 1493–1509. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Jimenez, F.J.; Alastrue-Agudo, A.; Stojkovic, M.; Erceg, S.; Moreno-Manzano, V. Connexin 50 expression in ependymal stem progenitor cells after spinal cord injury activation. Int. J. Mol. Sci. 2015, 16, 26608–26618. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Jiménez, F.J.; Alastrue-Agudo, A.; Erceg, S.; Stojkovic, M.; Moreno-Manzano, V. FM19G11 favors spinal cord injury regeneration and stem cell self-renewal by mitochondrial uncoupling and glucose metabolism induction. Stem Cells 2012, 30, 2221–2233. [Google Scholar] [CrossRef] [PubMed]
- Valdes-Sánchez, T.; Rodriguez-Jimenez, F.J.; García-Cruz, D.M.; Escobar-Ivirico, J.L.; Alastrue-Agudo, A.; Erceg, S.; Monleón, M.; Moreno-Manzano, V. Methacrylate-endcapped caprolactone and FM19G11 provide a proper niche for spinal cord-derived neural cells. J. Tissue Eng. Regen. Med. 2015, 9, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Manzano, V.; Rodríguez-Jiménez, F.J.; Aceña-Bonilla, J.L.; Fustero-Lardíes, S.; Erceg, S.; Dopazo, J.; Montaner, D.; Stojkovic, M.; Sánchez-Puelles, J.M. FM19G11, a new hypoxia-inducible factor (HIF) modulator, affects stem cell differentiation status. J. Biol. Chem. 2010, 285, 1333–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazdic, M.; Volarevic, V.; Arsenijevic, N.; Stojkovic, M. Mesenchymal stem cells: A friend or foe in immune-mediated diseases. Stem Cell Rev. 2015, 11, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Ruff, C.A.; Wilcox, J.T.; Fehlings, M.G. Cell-based transplantation strategies to promote plasticity following spinal cord injury. Exp. Neurol. 2012, 235, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Radtke, C.; Tan, A.M.; Zhao, P.; Hamada, H.; Houkin, K.; Honmou, O.; Kocsis, J.D. BDNF-hypersecreting human mesenchymal stem cells promote functional recovery, axonal sprouting, and protection of corticospinal neurons after spinal cord injury. J. Neurosci. 2009, 29, 14932–14941. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.K.; Riegler, J.; Wu, J.C. Stem cell imaging: From bench to bedside. Cell Stem Cell 2014, 14, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Paspala, S.A.; Vishwakarma, S.K.; Murthy, T.V.; Rao, T.N.; Khan, A.A. Potential role of stem cells in severe spinal cord injury: Current perspectives and clinical data. Stem Cells Cloning 2012, 5, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.K.; Bulte, J.W. Seeing stem cells at work in vivo. Stem Cell Rev. 2014, 10, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lepski, G. Cell transplantation for spinal cord injury: A systematic review. BioMed Res. Int. 2013, 2013, 786475. [Google Scholar] [CrossRef] [PubMed]
- Callera, F.; de Melo, C.M. Magnetic resonance tracking of magnetically labeled autologous bone marrow CD34+ cells transplanted into the spinal cord via lumbar puncture technique in patients with chronic spinal cord injury: CD34+ cells’ migration into the injured site. Stem Cells Dev. 2007, 16, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Obenaus, A.; Dilmac, N.; Tone, B.; Tian, H.R.; Hartman, R.; Digicaylioglu, M.; Snyder, E.Y.; Ashwal, S. Long-term magnetic resonance imaging of stem cells in neonatal ischemic injury. Ann. Neurol. 2011, 69, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Guzman, R.; Uchida, N.; Bliss, T.M.; He, D.; Christopherson, K.K.; Stellwagen, D.; Capela, A.; Greve, J.; Malenka, R.C.; Moseley, M.E.; et al. Long-term monitoring of transplanted human neural stem cells in developmental and pathological contexts with MRI. Proc. Natl. Acad. Sci. USA 2007, 104, 10211–10216. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Lara, L.E.; Xu, X.; Hofstetrova, K.; Pniak, A.; Chen, Y.; McFadden, C.D.; Martinez-Santiesteban, F.M.; Rutt, B.K.; Brown, A.; Foster, P.J. The use of cellular magnetic resonance imaging to track the fate of iron-labeled multipotent stromal cells after direct transplantation in a mouse model of spinal cord injury. Mol. Imaging Biol. 2011, 13, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhou, L.; Xing Wu, F. Tracking neural stem cells in patients with brain trauma. N. Engl. J. Med. 2006, 355, 2376–2378. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.E.; Muja, N.; Fainstein, N.; Bulte, J.W.M.; Ben-Hur, T. Conserved fate and function of ferumoxides-labeled neural precursor cells in vitro and in vivo. J. Neurosci. Res. 2010, 88, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Amemori, T.; Romanyuk, N.; Jendelova, P.; Herynek, V.; Turnovcova, K.; Prochazka, P.; Kapcalova, M.; Cocks, G.; Price, J.; Sykova, E. Human conditionally immortalized neural stem cells improve locomotor function after spinal cord injury in the rat. Stem Cell Res. Ther. 2013, 4, 68. [Google Scholar] [CrossRef] [PubMed]
- Cromer Berman, S.M.; Kshitiz; Wang, C.J.; Orukari, I.; Levchenko, A.; Bulte, J.W.; Walczak, P. Cell motility of neural stem cells is reduced after SPIO-labeling, which is mitigated after exocytosis. Magn. Reson. Med. 2013, 69, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.L.; Lu, P.G.; Zhang, L.J.; Li, F.; Chen, Z.; Wu, N.; Meng, H.; Lin, J.K.; Feng, H. In Vivo magnetic resonance imaging tracking of SPIO-labeled human umbilical cord mesenchymal stem cells. J. Cell. Biochem. 2012, 113, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Urdzíková, L.; Jendelová, P.; Glogarová, K.; Burian, M.; Hájek, M.; Syková, E. Transplantation of bone marrow stem cells as well as mobilization by granulocyte-colony stimulating factor promotes recovery after spinal cord injury in rats. J. Neurotrauma 2006, 23, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Chotivichit, A.; Ruangchainikom, M.; Chiewvit, P.; Wongkajornsilp, A.; Sujirattanawimol, K. Chronic spinal cord injury treated with transplanted autologous bone marrow-derived mesenchymal stem cells tracked by magnetic resonance imaging: A case report. J. Med. Case Rep. 2015, 9, 79. [Google Scholar] [CrossRef] [PubMed]
- Vaněček, V.; Zablotskii, V.; Forostyak, S.; Růžička, J.; Herynek, V.; Babič, M.; Jendelová, P.; Kubinová, S.; Dejneka, A.; Syková, E. Highly efficient magnetic targeting of mesenchymal stem cells in spinal cord injury. Int. J. Nanomed. 2012, 7, 3719–3730. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.-P.; Xu, C.; Liu, Y.; Li, J.-D.; Xie, J. Visual bone marrow mesenchymal stem cell transplantation in the repair of spinal cord injury. Neural Regen. Res. 2015, 10, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, E.M.; Lamanna, J.; Boulis, N.M. Stem cell therapy for the spinal cord. Stem Cell Res. Ther. 2012, 3, 24. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Suzuki, Y.; Huang, M.; Cao, F.; Xie, X.; Connolly, A.J.; Yang, P.C.; Wu, J.C. Comparison of reporter gene and iron particle labeling for tracking fate of human embryonic stem cells and differentiated endothelial cells in living subjects. Stem Cells 2008, 26, 864–873. [Google Scholar] [CrossRef] [PubMed]
- DeHaro, J.; Zurita, M.; Ayllón, L.; Vaquero, J. Detection of 111In-oxine-labeled bone marrow stromal cells after intravenous or intralesional administration in chronic paraplegic rats. Neurosci. Lett. 2005, 377, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Li, S.C.; Tachiki, L.M.; Luo, J.; Dethlefs, B.A.; Chen, Z.; Loudon, W.G. A biological global positioning system: Considerations for tracking stem cell behaviors in the whole body. Stem Cell Rev. 2010, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.C.; Hsu, C.H.; Wu, A.T.; Yang, L.Y.; Chen, W.H.; Chiu, W.T.; Lai, W.F.; Wu, C.H.; Gelovani, J.G.; Deng, W.P. A novel cell-based therapy for contusion spinal cord injury using GDNFdelivering NIH3T3 cells with dual reporter genes monitored by molecular imaging. J. Nucl. Med. 2008, 49, 1512–1519. [Google Scholar] [CrossRef] [PubMed]
- Berman, S.C.; Galpoththawela, C.; Gilad, A.A.; Bulte, J.W.; Walczak, P. Long-term MR cell tracking of neural stem cells grafted in immunocompetent versus immunodeficient mice reveals distinct differences in contrast between live and dead cells. Magn. Reson. Med. 2011, 65, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Tsuji, O.; Kumagai, G.; Hara, C.M.; Okano, H.J.; Miyawaki, A.; Toyama, Y.; Okano, H.; Nakamura, M. Comparative study of methods for administering neural stem/progenitor cells to treat spinal cord injury in mice. Cell Transplant. 2011, 20, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Ishii, K.; Yamane, J.; Iwanami, A.; Ikegami, T.; Katoh, H.; Iwamoto, Y.; Nakamura, M.; Miyoshi, H.; Okano, H.J.; et al. In vivo imaging of engrafted neural stem cells: Its application in evaluating the optimal timing of transplantation for spinal cord injury. FASEB J. 2005, 19, 1839–1841. [Google Scholar] [CrossRef] [PubMed]
- Lukovic, D.; Moreno Manzano, V.; Stojkovic, M.; Bhattacharya, S.S.; Erceg, S. Concise review: Human pluripotent stem cells in the treatment of spinal cord injury. Stem Cells 2012, 30, 1787–1792. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Tian, M.; Zhang, H. Molecular imaging in stem cell therapy for spinal cord injury. BioMed Res. Int. 2014, 2014, 759514. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gazdic, M.; Volarevic, V.; Arsenijevic, A.; Erceg, S.; Moreno-Manzano, V.; Arsenijevic, N.; Stojkovic, M. Stem Cells and Labeling for Spinal Cord Injury. Int. J. Mol. Sci. 2017, 18, 6. https://doi.org/10.3390/ijms18010006
Gazdic M, Volarevic V, Arsenijevic A, Erceg S, Moreno-Manzano V, Arsenijevic N, Stojkovic M. Stem Cells and Labeling for Spinal Cord Injury. International Journal of Molecular Sciences. 2017; 18(1):6. https://doi.org/10.3390/ijms18010006
Chicago/Turabian StyleGazdic, Marina, Vladislav Volarevic, Aleksandar Arsenijevic, Slaven Erceg, Victoria Moreno-Manzano, Nebojsa Arsenijevic, and Miodrag Stojkovic. 2017. "Stem Cells and Labeling for Spinal Cord Injury" International Journal of Molecular Sciences 18, no. 1: 6. https://doi.org/10.3390/ijms18010006