Neuroprotective Effects of Citicoline in in Vitro Models of Retinal Neurodegeneration

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
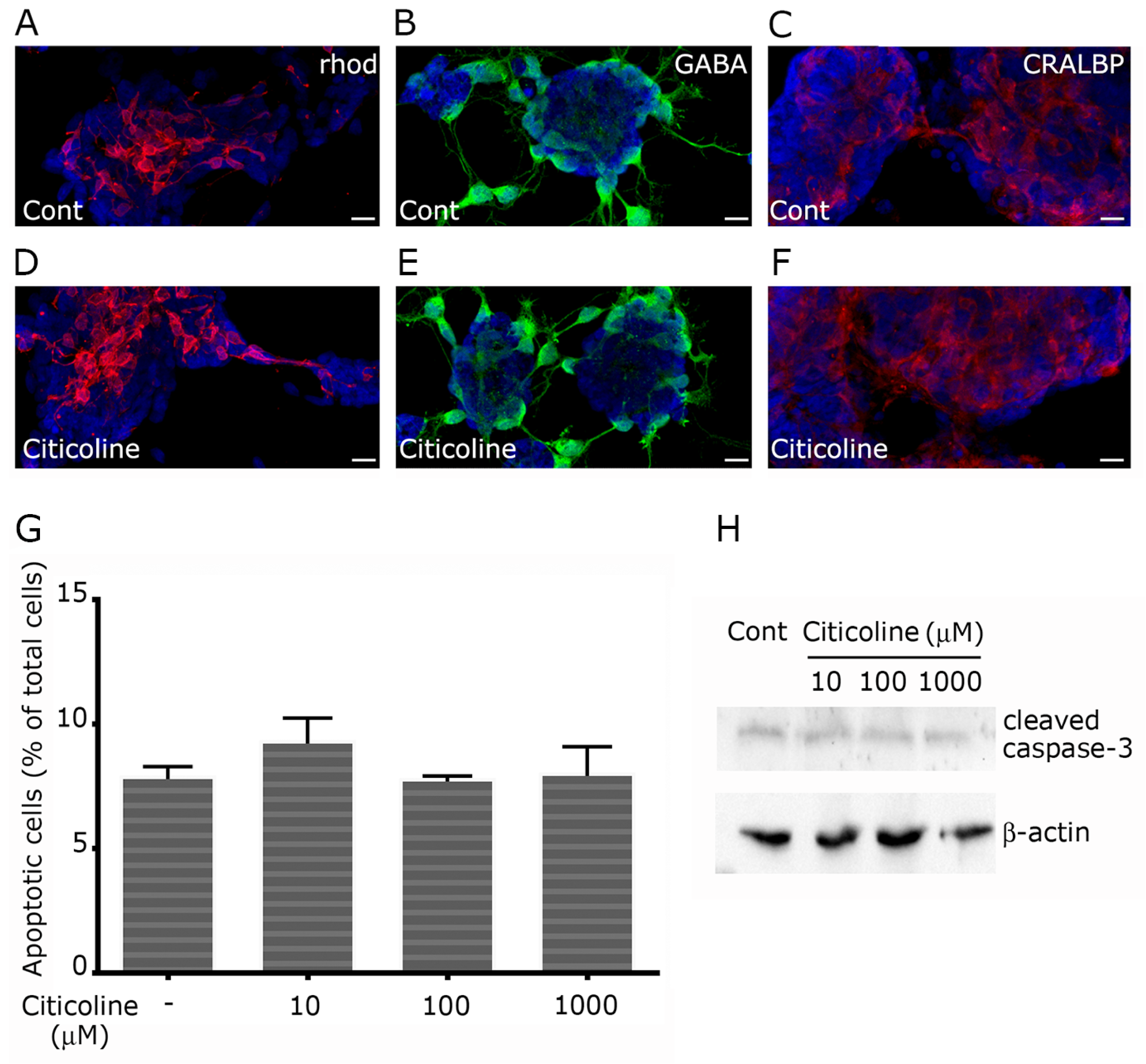
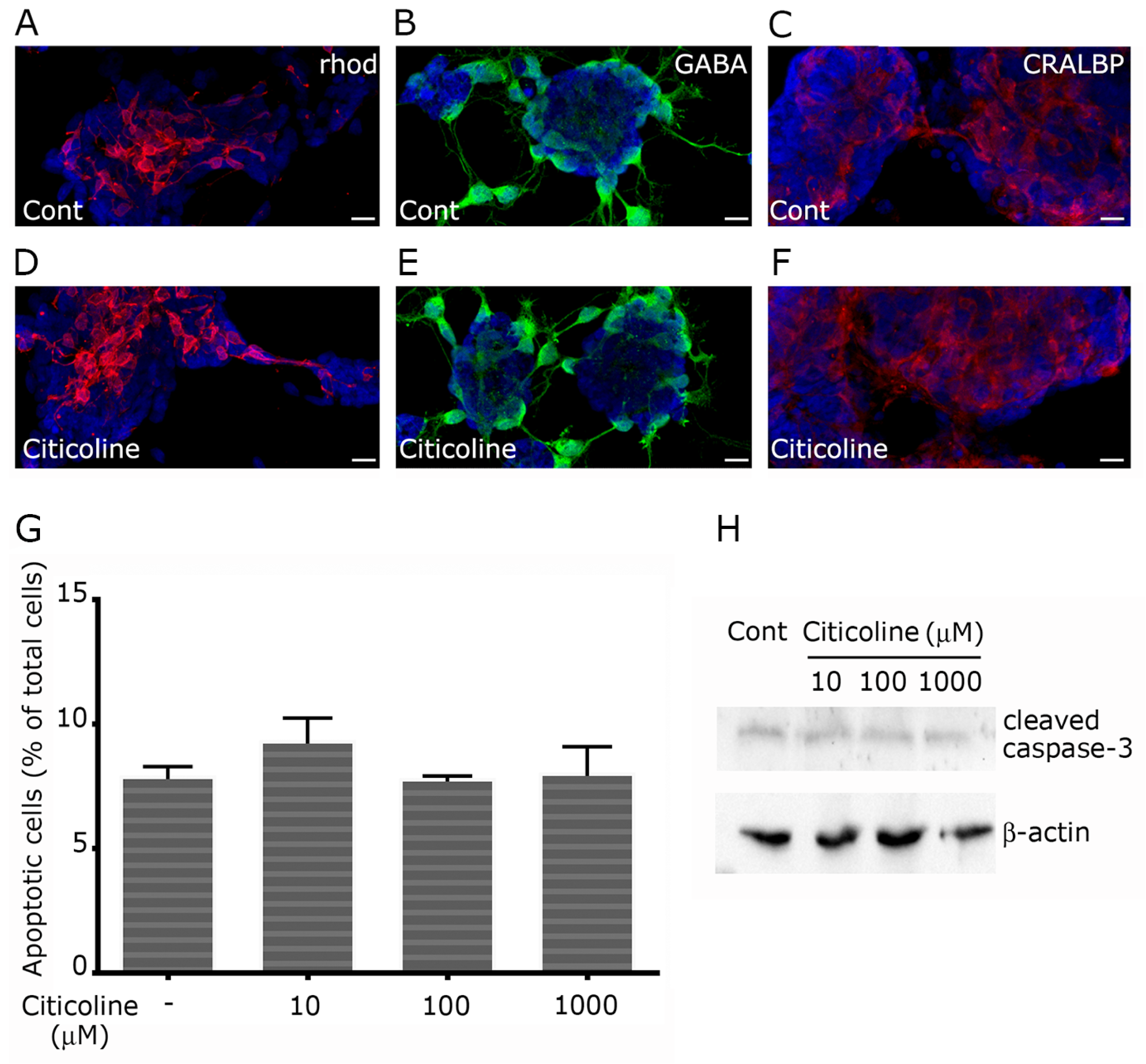
2.1. Citicoline Is Well Tolerated in Primary Retinal Cultures
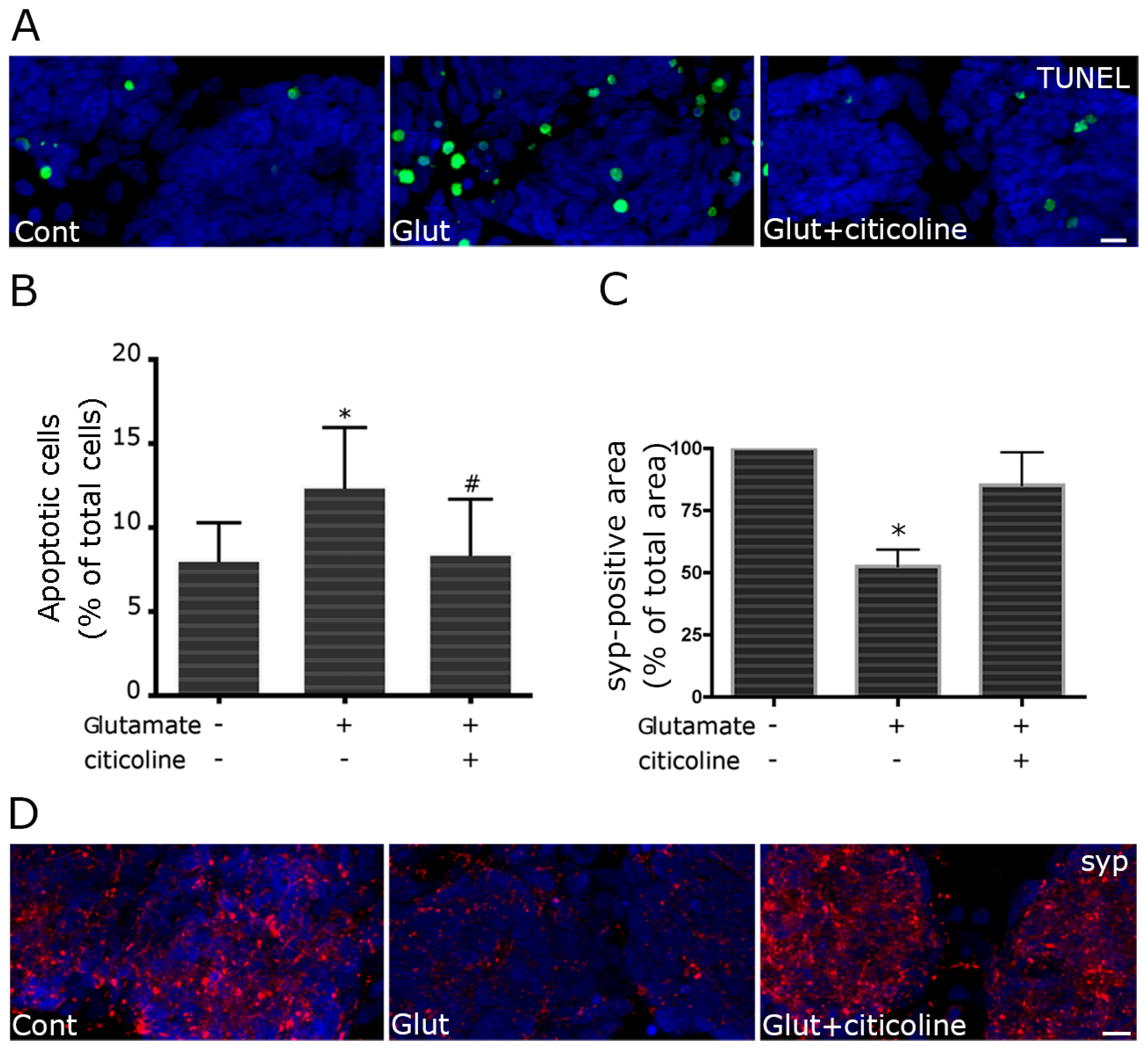
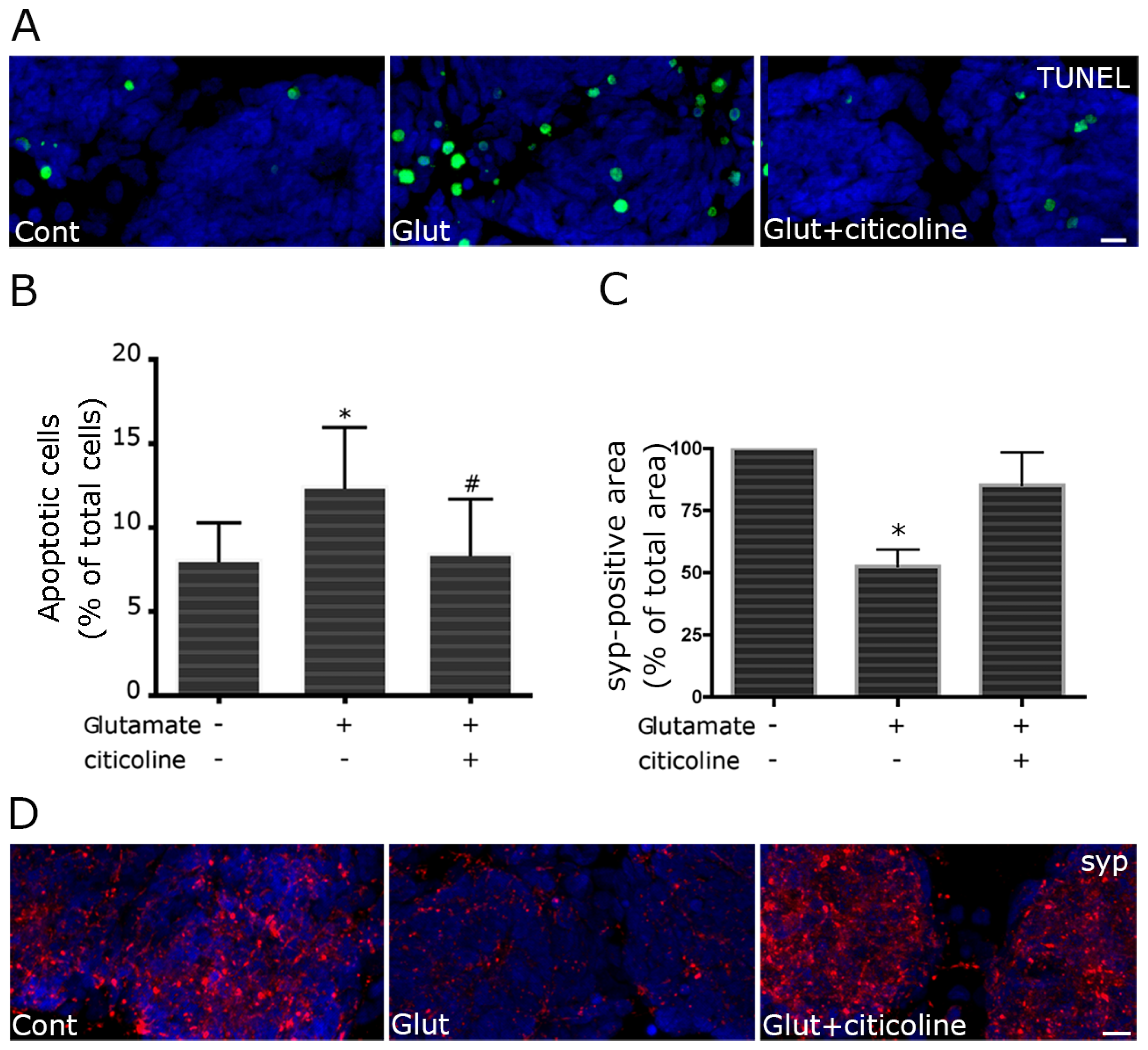
2.2. Citicoline Protects against Excitotoxic Cell Damage
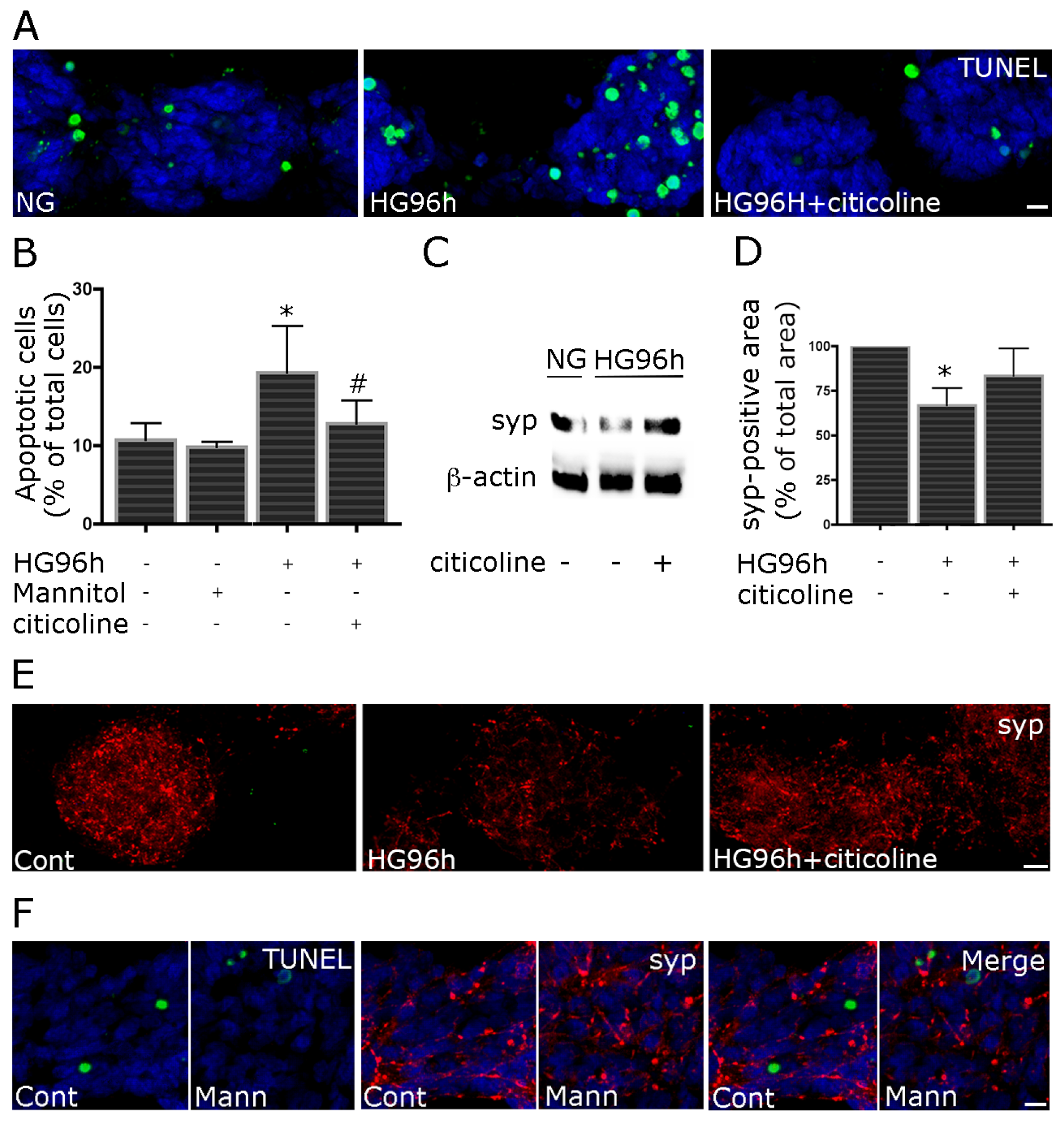
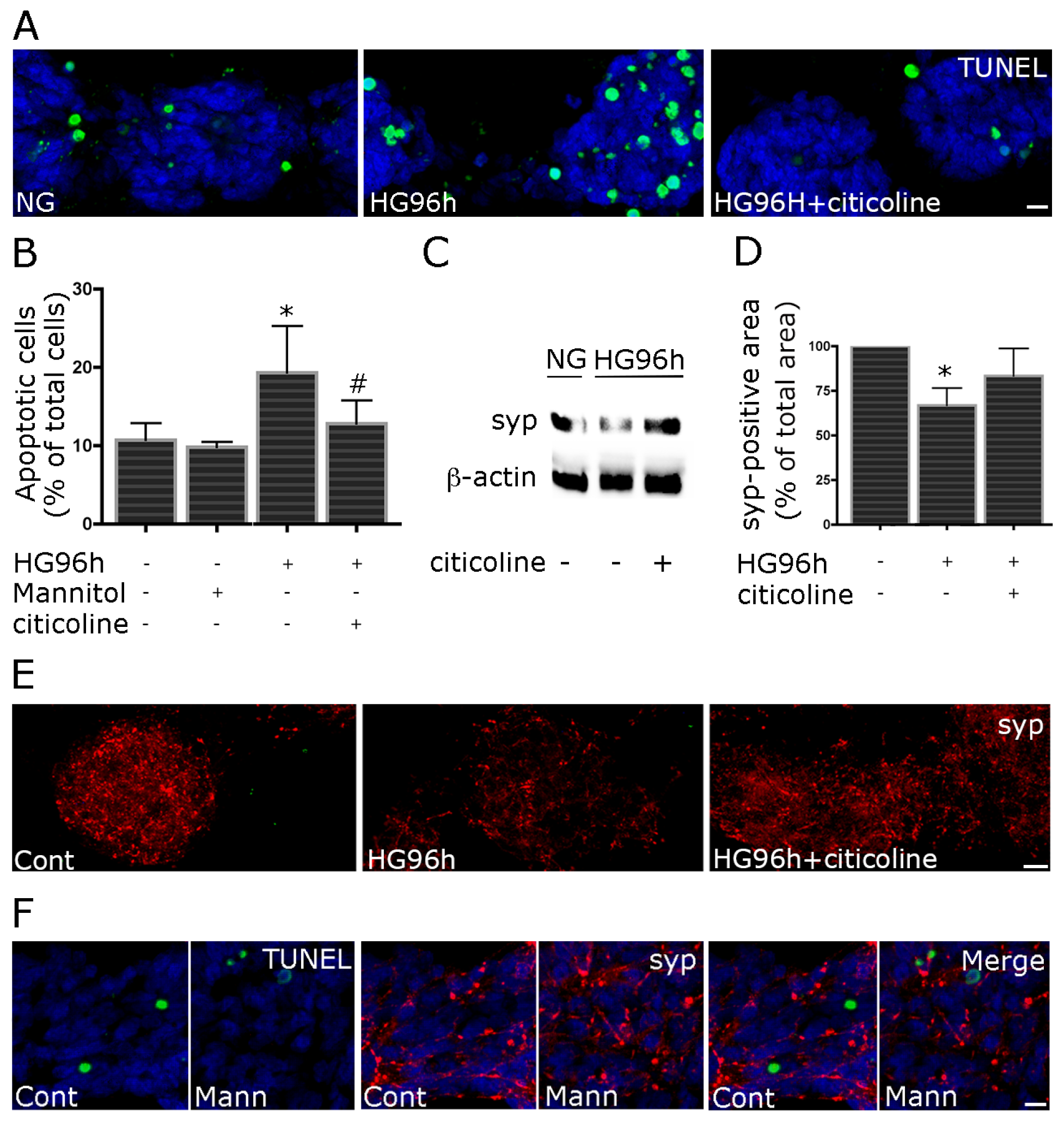
2.3. Citocoline Protects against HG-Induced Neurotoxicity
3. Experimental Section
3.1. Retinal Cultures
3.2. Cell Culture Treatments
3.3. Apoptosis Detection
3.4. Immunocytochemistry
3.5. Morphometric Analysis
3.6. Electrophoresis and Western Blot Analysis
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsA.M., L.G., C.M. and M.Vi. performed the experiments; A.M., M.Va., F.P. and F.M.A. discussed the results; A.M. and F.M.A. wrote the manuscript.
References
- Diederich, K.; Frauenknecht, K.; Minnerup, J.; Schneider, B.K.; Schmidt, A.; Altach, E.; Eggert, V.; Sommer, C.J.; Schäbitz, W.R. Citicoline enhances neuroregenerative processes after experimental stroke in rats. Stroke 2012, 43, 1931–1940. [Google Scholar]
- Putignano, S.; Gareri, P.; Castagna, A.; Cerqua, G.; Cervera, P.; Cotroneo, A.M.; Fiorillo, F.; Grella, R.; Lacava, R.; Maddonni, A.; et al. Retrospective and observational study to assess the efficacy of citicoline in elderly patients suffering from stupor related to complex geriatric syndrome. Clin. Interv. Aging 2012, 7, 113–118. [Google Scholar]
- Cotroneo, A.M.; Castagna, A.; Putignano, S.; Lacava, R.; Fantò, F.; Monteleone, F.; Rocca, F.; Malara, A.; Gareri, P. Effectiveness and safety of citicoline in mild vascular cognitive impairment: the IDEALE study. Clin. Interv. Aging 2013, 8, 131–137. [Google Scholar]
- Parisi, V.; Coppola, G.; Centofanti, M.; Oddone, F.; Angrisani, A.M.; Ziccardi, L.; Ricci, B.; Quaranta, L.; Manni, G. Evidence of the neuroprotective role of citicoline in glaucoma patients. Prog. Brain Res 2008, 173, 541–554. [Google Scholar] [Green Version]
- Parisi, V.; Manni, G.; Colacino, G.; Bucci, M.G. Cytidine-5′-diphosphocholine (citicoline) improves retinal and cortical responses in patients with glaucoma. Ophthalmology 1999, 106, 1126–1134. [Google Scholar]
- Rejdak, R.; Toczolowski, J.; Kurkowski, J.; Kaminski, M.L.; Rejdak, K.; Stelmasiak, Z.; Grieb, P. Oral citicoline treatment improves visual pathway function in glaucoma. Med. Sci. Monit 2003, 9, 24–28. [Google Scholar]
- Ottobelli, L.; Manni, G.L.; Centofanti, M.; Iester, M.; Allevena, F.; Rossetti, L. Citicoline oral solution in glaucoma: Is there a role in slowing disease progression? Ophthalmologica 2013, 229, 219–226. [Google Scholar]
- Oshitari, T.; Yoshida-Hata, N.; Yamamoto, S. Effect of neurotrophic factors on neuronal apoptosis and neurite regeneration in cultured rat retinas exposed to high glucose. Brain Res 2010, 1346, 43–51. [Google Scholar]
- Mir, C.; Clotet, J.; Aledo, R.; Durany, N.; Argemi, J.; Lozano, R.; Cervos-Navarro, J.; Casals, N. CDP-choline prevents glutamate-mediated cell death in cerebellar granule neurons. J. Mol. Neurosci 2003, 20, 53–60. [Google Scholar]
- Matyja, E.; Taraszewska, A.; Nagańska, E.; Grieb, P.; Rafałowska, J. CDP-cholineprotects motor neurons against apoptotic changes in a model of chronic glutamate excitotoxicity in vitro. Folia Neuropathol 2008, 46, 139–148. [Google Scholar]
- Choi, D.W. Glutamate neurotoxicity and diseases of the nervous system. Neuron 1988, 1, 623–634. [Google Scholar]
- Almasieh, M.; Wilson, A.M.; Morquette, B.; Cueva Vargas, J.L.; di Polo, A. The molecular basis of retinal ganglion cell death in glaucoma. Prog. Retin. Eye Res 2012, 31, 152–181. [Google Scholar]
- Han, Y.S.; Chung, I.Y.; Park, J.M.; Yu, J.M. Neuroprotective effect of citicoline on retinal cell damage induced by kainic acid in rats. Korean J. Ophthalmol 2005, 19, 219–226. [Google Scholar]
- Baltmr, A.; Duggan, J. Niz ari, S., Salt, T.E., Cordeiro, M.F. Neuroprotection in glaucoma—Is there a future role? Exp. Eye Res 2010, 91, 554–566. [Google Scholar]
- Antonetti, D.A.; Barber, A.J.; Bronson, S.K.; Freeman, W.M.; Gardner, T.W.; Jefferson, L.S.; Kester, M.; Kimball, S.R.; Krady, J.K.; LaNoue, K.F.; et al. Diabetic retinopathy: Seeing beyond glucose-induced microvascular disease. Diabetes 2006, 55, 2401–2411. [Google Scholar]
- Lieth, E.; Gardner, T.W.; Barber, A.J.; Antonetti, D.A. Penn State Retina Research Group. Retinal neurodegeneration: Early pathology in diabetes. Clin. Exp. Ophthalmol 2000, 28, 3–8. [Google Scholar]
- Parisi, V.; Uccioli, L. Visual electrophysiological responses in persons with type 1 diabetes. Diabetes Metab. Res. Rev 2001, 17, 12–18. [Google Scholar]
- Hernández, C.; Simó, R. Neuroprotection in diabetic retinopathy. Curr. Diabetes Rep 2012, 12, 329–337. [Google Scholar]
- Ola, M.S.; Nawaz, M.I.; Khan, H.A.; Alhomida, A.S. Neurodegeneration and neuroprotection in diabetic retinopathy. Int. J. Mol. Sci 2013, 14, 2559–2572. [Google Scholar]
- VanGuilder, H.D.; Brucklacher, R.M.; Patel, K.; Ellis, R.W.; Freeman, W.M.; Barber, A.J. Diabetes downregulates presynaptic proteins and reduces basal synapsin I phosphorylation in rat retina. Eur. J. Neurosci 2008, 28, 1–11. [Google Scholar]
- Santiago, A.R.; Rosa, S.C.; Santos, P.F.; Cristóvão, A.J.; Barber, A.J.; Ambrósio, A.F. Elevated glucose changes the expression of ionotropic glutamate receptor subunits and impairs calcium homeostasis in retinal neural cells. Investig. Ophthalmol. Vis. Sci 2006, 47, 4130–4137. [Google Scholar]
- Stem, M.S.; Gardner, T.W. Neurodegeneration in the pathogenesis of diabeticretinopathy: Molecular mechanisms and therapeutic implications. Curr. Med. Chem 2013, 20, 3241–3250. [Google Scholar]
- Adibhatla, R.M.; Hatcher, J.F.; Dempsey, R.J. Citicoline: Neuroprotective mechanisms in cerebral ischemia. J. Neurochem 2002, 80, 12–23. [Google Scholar]
- Xie, B.; Jiao, Q.; Cheng, Y.; Zhong, Y.; Shen, X. Effect of pigment epithelium-derived factor on glutamate uptake in retinal Muller cells under high-glucose conditions. Investig. Ophthalmol. Vis. Sci 2012, 53, 1023–1032. [Google Scholar]
- Malchiodi-Albedi, F.; Feher, J.; Caiazza, S.; Formisano, G.; Perilli, R.; Falchi, M.; Petrucci, T.C.; Scorcia, G.; Tombran-Tink, J. PEDF (pigment epithelium-derived factor) promotes increase and maturation of pigment granules in pigment epithelial cells in neonatal albino rat retinal cultures. Int. J. Dev. Neurosci 1998, 1, 423–432. [Google Scholar]
- Matteucci, A.; Cammarota, R.; Paradisi, S.; Varano, M.; Balduzzi, M.; Leo, L.; Bellenchi, G.C.; de Nuccio, C.; Carnovale-Scalzo, G.; Scorcia, G.; et al. Curcumin protects against NMDA-induced toxicity: A possible role for NR2A subunit. Investig. Ophthalmol. Vis. Sci 2011, 52, 1070–1077. [Google Scholar]
- Malchiodi-Albedi, F.; Paradisi, S.; di Nottia, M.; Simone, D.; Travaglione, S.; Falzano, L.; Guidotti, M.; Frank, C.; Cutarelli, A.; Fabbri, A.; et al. CNF1 improves astrocytic ability to support neuronal growth and differentiation in vitro. PLoS One 2012, 7, e34115. [Google Scholar]
- Gaddini, L.; Villa, M.; Matteucci, A.; Mallozzi, C.; Petrucci, T.C.; di Stasi, A.M.; Leo, L.; Malchiodi-Albedi, F.; Pricci, F. Early effects of high glucose in retinal tissue cultures Renin-Angiotensin system-dependent and -independent signaling. Neurobiol. Dis 2009, 35, 278–285. [Google Scholar]
- Hurtado, O.; Hernandez-Jimenez, M.; Zarruk, J.G.; Cuartero, M.I.; Ballesteros, I.; Camarero, G.; Moraga, A.; Pradillo, J.M.; Moro, M.A.; Lizasoain, I. Citicoline (CDP-choline) increases Sirtuin1 expression concomitant to neuroprotection in experimental stroke. J. Neurochem 2013, 26, 819–826. [Google Scholar]
- Hurtado, O.; Lizasoain, I.; Moro, M.A. Neuroprotection and recovery: Recent data at the bench on citicoline. Stroke 2011, 42, S33–S35. [Google Scholar]
- Hurtado, O.; Pradillo, J.M.; Fernandez-Lopez, D.; Morales, J.R.; Sobrino, T.; Castillo, J.; Alborch, E.; Moro, M.A.; Lizasoain, I. Delayed post-ischemic administration of CDP-choline increases EAAT2 association to lipid rafts and affords neuroprotection in experimental stroke. Neurobiol. Dis 2008, 29, 123–131. [Google Scholar]
- Kitada, M.; Kume, S.; Kanasaki, K.; Takeda-Watanabe, A.; Koya, D. Sirtuins as possible drug targets in type 2 diabetes. Curr. Drug Targets 2013, 14, 622–636. [Google Scholar]
- Saver, J.L. Citicoline: Update on a promising and widely available agent for neuroprotection and neurorepair. Rev. Neurol. Dis 2008, 5, 167–177. [Google Scholar]
- Secades, J.J.; Lorenzo, J.L. Citicoline: Pharmacological and clinical review, 2006 update. Methods Find Exp. Clin. Pharmacol 2006, 28, 1–56. [Google Scholar]
- Grieb, P. Neuroprotective properties of citicoline: Facts, doubts and unresolved issues. CNS Drugs 2014, 28, 185–193. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Matteucci, A.; Varano, M.; Gaddini, L.; Mallozzi, C.; Villa, M.; Pricci, F.; Malchiodi-Albedi, F. Neuroprotective Effects of Citicoline in in Vitro Models of Retinal Neurodegeneration. Int. J. Mol. Sci. 2014, 15, 6286-6297. https://doi.org/10.3390/ijms15046286
Matteucci A, Varano M, Gaddini L, Mallozzi C, Villa M, Pricci F, Malchiodi-Albedi F. Neuroprotective Effects of Citicoline in in Vitro Models of Retinal Neurodegeneration. International Journal of Molecular Sciences. 2014; 15(4):6286-6297. https://doi.org/10.3390/ijms15046286
Chicago/Turabian StyleMatteucci, Andrea, Monica Varano, Lucia Gaddini, Cinzia Mallozzi, Marika Villa, Flavia Pricci, and Fiorella Malchiodi-Albedi. 2014. "Neuroprotective Effects of Citicoline in in Vitro Models of Retinal Neurodegeneration" International Journal of Molecular Sciences 15, no. 4: 6286-6297. https://doi.org/10.3390/ijms15046286