Identification of the Novel Interacting Partners of the Mammalian Target of Rapamycin Complex 1 in Human CCRF-CEM and HEK293 Cells


Abstract
:1. Introduction
2. Results
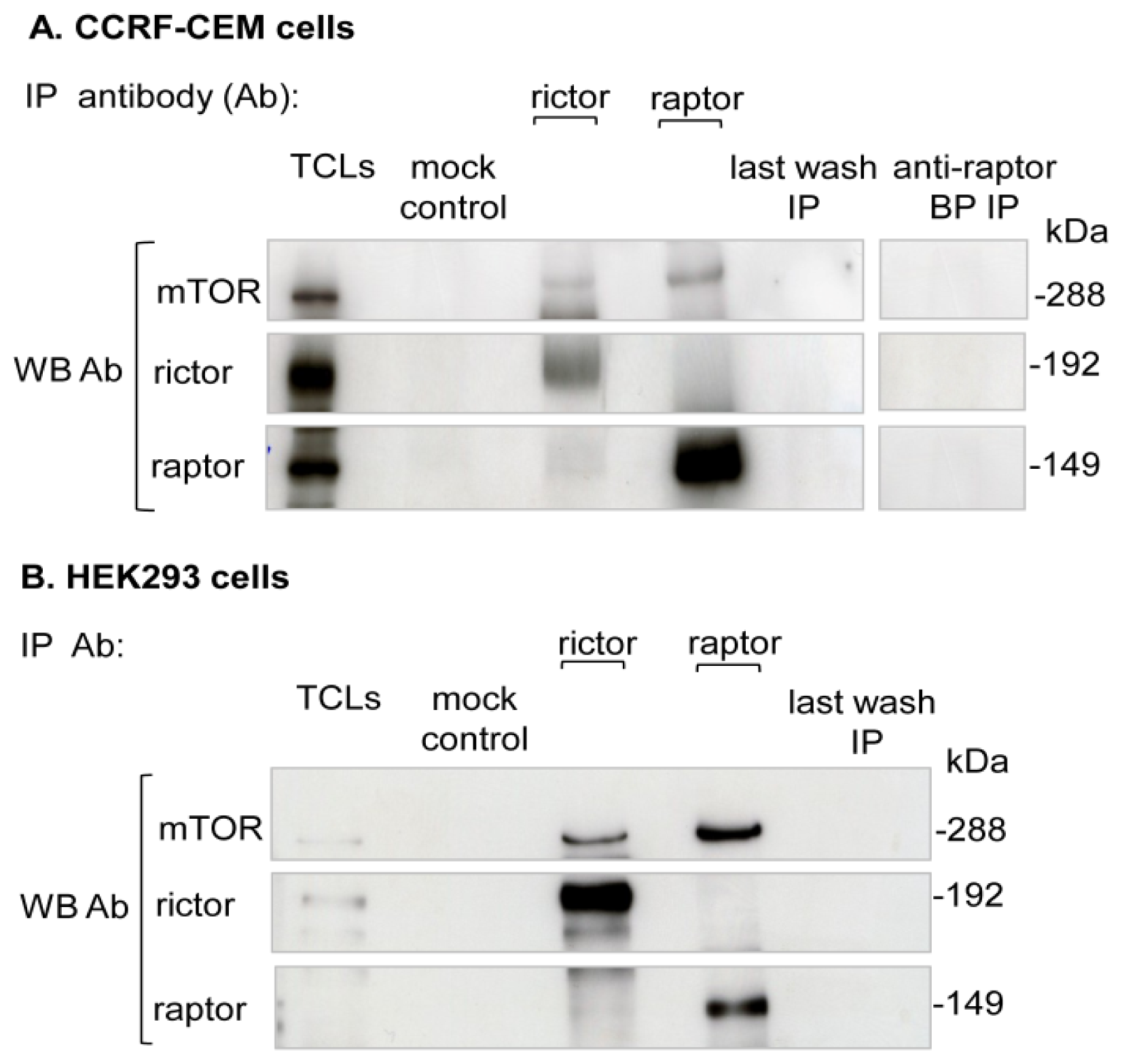
2.1. Purification of Endogenous mTORC1
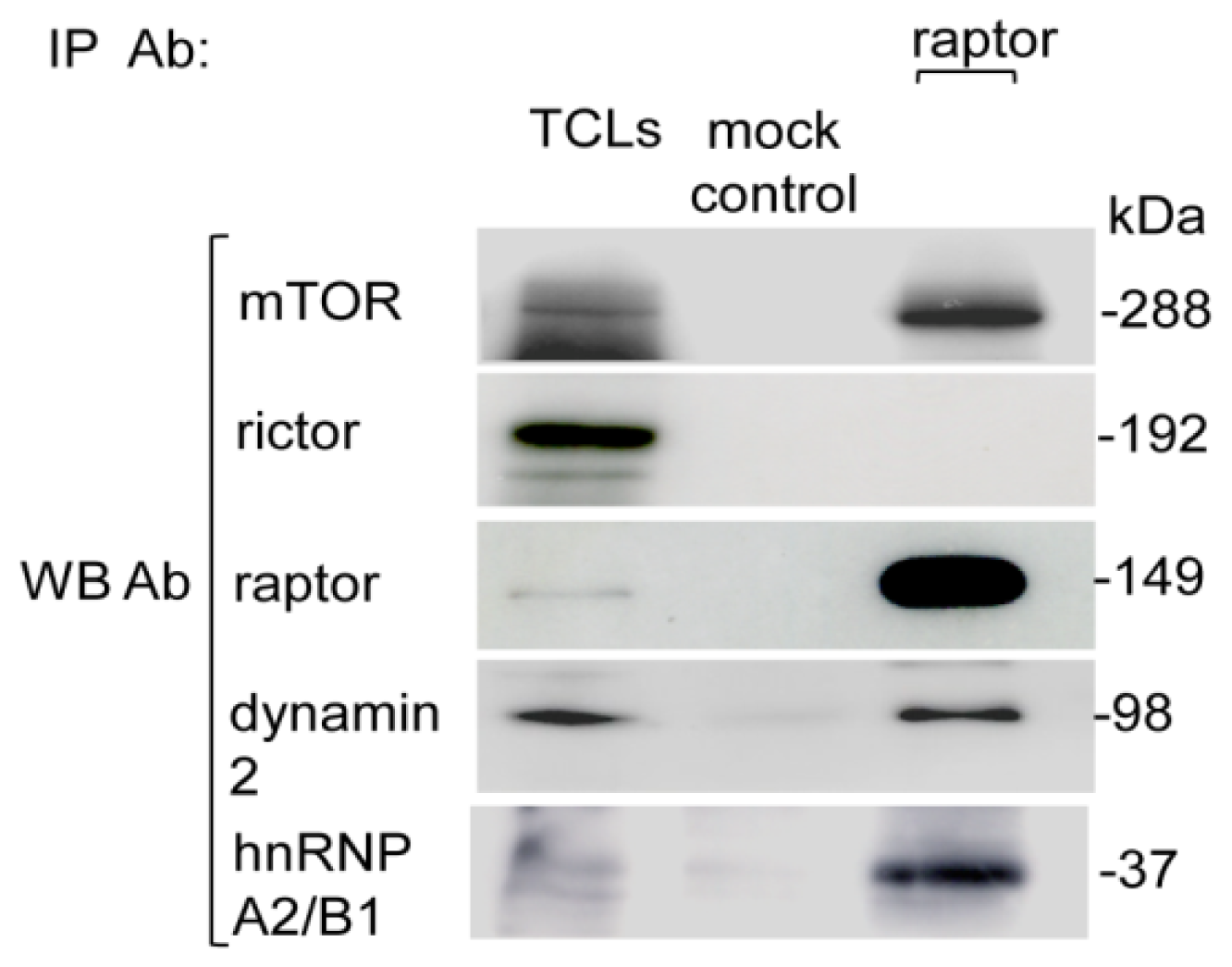
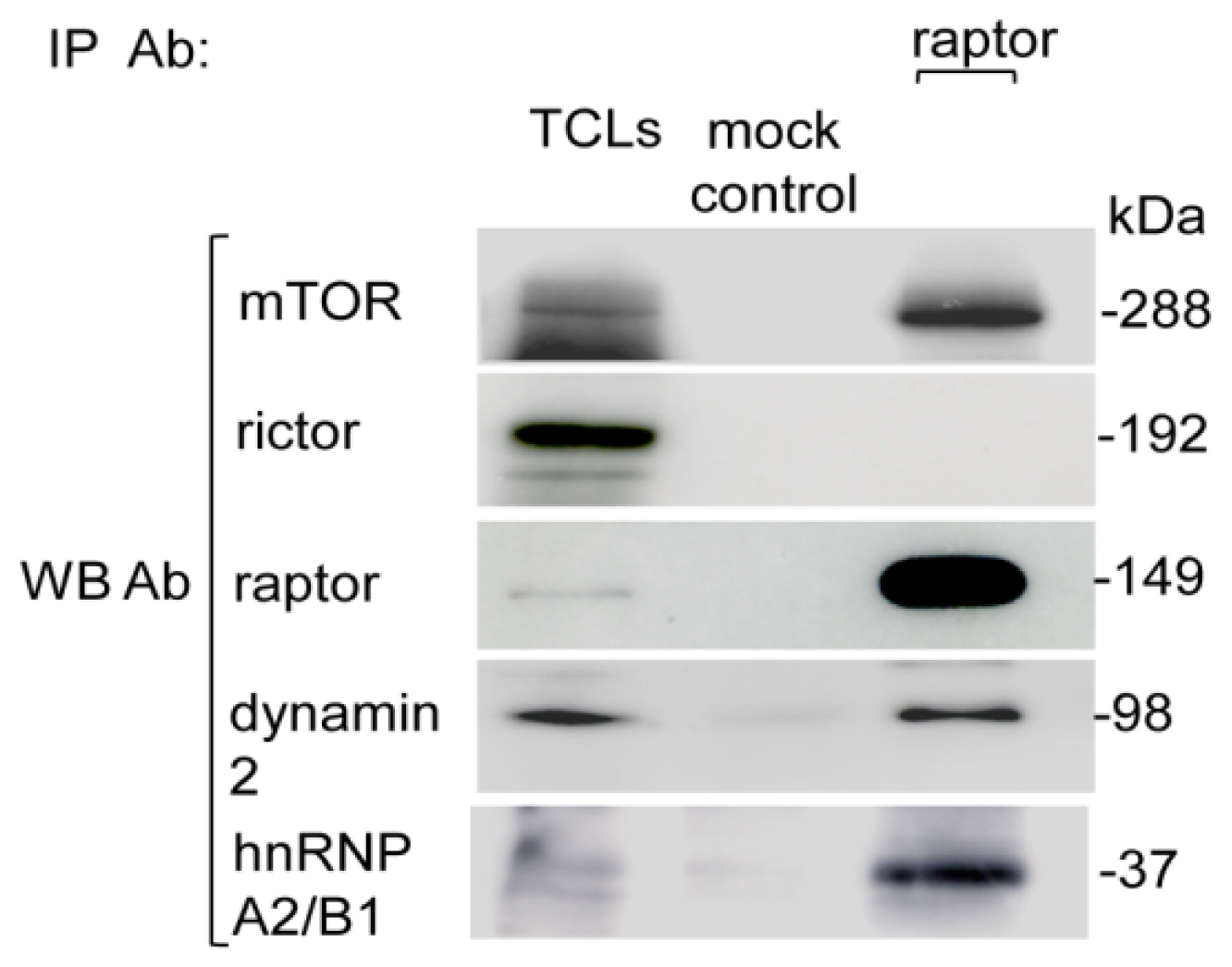
2.2. Purification of the Myc-Tag Raptor Component of mTORC1
2.3. Identification of mTORC1 Interacting Proteins Using Nano-LC ESI Q-TOF MS/MS Analysis
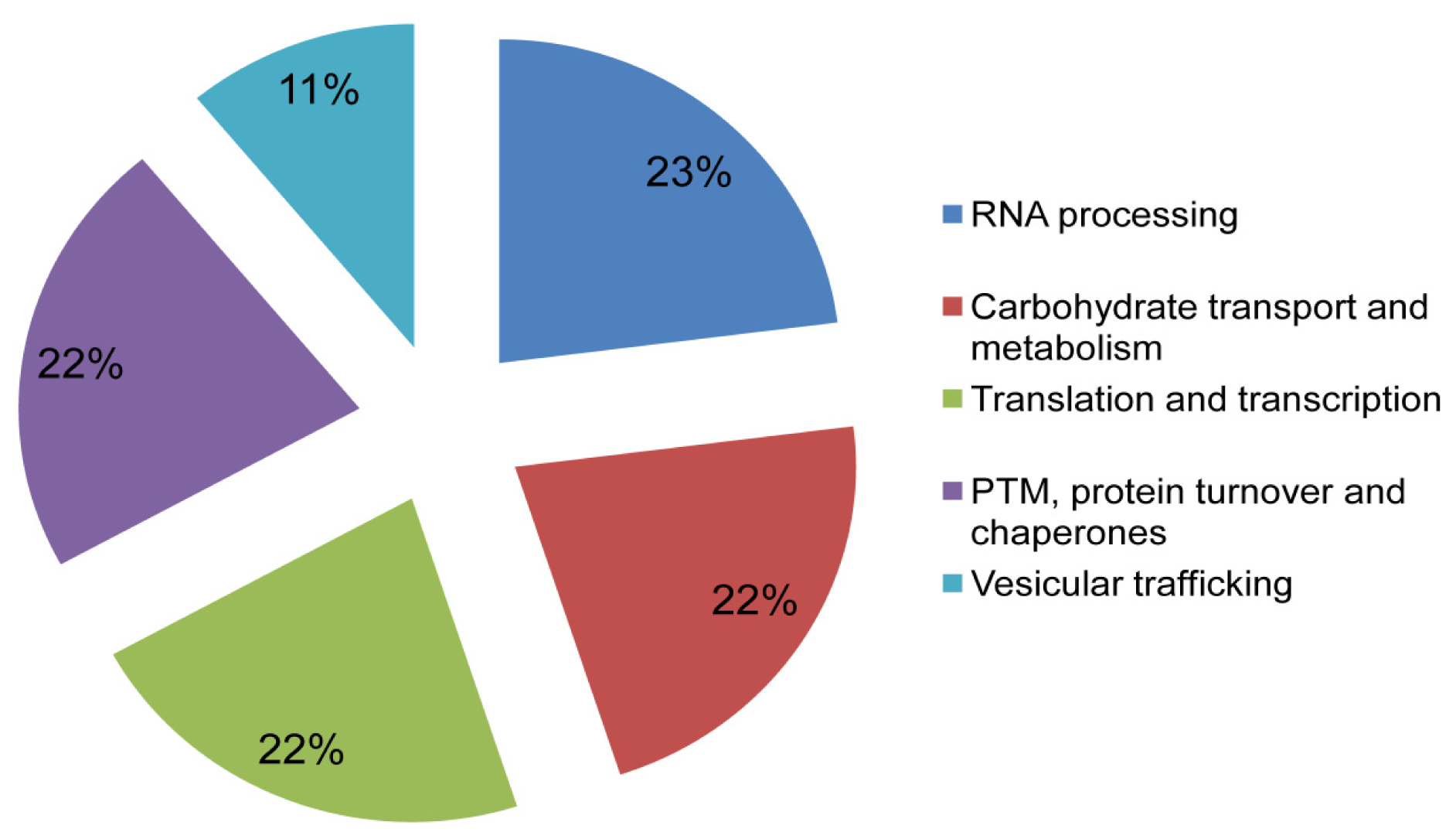
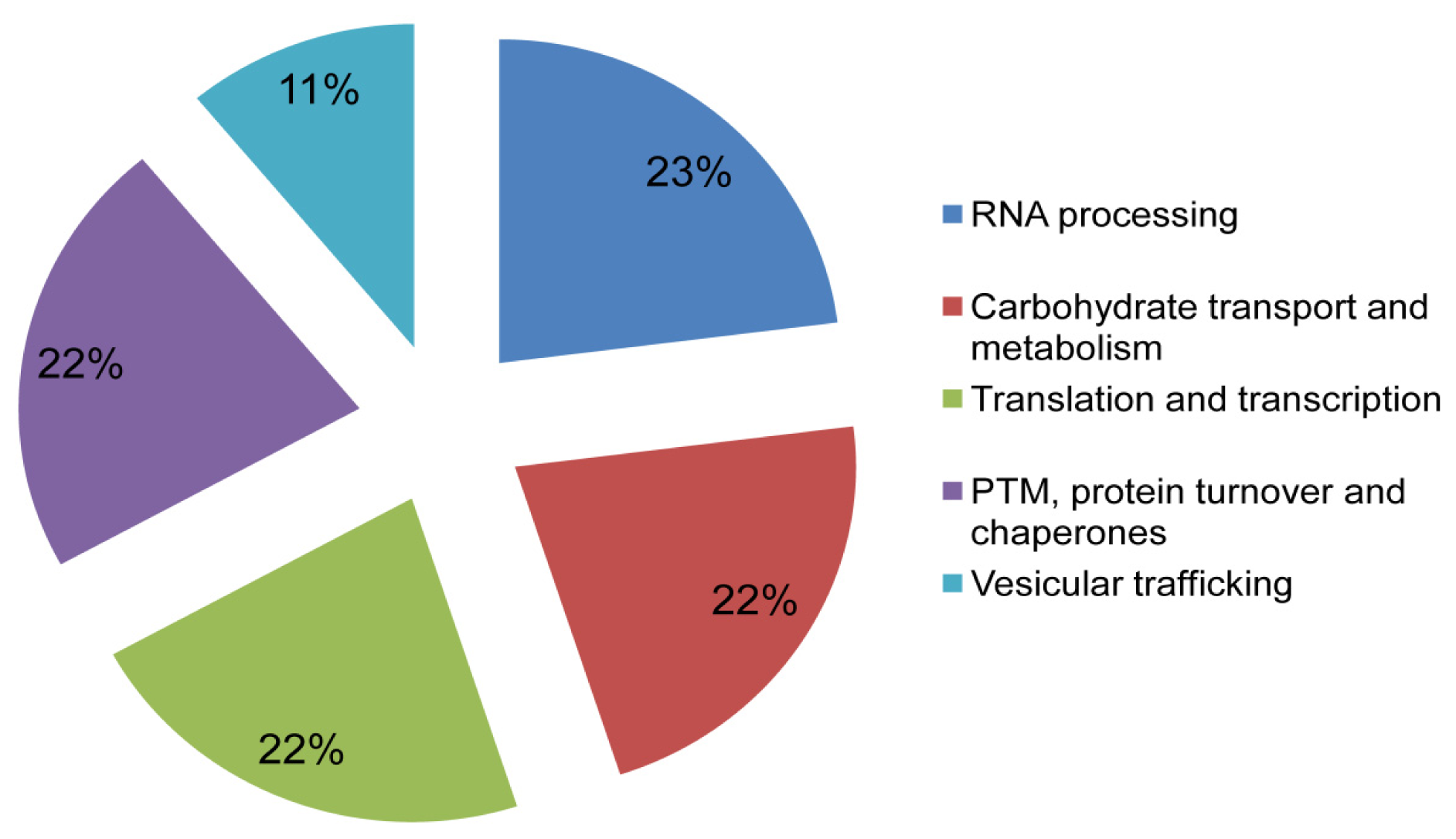
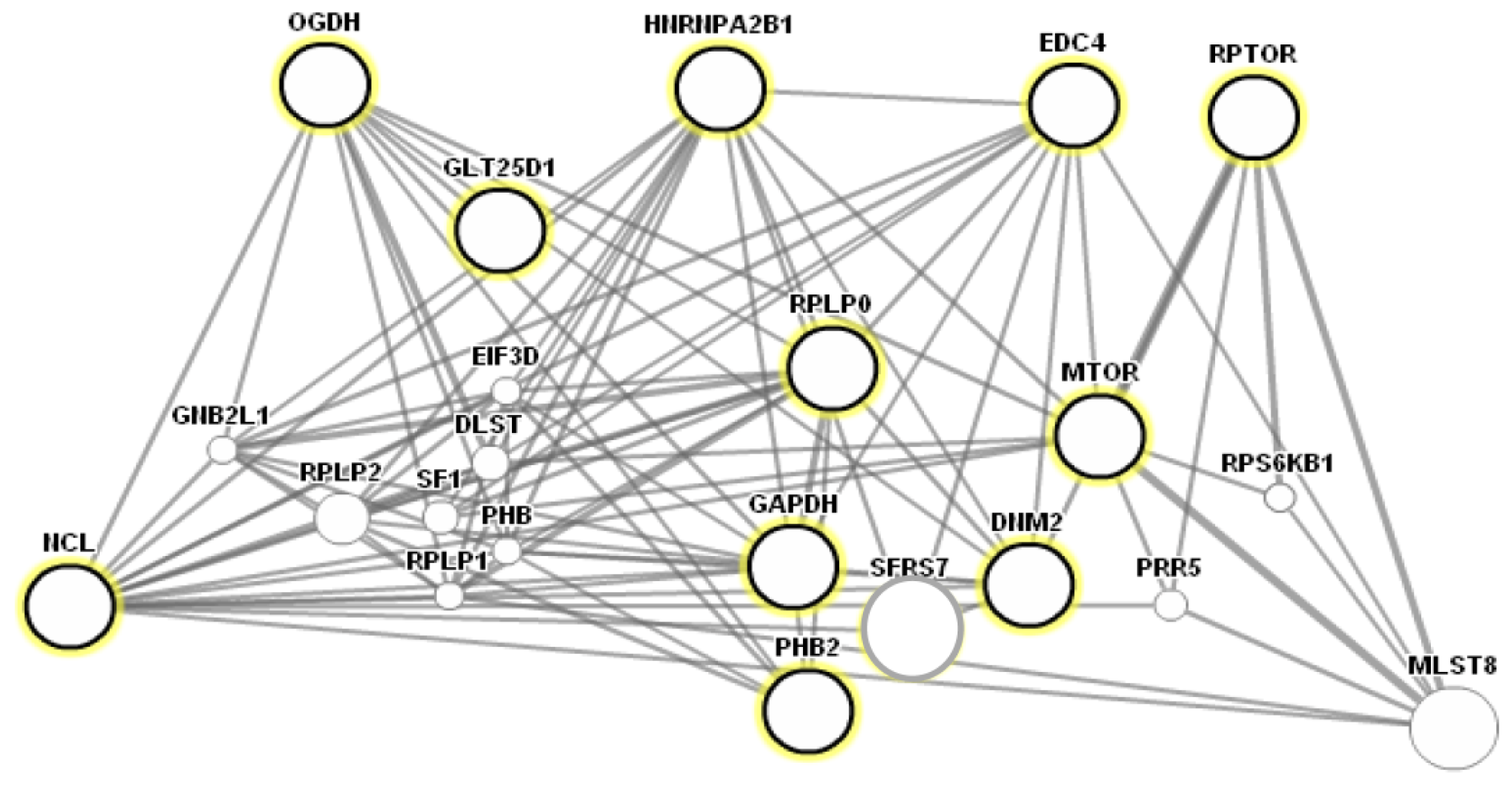
2.4. Functional Annotation and Protein-Protein Interaction Prediction
3. Discussion
4. Experimental Section
4.1. Antibodies and Reagents
4.2. Cell Culture
4.3. Cell Lysis and Endogenous mTORC1 Purification
4.4. Mammalian Cells Transfection and Myc-Tag mTORC1 Purification
4.5. SDS-PAGE and Immunoblot Analysis
4.6. Protein Visualization and In-Gel Digestion of Proteins
4.7. Peptide Sequence Analysis by Nano-LC ESI Q-TOF MS/MS and Database Search
4.8. Functional Annotation and Protein-Protein Interaction Prediction
5. Conclusions
Supplementary Information
ijms-15-04823-s001.pdfAcknowledgments
Conflicts of Interest
- Author ContributionsHazir Rahman designed and performed all the experiments, interpreted the findings and drafted the manuscript. Muhammad Qasim contributed in the interpretation of the study and in manuscript writing. Abdul R. Asif and Michael Oellerich participated in the design, supervision and interpretation of the study. All the authors read and approved the final manuscript.
References
- Laplante, M.; Sabatini, D.M. mTOR Signaling in Growth Control and Disease. Cell 2012, 149, 274–293. [Google Scholar]
- Anya, A.; Marina, H. mTOR signaling for biological control and cancer. J. Cell. Physiol. 2013, 228, 1658–1664. [Google Scholar]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar]
- Chen, J.; Zheng, X.F.; Brown, E.J.; Schreiber, S.L. Identification of an 11-kDa FKBP12-rapamycin-binding domain within the 289-kDa FKBP12-rapamycin-associated protein and characterization of a critical serine residue. Proc. Natl. Acad. Sci. USA 1995, 92, 4947–4951. [Google Scholar]
- Sabatini, D.M.; Erdjument-Bromage, H.; Lui, M.; Tempst, P.; Snyder, S.H. RAFT1: A mammalian protein that binds to FKBP12 in a rapamycin-dependent fashion and is homologous to yeast TORs. Cell 1994, 78, 35–43. [Google Scholar]
- Heitman, J.; Koller, A.; Kunz, J.; Henriquez, R.; Schmidt, A.; Movva, N.R.; Hall, M.N. The immunosuppressant FK506 inhibits amino acid import in Saccharomyces cerevisiae. Mol. Cell. Biol. 1993, 13, 5010–5019. [Google Scholar]
- Kim, D.H.; Sarbassov, D.D.; Ali, S.M.; King, J.E.; Latek, R.R.; Erdjument-bromage, H.; Tempst, P.; Sabatini, D.M. mTOR interacts with raptor to form a nutrient-sensitive complex that signals to the cell growth machinery. Cell 2002, 110, 163–175. [Google Scholar]
- Sarbassov, D.D.; Ali, S.M.; Kim, D.H.; Guertin, D.A.; Latek, R.R.; Erdjument-Bromage, H.; Tempst, F.; Sabatini, D.M. Rictor a novel binding partner of mTOR defines a rapamycin-insensitive and raptor-independent pathway that regulates the cytoskeleton. Curr. Biol. 2004, 14, 1296–1302. [Google Scholar]
- Peterson, T.R.; Laplante, M.; Thoreen, C.C.; Sancak, Y.; Kang, S.A.; Kuehl, W.M.; Nathanael, S.J.; Sabatini, D.M. DEPTOR is an mTOR inhibitor frequently overexpressed in multiple myeloma cells and required for their survival. Cell 2009, 137, 873–886. [Google Scholar]
- Kim, D.H.; Sarbassov, D.D.; Ali, S.M.; Latek, R.R.; Guntur, K.V.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D.M. GbetaL a positive regulator of the rapamycin-sensitive pathway required for the nutrient-sensitive interaction between raptor and mTOR. Mol. Cell 2003, 11, 895–904. [Google Scholar]
- Sancak, Y.; Thoreen, C.C.; Peterson, T.R.; Lindquist, R.A.; Kang, S.A.; Spooner, E.; Carr, S.A.; Sabatini, D.M. PRAS40 is an insulin-regulated inhibitor of the mTORC1 protein kinase. Mol. Cell 2007, 25, 903–915. [Google Scholar]
- Jenna, L.J.; Ryanl, C.R.; Kum, L.G. Amino acid signalling upstream of mTOR. Nat. Rev. Mol. Cell Biol. 2013, 14, 133–139. [Google Scholar]
- Angus, W.; Heth, R.T.; Giorgio, R. Immunoregulatory Functions of mTOR Inhibition. Nat. Rev. Immunol. 2009, 9, 324–337. [Google Scholar]
- Hui, Y.; Xianghui, W.; Yan, Z.; Huanrong, L.; Jiongbo, L.; Kun, S.; Yiwei, C.; Guangwei, L. Modulation of TSC-mTOR signaling on immune cells in immunity and autoimmunity. J. Cell. Physiol. 2014, 229, 17–26. [Google Scholar]
- Hara, K.; Maruki, Y.; Long, X.; Yoshino, K.; Oshiro, N.; Hidayat, S. Raptor a binding partner of target of rapamycin (TOR) mediates TOR action. Cell 2002, 110, 177–189. [Google Scholar]
- Wang, X.; Proud, C.G. mTORC1 signaling: What we still don’t know. J. Mol. Cell Biol. 2011, 3, 206–220. [Google Scholar]
- Mitsugu, S.; Michael, N.H. Making new contacts: The mTOR network in metabolism and signalling crosstalk. Nat. Rev. Mol. Cell Biol. 2014, 15, 155–162. [Google Scholar]
- Zhou, J.; Allred, D.C.; Avis, I.; Martinez, A.; Vos, M.D.; Smith, L.; Treston, A.M.; Mulshine, J.L. Differential expression of the early lung cancer detection marker heterogeneous nuclear ribonucleoprotein-A2/B1 (hnRNP-A2/B1) in normal breast and neoplastic breast cancer. Breast Cancer Res. Treat. 2001, 66, 217–224. [Google Scholar]
- Drenan, R.M.; Liu, X.; Bertram, P.G.; Zheng, X.F. FKBP12-rapamycin-associated protein or mammalian target of rapamycin (FRAP/mTOR) localization in the endoplasmic reticulum and the Golgi apparatus. J. Biol. Chem. 2004, 279, 772–778. [Google Scholar]
- Li, L.; Kim, E.; Yuan, H.; Inoki, K.; Gorassha, P.; Schiesher, R.L. Regulation of mTORC1 by the Rab and Arf GTPases. J. Biol. Chem. 2010, 285, 19705–19709. [Google Scholar]
- Durieux, A.C.; Prudhon, B.; Guicheney, P.; Bitoun, M. Dynamin 2 and human diseases. J. Mol. Med. 2010, 88, 339–350. [Google Scholar]
- Heymann, J.A.; Hinshaw, J.E. Dynamins at a glance. J. Cell Sci. 2009, 122, 3427–3431. [Google Scholar]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2004, 32, 115–119. [Google Scholar]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, 214–220. [Google Scholar]
- Bonetta, L. Protein-protein interactions: Interactome under construction. Nature 2010, 468, 851–854. [Google Scholar]
- Yang, Q.; Guan, K.L. Expanding mTOR signaling. Cell Res. 2007, 17, 666–681. [Google Scholar]
- Sancak, Y.; Bar-Peled, L.; Zoncu, R.; Markhard, A.L.; Nada, S.; Sabatini, D.M. Ragulator-Rag complex targets mTORC1 to the lysosomal surface and is necessary for its activation by amino acids. Cell 2010, 141, 290–303. [Google Scholar]
- Lamond, A.I.; Mann, M. Cell biology and the genome projects a concerted strategy for characterizing multiprotein complexes by using mass spectrometry. Trends Cell Biol. 1997, 7, 139–142. [Google Scholar]
- Goh, E.T.; Pardo, O.E.; Michael, N.; Niewiarowski, A.; Totty, N.; Volkova, D.; Tsaneva, I.R.; Seckl, M.J.; Gout, I. Involvement of heterogeneous ribonucleoprotein F in the regulation of cell proliferation via the mammalian target of rapamycin/S6 kinase 2 pathway. J. Biol. Chem. 2010, 285, 17065–17076. [Google Scholar]
- Fenger-Gron, M.; Fillman, C.; Norrild, B.; Lykke-Andersen, J. Multiple processing body factors and the ARE binding protein TTP activate mRNA decapping. Mol. Cell 2005, 20, 905–915. [Google Scholar]
- Rodriguez-Gabriel, M.A.; Remacha, M.; Ballesta, J.P. Phosphorylation of ribosomal protein P0 is not essential for ribosome function but can affect translation. Biochemistry 1998, 37, 16620–16626. [Google Scholar]
- Sengupta, S.; Peterson, T.R.; Sabatini, D.M. Regulation of the mTOR complex 1 pathway by nutrients growth factors and stress. Mol. Cell 2010, 40, 310–322. [Google Scholar]
- Srivastava, M.; Pollard, H.B. Molecular dissection of nucleolin’s role in growth and cell proliferation: New insights. FASEB J. 1999, 13, 1911–1922. [Google Scholar]
- Csermely, P.; Schnaider, T.; Cheatham, B.; Olson, M.O.; Kahn, C.R. Insulin induces the phosphorylation of nucleolin A possible mechanism of insulin-induced RNA efflux from nuclei. J. Biol. Chem. 1993, 268, 9747–9752. [Google Scholar]
- Zoncu, R.; Efeyan, A.; Sabatini, D.M. mTOR: From growth signal integration to cancer diabetes and ageing. Nat. Rev. Mol. Cell Biol 2011, 12, 21–35. [Google Scholar]
- Blum, H.; Beier, H.; Gross, H.J. Improved silver staining of plant-proteins, RNA and DNA in polyacrylamide gels. Electrophoresis 1987, 8, 93–99. [Google Scholar]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anaytical Chem. 1996, 68, 850–858. [Google Scholar]
- Rahman, H.; Qasim, M.; Asif, A.R.; Oellerich, M. Fetal calf serum heat inactivation and lipopolysaccharides contamination influence human T lymphoblasts proteome and phosphoproteome. Proteome Sci. 2011, 9, 71. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession No. | Protein Name | Mass (kDa) | Protein Function | c Mascot Score | d pI | ePeptide Matches (sequences) |
|---|---|---|---|---|---|---|
| Q8N122 | a Raptor | 149 | Translation and transcription | 450 | 6.43 | 14 (11) |
| P42345 | b mTOR | 288 | Translation and transcription | 254 | 6.73 | 18 (11) |
| P22626 | hnRNPA2/B1 | 37 | mRNA processing | 106 | 8.97 | 3 (3) |
| Q6P2E9 | Edc4 | 151.6 | mRNA processing | 92 | 5.55 | 3 (3) |
| P05388 | RPLP0 | 34.3 | Translation and transcription | 105 | 5.41 | 3 (3) |
| P19338 | Nucleolin | 76.5 | Translation and transcription | 133 | 4.6 | 8 (5) |
| P50570 | DNM2 | 98 | Vesicular trafficking | 63 | 7.04 | 5 (3) |
| P04406 | GAPDH | 36 | Carbohydrate transport and metabolism | 114 | 8.57 | 7 (4) |
| Q8NBJ5 | GLT25D1 | 71.5 | Post-translational modification, protein turnover and chaperone functions | 148 | 6.85 | 6 (6) |
| Q99623 | PHB2 | 33.2 | Post-translational modification, protein turnover and chaperone functions | 147 | 9.83 | 3 (3) |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rahman, H.; Qasim, M.; Oellerich, M.; Asif, A.R. Identification of the Novel Interacting Partners of the Mammalian Target of Rapamycin Complex 1 in Human CCRF-CEM and HEK293 Cells. Int. J. Mol. Sci. 2014, 15, 4823-4836. https://doi.org/10.3390/ijms15034823
Rahman H, Qasim M, Oellerich M, Asif AR. Identification of the Novel Interacting Partners of the Mammalian Target of Rapamycin Complex 1 in Human CCRF-CEM and HEK293 Cells. International Journal of Molecular Sciences. 2014; 15(3):4823-4836. https://doi.org/10.3390/ijms15034823
Chicago/Turabian StyleRahman, Hazir, Muhammad Qasim, Michael Oellerich, and Abdul R. Asif. 2014. "Identification of the Novel Interacting Partners of the Mammalian Target of Rapamycin Complex 1 in Human CCRF-CEM and HEK293 Cells" International Journal of Molecular Sciences 15, no. 3: 4823-4836. https://doi.org/10.3390/ijms15034823