The Role of Chemokines in Hepatitis C Virus-Mediated Liver Disease

Abstract
:1. Introduction
1.1. Hepatitis C Virus
1.2. Chemokines
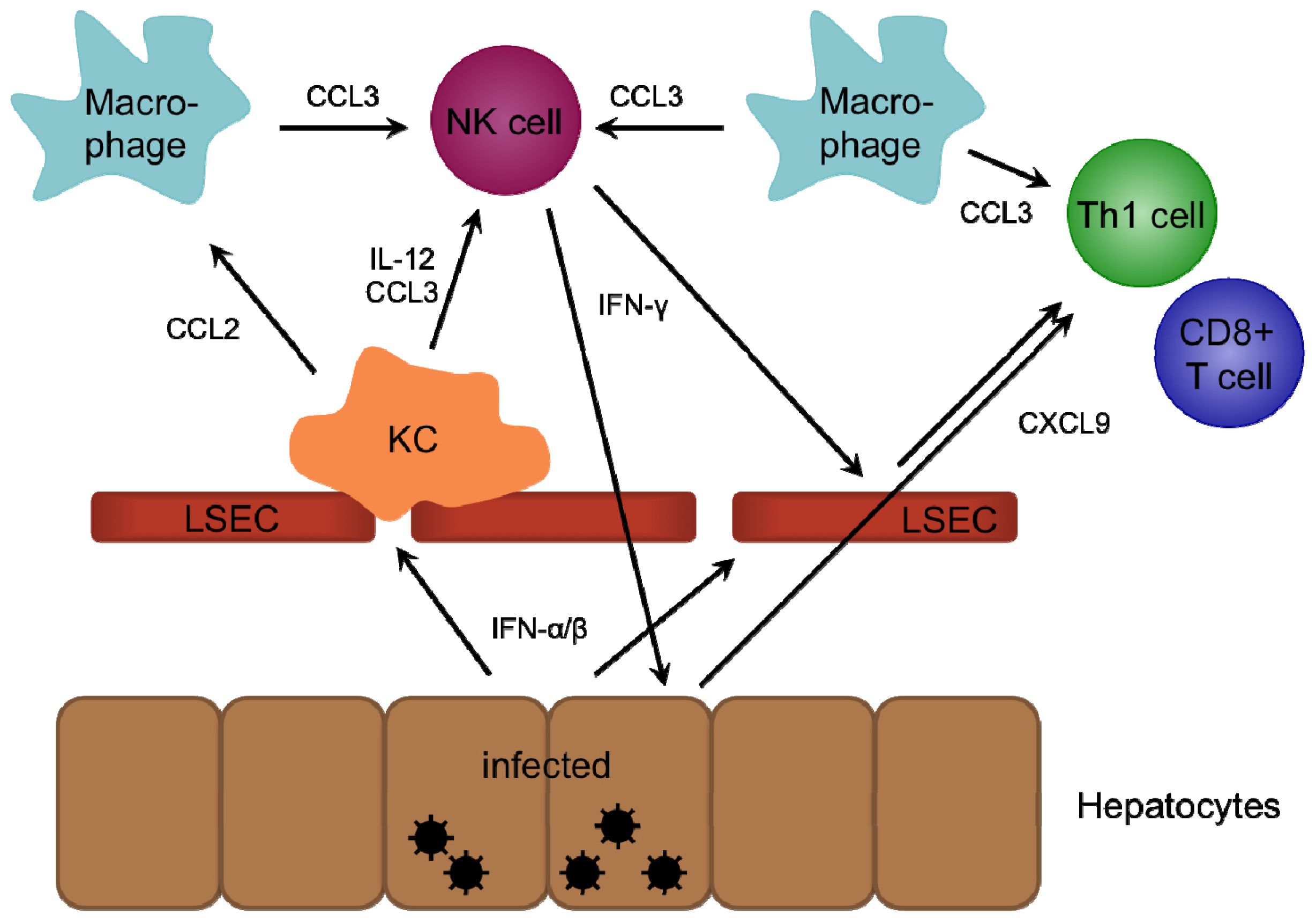
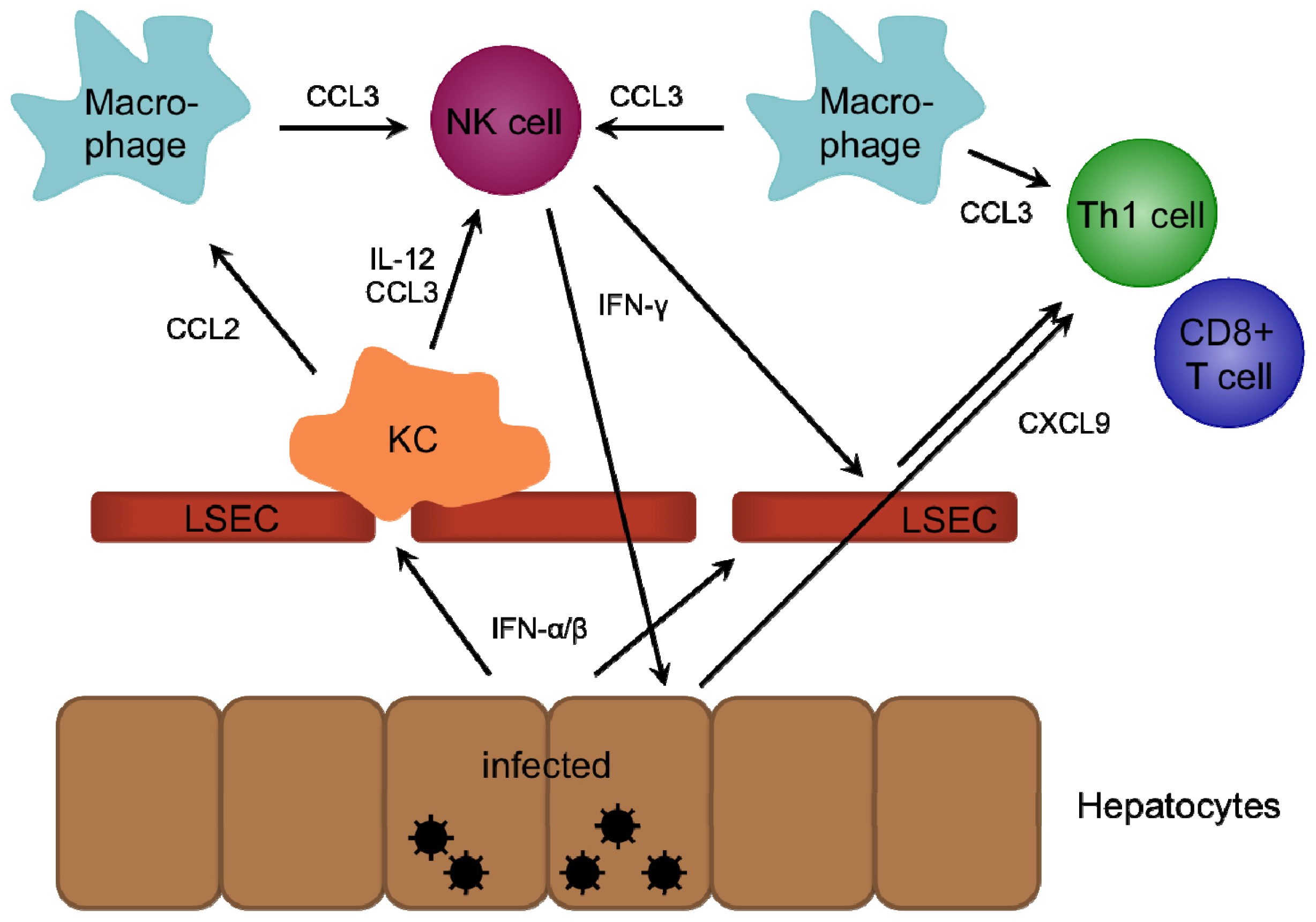
2. Role of Chemokines in Intrahepatic Immunity
2.1. CXCR3-Binding Chemokines
2.2. CXCR6-Binding Chemokines
2.3. CCR1/5-Binding Chemokines
2.4. CCR4-Binding Chemokines
2.5. CCR2-Binding Chemokines
2.6. CXCR1/2-Binding Chemokines
2.7. CXCR4/7-Binding Chemokines
3. HCV-Mediated Modulation of the Chemokine Response
3.1. CXCR3-Dependent Lymphocyte Recruitment in HCV Infection
3.2. Involvement of CXCR6 Positive Immune Cells in HCV Infection
3.3. Recruitment of CCR1/5 Positive Lymphocytes in HCV Infection
3.4. Hepatic Homing of CCR4 Positive Lymphocytes in HCV Infection
3.5. CCR2-Dependent Recruitment of Monocytes and Macrophages in HCV Infection
3.6. CXCR1/2-Expressing Immune Cells during HCV Infection
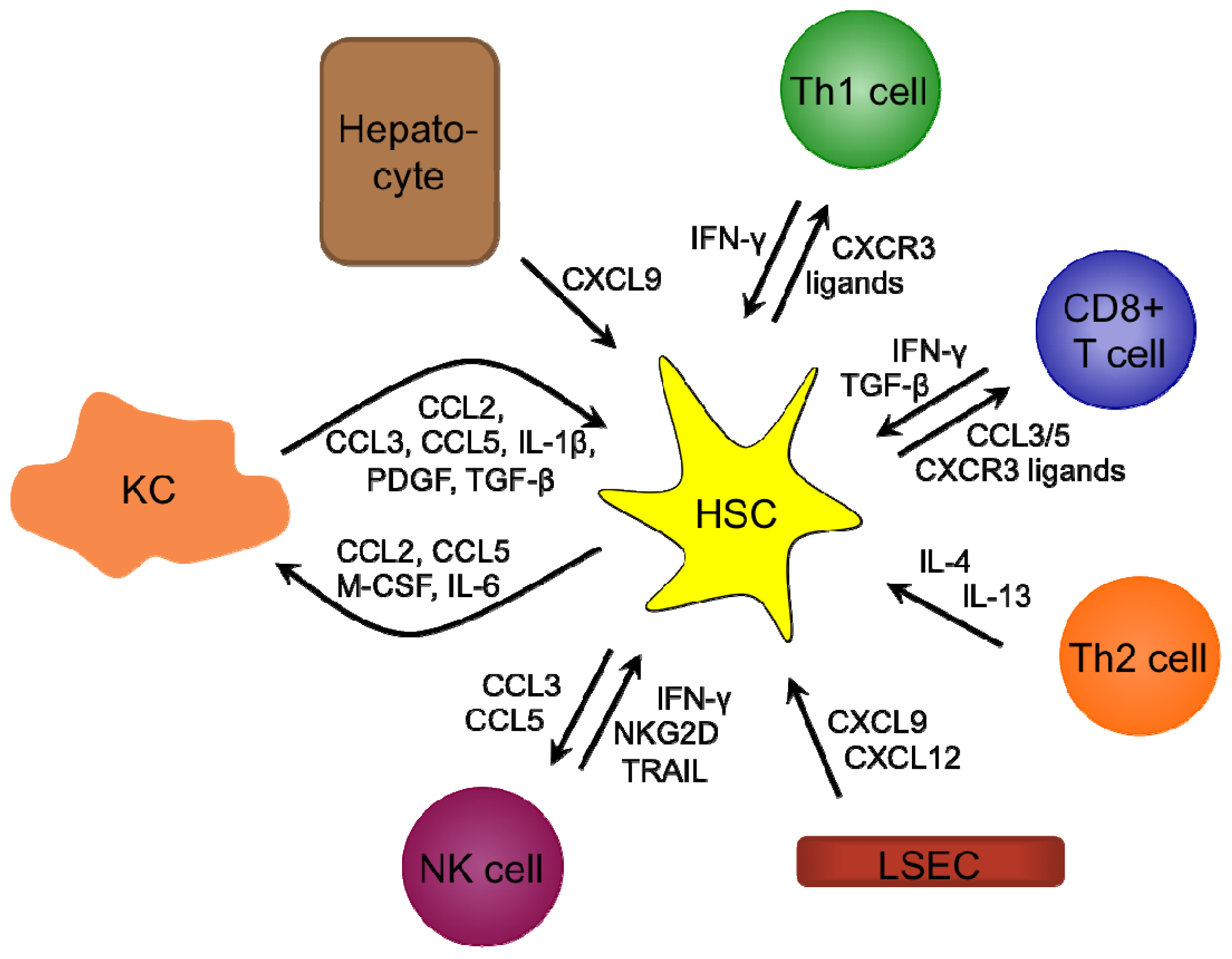
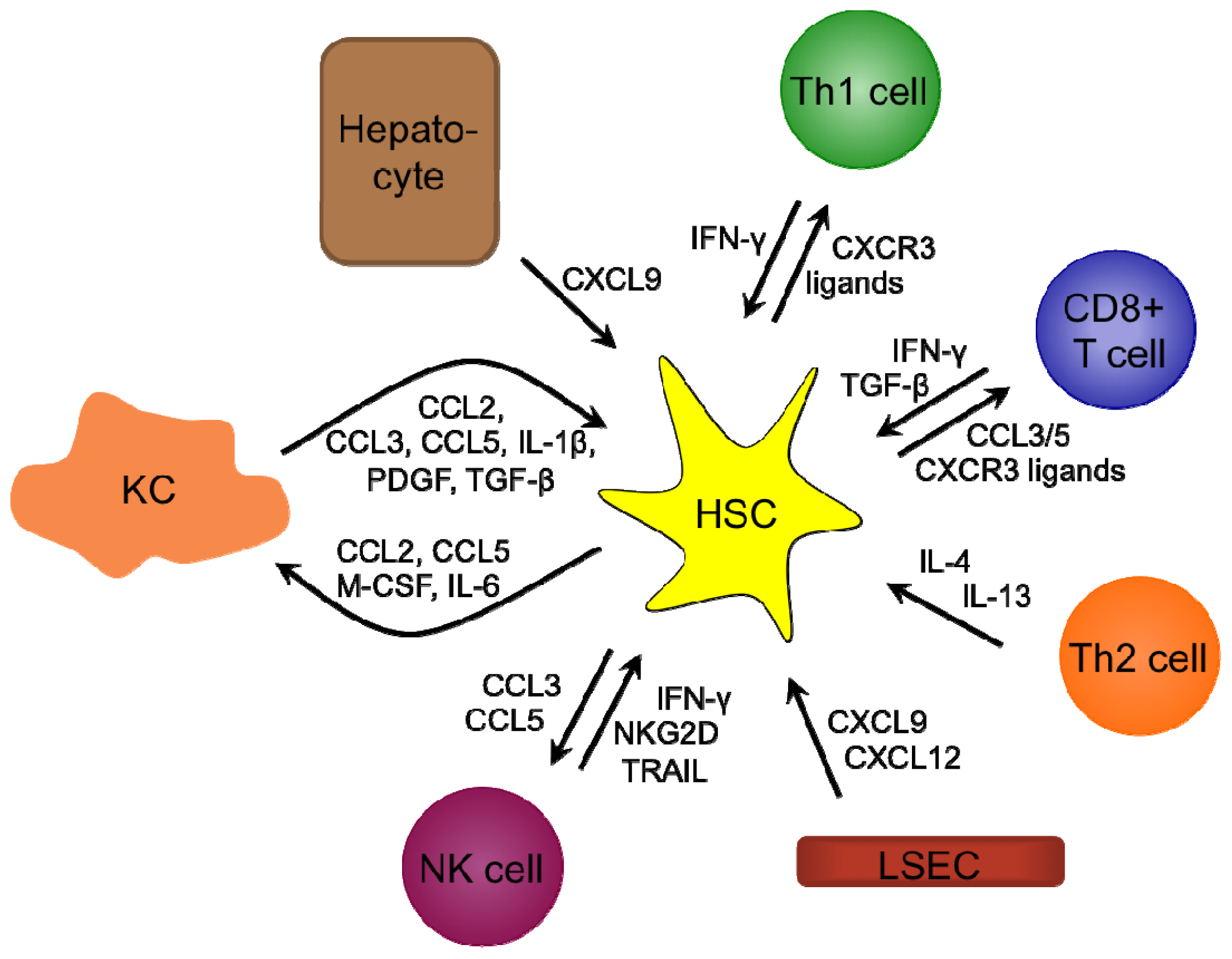
3.7. Liver Fibrosis
3.8. Liver Cancer
4. Chemokine Signaling as Therapeutic Target for Liver-Related Diseases
4.1. Acute Liver Injury
4.2. Metabolic Syndrome
4.3. Liver Fibrosis
4.4. Liver Cancer
5. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| ACKR | atypical chemokine receptor |
| DAA | direct acting antiviral |
| DC | dendritic cell |
| gt | genotype |
| HCC | hepatocellular carcinoma |
| HCV | hepatitis C virus |
| HSC | hepatic stellate cell |
| IFN | interferon |
| IL | interleukin |
| IRF | interferon regulatory factor |
| LPS | lipopolysaccaride |
| LSEC | liver sinusoidal endothelial cell |
| MAVS | mitochondrial antiviral signaling protein |
| MDSC | myeloid-derived suppressor cell |
| NFκB | nuclear factor κB |
| KC | Kupffer cell |
| NK cell | natural killer cell |
| NS | non-structural |
| pegIFN | pegylated interferon-α |
| PAMP | pathogen associated molecular pattern |
| pDC | plasmacytoid dendritic cell |
| PKR | protein kinase R |
| PRR | pattern recognition receptor |
| RBV | ribavirin |
| RIG-I | retinoic acid inducible gene-I |
| TCPTP | T cell protein tyrosine phosphatase |
| TGF | transforming growth factor |
| Th | T-helper |
| TLR | Toll-like receptor |
| TNF | tumor necrosis factor |
| TRIF | TIR-domain-containing adaptor-inducing-interferon-β |
| Treg | regulatory T cell |
References
- Lavanchy, D. The global burden of hepatitis C. Liver Int. 2009, 29, 74–81. [Google Scholar]
- Lavanchy, D. Evolving epidemiology of hepatitis C virus. Clin. Microbiol. Infect. 2011, 17, 107–115. [Google Scholar]
- Hoofnagle, J.H. Hepatitis C: The clinical spectrum of disease. Hepatology 1997, 26, 15S–20S. [Google Scholar]
- Arzumanyan, A.; Reis, H.M.G.P.V.; Feitelson, M.A. Pathogenic mechanisms in HBV- and HCV-associated hepatocellular carcinoma. Nat. Rev. Cancer 2013, 13, 123–135. [Google Scholar]
- Moradpour, D.; Penin, F.; Rice, C.M. Replication of hepatitis C virus. Nat. Rev. Microbiol. 2007, 5, 453–463. [Google Scholar]
- Penin, F.; Dubuisson, J.; Rey, F.A.; Moradpour, D.; Pawlotsky, J.-M. Structural biology of hepatitis C virus. Hepatology 2004, 39, 5–19. [Google Scholar]
- Nakano, T.; Lau, G.M.G.; Lau, G.M.L.; Sugiyama, M.; Mizokami, M. An updated analysis of hepatitis C virus genotypes and subtypes based on the complete coding region. Liver Int. 2011, 32, 339–345. [Google Scholar]
- Micallef, J.M.; Kaldor, J.M.; Dore, G.J. Spontaneous viral clearance following acute hepatitis C infection: A systematic review of longitudinal studies. J. Viral Hepat. 2006, 13, 34–41. [Google Scholar]
- Bode, J.G.; Brenndorfer, E.D.; Haussinger, D. Subversion of innate host antiviral strategies by the hepatitis C virus. Arch. Biochem. Biophys. 2007, 462, 254–265. [Google Scholar]
- Loo, Y.-M.; Owen, D.M.; Li, K.; Erickson, A.K.; Johnson, C.L.; Fish, P.M.; Carney, D.S.; Wang, T.; Ishida, H.; Yoneyama, M.; et al. Viral and therapeutic control of IFN-β promoter stimulator 1 during hepatitis C virus infection. Proc. Natl. Acad. Sci. USA 2006, 103, 6001–6006. [Google Scholar]
- Saito, T.; Owen, D.M.; Jiang, F.; Marcotrigiano, J.; Gale, M., Jr. Innate immunity induced by composition-dependent RIG-I recognition of hepatitis C virus RNA. Nature 2008, 454, 523–527. [Google Scholar]
- Wang, N.; Liang, Y.; Devaraj, S.; Wang, J.; Lemon, S.M.; Li, K. Toll-like receptor 3 mediates establishment of an antiviral state against hepatitis C virus in hepatoma cells. J. Virol. 2009, 83, 9824–9834. [Google Scholar]
- Wu, J.; Lu, M.; Meng, Z.; Trippler, M.; Broering, R.; Szczeponek, A.; Krux, F.; Dittmer, U.; Roggendorf, M.; Gerken, G.; et al. Toll-like receptor-mediated control of HBV replication by nonparenchymal liver cells in mice. Hepatology 2007, 46, 1769–1778. [Google Scholar]
- Seki, E.; Brenner, D.A. Toll-like receptors and adaptor molecules in liver disease: Update. Hepatology 2008, 48, 322–335. [Google Scholar]
- Li, K.; Li, N.L.; Wei, D.; Pfeffer, S.R.; Fan, M.; Pfeffer, L.M. Activation of chemokine and inflammatory cytokine response in hepatitis C virus-infected hepatocytes depends on toll-like receptor 3 sensing of hepatitis C virus double-stranded RNA intermediates. Hepatology 2012, 55, 666–675. [Google Scholar]
- Shimoike, T.; McKenna, S.A.; Lindhout, D.A.; Puglisi, J.D. Translational insensitivity to potent activation of PKR by HCV IRES RNA. Antivir. Res. 2009, 83, 228–237. [Google Scholar]
- Arnaud, N.; Dabo, S.; Akazawa, D.; Fukasawa, M.; Shinkai-Ouchi, F.; Hugon, J.; Wakita, T.; Meurs, E.F. Hepatitis C virus reveals a novel early control in acute immune response. PLoS Pathog. 2011, 7, e1002289. [Google Scholar]
- Meylan, E.; Curran, J.; Hofmann, K.; Moradpour, D.; Binder, M.; Bartenschlager, R.; Tschopp, J. Cardif is an adaptor protein in the RIG-I antiviral pathway and is targeted by hepatitis C virus. Nature 2005, 437, 1167–1172. [Google Scholar]
- Li, X.D.; Sun, L.; Seth, R.B.; Pineda, G.; Chen, Z.J. Hepatitis C virus protease NS3/4A cleaves mitochondrial antiviral signaling protein off the mitochondria to evade innate immunity. Proc. Natl. Acad. Sci. USA 2005, 102, 17717–17722. [Google Scholar]
- Li, K.; Foy, E.; Ferreon, J.C.; Nakamura, M.; Ferreon, A.C.; Ikeda, M.; Ray, S.C.; Gale, M., Jr; Lemon, S.M. Immune evasion by hepatitis C virus NS3/4A protease-mediated cleavage of the Toll-like receptor 3 adaptor protein TRIF. Proc. Natl. Acad. Sci. USA 2005, 102, 2992–2997. [Google Scholar]
- Gale, M.J.; Korth, M.J.; Tang, N.M.; Tan, S.L.; Hopkins, D.A.; Dever, T.E.; Polyak, S.J.; Gretch, D.R.; Katze, M.G. Evidence that hepatitis C virus resistance to interferon is mediated through repression of the PKR protein kinase by the nonstructural 5A protein. Virology 1997, 230, 217–227. [Google Scholar]
- Pavio, N.; Taylor, D.R.; Lai, M.M.C. Detection of a novel unglycosylated form of hepatitis C virus E2 envelope protein that is located in the cytosol and interacts with PKR. J. Virol. 2002, 76, 1265–1272. [Google Scholar]
- Taylor, D.R.; Shi, S.T.; Romano, P.R.; Barber, G.N.; Lai, M.M. Inhibition of the interferon-inducible protein kinase PKR by HCV E2 protein. Science 1999, 285, 107–110. [Google Scholar]
- Ge, D.; Fellay, J.; Thompson, A.J.; Simon, J.S.; Shianna, K.V.; Urban, T.J.; Heinzen, E.L.; Qiu, P.; Bertelsen, A.H.; Muir, A.J.; et al. Genetic variation in IL28B predicts hepatitis C treatment-induced viral clearance. Nature 2009, 461, 399–401. [Google Scholar]
- Poordad, F.; McCone, J.; Bacon, B.R.; Bruno, S.; Manns, M.P.; Sulkowski, M.S.; Jacobson, I.M.; Reddy, K.R.; Goodman, Z.D.; Boparai, N.; et al. Boceprevir for untreated chronic HCV genotype 1 infection. N. Engl. J. Med. 2011, 364, 1195–1206. [Google Scholar]
- Jacobson, I.M.; McHutchison, J.G.; Dusheiko, G.; di Bisceglie, A.M.; Reddy, K.R.; Bzowej, N.H.; Marcellin, P.; Muir, A.J.; Ferenci, P.; Flisiak, R.; et al. Telaprevir for previously untreated chronic hepatitis C virus infection. N. Engl. J. Med. 2011, 364, 2405–2416. [Google Scholar]
- Fried, M.W.; Buti, M.; Dore, G.J.; Flisiak, R.; Ferenci, P.; Jacobson, I.; Marcellin, P.; Manns, M.; Nikitin, I.; Poordad, F.; et al. Once-daily simeprevir (TMC435) with pegylated interferon and ribavirin in treatment-naïve genotype 1 hepatitis C: The randomized PILLAR study. Hepatology 2013, 58, 1918–1929. [Google Scholar]
- Gane, E.J.; Stedman, C.A.; Hyland, R.H.; Ding, X.; Svarovskaia, E.; Symonds, W.T.; Hindes, R.G.; Berrey, M.M. Nucleotide polymerase inhibitor sofosbuvir plus ribavirin for hepatitis C. N. Engl. J. Med. 2013, 368, 34–44. [Google Scholar]
- Flisiak, R.; Jaroszewicz, J.; Parfieniuk-Kowerda, A. Emerging treatments for hepatitis C. Exp. Opin. Emerg. Drugs 2013, 18, 461–475. [Google Scholar]
- Sulkowski, M.S.; Gardiner, D.F.; Rodriguez-Torres, M.; Reddy, K.R.; Hassanein, T.; Jacobson, I.; Lawitz, E.; Lok, A.S.; Hinestrosa, F.; Thuluvath, P.J.; et al. Daclatasvir plus sofosbuvir for previously treated or untreated chronic HCV infection. N. Engl. J. Med. 2014, 370, 211–221. [Google Scholar]
- Westra, I.M.; Oosterhuis, D.; Groothuis, G.M.M.; Olinga, P. Precision-cut liver slices as a model for the early onset of liver fibrosis to test antifibrotic drugs. Toxicol. Appl. Pharmacol. 2013, 274, 1–11. [Google Scholar]
- Hong, F.; Chou, H.; Fiel, M.I.; Friedman, S.L. Antifibrotic activity of sorafenib in experimental hepatic fibrosis: Refinement of inhibitory targets dosing and window of efficacy in vivo. Digest. Dis. Sci. 2013, 58, 257–264. [Google Scholar]
- Wilhelm, S.M.; Adnane, L.; Newell, P.; Villanueva, A.; Llovet, J.M.; Lynch, M. Preclinical overview of sorafenib a multikinase inhibitor that targets both Raf and VEGF and PDGF receptor tyrosine kinase signaling. Mol. Cancer Ther. 2008, 7, 3129–3140. [Google Scholar]
- Wang, Y.; Gao, J.; Zhang, D.; Zhang, J.; Ma, J.; Jiang, H. New insights into the antifibrotic effects of sorafenib on hepatic stellate cells and liver fibrosis. J. Hepatol. 2010, 53, 132–144. [Google Scholar]
- Bowen, D.G.; Walker, C.M. Adaptive immune responses in acute and chronic hepatitis C virus infection. Nature 2005, 436, 946–952. [Google Scholar]
- Bode, J.G.; Brenndorfer, E.D.; Haussinger, D. Hepatitis C virus (HCV) employs multiple strategies to subvert the host innate antiviral response. Biol. Chem. 2008, 389, 1283–1298. [Google Scholar]
- Brenndorfer, E.D.; Sallberg, M. Hepatitis C virus-mediated modulation of cellular immunity. Arch. Immun. Ther. Exp. 2012, 60, 315–329. [Google Scholar]
- Zlotnik, A.; Yoshie, O. The chemokine superfamily revisited. Immunity 2012, 36, 705–716. [Google Scholar]
- Luster, A.D. Chemokines—Chemotactic cytokines that mediate inflammation. N. Engl. J. Med. 1998, 338, 436–445. [Google Scholar]
- Moser, B.; Wolf, M.; Walz, A.; Loetscher, P. Chemokines: Multiple levels of leukocyte migration control. Trends Immun. 2004, 25, 75–84. [Google Scholar]
- Zhu, Y.; Murakami, F. Chemokine CXCL12 and its receptors in the developing central nervous system: Emerging themes and future perspectives. Dev. Neurobiol. 2012, 72, 1349–1362. [Google Scholar]
- Zou, Y.R.; Kottmann, A.H.; Kuroda, M.; Taniuchi, I.; Littman, D.R. Function of the chemokine receptor CXCR4 in haematopoiesis and in cerebellar development. Nature 1998, 393, 595–599. [Google Scholar]
- Calderon, L.; Boehm, T. Three chemokine receptors cooperatively regulate homing of hematopoietic progenitors to the embryonic mouse thymus. Proc. Natl. Acad. Sci. USA 2011, 108, 7517–7122. [Google Scholar]
- Esche, C.; Stellato, C.; Beck, L.A. Chemokines: Key players in innate and adaptive immunity. J. Investig. Dermatol. 2005, 125, 615–628. [Google Scholar]
- Dimberg, A. Chemokines in angiogenesis. Curr. Top. Microbiol. Immun. 2010, 341, 59–80. [Google Scholar]
- Flad, H.D.; Brandt, E. Platelet-derived chemokines: Pathophysiology and therapeutic aspects. Cell. Mol. Life Sci. 2010, 67, 2363–2386. [Google Scholar]
- Zlotnik, A.; Yoshie, O.; Nomiyama, H. The chemokine and chemokine receptor superfamilies and their molecular evolution. Genome Biol. 2006, 7, 243. [Google Scholar]
- Oo, Y.H.; Adams, D.H. The role of chemokines in the recruitment of lymphocytes to the liver. J. Autoimmun. 2010, 34, 45–54. [Google Scholar]
- Schall, T.J.; Proudfoot, A.E. I Overcoming hurdles in developing successful drugs targeting chemokine receptors. Nat. Publ. Gr. 2011, 11, 355–363. [Google Scholar]
- Loetscher, P.; Pellegrino, A.; Gong, J.H.; Mattioli, I.; Loetscher, M.; Bardi, G.; Baggiolini, M.; Clark-Lewis, I. The ligands of CXC chemokine receptor 3 I-TAC Mig and IP10 are natural antagonists for CCR3. J. Biol. Chem. 2001, 276, 2986–2991. [Google Scholar]
- Nibbs, R.J.B.; Graham, G.J. Immune regulation by atypical chemokine receptors. Nat. Rev. Immun. 2013, 13, 815–829. [Google Scholar]
- Fra, A.M.; Locati, M.; Otero, K.; Sironi, M.; Signorelli, P.; Massardi, M.L.; Gobbi, M.; Vecchi, A.; Sozzani, S.; Mantovani, A. Cutting edge: Scavenging of inflammatory CC chemokines by the promiscuous putatively silent chemokine receptor D6. J. Immunol. 2003, 170, 2279–2282. [Google Scholar]
- Nibbs, R.J.; Wylie, S.M.; Yang, J.; Landau, N.R.; Graham, G.J. Cloning and characterization of a novel promiscuous human β-chemokine receptor D6. J. Biol. Chem. 1997, 272, 32078–32083. [Google Scholar]
- Berres, M.L.; Trautwein, C.; Zaldivar, M.M.; Schmitz, P.; Pauels, K.; Lira, S.A.; Tacke, F.; Wasmuth, H.E. The chemokine scavenging receptor D6 limits acute toxic liver injury in vivo. Biol. Chem. 2009, 390, 1039–1045. [Google Scholar]
- Wasmuth, H.; Tacke, F.; Trautwein, C. Chemokines in liver inflammation and fibrosis. Semin. Liver Dis. 2010, 30, 215–225. [Google Scholar]
- Nemeth, E.; Baird, A.W.; O’Farrelly, C. Microanatomy of the liver immune system. Semin. Immunopathol. 2009, 31, 333–343. [Google Scholar]
- Tiegs, G.; Lohse, A.W. Immune tolerance: What is unique about the liver. J. Autoimmun. 2010, 34, 1–6. [Google Scholar]
- Bonecchi, R.; Bianchi, G.; Bordignon, P.P.; D’Ambrosio, D.; Lang, R.; Borsatti, A.; Sozzani, S.; Allavena, P.; Gray, P.A.; Mantovani, A.; et al. Differential expression of chemokine receptors and chemotactic responsiveness of type 1 T helper cells (Th1s) and Th2s. J. Exp. Med. 1998, 187, 129–134. [Google Scholar]
- Crispe, I.N. Hepatic T cells and liver tolerance. Nat. Rev. Immunol. 2003, 3, 51–62. [Google Scholar]
- Viola, A.; Luster, A.D. Chemokines and their receptors: Drug targets in immunity and inflammation. Ann. Rev. Pharmacol. Toxicol. 2008, 48, 171–197. [Google Scholar]
- Zimmermann, H.W.; Tacke, F. Modification of chemokine pathways and immune cell infiltration as a novel therapeutic approach in liver inflammation and fibrosis. Inflamm. Allerg. Drug Targets 2011, 10, 509–536. [Google Scholar]
- Bautista, A.P. Chronic alcohol intoxication primes Kupffer cells and endothelial cells for enhanced CC-chemokine production and concomitantly suppresses phagocytosis and chemotaxis. Front. Biosci. 2002, 7, a117–a125. [Google Scholar]
- Hokeness, K.L.; Kuziel, W.A.; Biron, C.A.; Salazar-Mather, T.P. Monocyte chemoattractant protein-1 and CCR2 interactions are required for IFN-α/β-induced inflammatory responses and antiviral defense in liver. J. Immunol. 2005, 174, 1549–1556. [Google Scholar]
- Loetscher, P.; Uguccioni, M.; Bordoli, L.; Baggiolini, M.; Moser, B.; Chizzolini, C.; Dayer, J.M. CCR5 is characteristic of Th1 lymphocytes. Nature 1998, 391, 344–345. [Google Scholar]
- Megjugorac, N.J.; Young, H.A.; Amrute, S.B.; Olshalsky, S.L.; Fitzgerald-Bocarsly, P. Virally stimulated plasmacytoid dendritic cells produce chemokines and induce migration of T and NK cells. J. Leukoc. Biol. 2004, 75, 504–514. [Google Scholar]
- Piqueras, B.; Connolly, J.; Freitas, H.; Palucka, A.K.; Banchereau, J. Upon viral exposure myeloid and plasmacytoid dendritic cells produce 3 waves of distinct chemokines to recruit immune effectors. Blood 2006, 107, 2613–2618. [Google Scholar]
- Salazar-Mather, T.P.; Lewis, C.A.; Biron, C.A. Type I interferons regulate inflammatory cell trafficking and macrophage inflammatory protein 1α delivery to the liver. J. Clin. Investig. 2002, 110, 321–330. [Google Scholar]
- Salazar-Mather, T.P.; Orange, J.S.; Biron, C.A. Early murine cytomegalovirus (MCMV) infection induces liver natural killer (NK) cell inflammation and protection through macrophage inflammatory protein 1α (MIP-1α)-dependent pathways. J. Exp. Med. 1998, 187, 1–14. [Google Scholar]
- Sallusto, F.; Schaerli, P.; Loetscher, P.; Schaniel, C.; Lenig, D.; Mackay, C.R.; Qin, S.; Lanzavecchia, A. Rapid and coordinated switch in chemokine receptor expression during dendritic cell maturation. Eur. J. Immunol. 1998, 28, 2760–2769. [Google Scholar]
- Seki, E.; De Minicis, S.; Österreicher, C.H.; Kluwe, J.; Osawa, Y.; Brenner, D.A.; Schwabe, R.F. TLR4 enhances TGF-β signaling and hepatic fibrosis. Nat. Med. 2007, 13, 1324–1332. [Google Scholar]
- Zimmermann, H.W.; Seidler, S.; Nattermann, J.; Gassler, N.; Hellerbrand, C.; Zernecke, A.; Tischendorf, J.J.W.; Luedde, T.; Weiskirchen, R.; Trautwein, C.; et al. Functional contribution of elevated circulating and hepatic non-classical CD14CD16 monocytes to inflammation and human liver fibrosis. PLoS One 2010, 5, e11049. [Google Scholar]
- Carr, M.W.; Roth, S.J.; Luther, E.; Rose, S.S.; Springer, T.A. Monocyte chemoattractant protein 1 acts as a T-lymphocyte chemoattractant. Proc. Natl. Acad. Sci. USA 1994, 91, 3652–3656. [Google Scholar]
- Decalf, J.; Fernandes, S.; Longman, R.; Ahloulay, M.; Audat, F.; Lefrerre, F.; Rice, C.M.; Pol, S.; Albert, M.L. Plasmacytoid dendritic cells initiate a complex chemokine and cytokine network and are a viable drug target in chronic HCV patients. J. Exp. Med. 2007, 204, 2423–2437. [Google Scholar]
- Fuentes, M.E.; Durham, S.K.; Swerdel, M.R.; Lewin, A.C.; Barton, D.S.; Megill, J.R.; Bravo, R.; Lira, S.A. Controlled recruitment of monocytes and macrophages to specific organs through transgenic expression of monocyte chemoattractant protein-1. J. Immunol. 1995, 155, 5769–5776. [Google Scholar]
- Gu, L.; Tseng, S.; Horner, R.M.; Tam, C.; Loda, M.; Rollins, B.J. Control of TH2 polarization by the chemokine monocyte chemoattractant protein-1. Nature 2000, 404, 407–411. [Google Scholar]
- Gunn, M.D.; Nelken, N.A.; Liao, X.; Williams, L.T. Monocyte chemoattractant protein-1 is sufficient for the chemotaxis of monocytes and lymphocytes in transgenic mice but requires an additional stimulus for inflammatory activation. J. Immunol. 1997, 158, 376–383. [Google Scholar]
- Maghazachi, A.A.; al-Aoukaty, A.; Schall, T.J. C-C chemokines induce the chemotaxis of NK and IL-2-activated NK cells Role for G proteins. J. Immunol. 1994, 153, 4969–4977. [Google Scholar]
- Alferink, J.; Lieberam, I.; Reindl, W.; Behrens, A.; Weiss, S.; Hüser, N.; Gerauer, K.; Ross, R.; Reske-Kunz, A.B.; Ahmad-Nejad, P.; et al. Compartmentalized production of CCL17 in vivo: Strong inducibility in peripheral dendritic cells contrasts selective absence from the spleen. J. Exp. Med. 2003, 197, 585–599. [Google Scholar]
- Andrew, D.P.; Chang, M.S.; McNinch, J.; Wathen, S.T.; Rihanek, M.; Tseng, J.; Spellberg, J.P.; Elias, C.G. STCP-1 (MDC) CC chemokine acts specifically on chronically activated Th2 lymphocytes and is produced by monocytes on stimulation with Th2 cytokines IL-4 and IL-13. J. Immunol. 1998, 161, 5027–5038. [Google Scholar]
- Hirahara, K.; Liu, L.; Clark, R.A.; Yamanaka, K.-i.; Fuhlbrigge, R.C.; Kupper, T.S. The majority of human peripheral blood CD4+ CD25highFoxp3+ regulatory T cells bear functional skin-homing receptors. J. Immunol. 2006, 177, 4488–4494. [Google Scholar]
- Imai, T.; Nagira, M.; Takagi, S.; Kakizaki, M.; Nishimura, M.; Wang, J.; Gray, P.W.; Matsushima, K.; Yoshie, O. Selective recruitment of CCR4-bearing Th2 cells toward antigen-presenting cells by the CC chemokines thymus and activation-regulated chemokine and macrophage-derived chemokine. Int. Immunol. 1999, 11, 81–88. [Google Scholar]
- Mariani, M.; Lang, R.; Binda, E.; Panina-Bordignon, P.; D’Ambrosio, D. Dominance of CCL22 over CCL17 in induction of chemokine receptor CCR4 desensitization and internalization on human Th2 cells. Eur. J. Immunol. 2004, 34, 231–240. [Google Scholar]
- Sallusto, F.; Lenig, D.; Mackay, C.R.; Lanzavecchia, A. Flexible programs of chemokine receptor expression on human polarized T helper 1 and 2 lymphocytes. J. Exp. Med. 1998, 187, 875–883. [Google Scholar]
- Ahuja, S.K.; Murphy, P.M. The CXC chemokines growth-regulated oncogene GROα GROβ GROγ neutrophil-activating peptide-2 and epithelial cell-derived neutrophil-activating peptide-78 are potent agonists for the type B but not the type A human interleukin-8 receptor. J. Biol. Chem. 1996, 271, 20545–20550. [Google Scholar]
- Clarke, C.; Kuboki, S.; Sakai, N.; Kasten, K.R.; Tevar, A.D.; Schuster, R.; Blanchard, J.; Caldwell, C.C.; Edwards, M.J.; Lentsch, A.B. CXC chemokine receptor-1 is expressed by hepatocytes and regulates liver recovery after hepatic ischemia/reperfusion injury. Hepatology 2011, 53, 261–271. [Google Scholar]
- Hess, C.; Means, T.K.; Autissier, P.; Woodberry, T.; Altfeld, M.; Addo, M.M.; Frahm, N.; Brander, C.; Walker, B.D.; Luster, A.D. IL-8 responsiveness defines a subset of CD8 T cells poised to kill. Blood 2004, 104, 3463–3471. [Google Scholar]
- Heydtmann, M.; Adams, D.H. Chemokines in the immunopathogenesis of hepatitis C infection. Hepatology 2009, 49, 676–688. [Google Scholar]
- Huber, A.R.; Kunkel, S.L.; Todd, R.F.; Weiss, S.J. Regulation of transendothelial neutrophil migration by endogenous interleukin-8. Science 1991, 254, 99–102. [Google Scholar]
- Matsushima, K.; Oppenheim, J.J. Interleukin 8 and MCAF: Novel inflammatory cytokines inducible by IL 1 and TNF. Cytokine 1989, 1, 2–13. [Google Scholar]
- Ohkubo, K.; Masumoto, T.; Horiike, N.; Onji, M. Induction of CINC (interleukin-8) production in rat liver by non-parenchymal cells. J. Gastroenterol. Hepatol. 1998, 13, 696–702. [Google Scholar]
- Pelletier, M.; Maggi, L.; Micheletti, A.; Lazzeri, E.; Tamassia, N.; Costantini, C.; Cosmi, L.; Lunardi, C.; Annunziato, F.; Romagnani, S.; et al. Evidence for a cross-talk between human neutrophils and Th17 cells. Blood 2010, 115, 335–343. [Google Scholar]
- Strieter, R.M.; Kunkel, S.L.; Showell, H.J.; Remick, D.G.; Phan, S.H.; Ward, P.A.; Marks, R.M. Endothelial cell gene expression of a neutrophil chemotactic factor by TNF-α LPS and IL-1β. Science 1989, 243, 1467–1469. [Google Scholar]
- Thornton, A.J.; Strieter, R.M.; Lindley, I.; Baggiolini, M.; Kunkel, S.L. Cytokine-induced gene expression of a neutrophil chemotactic factor/IL-8 in human hepatocytes. J. Immunol. 1990, 144, 2609–2613. [Google Scholar]
- Van Damme, J.; Decock, B.; Lenaerts, J.P.; Conings, R.; Bertini, R.; Mantovani, A.; Billiau, A. Identification by sequence analysis of chemotactic factors for monocytes produced by normal and transformed cells stimulated with virus double-stranded RNA or cytokine. Eur. J. Immunol. 1989, 19, 2367–2373. [Google Scholar]
- Wagoner, J.; Austin, M.; Green, J.; Imaizumi, T.; Casola, A.; Brasier, A.; Khabar, K.S.A.; Wakita, T.; Gale, M.; Polyak, S.J. Regulation of CXCL-8 (interleukin-8) induction by double-stranded RNA signaling pathways during hepatitis C virus infection. J. Virol. 2007, 81, 309–318. [Google Scholar]
- Brownell, J.; Polyak, S.J. Molecular pathways: Hepatitis C virus CXCL10 and the inflammatory road to liver cancer. Clin. Cancer Res. 2013, 19, 1347–1352. [Google Scholar]
- Cole, K.E.; Strick, C.A.; Paradis, T.J.; Ogborne, K.T.; Loetscher, M.; Gladue, R.P.; Lin, W.; Boyd, J.G.; Moser, B.; Wood, D.E.; et al. Interferon-inducible T cell alpha chemoattractant (I-TAC): A novel non-ELR CXC chemokine with potent activity on activated T cells through selective high affinity binding to CXCR3. J. Exp. Med. 1998, 187, 2009–2021. [Google Scholar]
- Crispe, I.N. Migration of lymphocytes into hepatic sinusoids. J. Hepatol. 2012, 57, 218–220. [Google Scholar]
- Helbig, K.J.; Ruszkiewicz, A.; Semendric, L.; Harley, H.A.J.; McColl, S.R.; Beard, M.R. Expression of the CXCR3 ligand I-TAC by hepatocytes in chronic hepatitis C and its correlation with hepatic inflammation. Hepatology 2004, 39, 1220–1229. [Google Scholar]
- Hokeness, K.L.; Deweerd, E.S.; Munks, M.W.; Lewis, C.A.; Gladue, R.P.; Salazar-Mather, T.P. CXCR3-dependent recruitment of antigen-specific T lymphocytes to the liver during murine cytomegalovirus infection. J. Virol. 2007, 81, 1241–1250. [Google Scholar]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011, 11, 519–531. [Google Scholar]
- Lasagni, L.; Francalanci, M.; Annunziato, F.; Lazzeri, E.; Giannini, S.; Cosmi, L.; Sagrinati, C.; Mazzinghi, B.; Orlando, C.; Maggi, E.; et al. An Alternatively spliced bariant of CXCR3 mediates the inhibition of endothelial cell growth induced by IP-10 mig and I-TAC and acts as functional receptor for platelet factor 4. J. Exp. Med. 2003, 197, 1537–1549. [Google Scholar]
- Mueller, A.; Meiser, A.; McDonagh, E.M.; Fox, J.M.; Petit, S.J.; Xanthou, G.; Williams, T.J.; Pease, J.E. CXCL4-induced migration of activated T lymphocytes is mediated by the chemokine receptor CXCR3. J. Leukoc. Biol. 2008, 83, 875–882. [Google Scholar]
- Bernhagen, J.; Krohn, R.; Lue, H.; Gregory, J.L.; Zernecke, A.; Koenen, R.R.; Dewor, M.; Georgiev, I.; Schober, A.; Leng, L.; et al. MIF is a noncognate ligand of CXC chemokine receptors in inflammatory and atherogenic cell recruitment. Nat. Med. 2007, 13, 587–596. [Google Scholar]
- Burns, J.M.; Summers, B.C.; Wang, Y.; Melikian, A.; Berahovich, R.; Miao, Z.; Penfold, M.E.T.; Sunshine, M.J.; Littman, D.R.; Kuo, C.J.; et al. A novel chemokine receptor for SDF-1 and I-TAC involved in cell survival cell adhesion and tumor development. J. Exp. Med. 2006, 203, 2201–2213. [Google Scholar]
- Gupta, S.K.; Pillarisetti, K. Cutting edge: CXCR4-Lo: Molecular cloning and functional expression of a novel human CXCR4 splice variant. J. Immunol. 1999, 163, 2368–2372. [Google Scholar]
- Hong, F.; Tuyama, A.; Lee, T.F.; Loke, J.; Agarwal, R.; Cheng, X.; Garg, A.; Fiel, M.I.; Schwartz, M.; Walewski, J.; et al. Hepatic stellate cells express functional CXCR4: Role in stromal cell-derived factor-1α-mediated stellate cell activation. Hepatology 2009, 49, 2055–2067. [Google Scholar]
- Levoye, A.; Balabanian, K.; Baleux, F.; Bachelerie, F.; Lagane, B. CXCR7 heterodimerizes with CXCR4 and regulates CXCL12-mediated G protein signaling. Blood 2009, 113, 6085–6093. [Google Scholar]
- Maksym, R.B.; Tarnowski, M.; Grymula, K.; Tarnowska, J.; Wysoczynski, M.; Liu, R.; Czerny, B.; Ratajczak, J.; Kucia, M.; Ratajczak, M.Z. The role of stromal-derived factor-1—CXCR7 axis in development and cancer. Eur. J. Pharmacol. 2009, 625, 31–40. [Google Scholar]
- Saini, V.; Marchese, A.; Majetschak, M. CXC chemokine receptor 4 is a cell surface receptor for extracellular ubiquitin. J. Biol. Chem. 2010, 285, 15566–15576. [Google Scholar]
- Geissmann, F.; Cameron, T.O.; Sidobre, S.; Manlongat, N.; Kronenberg, M.; Briskin, M.J.; Dustin, M.L.; Littman, D.R. Intravascular immune surveillance by CXCR6+ NKT cells patrolling liver sinusoids. PLoS Biol. 2005, 3, e113. [Google Scholar]
- Germanov, E.; Veinotte, L.; Cullen, R.; Chamberlain, E.; Butcher, E.C.; Johnston, B. Critical role for the chemokine receptor CXCR6 in homeostasis and activation of CD1d-restricted NKT cells. J. Immunol. 2008, 181, 81–91. [Google Scholar]
- Heydtmann, M.; Lalor, P.F.; Eksteen, J.A.; Hubscher, S.G.; Briskin, M.; Adams, D.H. CXC chemokine ligand 16 promotes integrin-mediated adhesion of liver-infiltrating lymphocytes to cholangiocytes and hepatocytes within the inflamed human liver. J. Immunol. 2005, 174, 1055–1062. [Google Scholar]
- Shields, P.L.; Morland, C.M.; Salmon, M.; Qin, S.; Hubscher, S.G.; Adams, D.H. Chemokine and chemokine receptor interactions provide a mechanism for selective T cell recruitment to specific liver compartments within hepatitis C-infected liver. J. Immunol. 1999, 163, 6236–6243. [Google Scholar]
- Nguyen, N.; de Esch, C.; Cameron, B.; Kumar, R.K.; Zekry, A.; Lloyd, A.R. Positioning of leukocyte subsets in the portal and lobular compartments of HCV-infected liver correlates with local chemokine expression. J. Gastroenterol. Hepatol. 2014, 2014. [Google Scholar] [CrossRef]
- Zeremski, M.; Petrovic, L.M.; Talal, A.H. The role of chemokines as inflammatory mediators in chronic hepatitis C virus infection. J. Viral. Hepat. 2007, 14, 675–687. [Google Scholar]
- Lichterfeld, M.; Leifeld, L.; Nischalke, H.D.; Rockstroh, J.K.; Hess, L.; Sauerbruch, T.; Spengler, U. Reduced CC chemokine receptor (CCR) 1 and CCR5 surface expression on peripheral blood T lymphocytes from patients with chronic hepatitis C infection. J. Infect. Dis. 2002, 185, 1803–1807. [Google Scholar]
- Apolinario, A.; Majano, P.L.; Lorente, R.; Nunez, O.; Clemente, G.; Garcia-Monzon, C. Gene expression profile of T-cell-specific chemokines in human hepatocyte-derived cells: Evidence for a synergistic inducer effect of cytokines and hepatitis C virus proteins. J. Viral. Hepat. 2005, 12, 27–37. [Google Scholar]
- Sillanpaa, M.; Kaukinen, P.; Melen, K.; Julkunen, I. Hepatitis C virus proteins interfere with the activation of chemokine gene promoters and downregulate chemokine gene expression. J. Gen. Virol. 2008, 89, 432–443. [Google Scholar]
- Brenndorfer, E.D.; Brass, A.; Karthe, J.; Ahlen, G.; Bode, J.G.; Sallberg, M. Cleavage of the T cell protein tyrosine phosphatase by the hepatitis C virus nonstructural 3/4A protease induces a Th1 to Th2 shift reversible by ribavirin therapy. J. Immunol. 2014, 192, 1671–80. [Google Scholar]
- Asselah, T.; Bieche, I.; Laurendeau, I.; Paradis, V.; Vidaud, D.; Degott, C.; Martinot, M.; Bedossa, P.; Valla, D.; Vidaud, M.; et al. Liver gene expression signature of mild fibrosis in patients with chronic hepatitis, C. Gastroenterology 2005, 129, 2064–2075. [Google Scholar]
- Zhdanov, K.V.; Gusev, D.A.; Chirskii, V.S.; Sysoev, K.A.; Iakubovskaia, L.A.; Shakhmanov, D.M.; Totolian, A.A. Chronic HCV-infection and expression of mRNA of CC-chemokines and their receptors. Zhurnal Mikrobiol. Epidemiol. Immunobiol. 2008, 2008, 73–78. (In Russian). [Google Scholar]
- Brenndorfer, E.D.; Weiland, M.; Frelin, L.; Derk, E.; Ahlen, G.; Jiao, J.; Bode, J.G.; Sallberg, M. Anti-tumor necrosis factor α treatment promotes apoptosis and prevents liver regeneration in a transgenic mouse model of chronic hepatitis C. Hepatology 2010, 52, 1553–1563. [Google Scholar]
- Soo, H.M.; Garzino-Demo, A.; Hong, W.; Tan, Y.H.; Tan, Y.J.; Goh, P.; Lim, S.G.; Lim, S.P. Expression of a full-length hepatitis C virus cDNA up-regulates the expression of CC chemokines MCP-1 and RANTES. Virology 2002, 303, 253–277. [Google Scholar]
- Brenndorfer, E.D.; Brass, A.; Soderholm, J.; Frelin, L.; Aleman, S.; Bode, J.G.; Sallberg, M. Hepatitis C virus non-structural 3/4A protein interferes with intrahepatic interferon-γ production. Gut 2012, 61, 589–596. [Google Scholar]
- Riezu-Boj, J.I.; Larrea, E.; Aldabe, R.; Guembe, L.; Casares, N.; Galeano, E.; Echeverria, I.; Sarobe, P.; Herrero, I.; Sangro, B.; et al. Hepatitis C virus induces the expression of CCL17 and CCL22 chemokines that attract regulatory T cells to the site of infection. J. Hepatol. 2011, 54, 422–431. [Google Scholar]
- Gruener, N.H.; Lechner, F.; Jung, M.C.; Diepolder, H.; Gerlach, T.; Lauer, G.; Walker, B.; Sullivan, J.; Phillips, R.; Pape, G.R.; et al. Sustained dysfunction of antiviral CD8+ T lymphocytes after infection with hepatitis C virus. J. Virol. 2001, 75, 5550–5558. [Google Scholar]
- Kaplanski, G.; Farnarier, C.; Payan, M.J.; Bongrand, P.; Durand, J.M. Increased levels of soluble adhesion molecules in the serum of patients with hepatitis C Correlation with cytokine concentrations and liver inflammation and fibrosis. Dig. Dis. Sci. 1997, 42, 2277–2284. [Google Scholar]
- Polyak, S.J.; Khabar, K.S.; Rezeiq, M.; Gretch, D.R. Elevated levels of interleukin-8 in serum are associated with hepatitis C virus infection and resistance to interferon therapy. J. Virol. 2001, 75, 6209–6211. [Google Scholar]
- Bataller, R.; Paik, Y.H.; Lindquist, J.N.; Lemasters, J.J.; Brenner, D.A. Hepatitis C virus core and nonstructural proteins induce fibrogenic effects in hepatic stellate cells. Gastroenterology 2004, 126, 529–540. [Google Scholar]
- Kadoya, H.; Nagano-Fujii, M.; Deng, L.; Nakazono, N.; Hotta, H. Nonstructural proteins 4A and 4B of hepatitis C virus transactivate the interleukin 8 promoter. Microbiol. Immunol. 2005, 49, 265–273. [Google Scholar]
- Zeremski, M.; Petrovic, L.M.; Chiriboga, L.; Brown, Q.B.; Yee, H.T.; Kinkhabwala, M.; Jacobson, I.M.; Dimova, R.; Markatou, M.; Talal, A.H. Intrahepatic levels of CXCR3-associated chemokines correlate with liver inflammation and fibrosis in chronic hepatitis C. Hepatology 2008, 48, 1440–1450. [Google Scholar]
- Helbig, K.J.; Ruszkiewicz, A.; Lanford, R.E.; Berzsenyi, M.D.; Harley, H.A.; McColl, S.R.; Beard, M.R. Differential expression of the CXCR3 ligands in chronic hepatitis C virus (HCV) infection and their modulation by HCV in vitro. J. Virol. 2009, 83, 836–846. [Google Scholar]
- Larrubia, J.R.; Calvino, M.; Benito, S.; Sanz-de-Villalobos, E.; Perna, C.; Perez-Hornedo, J.; Gonzalez-Mateos, F.; Garcia-Garzon, S.; Bienvenido, A.; Parra, T. The role of CCR5/CXCR3 expressing CD8+ cells in liver damage and viral control during persistent hepatitis C virus infection. J. Hepatol. 2007, 47, 632–641. [Google Scholar]
- Kaukinen, P.; Sillanpaa, M.; Kotenko, S.; Lin, R.; Hiscott, J.; Melen, K.; Julkunen, I. Hepatitis C virus NS2 and NS3/4A proteins are potent inhibitors of host cell cytokine/chemokine gene expression. Virol. J. 2006, 3, 66. [Google Scholar]
- Zekri, A.R.; Bahnassy, A.A.; Mohamed, W.S.; Alam El-Din, H.M.; Shousha, H.I.; Zayed, N.; Eldahshan, D.H.; Abdel-Aziz, A.O. Dynamic interplay between CXCL levels in chronic hepatitis C patients treated by interferon. Virol. J. 2013, 10, 218. [Google Scholar]
- Brownell, J.; Wagoner, J.; Lovelace, E.S.; Thirstrup, D.; Mohar, I.; Smith, W.; Giugliano, S.; Li, K.; Crispe, I.N.; Rosen, H.R.; et al. Independent parallel pathways to CXCL10 induction in HCV-infected hepatocytes. J. Hepatol. 2013, 59, 701–708. [Google Scholar]
- Zeremski, M.; Hooker, G.; Shu, M.A.; Winkelstein, E.; Brown, Q.; des Jarlais, D.C.; Tobler, L.H.; Rehermann, B.; Busch, M.P.; Edlin, B.R.; et al. Induction of CXCR3- and CCR5-associated chemokines during acute hepatitis C virus infection. J. Hepatol. 2011, 55, 545–553. [Google Scholar]
- Harvey, C.E.; Post, J.J.; Palladinetti, P.; Freeman, A.J.; Ffrench, R.A.; Kumar, R.K.; Marinos, G.; Lloyd, A.R. Expression of the chemokine IP-10 (CXCL10) by hepatocytes in chronic hepatitis C virus infection correlates with histological severity and lobular inflammation. J. Leukoc. Biol. 2003, 74, 360–369. [Google Scholar]
- Helbig, K.J.; George, J.; Beard, M.R. A novel I-TAC promoter polymorphic variant is functional in the presence of replicating HCV in vitro. J. Clin. Virol. 2005, 32, 137–143. [Google Scholar]
- Spengler, U.; Nattermann, J. Immunopathogenesis in hepatitis C virus cirrhosis. Clin. Sci. 2007, 112, 141–155. [Google Scholar]
- Wang, J.; Holmes, T.H.; Cheung, R.; Greenberg, H.B.; He, X.S. Expression of chemokine receptors on intrahepatic and peripheral lymphocytes in chronic hepatitis C infection: Its relationship to liver inflammation. J. Infect. Dis. 2004, 190, 989–997. [Google Scholar]
- Northfield, J.W.; Kasprowicz, V.; Lucas, M.; Kersting, N.; Bengsch, B.; Kim, A.; Phillips, R.E.; Walker, B.D.; Thimme, R.; Lauer, G.; et al. CD161 expression on hepatitis C virus-specific CD8+ T cells suggests a distinct pathway of T cell differentiation. Hepatology 2008, 47, 396–406. [Google Scholar]
- Apolinario, A.; Majano, P.L.; Alvarez-Perez, E.; Saez, A.; Lozano, C.; Vargas, J.; Garcia-Monzon, C. Increased expression of T cell chemokines and their receptors in chronic hepatitis C: Relationship with the histological activity of liver disease. Am. J. Gastroenterol. 2002, 97, 2861–2870. [Google Scholar]
- Palmer, C.; Corpuz, T.; Guirguis, M.; O’Toole, S.; Yan, K.; Bu, Y.; Jorgenson, J.; Talbot, M.; Loi, K.; Lloyd, A.; et al. The effect of obesity on intrahepatic cytokine and chemokine expression in chronic hepatitis C infection. Gut 2010, 59, 397–404. [Google Scholar]
- Mihm, S.; Schweyer, S.; Ramadori, G. Expression of the chemokine IP-10 correlates with the accumulation of hepatic IFN-γ and IL-18 mRNA in chronic hepatitis C but not in hepatitis B. J. Med. Virol. 2003, 70, 562–570. [Google Scholar]
- Woitas, R.P.; Ahlenstiel, G.; Iwan, A.; Rockstroh, J.K.; Brackmann, H.H.; Kupfer, B.; Matz, B.; Offergeld, R.; Sauerbruch, T.; Spengler, U. Frequency of the HIV-protective CC chemokine receptor 5-Delta32/Delta32 genotype is increased in hepatitis C. Gastroenterology 2002, 122, 1721–1728. [Google Scholar]
- Brenndorfer, E.D.; Karthe, J.; Frelin, L.; Cebula, P.; Erhardt, A.; Schulte am Esch, J.; Hengel, H.; Bartenschlager, R.; Sallberg, M.; Haussinger, D.; et al. Nonstructural 3/4A protease of hepatitis C virus activates epithelial growth factor-induced signal transduction by cleavage of the T-cell protein tyrosine phosphatase. Hepatology 2009, 49, 1810–1820. [Google Scholar]
- Rahbin, N.; Frelin, L.; Aleman, S.; Hultcrantz, R.; Sallberg, M.; Brenndorfer, E.D. Non-structural 3 protein expression is associated with T cell protein tyrosine phosphatase and viral RNA levels in chronic hepatitis C patients. Biochem. Biophys. Res. Commun. 2013, 433, 31–35. [Google Scholar]
- Shimoda, K.; Begum, N.A.; Shibuta, K.; Mori, M.; Bonkovsky, H.L.; Banner, B.F.; Barnard, G.F. Interleukin-8 and hIRH (SDF1-α/PBSF) mRNA expression and histological activity index in patients with chronic hepatitis C. Hepatology 1998, 28, 108–115. [Google Scholar]
- Tsukada, S.; Parsons, C.J.; Rippe, R.A. Mechanisms of liver fibrosis. Clin. Chim. Acta 2006, 364, 33–60. [Google Scholar]
- Imamura, M.; Ogawa, T.; Sasaguri, Y.; Chayama, K.; Ueno, H. Suppression of macrophage infiltration inhibits activation of hepatic stellate cells and liver fibrogenesis in rats. Gastroenterology 2005, 128, 138–146. [Google Scholar]
- Muhlbauer, M.; Bosserhoff, A.K.; Hartmann, A.; Thasler, W.E.; Weiss, T.S.; Herfarth, H.; Lock, G.; Scholmerich, J.; Hellerbrand, C. A novel MCP-1 gene polymorphism is associated with hepatic MCP-1 expression and severity of HCV-related liver disease. Gastroenterology 2003, 125, 1085–1093. [Google Scholar]
- Schwabe, R.F.; Bataller, R.; Brenner, D.A. Human hepatic stellate cells express CCR5 and RANTES to induce proliferation and migration. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G949–G958. [Google Scholar]
- Seki, E.; De Minicis, S.; Gwak, G.Y.; Kluwe, J.; Inokuchi, S.; Bursill, C.A.; Llovet, J.M.; Brenner, D.A.; Schwabe, R.F. CCR1 and CCR5 promote hepatic fibrosis in mice. J. Clin. Investig. 2009, 119, 1858–1870. [Google Scholar]
- Radaeva, S.; Sun, R.; Jaruga, B.; Nguyen, V.T.; Tian, Z.; Gao, B. Natural killer cells ameliorate liver fibrosis by killing activated stellate cells in NKG2D-dependent and tumor necrosis factor-related apoptosis-inducing ligand-dependent manners. Gastroenterology 2006, 130, 435–452. [Google Scholar]
- Kawarabayashi, N.; Seki, S.; Hatsuse, K.; Ohkawa, T.; Koike, Y.; Aihara, T.; Habu, Y.; Nakagawa, R.; Ami, K.; Hiraide, H.; et al. Decrease of CD56+ T cells and natural killer cells in cirrhotic livers with hepatitis C may be involved in their susceptibility to hepatocellular carcinoma. Hepatology 2000, 32, 962–969. [Google Scholar]
- Muhanna, N.; Doron, S.; Wald, O.; Horani, A.; Eid, A.; Pappo, O.; Friedman, S.L.; Safadi, R. Activation of hepatic stellate cells after phagocytosis of lymphocytes: A novel pathway of fibrogenesis. Hepatology 2008, 48, 963–977. [Google Scholar]
- Jeong, W.I.; Park, O.; Radaeva, S.; Gao, B. STAT1 inhibits liver fibrosis in mice by inhibiting stellate cell proliferation and stimulating NK cell cytotoxicity. Hepatology 2006, 44, 1441–1451. [Google Scholar]
- Wynn, T.A. Fibrotic disease and the T(H)1/T(H)2 paradigm. Nat. Rev. 2004, 4, 583–594. [Google Scholar]
- Chew, V.; Chen, J.; Lee, D.; Loh, E.; Lee, J.; Lim, K.H.; Weber, A.; Slankamenac, K.; Poon, R.T.; Yang, H.; et al. Chemokine-driven lymphocyte infiltration: an early intratumoural event determining long-term survival in resectable hepatocellular carcinoma. Gut 2012, 61, 427–438. [Google Scholar]
- Fu, J.; Xu, D.; Liu, Z.; Shi, M.; Zhao, P.; Fu, B.; Zhang, Z.; Yang, H.; Zhang, H.; Zhou, C.; et al. Increased regulatory T cells correlate with CD8 T-cell impairment and poor survival in hepatocellular carcinoma patients. Gastroenterology 2007, 132, 2328–2339. [Google Scholar]
- Hoechst, B.; Ormandy, L.A.; Ballmaier, M.; Lehner, F.; Kruger, C.; Manns, M.P.; Greten, T.F.; Korangy, F. A new population of myeloid-derived suppressor cells in hepatocellular carcinoma patients induces CD4+ CD25+ Foxp3+ T cells. Gastroenterology 2008, 135, 234–243. [Google Scholar]
- Ajuebor, M.N.; Hogaboam, C.M.; Le, T.; Proudfoot, A.E.; Swain, M.G. CCL3/MIP-1α is pro-inflammatory in murine T cell-mediated hepatitis by recruiting CCR1-expressing CD4+ T cells to the liver. Eur. J. Immunol. 2004, 34, 2907–2918. [Google Scholar]
- Ajuebor, M.N.; Wondimu, Z.; Hogaboam, C.M.; Le, T.; Proudfoot, A.E.; Swain, M.G. CCR5 deficiency drives enhanced natural killer cell trafficking to and activation within the liver in murine T cell-mediated hepatitis. Am. J. Pathol. 2007, 170, 1975–1988. [Google Scholar]
- Moreno, C.; Gustot, T.; Nicaise, C.; Quertinmont, E.; Nagy, N.; Parmentier, M.; Le Moine, O.; Deviere, J.; Louis, H. CCR5 deficiency exacerbates T-cell-mediated hepatitis in mice. Hepatology 2005, 42, 854–862. [Google Scholar]
- Mitchell, C.; Couton, D.; Couty, J.P.; Anson, M.; Crain, A.M.; Bizet, V.; Renia, L.; Pol, S.; Mallet, V.; Gilgenkrantz, H. Dual role of CCR2 in the constitution and the resolution of liver fibrosis in mice. Am. J. Pathol. 2009, 174, 1766–1775. [Google Scholar]
- Ishiwata, Y.; Yokochi, S.; Hashimoto, H.; Ninomiya, F.; Suzuki, T. Protection against concanavalin A-induced murine liver injury by the organic germanium compound propagermanium. Scand. J. Immunol. 1998, 48, 605–614. [Google Scholar]
- Yokochi, S.; Ishiwata, Y.; Hashimoto, H.; Ninomiya, F.; Suzuki, T. Hepatoprotective effect of propagermanium on Corynebacterium parvum and lipopolysaccharide-induced liver injury in mice. Scand. J. Immunol. 1998, 48, 183–191. [Google Scholar]
- Hogaboam, C.M.; Bone-Larson, C.L.; Steinhauser, M.L.; Matsukawa, A.; Gosling, J.; Boring, L.; Charo, I.F.; Simpson, K.J.; Lukacs, N.W.; Kunkel, S.L. Exaggerated hepatic injury due to acetaminophen challenge in mice lacking C-C chemokine receptor 2. Am. J. Pathol. 2000, 156, 1245–1252. [Google Scholar]
- Wiederholt, T.; von Westernhagen, M.; Zaldivar, M.M.; Berres, M.L.; Schmitz, P.; Hellerbrand, C.; Muller, T.; Berg, T.; Trautwein, C.; Wasmuth, H.E. Genetic variations of the chemokine scavenger receptor D6 are associated with liver inflammation in chronic hepatitis C. Hum. Immunol. 2008, 69, 861–866. [Google Scholar]
- Jaeschke, H.; Hasegawa, T. Role of neutrophils in acute inflammatory liver injury. Liver Int. 2006, 26, 912–919. [Google Scholar]
- Kuboki, S.; Shin, T.; Huber, N.; Eismann, T.; Galloway, E.; Schuster, R.; Blanchard, J.; Edwards, M.J.; Lentsch, A.B. Hepatocyte signaling through CXC chemokine receptor-2 is detrimental to liver recovery after ischemia/reperfusion in mice. Hepatology 2008, 48, 1213–1223. [Google Scholar]
- Bertini, R.; Allegretti, M.; Bizzarri, C.; Moriconi, A.; Locati, M.; Zampella, G.; Cervellera, M.N.; di Cioccio, V.; Cesta, M.C.; Galliera, E.; et al. Noncompetitive allosteric inhibitors of the inflammatory chemokine receptors CXCR1 and CXCR2: prevention of reperfusion injury. Proc. Natl. Acad. Sci. USA 2004, 101, 11791–11796. [Google Scholar]
- Erhardt, A.; Wegscheid, C.; Claass, B.; Carambia, A.; Herkel, J.; Mittrucker, H.W.; Panzer, U.; Tiegs, G. CXCR3 deficiency exacerbates liver disease and abrogates tolerance in a mouse model of immune-mediated hepatitis. J. Immunol. 2011, 186, 5284–5293. [Google Scholar]
- Zaldivar, M.M.; Berres, M.L.; Sahin, H.; Nellen, A.; Heinrichs, D.; Schmitz, P.; Gassler, N.; Streetz, K.L.; Trautwein, C.; Wasmuth, H.E. The chemokine receptor CXCR3 limits injury after acute toxic liver damage. Lab. Investig. 2012, 92, 724–734. [Google Scholar]
- Zhai, Y.; Shen, X.D.; Hancock, W.W.; Gao, F.; Qiao, B.; Lassman, C.; Belperio, J.A.; Strieter, R.M.; Busuttil, R.W.; Kupiec-Weglinski, J.W. CXCR3+ CD4+ T cells mediate innate immune function in the pathophysiology of liver ischemia/reperfusion injury. J. Immunol. 2006, 176, 6313–6322. [Google Scholar]
- Ajuebor, M.N.; Jin, Y.; Gremillion, G.L.; Strieter, R.M.; Chen, Q.; Adegboyega, P.A. γδT cells initiate acute inflammation and injury in adenovirus-infected liver via cytokine-chemokine cross talk. J. Virol. 2008, 82, 9564–9576. [Google Scholar]
- Wehr, A.; Baeck, C.; Heymann, F.; Niemietz, P.M.; Hammerich, L.; Martin, C.; Zimmermann, H.W.; Pack, O.; Gassler, N.; Hittatiya, K.; et al. Chemokine receptor CXCR6-dependent hepatic NK T Cell accumulation promotes inflammation and liver fibrosis. J. Immunol. 2013, 190, 5226–5236. [Google Scholar]
- Xu, H.B.; Gong, Y.P.; Cheng, J.; Chu, Y.W.; Xiong, S.D. CXCL16 participates in pathogenesis of immunological liver injury by regulating T lymphocyte infiltration in liver tissue. World J. Gastroenterol. 2005, 11, 4979–4985. [Google Scholar]
- Yang, S.J.; IglayReger, H.B.; Kadouh, H.C.; Bodary, P.F. Inhibition of the chemokine (C-C motif) ligand 2/chemokine (C-C motif) receptor 2 pathway attenuates hyperglycaemia and inflammation in a mouse model of hepatic steatosis and lipoatrophy. Diabetologia 2009, 52, 972–981. [Google Scholar]
- Tamura, Y.; Sugimoto, M.; Murayama, T.; Minami, M.; Nishikaze, Y.; Ariyasu, H.; Akamizu, T.; Kita, T.; Yokode, M.; Arai, H. C-C chemokine receptor 2 inhibitor improves diet-induced development of insulin resistance and hepatic steatosis in mice. J. Ather. Thromb. 2010, 17, 219–228. [Google Scholar]
- Barashi, N.; Weiss, I.D.; Wald, O.; Wald, H.; Beider, K.; Abraham, M.; Klein, S.; Goldenberg, D.; Axelrod, J.; Pikarsky, E.; et al. Inflammation-induced hepatocellular carcinoma is dependent on CCR5 in mice. Hepatology 2013, 58, 1021–1030. [Google Scholar]
- Karlmark, K.R.; Weiskirchen, R.; Zimmermann, H.W.; Gassler, N.; Ginhoux, F.; Weber, C.; Merad, M.; Luedde, T.; Trautwein, C.; Tacke, F. Hepatic recruitment of the inflammatory Gr1+ monocyte subset upon liver injury promotes hepatic fibrosis. Hepatology 2009, 50, 261–274. [Google Scholar]
- Seki, E.; de Minicis, S.; Inokuchi, S.; Taura, K.; Miyai, K.; van Rooijen, N.; Schwabe, R.F.; Brenner, D.A. CCR2 promotes hepatic fibrosis in mice. Hepatology 2009, 50, 185–197. [Google Scholar]
- Baeck, C.; Wehr, A.; Karlmark, K.R.; Heymann, F.; Vucur, M.; Gassler, N.; Huss, S.; Klussmann, S.; Eulberg, D.; Luedde, T.; et al. Pharmacological inhibition of the chemokine CCL2 (MCP-1) diminishes liver macrophage infiltration and steatohepatitis in chronic hepatic injury. Gut 2012, 61, 416–426. [Google Scholar]
- Stefanovic, L.; Brenner, D.A.; Stefanovic, B. Direct hepatotoxic effect of KC chemokine in the liver without infiltration of neutrophils. Exp. Biol. Med. 2005, 230, 573–586. [Google Scholar]
- Zimmermann, H.W.; Seidler, S.; Gassler, N.; Nattermann, J.; Luedde, T.; Trautwein, C.; Tacke, F. Interleukin-8 is activated in patients with chronic liver diseases and associated with hepatic macrophage accumulation in human liver fibrosis. PLoS One 2011, 6, e21381. [Google Scholar]
- Zaldivar, M.M.; Pauels, K.; von Hundelshausen, P.; Berres, M.L.; Schmitz, P.; Bornemann, J.; Kowalska, M.A.; Gassler, N.; Streetz, K.L.; Weiskirchen, R.; et al. CXC chemokine ligand 4 (Cxcl4) is a platelet-derived mediator of experimental liver fibrosis. Hepatology 2010, 51, 1345–1353. [Google Scholar]
- Hintermann, E.; Bayer, M.; Pfeilschifter, J.M.; Luster, A.D.; Christen, U. CXCL10 promotes liver fibrosis by prevention of NK cell mediated hepatic stellate cell inactivation. J. Autoimmun. 2010, 35, 424–435. [Google Scholar]
- Wasmuth, H.E.; Lammert, F.; Zaldivar, M.M.; Weiskirchen, R.; Hellerbrand, C.; Scholten, D.; Berres, M.L.; Zimmermann, H.; Streetz, K.L.; Tacke, F.; et al. Antifibrotic effects of CXCL9 and its receptor CXCR3 in livers of mice and humans. Gastroenterology 2009, 137, 309–319. [Google Scholar]
- Ding, B.S.; Cao, Z.; Lis, R.; Nolan, D.J.; Guo, P.; Simons, M.; Penfold, M.E.; Shido, K.; Rabbany, S.Y.; Rafii, S. Divergent angiocrine signals from vascular niche balance liver regeneration and fibrosis. Nature 2014, 505, 97–102. [Google Scholar]
- Yang, X.; Lu, P.; Fujii, C.; Nakamoto, Y.; Gao, J.L.; Kaneko, S.; Murphy, P.M.; Mukaida, N. Essential contribution of a chemokine CCL3 and its receptor CCR1 to hepatocellular carcinoma progression. Int. J. Cancer 2006, 118, 1869–1876. [Google Scholar]
- Kapanadze, T.; Gamrekelashvili, J.; Ma, C.; Chan, C.; Zhao, F.; Hewitt, S.; Zender, L.; Kapoor, V.; Felsher, D.W.; Manns, M.P.; et al. Regulation of accumulation and function of myeloid derived suppressor cells in different murine models of hepatocellular carcinoma. J Hepatol. 2013, 59, 1007–1013. [Google Scholar]
- Chen, Y.; Huang, Y.; Reiberger, T.; Duyverman, A.M.; Huang, P.; Samuel, R.; Hiddingh, L.; Roberge, S.; Koppel, C.; Lauwers, G.Y.; et al. Differential effects of sorafenib on liver versus tumor fibrosis mediated by SDF1α/CXCR4 axis and Gr-1+ myeloid cell infiltration in mice. Hepatology 2014, 2014. [Google Scholar] [CrossRef]
- Zheng, K.; Li, H.Y.; Su, X.L.; Wang, X.Y.; Tian, T.; Li, F.; Ren, G.S. Chemokine receptor CXCR7 regulates the invasion angiogenesis and tumor growth of human hepatocellular carcinoma cells. J. Exp. Clin. Cancer Res. 2010, 29, 31. [Google Scholar]
- Fatkenheuer, G.; Pozniak, A.L.; Johnson, M.A.; Plettenberg, A.; Staszewski, S.; Hoepelman, A.I.; Saag, M.S.; Goebel, F.D.; Rockstroh, J.K.; Dezube, B.J.; et al. Efficacy of short-term monotherapy with maraviroc a new CCR5 antagonist in patients infected with HIV-1. Nat. Med. 2005, 11, 1170–1172. [Google Scholar]
- Ramos, C.D.; Canetti, C.; Souto, J.T.; Silva, J.S.; Hogaboam, C.M.; Ferreira, S.H.; Cunha, F.Q. MIP-1α [CCL3] acting on the CCR1 receptor mediates neutrophil migration in immune inflammation via sequential release of TNF-α and LTB4. J. Leukoc. Biol. 2005, 78, 167–177. [Google Scholar]
- Bonecchi, R.; Polentarutti, N.; Luini, W.; Borsatti, A.; Bernasconi, S.; Locati, M.; Power, C.; Proudfoot, A.; Wells, T.N.; Mackay, C.; et al. Up-regulation of CCR1 and CCR3 and induction of chemotaxis to CC chemokines by IFN-γ in human neutrophils. J. Immunol. 1999, 162, 474–479. [Google Scholar]

{kind=link}
{kind=link}
| Chemokine receptor | Expression on | Ligands | Secretion by | Function | References |
|---|---|---|---|---|---|
| CCR1 | immature DCs, monocytes, memory T cells, NK cells | CCL3, CCL5 | infiltrating macrophages, KCs, DCs, activated HSCs, LSECs | recruitment of NK cells, immature DCs | [38,58–71] |
| CCR2 | monocytes, memory T cells, pDCs | CCL2, CCL7, CCL8, CCL13 | KCs, activated HSCs | recruitment of monocytes | [48,58,61,63,66,67,70, 72–77] |
| CCR4 | Th2 cells, Tregs | CCL17, CCL22 | DCs, macrophages, monocytes | recruitment of Th2 cells, Tregs | [38,78–83] |
| CCR5 | immature DCs, Th1 cells, NK cells, monocytes, pDCs, HSCs | CCL3, CCL4, CCL5, | infiltrating macrophages, KCs, DCs, activated HSCs, LSECs | recruitment of NK cells, immature DCs, effector T cells | [38,58–71] |
| CXCR1 | neutrophils, monocytes, mast cells, hepatocytes | CXCL6, CXCL7, CXCL8, | LSECs, KCs, activated hepatocytes, Th1 cells, Th17 cells | recruitment of neutrophils | [58,60,84–95] |
| CXCR2 | neutrophils, monocytes, mast cells | CXCL1-3, CXCL5, CXCL6, CXCL7, CXCL8 | LSECs, KCs, activated hepatocytes, Th1 cells, Th17 cells | recruitment of neutrophils | [58,60,84,87–95] |
| CXCR3 | CD8 T cells, Th1 cells, NK cells, NKT cells, Tregs, HSCs, mast cells | CXCL9, CXCL10, CXCL11, | hepatocytes, LSECs, HSCs, KCs, DCs, neutrophils | recruitment of NK, NKT and T cells | [58–61,96–101] |
| CXCR3-B | microvascular endothelial cells, activated T cells | CXCL4, CXCL9/10/11 | platelets | mediation of angiostatic effects | [46,102,103] |
| CXCR4 | widely expressed, HSCs | CXCL12 | activated HSCs | neoangiogenesisand HSC activation | [104–110] |
| CXCR6 | Th1 T cells, CD8 T cells, NKT cells, NK cells | CXCL16 | LSECs, hepatocytes, cholangiocytes, activated DCs | recruitment and retention of effector cells in the liver | [111–113] |
| Chemokine receptor | Ligands | Changes in HCV infection | References |
|---|---|---|---|
| CCR1 | CCL3, CCL5, CCL7, CCL8, CCL13-16, CCL23 | Increased intrahepatic CCL3 and 5 expression during chronic hepatitis C | [116] |
| Decreased proportion of CCR1+ CD8+ T cells in chronic hepatitis C patients | [117] | ||
| Increase of CCL5 expression by HCV core and NS5A in vitro, but decrease by NS3/4A | [118,119] | ||
| Decrease of the intrahepatic level of CCL3 by NS3/4A in vivo | [120] | ||
| CCR2 | CCL2, CCL7, CCL8, CCL13 | Increased expression of CCL2 and CCR2 in HCV-infected patients | [121,122] |
| Increase of CCL2 expression by HCV NS3/4A and NS5A | [123,124] | ||
| CCR4 | CCL17, CCL22 | Enhanced expression of CCL17 and CCL22 in chronic hepatitis C patients | [125,126] |
| Increased intrahepatic levels of CCL17 and CCL22 through liver-specific HCV NS3/4A expression | [125] | ||
| CCR5 | CCL3-5, CCL8 | High frequency of CCR5+ T cells during acute HCV infection | [127] |
| Increased CCL3, 4 and 5 expression during chronic hepatitis C | [116] | ||
| Decreased proportion of CCR5+ CD8+ T cells in chronic hepatitis C patients | [117] | ||
| Increase of CCL5 expression by HCV core and NS5A in vitro, but decrease by NS3/4A | [118,119] | ||
| Decrease of the intrahepatic level of CCL3 by NS3/4A in vivo | [120] | ||
| CXCR1/2 | CXCL6-8 (for CXCR1) and CXCL1-8 (for CXCR2) | Increased levels of CXCL8 in HCV-infected patients | [128,129] |
| Induction of CXCL8 expression by HCV core, NS4A, NS4B and NS5B, but decrease by NS3/4A | [119,130,131] | ||
| CXCR3 | CXCL4, CXCL9-11 | Increased intrahepatic CXCL9, CXCL10 and CXCL11 levels during chronic hepatitis C | [132] |
| Increased intrahepatic CXCL10 and CXCL11 levels in HCV-infected chimpanzees | [133] | ||
| High frequency of CXCR3+ CD8+ T cells during chronic HCV infection | [132,134] | ||
| Decrease of the intrahepatic levels of CXCL9 and 11 by NS3/4A in vivo | [125] | ||
| Reduction of CXCL10 expression by NS3/4A in vitro | [135] | ||
| CXCR6 | CXCL16 | Increase in CXCL16 levels during IFN therapy | [136] |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Brass, A.; Brenndörfer, E.D. The Role of Chemokines in Hepatitis C Virus-Mediated Liver Disease. Int. J. Mol. Sci. 2014, 15, 4747-4779. https://doi.org/10.3390/ijms15034747
Brass A, Brenndörfer ED. The Role of Chemokines in Hepatitis C Virus-Mediated Liver Disease. International Journal of Molecular Sciences. 2014; 15(3):4747-4779. https://doi.org/10.3390/ijms15034747
Chicago/Turabian StyleBrass, Anette, and Erwin Daniel Brenndörfer. 2014. "The Role of Chemokines in Hepatitis C Virus-Mediated Liver Disease" International Journal of Molecular Sciences 15, no. 3: 4747-4779. https://doi.org/10.3390/ijms15034747