Genetics of Oxidative Stress in Obesity


Abstract
:1. Introduction
2. Methodology
3. Oxidative Stress in Obesity
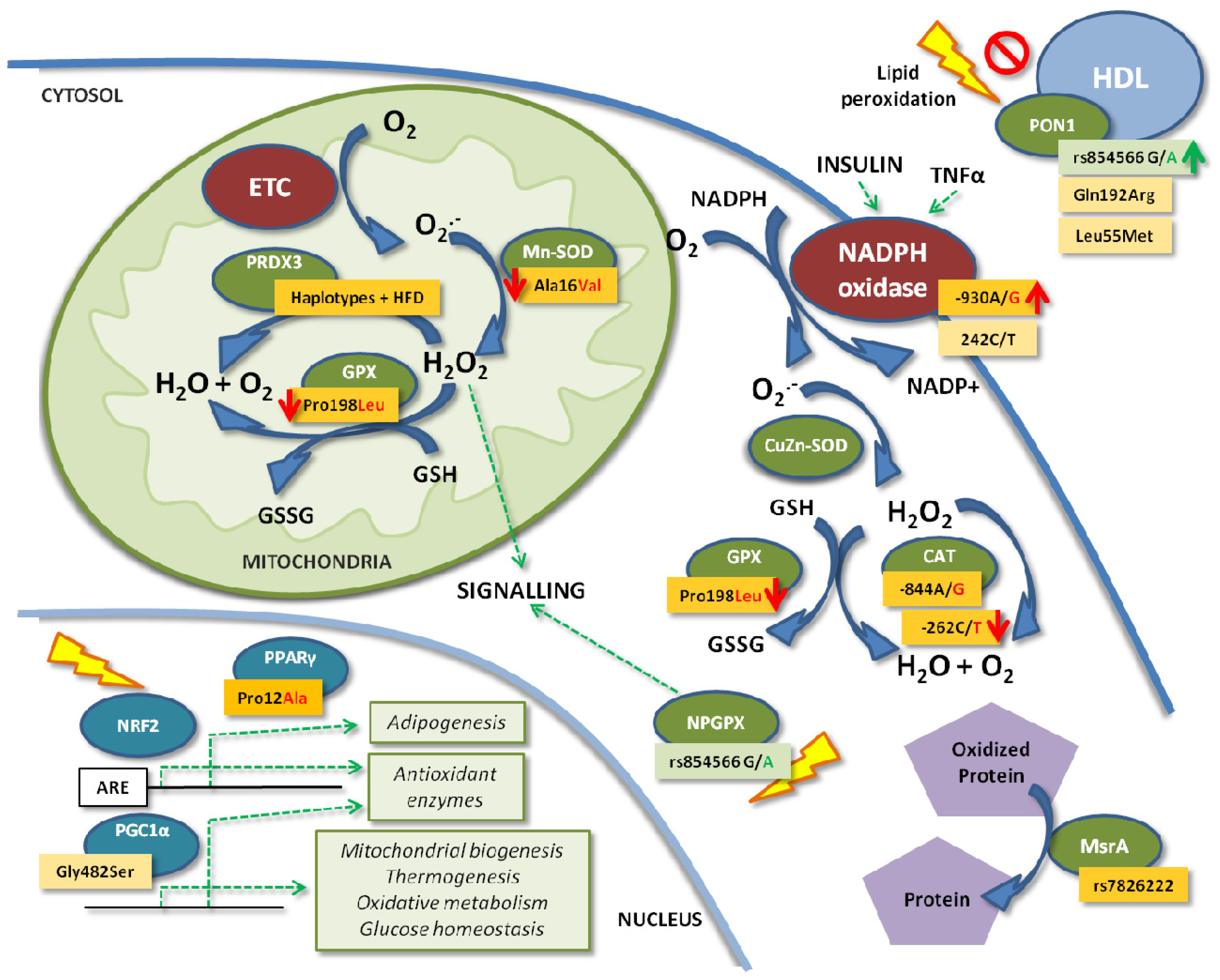
4. Enzymatic Antioxidant Defense Genes
4.1. Glutathione Peroxidases
4.2. Catalase
4.3. Paraoxonases
4.4. Peroxiredoxins
4.5. Superoxide Dismutases
4.6. MsrA
5. The ROS Producer: NADPH Oxidase
6. ROS Response Mechanisms: Transcription Factors
6.1. PPARγ
6.2. PGC1α
6.3. NRF2
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Shamseddeen, H.; Getty, J.Z.; Hamdallah, I.N.; Ali, M.R. Epidemiology and economic impact of obesity and type 2 diabetes. Surg. Clin. N. Am 2011, 91, 1163–1172. [Google Scholar]
- Gough, D.R.; Cotter, T.G. Hydrogen peroxide: a Jekyll and Hyde signalling molecule. Cell Death Dis 2011. [Google Scholar] [CrossRef]
- Da Costa, L.A.; Badawi, A.; El–Sohemy, A. Nutrigenetics and modulation of oxidative stress. Ann. Nutr. Metab 2012, 60, 27–36. [Google Scholar]
- Chang, Y.C.; Yu, Y.H.; Shew, J.Y.; Lee, W.J.; Hwang, J.J.; Chen, Y.H.; Chen, Y.R.; Wei, P.C.; Chuang, L.M.; Lee, W.H. Deficiency of NPGPx, an oxidative stress sensor, leads to obesity in mice and human. EMBO Mol. Med 2013, 5, 1165–1179. [Google Scholar]
- Balistreri, C.R.; Caruso, C.; Candore, G. The role of adipose tissue and adipokines in obesity-related inflammatory diseases. Mediators Inflamm 2010. [Google Scholar] [CrossRef]
- Trayhurn, P.; Wood, I.S. Adipokines: Inflammation and the pleiotropic role of white adipose tissue. Br. J. Nutr 2004, 92, 347–355. [Google Scholar]
- Bondia-Pons, I.; Ryan, L.; Martinez, J.A. Oxidative stress and inflammation interactions in human obesity. J. Physiol. Biochem 2012, 68, 701–711. [Google Scholar]
- Trayhurn, P. Hypoxia and adipose tissue function and dysfunction in obesity. Physiol. Rev 2013, 93, 1–21. [Google Scholar]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Invest 2004, 114, 1752–1761. [Google Scholar]
- Anderson, E.J.; Lustig, M.E.; Boyle, K.E.; Woodlief, T.L.; Kane, D.A.; Lin, C.T.; Price, J.W., 3rd; Kang, L.; Rabinovitch, P.S.; Szeto, H.H.; et al. Mitochondrial H2O2 emission and cellular redox state link excess fat intake to insulin resistance in both rodents and humans. J. Clin. Invest 2009, 119, 573–581. [Google Scholar]
- Bajnok, L.; Seres, I.; Varga, Z.; Jeges, S.; Peti, A.; Karanyi, Z.; Juhasz, A.; Csongradi, E.; Mezosi, E.; Nagy, E.V.; et al. Relationship of endogenous hyperleptinemia to serum paraoxonase 1, cholesteryl ester transfer protein, and lecithin cholesterol acyltransferase in obese individuals. Metabolism 2007, 56, 1542–1549. [Google Scholar]
- Keaney, J.F., Jr.; Larson, M.G.; Vasan, R.S.; Wilson, P.W.; Lipinska, I.; Corey, D.; Massaro, J.M.; Sutherland, P.; Vita, J.A.; Benjamin, E.J. Framingham study: Obesity and systemic oxidative stress: Clinical correlates of oxidative stress in the Framingham Study. Arterioscler. Thromb. Vasc. Biol 2003, 23, 434–439. [Google Scholar]
- Codoñer-Franch, P.; Tavárez-Alonso, S.; Murria-Estal, R.; Megías-Vericat, J.; Tortajada-Girbés, M.; Alonso-Iglesias, E. Nitric oxide production is increased in severely obese children and related to markers of oxidative stress and inflammation. Atherosclerosis 2011, 2215, 475–480. [Google Scholar]
- Ozgen, I.T.; Tascilar, M.E.; Bilir, P.; Boyraz, M.; Guncikan, M.N.; Akay, C.; Dundaroz, R. Oxidative stress in obese children and its relation with insulin resistance. J. Pediatr. Endocrinol. Metab 2012, 25, 261–266. [Google Scholar]
- Rupérez, A.I.; Olza, J.; Gil-Campos, M.; Leis, R.; Mesa, M.D.; Tojo, R.; Cañete, R.; Gil, A.; Aguilera, C.M. Are Catalase −844A/G Polymorphism and Activity Associated with Childhood Obesity? Antioxid. Redox Signal 2013, 19, 1970–1975. [Google Scholar]
- D’Archivio, M.; Annuzzi, G.; Varì, R.; Filesi, C.; Giacco, R.; Scazzocchio, B.; Santangelo, C.; Giovannini, C.; Rivellese, A.A.; Masella, R. Predominant role of obesity/insulin resistance in oxidative stress development. Eur. J. Clin. Invest 2012, 42, 70–78. [Google Scholar]
- Block, G.; Dietrich, M.; Norkus, E.P.; Morrow, J.D.; Hudes, M.; Caan, B.; Packer, L. Factors associated with oxidative stress in human populations. Am. J. Epidemiol 2002, 156, 274–285. [Google Scholar]
- Meigs, J.B.; Larson, M.G.; Fox, C.S.; Keaney, J.F., Jr.; Vasan, R.S.; Benjamin, E.J. Association of oxidative stress, insulin resistance, and diabetes risk phenotypes: The Framingham Offspring Study. Diabetes Care 2007, 30, 2529–2535. [Google Scholar]
- Karbownik-Lewinska, M.; Szosland, J.; Kokoszko-Bilska, A.; Stępniak, J.; Zasada, K.; Gesing, A.; Lewinski, A. Direct contribution of obesity to oxidative damage to macromolecules. Neuro Endocrinol. Lett 2012, 33, 453–461. [Google Scholar]
- Sankhla, M.; Sharma, T.K.; Mathur, K.; Rathor, J.S.; Butolia, V.; Gadhok, A.K.; Vardey, S.K.; Sinha, M.; Kaushik, G.G. Relationship of oxidative stress with obesity and its role in obesity induced metabolic syndrome. Clin. Lab 2012, 58, 385–392. [Google Scholar]
- Holvoet, P.; Lee, D.H.; Steffes, M.; Gross, M.; Jacobs, D.R., Jr. Association between circulating oxidized low-density lipoprotein and incidence of the metabolic syndrome. JAMA 2008, 299, 2287–2293. [Google Scholar]
- Van Guilder, G.P.; Hoetzer, G.L.; Greiner, J.J.; Stauffer, B.L.; Desouza, C.A. Influence of metabolic syndrome on biomarkers of oxidative stress and inflammation in obese adults. Obesity (Silver Spring) 2006, 14, 2127–2231. [Google Scholar]
- Aslan, M.; Horoz, M.; Sabuncu, T.; Celik, H.; Selek, S. Serum paraoxonase enzyme activity and oxidative stress in obese subjects. Pol. Arch. Med. Wewn 2011, 121, 181–185. [Google Scholar]
- Ferretti, G.; Bacchetti, T.; Moroni, C.; Savino, S.; Liuzzi, A.; Balzola, F.; Bicchiega, V. Paraoxonase activity in high-density lipoproteins: A comparison between healthy and obese females. J. Clin. Endocrinol. Metab 2005, 90, 1728–1733. [Google Scholar]
- Martínez-Salazar, M.F.; Almenares-López, D.; García-Jiménez, S.; Sánchez-Alemán, M.A.; Juantorena-Ugás, A.; Ríos, C.; Monroy-Noyola, A. Relationship between the paraoxonase (PON1) L55M and Q192R polymorphisms and obesity in a Mexican population: A pilot study. Genes Nutr 2011, 6, 361–368. [Google Scholar]
- Veiga, L.; Silva-Nunes, J.; Melão, A.; Oliveira, A.; Duarte, L.; Brito, M. Q192R polymorphism of the paraoxonase-1 gene as a risk factor for obesity in Portuguese women. Eur. J. Endocrinol 2011, 164, 213–218. [Google Scholar]
- Tabur, S.; Torun, A.N.; Sabuncu, T.; Turan, M.N.; Celik, H.; Ocak, A.R.; Aksoy, N. Non-diabetic metabolic syndrome and obesity do not affect serum paraoxonase and arylesterase activities but do affect oxidative stress and inflammation. Eur. J. Endocrinol 2010, 162, 535–541. [Google Scholar]
- Krzystek-Korpacka, M.; Patryn, E.; Boehm, D.; Berdowska, I.; Zielinski, B.; Noczynska, A. Advanced oxidation protein products (AOPPs) in juvenile overweight and obesity prior to and following weight reduction. Clin. Biochem 2008, 41, 943–949. [Google Scholar]
- Warolin, J.; Coenen, K.R.; Kantor, J.L.; Whitaker, L.E.; Wang, L.; Acra, S.A.; Roberts, L.J., 2nd; Buchowski, M.S. The relationship of oxidative stress, adiposity and metabolic risk factors in healthy Black and White American youth. Pediatr. Obes 2014, 9, 43–52. [Google Scholar]
- Oliver, S.R.; Rosa, J.S.; Milne, G.L.; Pontello, A.M.; Borntrager, H.L.; Heydari, S.; Galassetti, P.R. Increased oxidative stress and altered substrate metabolism in obese children. Int. J. Pediatr. Obes 2010, 5, 436–444. [Google Scholar]
- Araki, S.; Dobashi, K.; Yamamoto, Y.; Asayama, K.; Kusuhara, K. Increased plasma isoprostane is associated with visceral fat, high molecular weight adiponectin, and metabolic complications in obese children. Eur. J. Pediatr 2010, 169, 965–970. [Google Scholar]
- Codoñer-Franch, P.; Tavárez-Alonso, S.; Porcar-Almela, M.; Navarro-Solera, M.; Arilla-Codoñer, A.; Alonso-Iglesias, E. Plasma resistin levels are associated with homocysteine, endothelial activation, and nitrosative stress in obese youths. Clin. Biochem 2014, 47, 44–48. [Google Scholar]
- Kelly, A.S.; Jacobs, D.R., Jr.; Sinaiko, A.R.; Moran, A.; Steffen, L.M.; Steinberger, J. Relation of circulating oxidized LDL to obesity and insulin resistance in children. Pediatr. Diabetes 2010, 11, 552–555. [Google Scholar]
- Koncsos, P.; Seres, I.; Harangi, M.; Illyés, I.; Józsa, L.; Gönczi, F.; Bajnok, L.; Paragh, G. Human paraoxonase-1 activity in childhood obesity and its relation to leptin and adiponectin levels. Pediatr. Res 2010, 67, 309–313. [Google Scholar]
- Krzystek-Korpacka, M.; Patryn, E.; Hotowy, K.; Czapińska, E.; Majda, J.; Kustrzeba-Wójcicka, I.; Noczyńska, A.; Gamian, A. Paraoxonase (PON)-1 activity in overweight and obese children and adolescents: Association with obesity-related inflammation and oxidative stress. Adv. Clin. Exp. Med 2013, 22, 229–236. [Google Scholar]
- Ferré, N.; Feliu, A.; García-Heredia, A.; Marsillach, J.; París, N.; Zaragoza-Jordana, M.; Mackness, B.; Mackness, M.; Escribano, J.; Closa-Monasterolo, R.; et al. Impaired paraoxonase-1 status in obese children. Relationships with insulin resistance and metabolic syndrome. Clin. Biochem 2013, 46, 1830–1836. [Google Scholar]
- Rupérez, A.I.; López-Guarnido, O.; Gil, F.; Olza, J.; Gil-Campos, M.; Leis, R.; Tojo, R.; Cañete, R.; Gil, A.; Aguilera, C.M. Paraoxonase 1 activities and genetic variation in childhood obesity. Br. J. Nutr 2013, 110, 1639–1647. [Google Scholar]
- Pirgon, Ö.; Bilgin, H.; Çekmez, F.; Kurku, H.; Dündar, B.N. Association between insulin resistance and oxidative stress parameters in obese adolescents with non-alcoholic fatty liver disease. J. Clin. Res. Pediatr. Endocrinol 2013, 5, 33–39. [Google Scholar]
- Krieger-Brauer, H.I.; Medda, P.K.; Kather, H. The stimulus-sensitive H2O2-generating system present in human fat-cell plasma membranes is multireceptor-linked and under antagonistic control by hormones and cytokines. Biochem. J 1995, 307, 543–548. [Google Scholar]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol 2006, 194, 7–15. [Google Scholar]
- Houstis, N.; Rosen, E.D.; Lander, E.S. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature 2006, 440, 944–948. [Google Scholar]
- Styskal, J.; Nwagwu, F.A.; Watkins, Y.N.; Liang, H.; Richardson, A.; Musi, N.; Salmon, A.B. Methionine sulfoxide reductase A affects insulin resistance by protecting insulin receptor function. Free Radic. Biol. Med 2013, 56, 123–132. [Google Scholar]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory mechanisms in obesity. Annu. Rev. Immunol 2011, 29, 415–545. [Google Scholar]
- Huh, J.Y.; Kim, Y.; Jeong, J.; Park, J.; Kim, I.; Huh, K.H.; Kim, Y.S.; Woo, H.A.; Rhee, S.G.; Lee, K.J.; et al. Peroxiredoxin 3 is a key molecule regulating adipocyte oxidative stress, mitochondrial biogenesis, and adipokine expression. Antioxid. Redox Signal 2012, 16, 229–243. [Google Scholar]
- Luo, W.; Cao, J.; Li, J.; He, W. Adipose tissue-specific PPARγ deficiency increases resistance to oxidative stress. Exp. Gerontol 2008, 43, 154–163. [Google Scholar]
- Itoh, H.; Doi, K.; Tanaka, T.; Fukunaga, Y.; Hosoda, K.; Inoue, G.; Nishimura, H.; Yoshimasa, Y.; Yamori, Y.; Nakao, K. Hypertension and insulin resistance: Role of peroxisome proliferator-activated receptor gamma. Clin. Exp. Pharmacol. Physiol 1999, 26, 558–560. [Google Scholar]
- Austin, S.; St-Pierre, J. PGC1a and mitochondrial metabolism - emerging concepts and relevance in ageing and neurodegenerative disorders. J. Cell Sci 2012, 125, 4963–4971. [Google Scholar]
- Valle, I.; Alvarez-Barrientos, A.; Arza, E.; Lamas, S.; Monsalve, M. PGC-1a regulates the mitochondrial antioxidant defense system in vascular endothelial cells. Cardiovasc. Res 2005, 66, 562–573. [Google Scholar]
- Margis, R.; Dunand, C.; Teixeira, F.K.; Margis-Pinheiro, M. Glutathione peroxidase family - an evolutionary overview. FEBS J 2008, 275, 3959–3970. [Google Scholar]
- Yoshimura, S.; Watanabe, K.; Suemizu, H.; Onozawa, T.; Mizoguchi, J.; Tsuda, K.; Hatta, H.; Moriuchi, T. Tissue specific expression of the plasma glutathione peroxidase gene in rat kidney. J. Biochem 1991, 109, 918–923. [Google Scholar]
- Toppo, S.; Vanin, S.; Bosello, V.; Tosatto, S.C. Evolutionary and structural insights into the multifaceted glutathione peroxidase (Gpx) superfamily. Antioxid. Redox. Signal 2008, 10, 1501–1514. [Google Scholar]
- Wei, P.C.; Hsieh, Y.H.; Su, M.I.; Jiang, X.; Hsu, P.H.; Lo, W.T.; Weng, J.Y.; Jeng, Y.M.; Wang, J.M.; Chen, P.L.; et al. Loss of the oxidative stress sensor NPGPx compromises GRP78 chaperone activity and induces systemic disease. Mol. Cell 2012, 48, 747–759. [Google Scholar]
- Asayama, K.; Nakane, T.; Dobashi, K.; Kodera, K.; Hayashibe, H.; Uchida, N.; Nakazawa, S. Effect of obesity and troglitazone on expression of two glutathione peroxidases: Cellular and extracellular types in serum, kidney and adipose tissue. Free Radic. Res 2001, 34, 337–347. [Google Scholar]
- Lijnen, H.R.; Van Hul, M.; Hemmeryckx, B. Caloric restriction improves coagulation and inflammation profile in obese mice. Thromb. Res 2012, 129, 74–79. [Google Scholar]
- Loh, K.; Deng, H.; Fukushima, A.; Cai, X.; Boivin, B.; Galic, S.; Bruce, C.; Shields, B.J.; Skiba, B.; Ooms, L.M.; et al. Reactive oxygen species enhance insulin sensitivity. Cell Metab 2009, 10, 260–272. [Google Scholar]
- Merry, T.L.; Tran, M.; Stathopoulos, M.; Wiede, F.; Fam, B.C.; Dood, G.T.; Clarke, I.; Watt, M.J.; Andrikopoulos, S.; Tiganis, T. High fat fed obese Gpx1-deficient mice exhibit defective insulin secretion but protection from hepatic steatosis and liver damage. Antioxid. Redox Signal 2013. [Google Scholar] [CrossRef]
- De Haan, J.B.; Witting, P.K.; Stefanovic, N.; Pete, J.; Daskalakis, M.; Kola, I.; Stocker, R.; Smolich, J.J. Lack of the antioxidant glutathione peroxidase-1 does not increase atherosclerosis in C57BL/J6 mice fed a high-fat diet. J. Lipid Res 2006, 47, 1157–1167. [Google Scholar]
- Lee, Y.S.; Kim, A.Y.; Choi, J.W.; Kim, M.; Yasue, S.; Son, H.J.; Masuzaki, H.; Park, K.S.; Kim, J.B. Dysregulation of adipose glutathione peroxidase 3 in obesity contributes to local and systemic oxidative stress. Mol. Endocrinol 2008, 22, 2176–2189. [Google Scholar]
- Lundholm, L.; Putnik, M.; Otsuki, M.; Andersson, S.; Ohlsson, C.; Gustafsson, J.A.; Dahlman-Wright, K. Effects of estrogen on gene expression profiles in mouse hypothalamus and white adipose tissue: target genes include glutathione peroxidase 3 and cell death-inducing DNA fragmentation factor, alpha-subunit-like effector A. J. Endocrinol 2008, 196, 547–557. [Google Scholar]
- Kuzuya, M.; Ando, F.; Iguchi, A.; Shimokata, H. Glutathione peroxidase 1 Pro198Leu variant contributes to the metabolic syndrome in men in a large Japanese cohort. Am. J. Clin. Nutr 2008, 87, 1939–1944. [Google Scholar]
- Cominetti, C.; de Bortoli, M.C.; Purgatto, E.; Ong, T.P.; Moreno, F.S.; Garrido, A.B., Jr.; Cozzolino, S.M. Associations between glutathione peroxidase-1 Pro198Leu polymorphism, selenium status, and DNA damage levels in obese women after consumption of Brazil nuts. Nutrition 2011, 27, 891–896. [Google Scholar]
- Shuvalova, Y.A.; Kaminnyi, A.I.; Meshkov, A.N.; Kukharchuk, V.V. Pro198Leu polymorphism of GPx-1 gene and activity of erythrocytic glutathione peroxidase and lipid peroxidation products. Bull. Exp. Biol. Med 2010, 149, 743–745. [Google Scholar]
- Hamanishi, T.; Furuta, H.; Kato, H.; Doi, A.; Tamai, M.; Shimomura, H.; Sakagashira, S.; Nishi, M.; Sasaki, H.; Sanke, T.; et al. Functional variants in the glutathione peroxidase-1 (GPx-1) gene are associated with increased intima-media thickness of carotid arteries and risk of macrovascular diseases in japanese type 2 diabetic patients. Diabetes 2004, 53, 2455–2460. [Google Scholar]
- Rindler, P.M.; Plafker, S.M.; Szweda, L.I.; Kinter, M. High dietary fat selectively increases catalase expression within cardiac mitochondria. J. Biol. Chem 2013, 288, 1979–1990. [Google Scholar]
- Shin, M.J.; Park, E. Contribution of insulin resistance to reduced antioxidant enzymes and vitamins in nonobese Korean children. Clin. Chim. Acta 2006, 365, 200–205. [Google Scholar]
- Wang, Z.; Li, Y.; Wang, B.; He, Y.; Wang, Y.; Xi, H.; Li, Y.; Wang, Y.; Zhu, D.; Jin, J.; et al. A haplotype of the catalase gene confers an increased risk of essential hypertension in Chinese Han. Hum. Mutat 2010, 31, 272–278. [Google Scholar]
- Adkins, S.; Gan, K.N.; Mody, M.; La Du, B.N. Molecular basis for the polymorphic forms of human serum paraoxonase/arylesterase: Glutamine or arginine at position 191, for the respective A or B allozymes. Am. J. Hum. Genet 1993, 52, 598–608. [Google Scholar]
- Baráth, A.; Németh, I.; Karg, E.; Endreffy, E.; Bereczki, C.; Gellén, B.; Haszon, I.; Túri, S. Roles of paraoxonase and oxidative stress in adolescents with uraemic, essential or obesity-induced hypertension. Kidney Blood Press. Res 2006, 29, 144–151. [Google Scholar]
- Hiroi, M.; Nagahara, Y.; Miyauchi, R.; Misaki, Y.; Goda, T.; Kasezawa, N.; Sasaki, S.; Yamakawa-Kobayashi, K. The combination of genetic variations in the PRDX3 gene and dietary fat intake contribute to obesity risk. Obesity (Silver Spring) 2011, 19, 882–887. [Google Scholar]
- Lindgren, C.M.; Heid, I.M.; Randall, J.C.; Lamina, C.; Steinthorsdottir, V.; Qi, L.; Speliotes, E.K.; Thorleifsson, G.; Willer, C.J.; Herrera, B.M.; et al. Genome-wide association scan meta-analysis identifies three Loci influencing adiposity and fat distribution. PLoS Genet 2009, 5, e1000508. [Google Scholar]
- Bille, D.S.; Banasik, K.; Justesen, J.M.; Sandholt, C.H.; Sandbæk, A.; Lauritzen, T.; Jørgensen, T.; Witte, D.R.; Holm, J.C.; Hansen, T.; et al. Implications of central obesity-related variants in LYPLAL1, NRXN3, MSRA, and TFAP2B on quantitative metabolic traits in adult Danes. PLoS One 2011, 6, e20640. [Google Scholar]
- Graff, M.; Fernández-Rhodes, L.; Liu, S.; Carlson, C.; Wassertheil-Smoller, S.; Neuhouser, M.; Reiner, A.; Kooperberg, C.; Rampersaud, E.; Manson, J.E.; et al. Generalization of adiposity genetic loci to US Hispanic women. Nutr. Diabetes 2013, 3, e85. [Google Scholar]
- Yeung, E.; Qi, L.; Hu, F.B.; Zhang, C. Novel abdominal adiposity genes and the risk of type 2 diabetes: findings from two prospective cohorts. Int. J. Mol. Epidemiol. Genet 2011, 2, 138–144. [Google Scholar]
- Delahanty, R.J.; Beeghly-Fadiel, A.; Xiang, Y.B.; Long, J.; Cai, Q.; Wen, W.; Xu, W.H.; Cai, H.; He, J.; Gao, Y.T.; Zheng, W.; Shu, X.O. Association of obesity-related genetic variants with endometrial cancer risk: A report from the Shanghai Endometrial Cancer Genetics Study. Am. J. Epidemiol 2011, 174, 1115–1126. [Google Scholar]
- Hotta, K.; Kitamoto, T.; Kitamoto, A.; Mizusawa, S.; Matsuo, T.; Nakata, Y.; Kamohara, S.; Miyatake, N.; Kotani, K.; Komatsu, R.; et al. Association of variations in the FTO, SCG3 and MTMR9 genes with metabolic syndrome in a Japanese population. J. Hum. Genet 2011, 56, 647–651. [Google Scholar] [Green Version]
- Hotta, K.; Nakamura, M.; Nakamura, T.; Matsuo, T.; Nakata, Y.; Kamohara, S.; Miyatake, N.; Kotani, K.; Komatsu, R.; Itoh, N.; et al. Polymorphisms in NRXN3, TFAP2B, MSRA, LYPLAL1, FTO and MC4R and their effect on visceral fat area in the Japanese population. J. Hum. Genet 2010, 55, 738–742. [Google Scholar]
- Scherag, A.; Dina, C.; Hinney, A.; Vatin, V.; Scherag, S.; Vogel, C.I.; Müller, T.D.; Grallert, H.; Wichmann, H.E.; Balkau, B.; et al. Two new Loci for body-weight regulation identified in a joint analysis of genome-wide association studies for early-onset extreme obesity in French and german study groups. PLoS Genet 2010, 6, e1000916. [Google Scholar]
- Dorajoo, R.; Blakemore, A.I.; Sim, X.; Ong, R.T.; Ng, D.P.; Seielstad, M.; Wong, T.Y.; Saw, S.M.; Froguel, P.; Liu, J.; et al. Replication of 13 obesity loci among Singaporean Chinese, Malay and Asian-Indian populations. Int. J. Obes. (Lond) 2012, 36, 159–163. [Google Scholar]
- Scherag, A.; Kleber, M.; Boes, T.; Kolbe, A.L.; Ruth, A.; Grallert, H.; Illig, T.; Heid, I.M.; Toschke, A.M.; Grau, K.; et al. SDCCAG8 obesity alleles and reduced weight loss after a lifestyle intervention in overweight children and adolescents. Obesity (Silver Spring) 2012, 20, 466–470. [Google Scholar]
- Sutton, A.; Khoury, H.; Prip-Buus, C.; Cepanec, C.; Pessayre, D.; Degoul, F. The Ala16Val genetic dimorphism modulates the import of human manganese superoxide dismutase into rat liver mitochondria. Pharmacogenetics 2003, 13, 145–157. [Google Scholar]
- Montano, M.A.; Barrio Lera, J.P.; Gottlieb, M.G.; Schwanke, C.H.; da Rocha, M.I.; Manica-Cattani, M.F.; dos Santos, G.F.; da Cruz, I.B. Association between manganese superoxide dismutase (MnSOD) gene polymorphism and elderly obesity. Mol. Cell. Biochem 2009, 328, 33–40. [Google Scholar]
- Montano, M.A.; da Cruz, I.B.; Duarte, M.M.; da Krewer, C.C.; da Rocha, M.I.; Mânica-Cattani, M.F.; Soares, F.A.; Rosa, G.; Maris, A.F.; Battiston, F.G.; et al. Inflammatory cytokines in vitro production are associated with Ala16Val superoxide dismutase gene polymorphism of peripheral blood mononuclear cells. Cytokine 2012, 60, 30–33. [Google Scholar]
- Caple, F.; Williams, E.A.; Spiers, A.; Tyson, J.; Burtle, B.; Daly, A.K.; Mathers, J.C.; Hesketh, J.E. Inter- individual variation in DNA damage and base excision repair in young, healthy nonsmokers: Effects of dietary supplementation and genotype. Br. J. Nutr 2010, 103, 1585–1593. [Google Scholar]
- San José, G.; Moreno, M.U.; Olivan, S.; Beloqui, O.; Fortuno, A.; Díez, J.; Zalba, G. Functional effect of the p22phox −930A/G polymorphism on p22phox expression and NADPH oxidase activity in hypertension. Hypertension 2004, 44, 163–169. [Google Scholar]
- Ochoa, M.C.; Razquin, C.; Zalba, G.; Martínez-González, M.A.; Martínez, J.A.; Marti, A. G allele of the -930A>G polymorphism of the CYBA gene is associated with insulin resistance in obese subjects. J. Physiol. Biochem 2008, 64, 127–133. [Google Scholar]
- Hayaishi-Okano, R.; Yamasaki, Y.; Kajimoto, Y.; Sakamoto, K.; Ohtoshi, K.; Katakami, N.; Kawamori, D.; Miyatsuka, T.; Hatazaki, M.; Hazama, Y.; et al. Association of NAD(P)H oxidase p22 phox gene variation with advanced carotid atherosclerosis in Japanese type 2 diabetes. Diabetes Care 2003, 26, 458–463. [Google Scholar]
- Schreiber, R.; Ferreira-Sae, M.C.; Tucunduva, A.C.; Mill, J.G.; Costa, F.O.; Krieger, J.E.; Franchini, K.G.; Pereira, A.C.; Nadruz, W., Jr. CYBA C242T polymorphism is associated with obesity and diabetes mellitus in Brazilian hypertensive patients. Diabet. Med 2012, 29, e55–e61. [Google Scholar]
- Kim, C.; Zheng, T.; Lan, Q.; Chen, Y.; Foss, F.; Chen, X.; Holford, T.; Leaderer, B.; Boyle, P.; Chanock, S.J.; et al. Genetic polymorphisms in oxidative stress pathway genes and modification of BMI and risk of non-Hodgkin lymphoma. Cancer Epidemiol. Biomark. Prev 2012, 21, 866–868. [Google Scholar]
- Deeb, S.S.; Fajas, L.; Nemoto, M.; Pihlajamäki, J.; Mykkänen, L.; Kuusisto, J.; Laakso, M.; Fujimoto, W.; Auwerx, J. A Pro12Ala substitution in PPARgamma2 associated with decreased receptor activity, lower body mass index and improved insulin sensitivity. Nat. Genet 1998, 20, 284–287. [Google Scholar]
- Galbete, C.; Toledo, E.; Martínez-González, M.A.; Martínez, J.A.; Guillén-Grima, F.; Marti, A. Pro12Ala variant of the PPARG2 gene increases body mass index: An updated meta-analysis encompassing 49,092 subjects. Obesity (Silver Spring) 2013. [Google Scholar] [CrossRef]
- Fanelli, M.; Filippi, E.; Sentinelli, F.; Romeo, S.; Fallarino, M.; Buzzetti, R.; Leonetti, F.; Baroni, M.G. The Gly482Ser missense mutation of the peroxisome proliferator-activated receptor gamma coactivator-1 alpha (PGC-1 alpha) gene associates with reduced insulin sensitivity in normal and glucose-intolerant obese subjects. Dis. Markers 2005, 21, 175–180. [Google Scholar]
- Esterbauer, H.; Oberkofler, H.; Linnemayr, V.; Iglseder, B.; Hedegger, M.; Wolfsgruber, P.; Paulweber, B.; Fastner, G.; Krempler, F.; Patsch, W. Peroxisome proliferator-activated receptor-gamma coactivator-1 gene locus: associations with obesity indices in middle-aged women. Diabetes 2002, 51, 1281–1286. [Google Scholar]
- Weng, S.W.; Lin, T.K.; Wang, P.W.; Chen, I.Y.; Lee, H.C.; Chen, S.D.; Chuang, Y.C.; Liou, C.W. Gly482Ser polymorphism in the peroxisome proliferator-activated receptor gamma coactivator-1alpha gene is associated with oxidative stress and abdominal obesity. Metabolism 2010, 59, 581–586. [Google Scholar]
- Okauchi, Y.; Iwahashi, H.; Okita, K.; Yuan, M.; Matsuda, M.; Tanaka, T.; Miyagawa, J.; Funahashi, T.; Horikawa, Y.; Shimomura, I.; et al. PGC-1alpha Gly482Ser polymorphism is associated with the plasma adiponectin level in type 2 diabetic men. Endocr. J 2008, 55, 991–997. [Google Scholar]
- Goyenechea, E.; Crujeiras, A.B.; Abete, I.; Parra, D.; Martínez, J.A. Enhanced short-term improvement of insulin response to a low-caloric diet in obese carriers the Gly482Ser variant of the PGC-1alpha gene. Diabetes Res. Clin. Pract 2008, 82, 190–196. [Google Scholar]
- Ahn, J.; Gammon, M.D.; Santella, R.M.; Gaudet, M.M.; Britton, J.A.; Teitelbaum, S.L.; Terry, M.B.; Nowell, S.; Davis, W.; Garza, C.; et al. Associations between breast cancer risk and the catalase genotype, fruit and vegetable consumption, and supplement use. Am. J. Epidemiol 2005, 162, 943–952. [Google Scholar]
- Nadif, R.; Mintz, M.; Jedlicka, A.; Bertrand, J.P.; Kleeberger, S.R.; Kauffmann, F. Association of CAT polymorphisms with catalase activity and exposure to environmental oxidative stimuli. Free Radic. Res 2005, 39, 1345–1350. [Google Scholar]
- Bastaki, M.; Huen, K.; Manzanillo, P.; Chande, N.; Chen, C.; Balmes, J.R.; Tager, I.B.; Holland, N. Genotype-activity relationship for Mn-superoxide dismutase, glutathione peroxidases 1 and catalase in humans. Pharmacogenet. Genom 2006, 16, 279–286. [Google Scholar]
- Forsberg, L.; Lyrenás, L.; De Faire, U.; Morgenstern, R. A common functional C-T substitution polymorphism in the promoter region of the human catalase gene influences transcription factor binding, reporter gene transcription and is correlated to blood catalase levels. Free Radic. Biol. Med 2001, 30, 500–505. [Google Scholar]
- Labrecque, B.; Beaudry, D.; Mayhue, M.; Hallé, C.; Bordignon, V.; Murphy, B.D.; Palin, M.F. Molecular characterization and expression analysis of the porcine paraoxonase 3 (PON3) gene. Gene 2009, 443, 110–120. [Google Scholar]
- Costa, L.G.; Giordano, G.; Furlong, C.E. Pharmacological and dietary modulators of paraoxonase 1 (PON1) activity and expression: the hunt goes on. Biochem. Pharmacol 2011, 81, 337–344. [Google Scholar]
- Garin, M.C.; James, R.W.; Dussoix, P.; Blanché, H.; Passá, P.; Froguel, P.; Ruiz, J. Paraoxonase Polymorphism Met-Leu54 is associated with modified serum concentrations of the enzyme. A possible link between the paraoxonase gene and increased risk of cardiovascular disease in diabetes. J. Clin. Invest 1997, 99, 62–66. [Google Scholar]
- Cox, A.G.; Winterbourn, C.C.; Hampton, M.B. Mitochondrial peroxiredoxin involvement in antioxidant defense and redox signaling. Biochem. J 2010, 425, 313–325. [Google Scholar]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), MN-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic. Biol. Med 2002, 33, 337–349. [Google Scholar]
- Nakao, C.; Ookawara, T.; Sato, Y.; Kizaki, T.; Imazeki, N.; Matsubara, O.; Haga, S.; Suzuki, K.; Taniguchi, N.; Ohno, H. Extracellular superoxide dismutase in tissues from obese (ob/ob) mice. Free Radic. Res 2000, 33, 229–241. [Google Scholar]
- Adachi, T.; Inoue, M.; Hara, H.; Maehata, E.; Suzuki, S. Relationship of plasma extracellular-superoxide dismutase level with insulin resistance in type 2 diabetic patients. J. Endocrinol 2004, 181, 413–417. [Google Scholar]
- Jang, Y.C.; Pérez, V.I.; Song, W.; Lustgarten, M.S.; Salmon, A.B.; Mele, J.; Qi, W.; Liu, Y.; Liang, H.; Chaudhuri, A.; et al. Overexpression of Mn superoxide dismutase does not increase life span in mice. J. Gerontol. A Biol. Sci. Med. Sci 2009, 64, 1114–1125. [Google Scholar]
- Hoehn, K.L.; Salmon, A.B.; Hohnen-Behrens, C.; Turner, N.; Hoy, A.J.; Maghzal, G.J.; Stocker, R.; Van Remmen, H.; Kraegen, E.W.; Cooney, G.J.; et al. Insulin resistance is a cellular antioxidant defense mechanism. Proc. Natl. Acad. Sci. USA 2009, 106, 17787–17792. [Google Scholar]
- Liu, Y.; Qi, W.; Richardson, A.; van Remmen, H.; Ikeno, Y.; Salmon, A.B. Oxidative damage associated with obesity is prevented by overexpression of CuZn- or Mn-superoxide dismutase. Biochem. Biophys. Res. Commun 2013, 16, 78–83. [Google Scholar]
- Kurahashi, T.; Konno, T.; Otsuki, N.; Kwon, M.; Tsunoda, S.; Ito, J.; Fujii, J. A malfunction in triglyceride transfer from the intracellular lipid pool to apoB in enterocytes of SOD1-deficient mice. FEBS Lett 2012, 586, 4289–4295. [Google Scholar]
- Shimoda-Matsubayashi, S.; Matsumine, H.; Kobayashi, T.; Nakagawa-Hattori, Y. Structural dimorphism in the mitochondrial targeting sequence in the human manganese superoxide dismutase gene. Biochem. Biophys. Res. Commun 1996, 226, 561–565. [Google Scholar]
- El-Koofy, N.M.; El-Karaksy, H.M.; Mandour, I.M.; Anwar, G.M.; El-Raziky, M.S.; El-Hennawy, A.M. Genetic polymorphisms in non-alcoholic fatty liver disease in obese Egyptian children. Saudi J. Gastroenterol 2011, 17, 265–270. [Google Scholar]
- Bertelsen, M.; Anggard, E.E.; Carrier, M.J. Oxidative stress impairs insulin internalization inendothelial cells in vitro. Diabetologia 2001, 44, 605–613. [Google Scholar]
- Tirosh, A.; Potashnik, R.; Bashan, N.; Rudich, A. Oxidative stress disrupts insulin-induced cellular redistribution of insulin receptor substrate-1 and phosphatidylinositol 3-kinase in 3T3-L1 adipocytes: A putative cellular mechanism for impaired proteinkinase B activation and GLUT4 translocation. J. Biol.Chem 1999, 274, 10595–10602. [Google Scholar]
- Uthus, E.O.; Picklo, M.J., Sr. Obesity reduces methionine sulphoxide reductase activity in visceral adipose tissue. Free Radic. Res 2011, 45, 1052–1060. [Google Scholar]
- DeCoursey, T.E.; Ligeti, E. Regulation and termination of NADPH oxidase activity. Cell. Mol. Life Sci 2005, 62, 2173–2193. [Google Scholar]
- Eriksson, J.W. Metabolic stress in insulin’s target cells leads to ROS accumulation—A hypothetical common pathway causing insulin resistance. FEBS Lett 2007, 581, 3734–3742. [Google Scholar]
- Vidal-Puig, A.J.; Considine, R.V.; Jimenez-Liñan, M.; Werman, A.; Pories, W.J.; Caro, J.F.; Flier, J.S. Peroxisome proliferator-activated receptor gene expression in human tissues. Effects of obesity, weight loss, and regulation by insulin and glucocorticoids. J. Clin. Invest 1997, 99, 2416–2422. [Google Scholar]
- Berhouma, R.; Kouidhi, S.; Ammar, M.; Abid, H.; Ennafaa, H.; Benammar-Elgaaied, A. Correlation of peroxisome proliferator-activated receptor (PPAR-γ) mRNA expression with Pro12Ala polymorphism in obesity. Biochem. Genet 2013, 51, 256–263. [Google Scholar]
- Zielekiak, A.; Wójcik, M.; Woźniak, L.A. Structure and physiological functions of the human peroxisome proliferator-activated receptor γ. Arch. Immunol. Ther. Exp 2008, 56, 331–345. [Google Scholar]
- Nitz, I.; Ewert, A.; Klapper, M.; Döring, F. Analysis of PGC-1alpha variants Gly482Ser and Thr612Met concerning their PPARgamma2-coactivation function. Biochem. Biophys. Res. Commun 2007, 353, 481–486. [Google Scholar]
- Chartoumpekis, D.V.; Kensler, T.W. New player on an old field; the Keap1/Nrf2 pathway as a target for treatment of type 2 diabetes and metabolic syndrome. Curr. Diabetes Rev 2013, 9, 137–145. [Google Scholar]
- Vomhof-DeKrey, E.E.; Picklo, M.J. NAD(P)H:quinone oxidoreductase 1 activity reduces hypertrophy in 3T3-L1 adipocytes. Free Radic. Biol. Med 2012, 53, 690–700. [Google Scholar]
- Hou, Y.; Xue, P.; Bai, Y.; Liu, D.; Woods, C.G.; Yarborough, K.; Fu, J.; Zhang, Q.; Sun, G.; Collins, S.; et al. Nuclear factor erythroid-derived factor 2-related factor 2 regulates transcription of CCAAT/enhancer-binding protein β during adipogenesis. Free Radic. Biol. Med 2012, 52, 462–472. [Google Scholar]
- Chartoumpekis, D.V.; Ziros, P.G.; Sykiotis, G.P.; Zaravinos, A.; Psyrogiannis, A.I.; Kyriazopoulou, V.E.; Spandidos, D.A.; Habeos, I.G. Nrf2 activation diminishes during adipocyte differentiation of ST2 cells. Int. J. Mol. Med 2011, 28, 823–238. [Google Scholar]
- Chartoumpekis, D.V.; Ziros, P.G.; Psyrogiannis, A.I.; Papavassiliou, A.G.; Kyriazopoulou, V.E.; Sykiotis, G.P.; Habeos, I.G. Nrf2 represses FGF21 during long-term high-fat diet-induced obesity in mice. Diabetes 2011, 60, 2465–2473. [Google Scholar]
- Pi, J.; Leung, L.; Xue, P.; Wang, W.; Hou, Y.; Liu, D.; Yehuda-Shnaidman, E.; Lee, C.; Lau, J.; Kurtz, T.W.; et al. Deficiency in the nuclear factor E2-related factor-2 transcription factor results in impaired adipogenesis and protects against diet-induced obesity. J. Biol. Chem 2010, 285, 9292–9300. [Google Scholar]
- Shin, S.; Wakabayashi, J.; Yates, M.S.; Wakabayashi, N.; Dolan, P.M.; Aja, S.; Liby, K.T.; Sporn, M.B.; Yamamoto, M.; Kensler, T.W. Role of Nrf2 in prevention of high-fat diet-induced obesity by synthetic triterpenoid CDDO-imidazolide. Eur. J. Pharmacol 2009, 620, 138–144. [Google Scholar]
- Meher, A.K.; Sharma, P.R.; Lira, V.A.; Yamamoto, M.; Kensler, T.W.; Yan, Z.; Leitinger, N. Nrf2 deficiency in myeloid cells is not sufficient to protect mice from high-fat diet-induced adipose tissue inflammation and insulin resistance. Free Radic. Biol. Med 2012, 52, 1708–1715. [Google Scholar]
- Das, S.K.; Sharma, N.K.; Hasstedt, S.J.; Mondal, A.K.; Ma, L.; Langberg, K.A.; Elbein, S.C. An integrative genomics approach identifies activation of thioredoxin/thioredoxin reductase-1-mediated oxidative stress defense pathway and inhibition of angiogenesis in obese nondiabetic human subjects. J. Clin. Endocrinol. Metab 2011, 96, E1308–E1313. [Google Scholar]
- Cho, H.Y. Genomic Structure Variation of Nuclear Factor (Erythroid-Derived 2)-Like 2. Oxid. Med. Cell. Longev 2013. [Google Scholar] [CrossRef]
- Marzec, J.M.; Christie, J.D.; Reddy, S.P.; Jedlicka, A.E.; Vuong, H.; Lanken, P.N.; Aplenc, R.; Yamamoto, T.; Yamamoto, M.; Cho, H.Y.; Kleeberger, S.R. Functional polymorphisms in the transcription factor NRF2 in humans increase the risk of acute lung injury. FASEB J 2007, 21, 2237–2246. [Google Scholar]
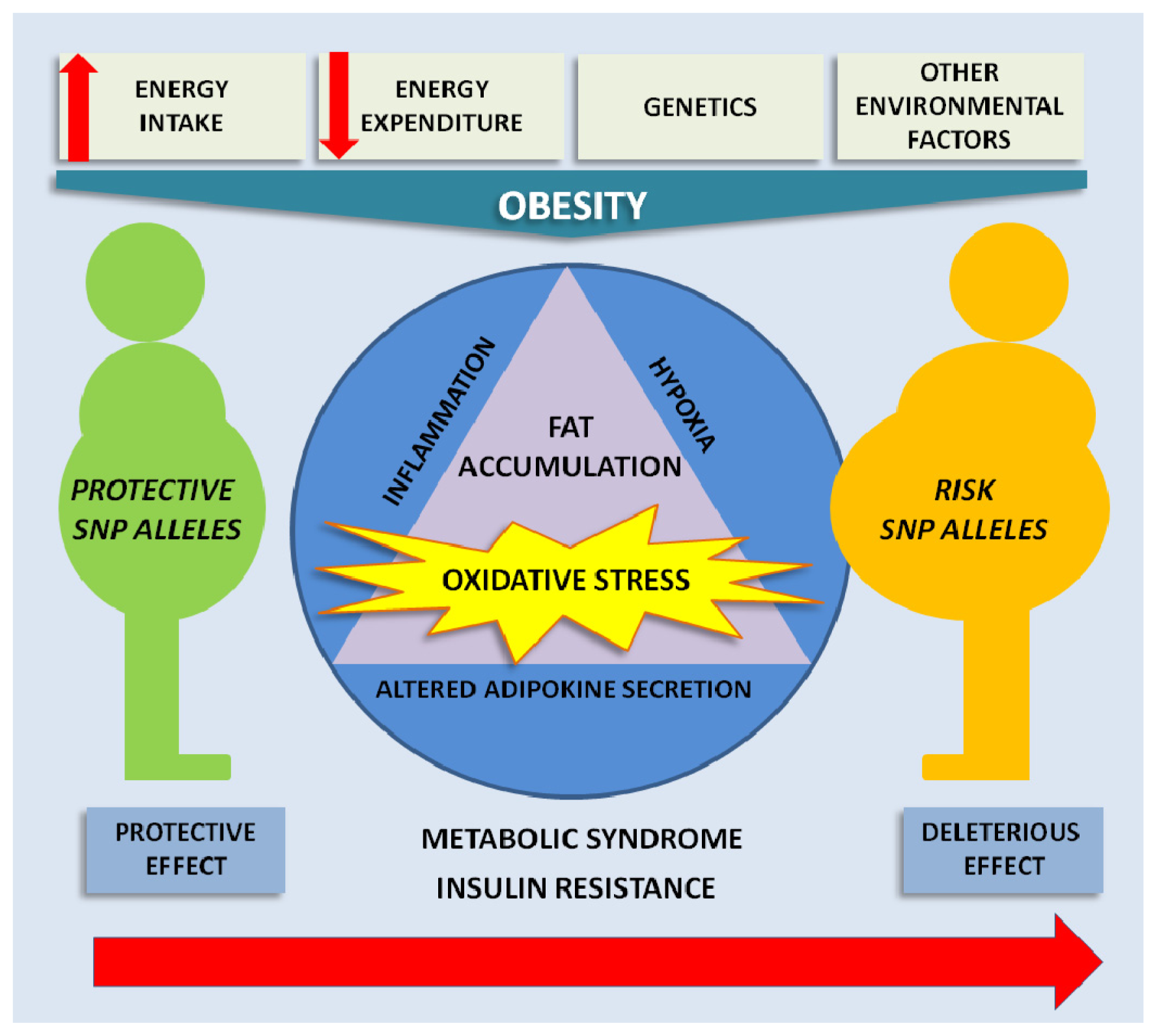

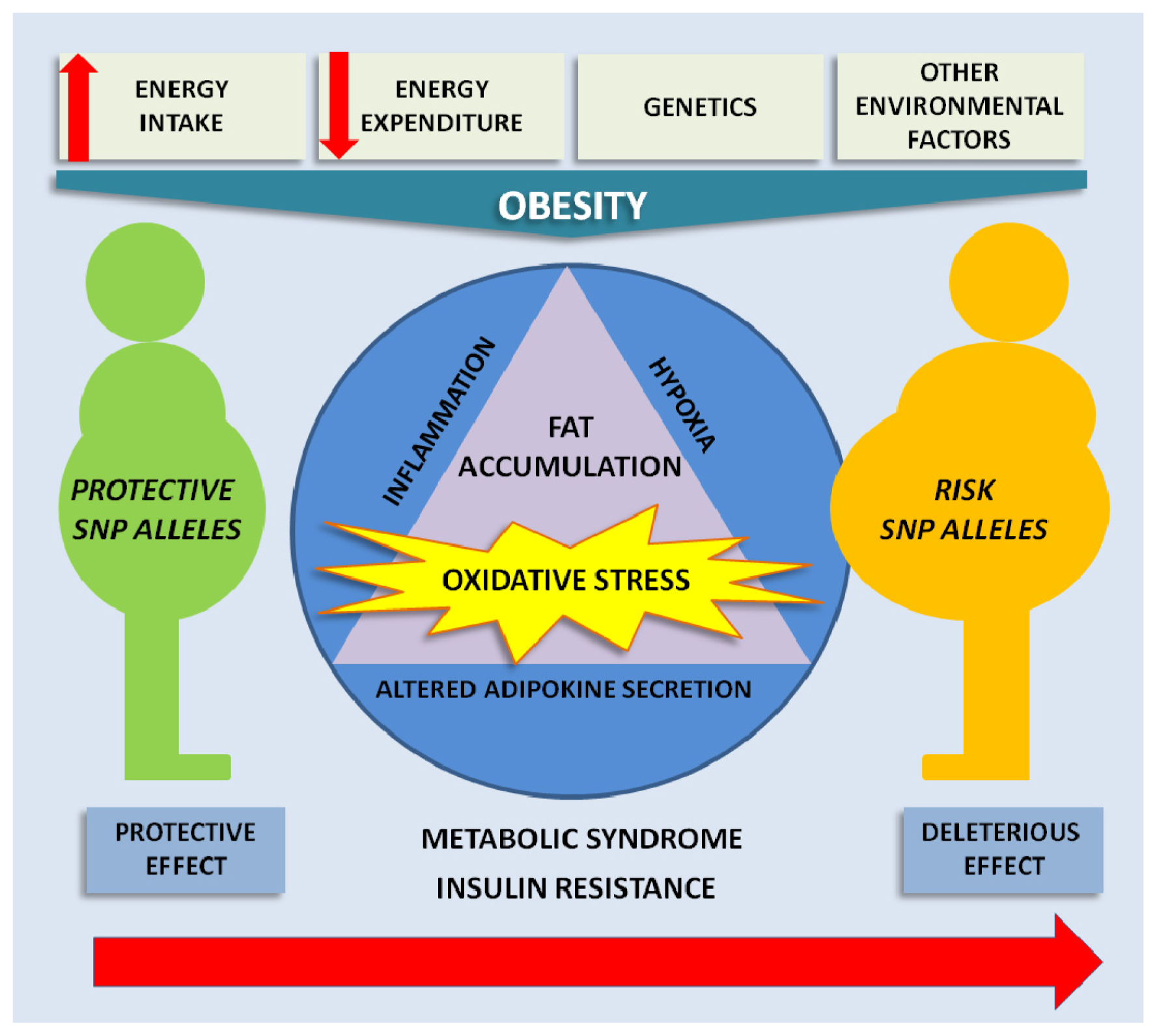{kind=link}
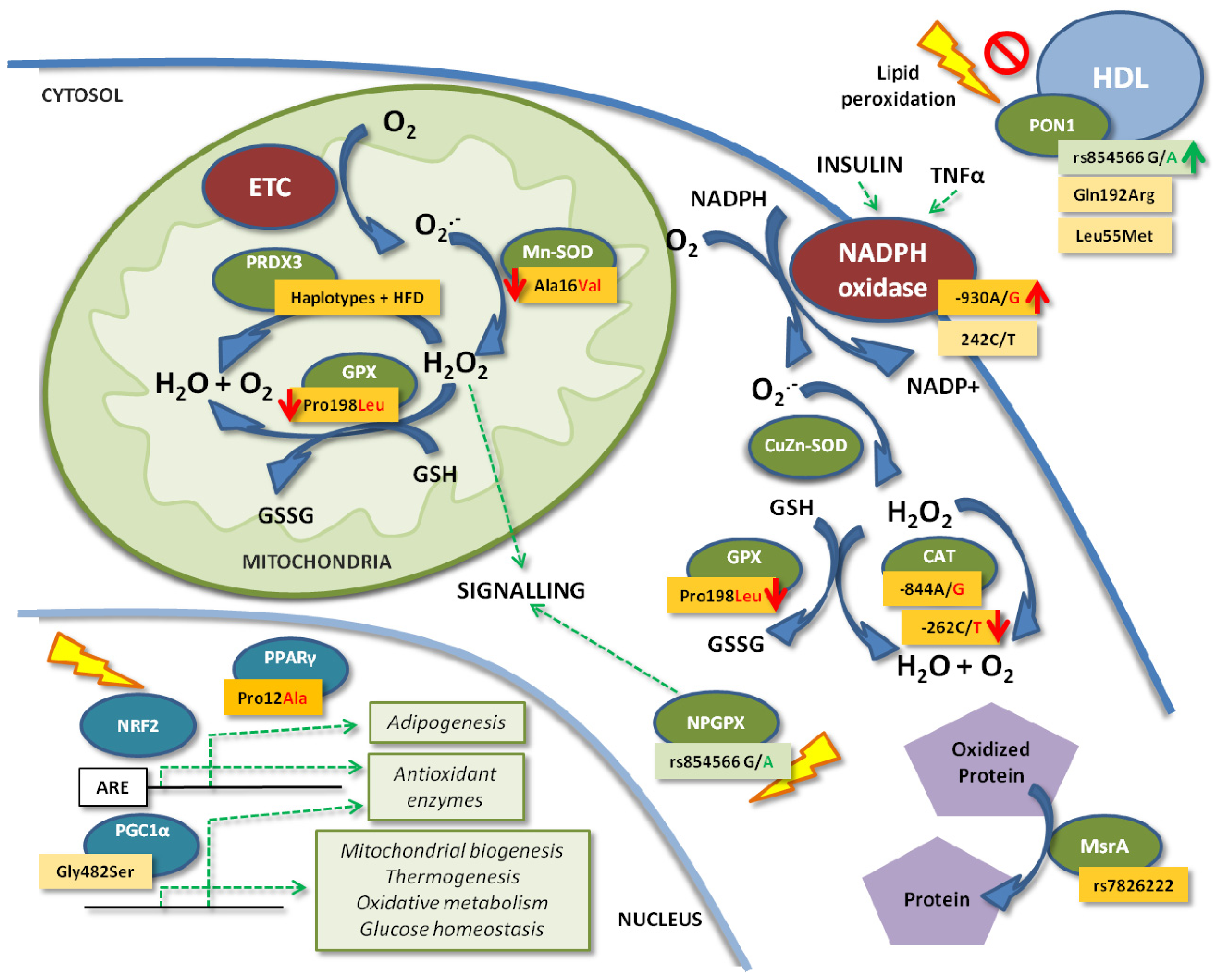{kind=link}
| Marker | References | |
|---|---|---|
| Adults | ↑ F2α-isoprostane | [12,16–18] |
| ↑ MDA/TBARS | [11,19,20] | |
| ↑ Ox-LDL | [16,21,22] | |
| ↓ PON1 activity | [11,23,24] (Non-significant: [25–27]) | |
| ↓ TAC | [27] | |
| Children | ↑ AOPPs | [28] |
| ↓ CAT activity | [15] | |
| ↑ F2α-isoprostane | [13,29–31] | |
| ↓ GPX activity | [14] (pubertal) (Non-significant: [15], prepubertal) | |
| ↑ MDA | [13,32] | |
| ↑ Nitrite/nitrate | [14] | |
| ↑ NO | [13] | |
| ↑ Ox-LDL | [33] | |
| ↓ PON1 activity | [34–36] (Non-significant: [37]) | |
| ↓ TAC | [38] | |
| Gene | Analyzed variant (dbSNP) | Other designations | Alleles 1/2 | Outcome | References |
|---|---|---|---|---|---|
| GPX1 | rs1050450 | 594C/T, Pro198Leu | C/T | Leu male carriers have higher waist-hip ratio, TAG, insulin, HOMA-β, SBP, DBP | [60] |
| Leu female carriers have higher body fat mass, insulin and HOMA-IR | |||||
| Leu carriers show higher DNA damage after Se supplementation | [61] | ||||
| Leu carriers have higher lipoperoxides and MDA in LDL | [62] | ||||
| GPX1 | Ala5/Ala6 + Pro198Leu | 40% decrease in GPX1 activity | [63] | ||
| GPX1 | −602A/G + 2C/T | 25% decrease in transcriptional activity | [63] | ||
| GPX7 | rs835337 | G/A | A allele Associated with decreased BMI in adults (British, Finnish and Han Chinese) and with increased GPX7 expression in abdominal adipose tissue and decreased serum MDA concentrations | [4] | |
| CAT | rs769214 | −844A/G | A/G | Associated with prepubertal obesity, higher weight, BMI Z-score, A-FABP | [15] |
| CAT | rs7943316 | −89T/A | A/T | Associated with prepubertal obesity | [15] |
| CAT | rs1049982 | −20C/T | C/T | Associated with prepubertal obesity | [15] |
| CAT | −844G/−89A/−20T | Lower CAT expression under high oxidative stress conditions | [66] | ||
| PON1 | rs662 | Q192R | G/A | 192RR (Arg-Arg) individuals show higher activity degrading paraoxon | [67] |
| R allele associated with higher obesity risk | [26] | ||||
| No association with obesity in Mexican adults | [25] | ||||
| No association with obesity in adolescents | [68] | ||||
| No association with obesity in prepubertal children | [37] | ||||
| PON1 | rs854560 | L55M | A/T | 55LL (Leu-Leu) individuals have increased serum PON1 concentrations | [35] |
| PON1 | rs854566 | G/A | Protection against prepubertal childhood obesity and lactonase activity | [37] | |
| PRDX3 | rs3740562* | A/G | Associated with higher BMI in Japanese adults when combined with high-fat diet | [69] | |
| PRDX3 | rs2271362* | C/T | Associated with higher BMI in Japanese adults when combined with high-fat diet | [69] | |
| PRDX3 | rs7768* | G/C | Associated with higher BMI in Japanese adults when combined with high-fat diet | [69] | |
| PRDX3 | rs3377* | A/C | Associated with higher BMI in Japanese adults when combined with high-fat diet | [69] | |
| MSRA | rs7826222 | rs545854 | G/C | Associated with WC in Europeans | [70,71] |
| Associated with BMI in Hispanic women | [72] | ||||
| Associated with higher type 2 diabetes risk | [73] | ||||
| Not associated with BMI in Chinese | [74] | ||||
| Not associated with metabolic syndrome or visceral fat in Japanese | [75,76] | ||||
| rs473034 | G/A | Associated with extreme childhood obesity | [77] | ||
| rs516175 | C/T | Associated with BMI Z-score in Singaporeans | [78] | ||
| No effect on lifestyle intervention in overweight children and adolescents | [79] | ||||
| rs13278851 | G/A | No effect on lifestyle intervention in overweight children and adolescents | [79] | ||
| rs17150703 | G/A | No effect on lifestyle intervention in overweight children and adolescents | [79] | ||
| SOD2 | rs4880 | Ala16Val | C/T | Val variant associated with MnSOD arrest in the inner mitochondrial membrane and lower dismutase efficiency | [80] |
| Val allele associated with higher obesity risk in elderly | [81] | ||||
| Val allele show higher levels of IL-1, IL-6, TNFα, IFNγ and lower levels of IL-10 | [82] | ||||
| Val allele carriers show lower DNA damage levels | [83] | ||||
| p22phox | rs9932581 | −930A/G | A/G | GG carriers have higher p22phox expression and NADPH oxidase activity | [84] |
| GG carriers have higher HOMA-IR and insulin but not higher obesity risk | [85] | ||||
| p22phox | rs4673 | 242C/T | C/T | T allele type 2 diabetic carriers have lower IMT and 8-OHdG values, non-diabetic carriers have lower HOMA-IR and insulin | [86] |
| CC carriers protected against diabetes and obesity, with lower plasma glucose levels and WC in hypertensive patients | [87] | ||||
| p22phox | rs7195830 | C/T | C allele associated with higher obesity risk in women | [88] | |
| p22phox | rs12709102 | T/C | T allele associated with higher obesity risk in women | [88] | |
| PPARγ | rs1801282 | Pro12Ala | C/G | Ala allele decreases receptor mediated transcriptional activity and is associated with lower BMI and increased insulin sensitivity | [89] |
| Ala allele associated with increased BMI (+0.065 kg/m2 per allele) | [90] | ||||
| PGC1α | rs8192678 | Gly482Ser | G/A | Ser allele associated with HOMA-IR in obese subjects | [91] |
| Ser variant associated with lower BMI, waist and hip circumference and total body fat in women | [92] | ||||
| Not associated with obesity or type 2 diabetes in overweight Chinese subjects but associated with high insulin, HOMA-IR and waist-hip ratio, as well as TBARS, in patients with hyperglycemia | [93] | ||||
| Ser male diabetic carriers have lower adiponectin plasma levels | [94] | ||||
| Ser carriers have higher HOMA-IR and insulin concentrations, but eight-week low calorie diet reduces the risk | [95] |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rupérez, A.I.; Gil, A.; Aguilera, C.M. Genetics of Oxidative Stress in Obesity. Int. J. Mol. Sci. 2014, 15, 3118-3144. https://doi.org/10.3390/ijms15023118
Rupérez AI, Gil A, Aguilera CM. Genetics of Oxidative Stress in Obesity. International Journal of Molecular Sciences. 2014; 15(2):3118-3144. https://doi.org/10.3390/ijms15023118
Chicago/Turabian StyleRupérez, Azahara I., Angel Gil, and Concepción M. Aguilera. 2014. "Genetics of Oxidative Stress in Obesity" International Journal of Molecular Sciences 15, no. 2: 3118-3144. https://doi.org/10.3390/ijms15023118
APA StyleRupérez, A. I., Gil, A., & Aguilera, C. M. (2014). Genetics of Oxidative Stress in Obesity. International Journal of Molecular Sciences, 15(2), 3118-3144. https://doi.org/10.3390/ijms15023118