Predicting the Function of 4-Coumarate:CoA Ligase (LJ4CL1) in Lonicera japonica

Abstract
:1. Introduction
2. Results and Discussion
2.1. Global Phylogeny and Duplication of AMP-Binding Proteins
2.2. Global Phylogeny and Duplication of GEICIRG-Containing Proteins
2.3. Expression of GEICIRG-Containing Proteins in L. japonica Flowers
2.4. Substrate-Binding Diversity in the Expressed GEICIRG-Containing Proteins in L. japonica
3. Experimental Section
3.1. Classification of AMP-Dependent Synthetase/Ligase Sequences and GEICIRG-Motif Proteins
3.2. Phylogeny of AMP-Dependent Synthetase/Ligase Sequences and GEICIRG-Motif Proteins
3.3. Indentification of Orthologs and Paralogs
3.4. Gene Expression Analyses
3.5. Protein Structure and Binding Site Prediction
3.6. LC-MS Analysis of L. japonica Flowers
3.7. Expression of 4CL Protein in E.coli and Enzyme Activity Assay
4. Conclusions
Supplementary Information
ijms-15-02386-s001.pdfAcknowledgments
Conflicts of Interest
References
- Hu, W.J.; Kawaoka, A.; Tsai, C.J. Compartmentalized expression of two structurally and functionally distinct 4-coumarate:CoA ligase genes in aspen (Populus tremuloides). Proc. Natl. Acad. Sci. USA 1998, 95, 5407–5412. [Google Scholar]
- Hamberger, B.; Hahlbrock, K. The 4-coumarate:CoA ligase gene family in Arabidopsis thaliana comprises one rare, sinapate-activating and three commonly occurring isoenzymes. Proc. Natl. Acad. Sci. USA 2004, 101, 2209–2214. [Google Scholar]
- Ververidis, F.; Trantas, E.; Douglas, C. Biotechnology of flavonoids and other phenylpropanoid-derived natural products. Part I: Chemical diversity, impacts on plant biology and human health. Biotechnol. J 2007, 2, 1214–1234. [Google Scholar]
- Blach-Olszewska, Z.; Jatczak, B.; Rak, A. Production of cytokines and stimulation of resistance to viral infection in human leukocytes by Scutellaria baicalensis flavones. J. Interf. Cytokine Res 2008, 28, 571–581. [Google Scholar]
- Kaneko, M.; Ohnishi, Y.; Horinouchi, S. Cinnamate: Coenzyme A ligase from the filamentous bacterium Streptomyces coelicolor A3(2). J. Bacteriol 2003, 185, 20–27. [Google Scholar]
- Cohen-Gihon, I.; Nussinov, R.; Sharan, R. Comprehensive analysis of co-occurring domain sets in yeast proteins. BMC Genomics 2007, 8, 161. [Google Scholar]
- Bairoch, A. Prosite—A dictionary of sites and patterns in proteins. Nucleic Acids Res 1991, 19, 2241–2245. [Google Scholar]
- Becker-Andre, M.; Schulze-Lefert, P.; Hahlbrock, K. Structural comparison, modes of expression, and putative cis-acting elements of the two 4-coumarate:CoA ligase genes in potato. J. Biol. Chem 1991, 266, 8551–8559. [Google Scholar]
- Fulda, M.; Heinz, E.; Wolter, F.P. The fadd gene of Escherichia coli K12 is located close to rnd at 39.6 min of the chromosomal map and is a new member of the amp-binding protein family. Mol. Gen. Genet 1994, 242, 241–249. [Google Scholar]
- Stuible, H.P.; Buttner, D.; Ehlting, J. Mutational analysis of 4-coumarate:CoA ligase identifies functionally important amino acids and verifies its close relationship to other adenylate-forming enzymes. FEBS Lett 2000, 467, 117–122. [Google Scholar]
- Cukovic, D.; Ehlting, J.; VanZiffle, J.A. Structure and evolution of 4-coumarate:Coenzyme A ligase (4CL) gene families. Biol. Chem 2001, 382, 645–654. [Google Scholar]
- Souza, C.D.; Kim, S.S.; Koch, S. A novel fatty acyl-coA synthetase is required for pollen development and sporopollenin biosynthesis in Arabidopsis. Plant Cell 2009, 21, 507–525. [Google Scholar]
- Costa, M.A.; Bedgar, D.L.; Moinuddin, S.G.A. Characterization in vitro and in vivo of the putative multigene 4-coumarate:CoA ligase network in Arabidopsis: Syringyl lignin and sinapate/sinapyl alcohol derivative formation. Phytochemistry 2005, 66, 2072–2091. [Google Scholar]
- Gaid, M.M.; Scharnhop, H.; Ramadan, H. 4-Coumarate:CoA ligase family members from elicitor-treated Sorbus aucuparia cell cultures. J. Plant Physiol 2011, 168, 944–951. [Google Scholar]
- Allina, S.M.; Pri-Hadash, A.; Theilmann, D.A. 4-coumarate:Coenzyme A ligase in hybrid poplar—Properties of native enzymes, cDNA cloning, and analysis of recombinant enzymes. Plant Physiol 1998, 116, 743–754. [Google Scholar]
- Silber, M.V.; Meimberg, H.; Ebel, J. Identification of a 4-coumarate:CoA ligase gene family in the moss Physcomitrella patens. Phytochemistry 2008, 69, 2449–2456. [Google Scholar]
- Hamada, K.; Nishida, T.; Yamauchi, K. 4-Coumarate:Coenzyme A ligase in black locust (Robinia pseudoacacia) catalyses the conversion of sinapate to sinapoyl-CoA. J. Plant Res 2004, 117, 303–310. [Google Scholar]
- Rastogi, S.; Kumar, R.; Chanotiya, C.S. 4-Coumarate:CoA ligase partitions metabolites for eugenol biosynthesis. Plant Cell Physiol 2013, 54, 1238–1252. [Google Scholar]
- Klempien, W.; Kaminaga, Y.; Qualley, A. Contribution of CoA ligases to benzenoid biosynthesis in petunia flowers. Plant Cell 2012. [Google Scholar] [CrossRef]
- Morita, H.; Mori, T.; Wanibuchi, K. Crystallization and preliminary X-ray analysis of 4-coumarate:CoA ligase from Arabidopsis thaliana. Acta Crystallogr. Sect. F-Struct. Biol. Cryst. Commun 2011, 67, 409–411. [Google Scholar]
- Hu, Y.L.; Gai, Y.; Yin, L. Crystal structures of a populus tomentosa 4-coumarate:CoA ligase shed light on its enzymatic mechanisms. Plant Cell 2010, 22, 3093–3104. [Google Scholar]
- Schneider, K.; Hovel, K.; Witzel, K. The substrate specificity-determining amino acid code of 4-coumarate:CoA ligase. Proc. Natl. Acad. Sci. USA 2003, 100, 8601–8606. [Google Scholar]
- Kumar, A.; Ellis, B.E. 4-Coumarate:CoA ligase gene family in Rubus idaeus: cDNA structures, evolution, and expression. Plant Mol. Biol 2003, 51, 327–340. [Google Scholar]
- Yang, J.M.; Chen, F.; Yu, O. Controlled silencing of 4-coumarate:CoA ligase alters lignocellulose composition without affecting stem growth. Plant Physiol. Biochem 2011, 49, 103–109. [Google Scholar]
- Wagner, A.; Donaldson, L.; Kim, H. Suppression of 4-coumarate-CoA ligase in the coniferous gymnosperm Pinus radiata. Plant Physiol 2009, 149, 370–383. [Google Scholar]
- Chen, C.Y.; Qi, L.W.; Li, H.J.; Li, P.; Yi, L.; Liang, H.; Tang, M.D. Simultaneous determination of iridoids, phenolicacids, flavonoids, and saponins in FlosLonicerae and FlosLonicerae Japonicae by HPLC-DAD-ELSDcoupled with principal component analysis. J. Sep. Sci 2007, 30, 3181–3192. [Google Scholar]
- Qi, L.W.; Chen, C.Y.; Li, P. Structural characterization and identification of iridoidglycosides, saponins, phenolic acids and flavonoids in Flos Lonicerae Japonicae by a fast liquidchromatographymethod with diode-array detection and time-of-flightmass spectrometry. Rapid Commun. Mass Spectrom 2009, 23, 3227–3242. [Google Scholar]
- Peng, Y.Y.; Liu, F.H.; Ye, J.N. Determination of phenolic acids and flavones in Lonicerajaponica thumb by capillary electrophoresis with electrochemical detection. Electroanalysis 2005, 17, 356–362. [Google Scholar]
- Kushwaha, H.R.; Singh, A.K.; Sopory, S.K. Genome wide expression analysis of CBS domain containing proteins in Arabidopsis thaliana (L.) Heynh and Oryza sativa L. reveals their developmental and stress regulation. BMC Genomics 2009, 10, 200. [Google Scholar]
- Yuan, Y.; Song, L.; Li, M. Genetic variation and metabolic pathway intricacy govern the active compound content and quality of the Chinese medicinal plant Lonicera japonica thunb. BMC Genomics 2012, 13, 195. [Google Scholar]
- Souza, C.A.; Barbazuk, B.; Ralph, S.G. Genome-wide analysis of a land plant-specific acyl:coenzyme A synthetase (ACS) gene family in Arabidopsis, poplar, rice and Physcomitrella. New Phytol 2008, 179, 987–1003. [Google Scholar]
- Stein, T.; Vater, J.; Kruft, V. The multiple carrier model of nonribosomal peptide biosynthesis at modular multienzymatic templates. J. Biol. Chem 1996, 271, 15428–15435. [Google Scholar]
- Huang, S.; Li, R.; Zhang, Z. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar]
- Tuskan, G.A.; Difazio, S.; Jansson, S. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar]
- The Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815.
- Jaillon, O.; Aury, J.M.; Noel, B. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar]
- Schnable, P.S.; Ware, D.; Fulton, R.S. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R. The Sorghum bicolor genome and the diversification of grasses. Nature 2009, 457, 551–556. [Google Scholar]
- Yu, J.; Hu, S.; Wang, J. A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 2002, 296, 92–100. [Google Scholar]
- Khurana, P.; Gokhale, R.S.; Mohanty, D. Genome scale prediction of substrate specificity for acyl adenylate superfamily of enzymes based on active site residue profiles. BMC Bioinforma 2010, 11, 57. [Google Scholar]
- Aerts, R.J.; Baumann, T.W. Distribution and utilization of chlorogenic acid in coffea seedlings. J. Exp. Bot 1994, 45, 497–503. [Google Scholar]
- Schoch, G.; Goepfert, S.; Morant, M. CYP98A3 from Arabidopsis thaliana is a 3′-hydroxylase of phenolic esters, a missing link in the phenylpropanoid pathway. J. Biol. Chem 2001, 276, 36566–36574. [Google Scholar]
- Bateman, A.; Birney, E.; Durbin, R. The Pfam protein families database. Nucleic Acids Res 2000, 28, 263–266. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 1997, 25, 3389–3402. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol 2011, 28, 2731–2739. [Google Scholar]
- Jeanmougin, F.; Thompson, J.D.; Gouy, M. Multiple sequence alignment with Clustal x. Trends Biochem. Sci 1998, 23, 403–405. [Google Scholar]
- Tatusov, R.L.; Koonin, E.V.; Lipman, D.J. A genomic perspective on protein families. Science 1997, 278, 631–637. [Google Scholar]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol 2010, 11, R25. [Google Scholar]
- Mortazavi, A.; Williams, B.A.; Mccue, K. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinforma 2008, 9, 40. [Google Scholar]
- Voo, K.S.; Whetten, R.W.; O’Malley, D.M.; Sederoff, R.R. 4-Coumarate: Coenzyme A ligase from loblolly pine xylem (isolation, characterization, and complementary DNA cloning). Plant Physiol 1995, 108, 85–97. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.R.; Farr, A.L.; Randall, R.J. Protein measurement with the Folinphenol reagent. J. Biol. Chem 1951, 193, 265–269. [Google Scholar]
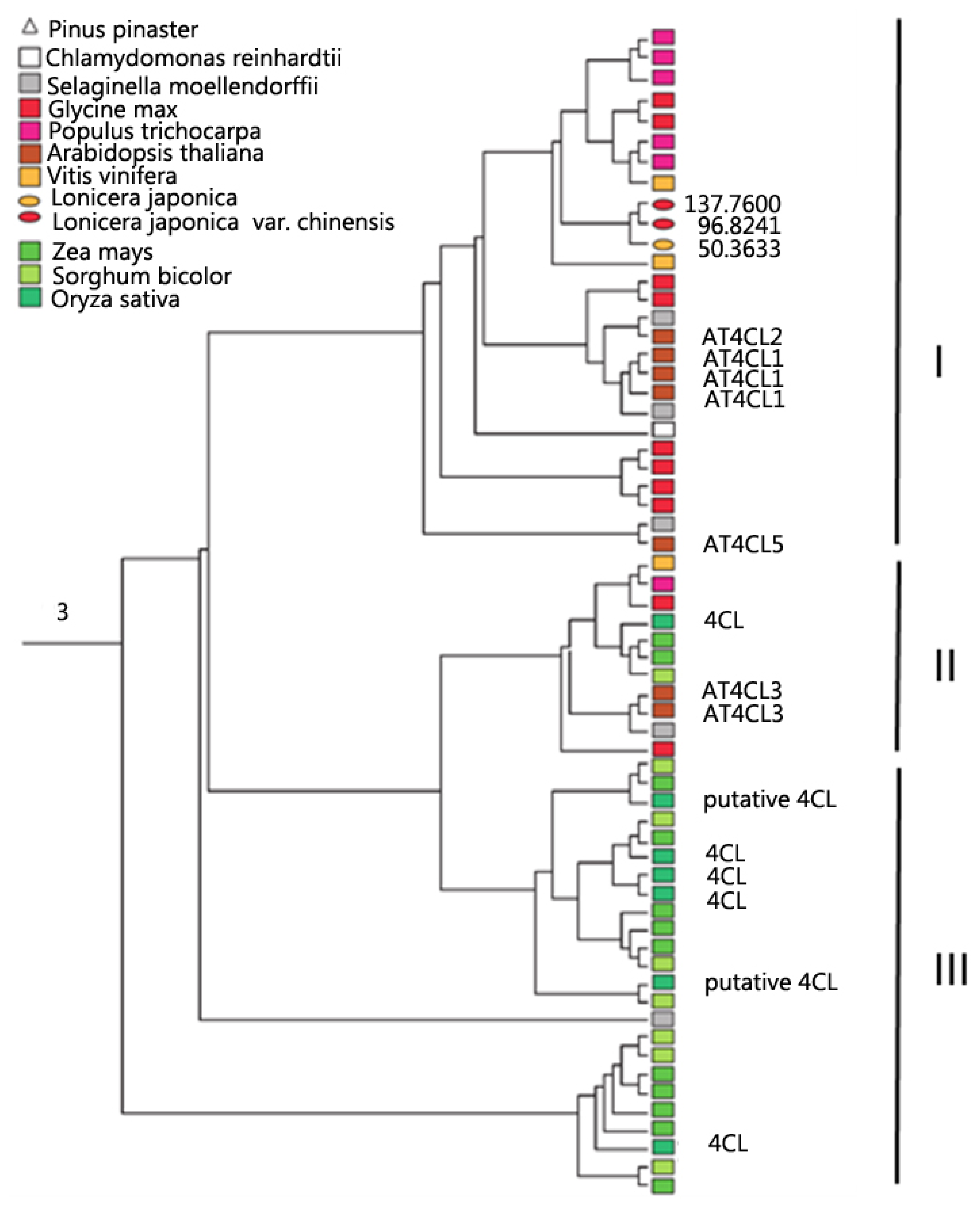


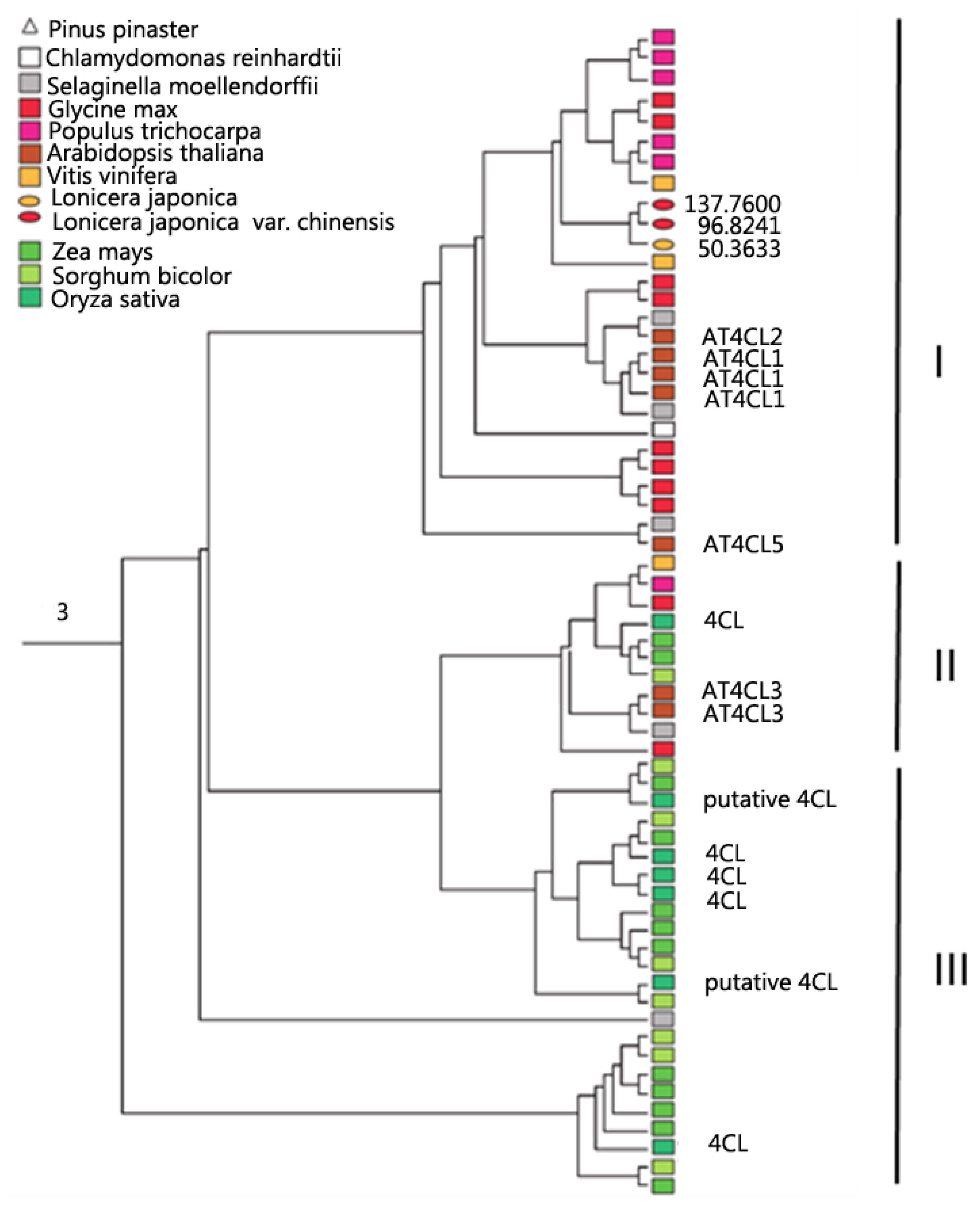{kind=link}
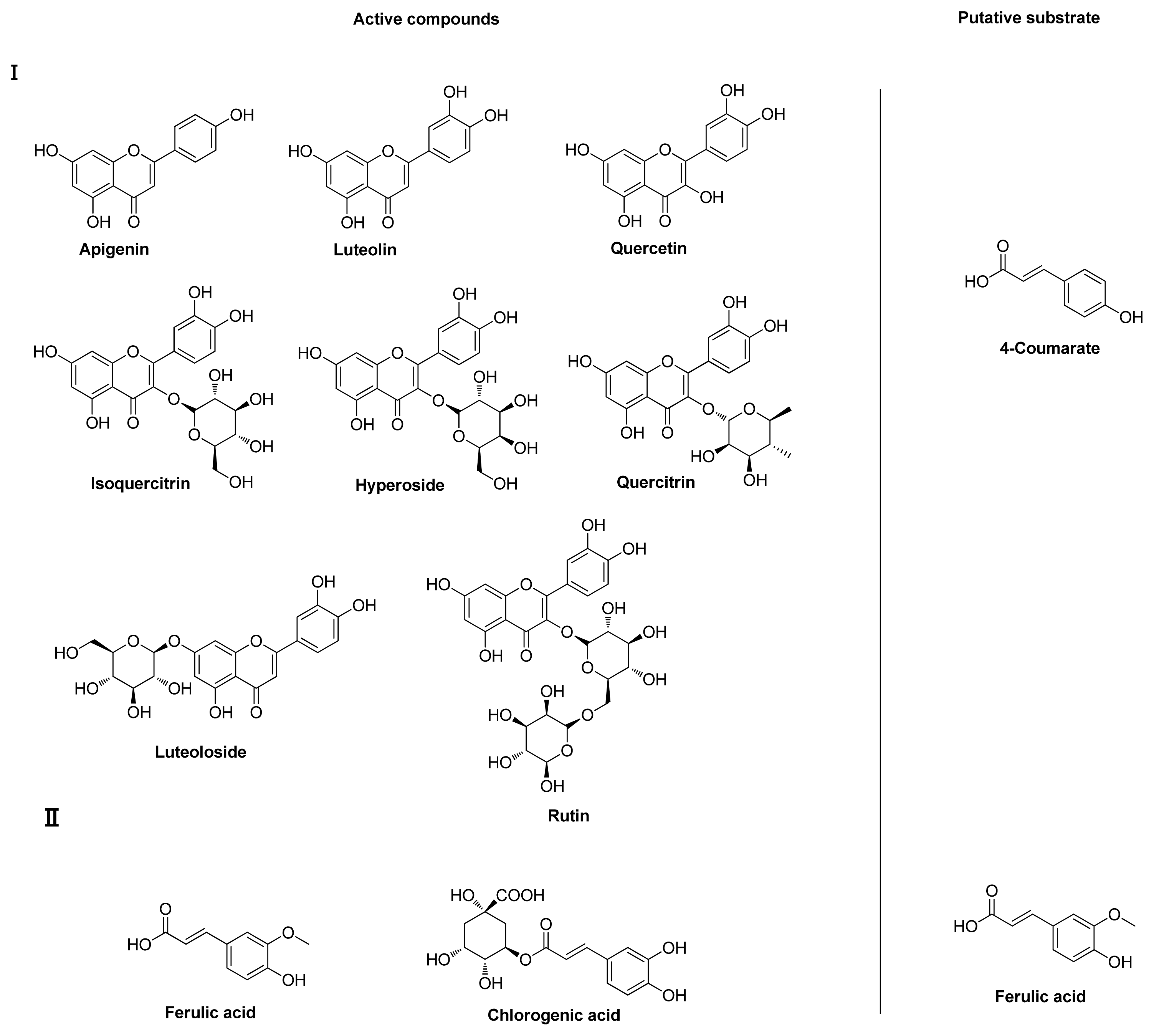{kind=link}
| Kingdom Group | Class | Clusters * | Number of copies | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | 1 | 1-1 | 1-2 | 1-3 | 1-4 | 2 | 2-1 | 2-2 | 2-3 | 2-4 | 3 | Total | |||
| Animal | Culex quinquefasciatus | 39 | 5 | 5 | 27 | 2 | 6 | 1 | 4 | 1 | 0 | 1 | 46 | ||
| Bacteria | Escherichia coli | 5 | 0 | 3 | 2 | 0 | 3 | 0 | 2 | 1 | 0 | 2 | 10 | ||
| Fungus | Penicillium marneffei | 20 | 3 | 3 | 12 | 2 | 25 | 3 | 7 | 15 | 0 | 5 | 50 | ||
| Aspergillus nidulans | 32 | 2 | 8 | 17 | 5 | 28 | 3 | 5 | 19 | 1 | 1 | 61 | |||
| Plant | Algae | Chlamydomonas reinhardtii | 18 | 11 | 2 | 0 | 5 | 9 | 0 | 8 | 0 | 1 | 0 | 27 | |
| Bryophyte | Physcomitrella patens | 43 | 8 | 16 | 14 | 5 | 2 | 0 | 0 | 1 | 1 | 0 | 45 | ||
| Pteridophyta | Selaginella moellendorffii | 70 | 15 | 26 | 26 | 3 | 14 | 3 | 9 | 2 | 0 | 0 | 84 | ||
| Gymnospermae | Pinus taeda | 4 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | ||
| Larix gmelinii | 18 | 0 | 0 | 18 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 18 | |||
| Pseudotsuga menziesii | 17 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 17 | |||
| Angiospermae | Dicotyledoneae | Glycine max | 99 | 30 | 29 | 35 | 5 | 12 | 5 | 6 | 1 | 0 | 0 | 111 | |
| Populus trichocarpa | 71 | 21 | 24 | 23 | 3 | 8 | 3 | 4 | 1 | 0 | 0 | 79 | |||
| Arabidopsis thaliana | 48 | 13 | 17 | 16 | 2 | 6 | 0 | 5 | 1 | 0 | 0 | 54 | |||
| Arabidopsis lyrata | 146 | 24 | 0 | 122 | 0 | 25 | 8 | 15 | 2 | 0 | 1 | 172 | |||
| Vitis vinifera | 34 | 8 | 9 | 14 | 3 | 6 | 1 | 4 | 1 | 0 | 0 | 40 | |||
| Lonicera japonica | 103 | 43 | 36 | 17 | 7 | 14 | 0 | 12 | 2 | 0 | 0 | 117 | |||
| Lonicera japonica var. chinensis | 73 | 33 | 20 | 16 | 4 | 12 | 0 | 10 | 2 | 0 | 1 | 86 | |||
| Monocotyledoneae | Zea mays | 82 | 23 | 21 | 35 | 3 | 15 | 3 | 9 | 3 | 0 | 0 | 97 | ||
| Sorghum bicolor | 48 | 16 | 13 | 17 | 2 | 10 | 3 | 6 | 1 | 0 | 0 | 58 | |||
| Oryza sativa | 60 | 31 | 12 | 16 | 1 | 16 | 3 | 9 | 4 | 0 | 0 | 76 | |||
| Total | 1252 | ||||||||||||||
| Clusters * | Number of copies | Total | Genome size | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|
| ACS | AAE | 4CL | ACS | ||||||
| Species | 1 | 2 | 3 | 4 | M | ||||
| Gymnospermae | Pinus pinaster | 0 | 0 | 1 | 0 | 1 | |||
| Algae | Chlamydomonas reinhardtii | 2 | 0 | 0 | 1 | 3 | |||
| Bryophyte | Physcomitrella patens | 0 | 3 | 12 | 0 | 15 | |||
| Pteridophyta | Selaginella moellendorffii | 3 | 2 | 5 | 6 | 16 | |||
| Angiospermae | Dicotyledoneae | Glycine max | 3 | 7 | 10 | 4 | 24 | 1100 | [33] |
| Populus trichocarpa | 5 | 1 | 6 | 2 | 14 | 485 | [34] | ||
| Arabidopsis thaliana | 2 | 1 | 7 | 5 | 15 | 135 | [35] | ||
| Vitis vinifera | 2 | 2 | 3 | 2 | 9 | 505 | [36] | ||
| Lonicera japonica | 5 | 4 | 1 | 5 | 15 | ||||
| Lonicera japonica var. chinensis | 4 | 2 | 2 | 3 | 11 | ~800 | Our group | ||
| Monocotyledoneae | Zea mays | 3 | 0 | 12 | 8 | 23 | 2300 | [37] | |
| Sorghum bicolor | 2 | 1 | 8 | 4 | 15 | 730 | [38] | ||
| Oryza sativa | 6 | 0 | 7 | 10 | 23 | 467 | [39] | ||
| Total | 37 | 23 | 74 | 50 | 184 | ||||
| Cluster * | Varieties | Lonicera japonica | Lonicera japonica var. chinensis | Total |
|---|---|---|---|---|
| Copy and RPKM | ||||
| 1 | Number of copies | 5 | 4 | 9 |
| RPKM | 320.4257 | 171.5426 | 491.9683 | |
| 2 | Number of copies | 4 | 2 | 6 |
| RPKM | 250.9583 | 306.7299 | 557.6882 | |
| 3 | Number of copies | 1 | 2 | 3 |
| RPKM | 50.3633 | 234.6021 | 284.9654 | |
| 4 | Number of copies | 5 | 3 | 8 |
| RPKM | 292.3302 | 236.8353 | 529.1655 | |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yuan, Y.; Yu, S.; Yu, J.; Zhan, Z.; Li, M.; Liu, G.; Wang, X.; Huang, L. Predicting the Function of 4-Coumarate:CoA Ligase (LJ4CL1) in Lonicera japonica. Int. J. Mol. Sci. 2014, 15, 2386-2399. https://doi.org/10.3390/ijms15022386
Yuan Y, Yu S, Yu J, Zhan Z, Li M, Liu G, Wang X, Huang L. Predicting the Function of 4-Coumarate:CoA Ligase (LJ4CL1) in Lonicera japonica. International Journal of Molecular Sciences. 2014; 15(2):2386-2399. https://doi.org/10.3390/ijms15022386
Chicago/Turabian StyleYuan, Yuan, Shulin Yu, Jun Yu, Zhilai Zhan, Minhui Li, Guiming Liu, Xumin Wang, and Luqi Huang. 2014. "Predicting the Function of 4-Coumarate:CoA Ligase (LJ4CL1) in Lonicera japonica" International Journal of Molecular Sciences 15, no. 2: 2386-2399. https://doi.org/10.3390/ijms15022386
APA StyleYuan, Y., Yu, S., Yu, J., Zhan, Z., Li, M., Liu, G., Wang, X., & Huang, L. (2014). Predicting the Function of 4-Coumarate:CoA Ligase (LJ4CL1) in Lonicera japonica. International Journal of Molecular Sciences, 15(2), 2386-2399. https://doi.org/10.3390/ijms15022386