Interactions of Borneol with DPPC Phospholipid Membranes: A Molecular Dynamics Simulation Study

Abstract
:1. Introduction
2. Results and Discussion
2.1. Concentration Effect
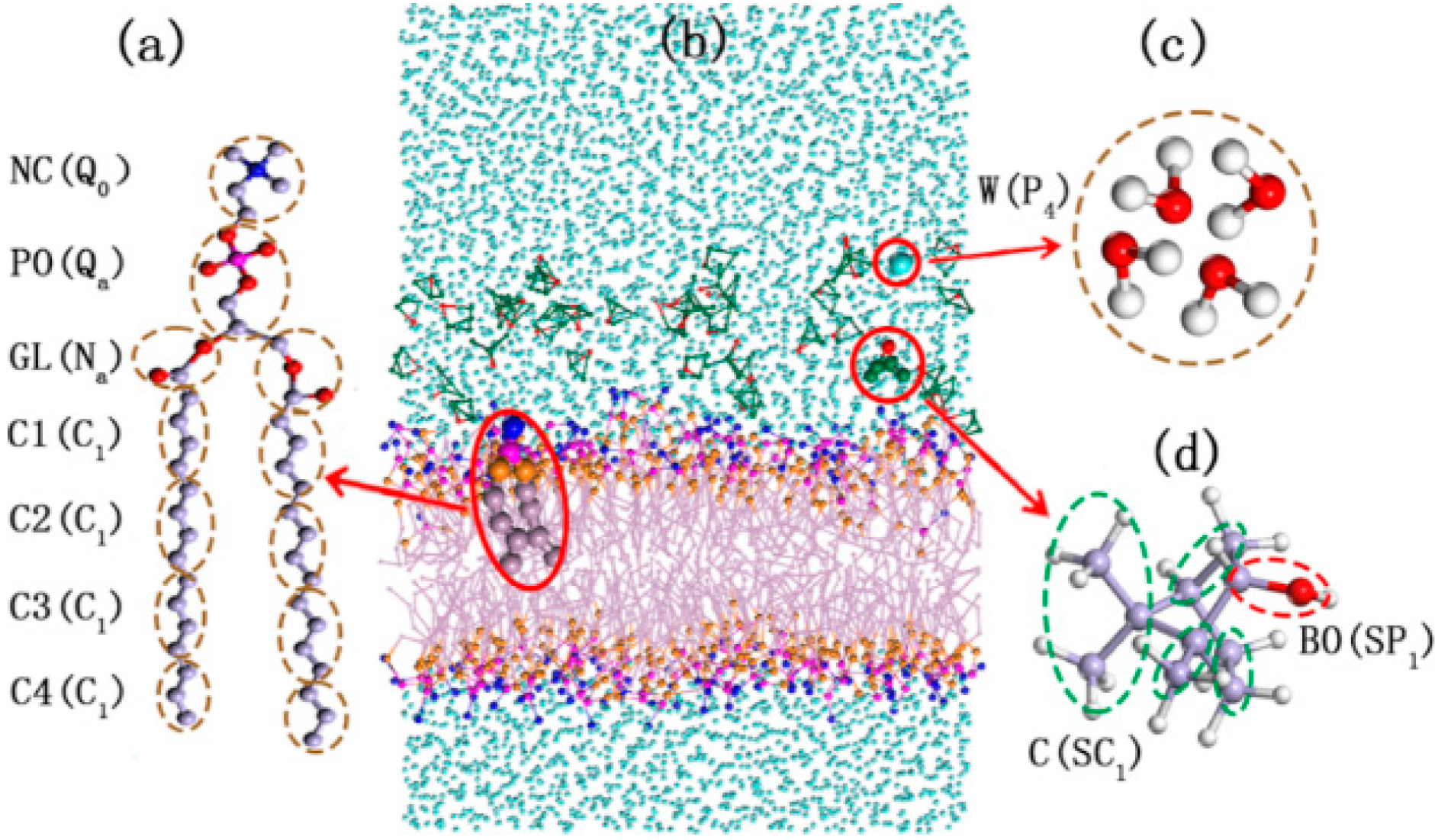
2.1.1. Validity of DPPC (1,2-Dipalmitoylsn-glycero-3-phosphatidylcholine) Model Membrane
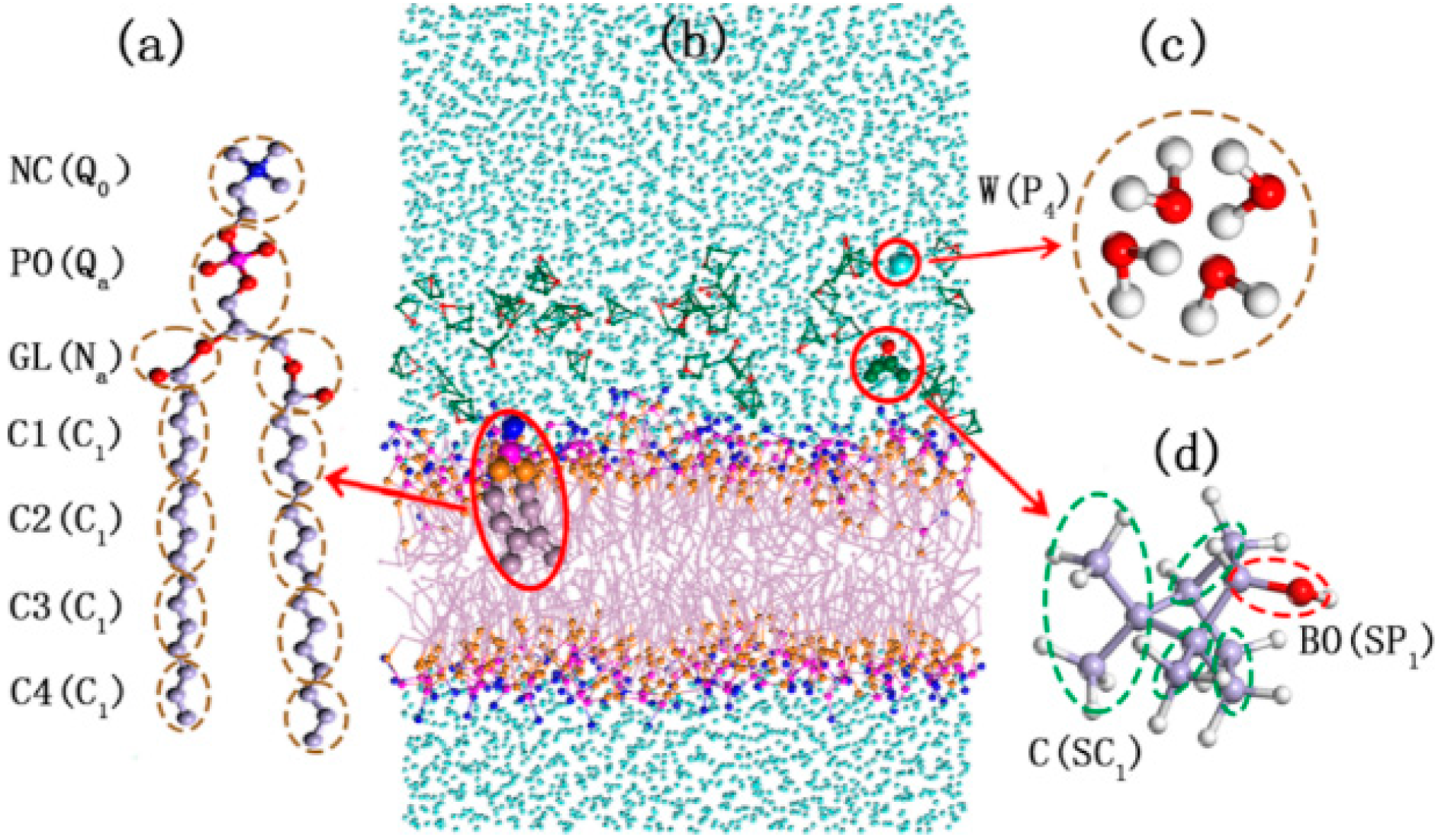
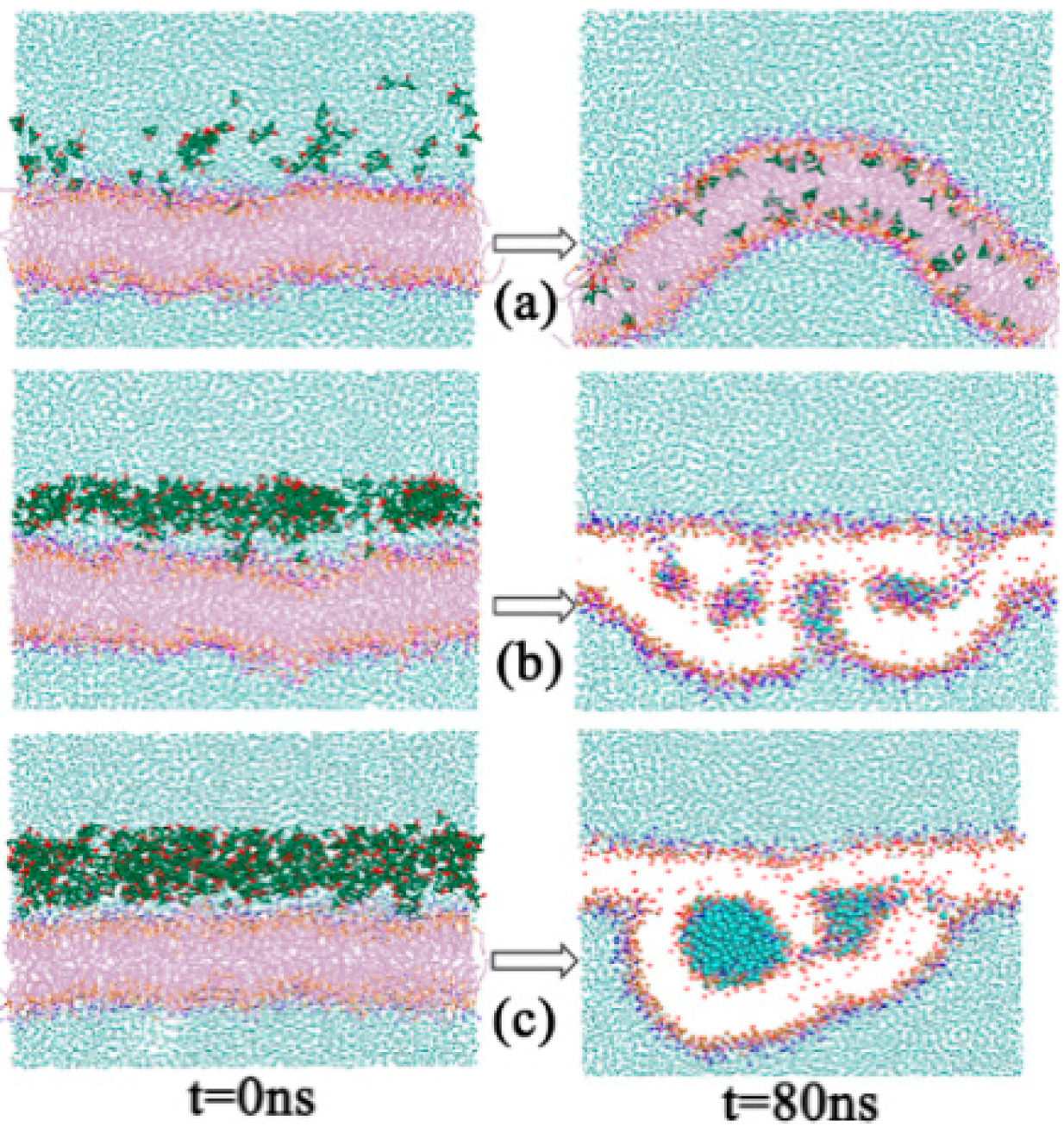
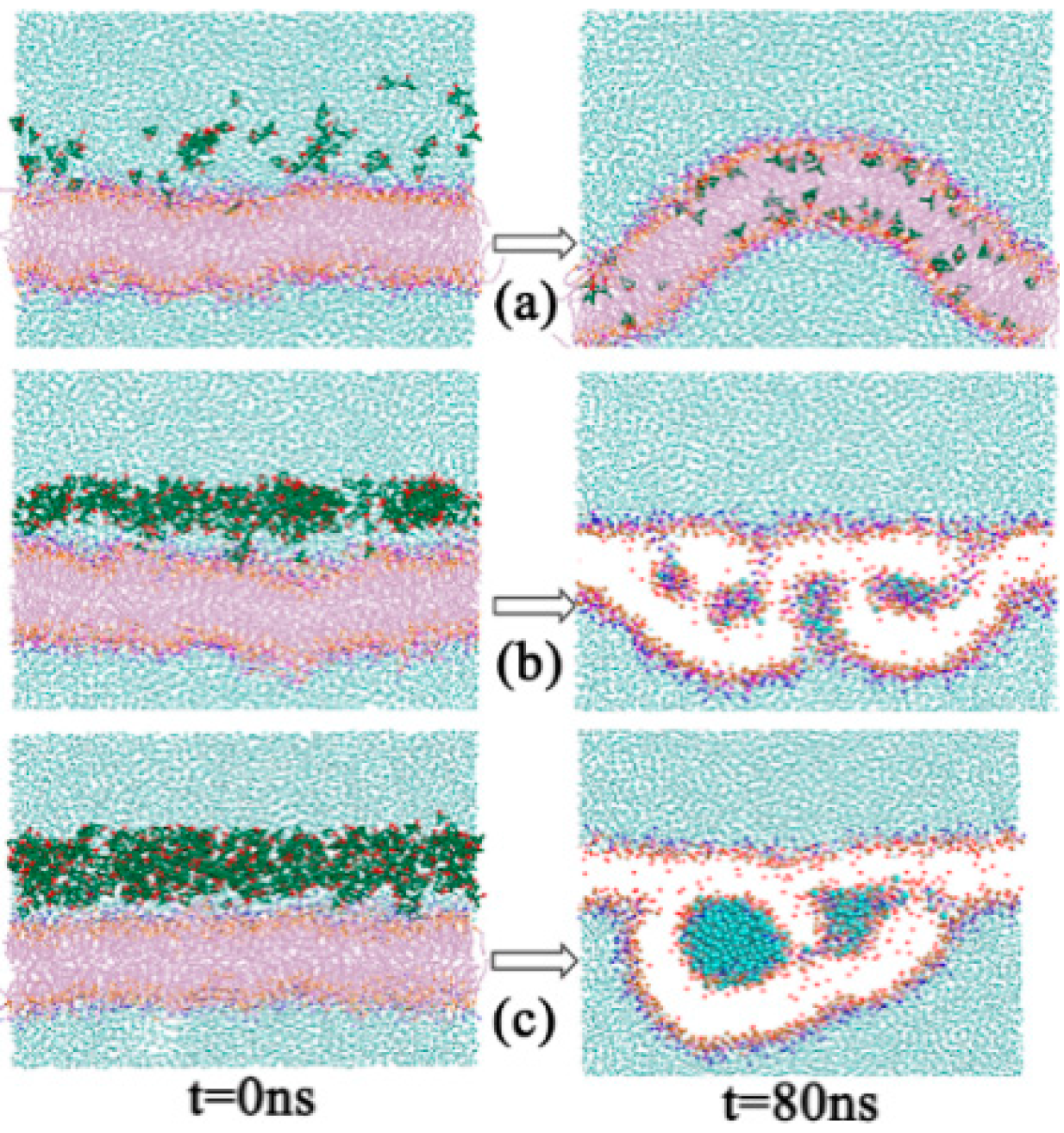
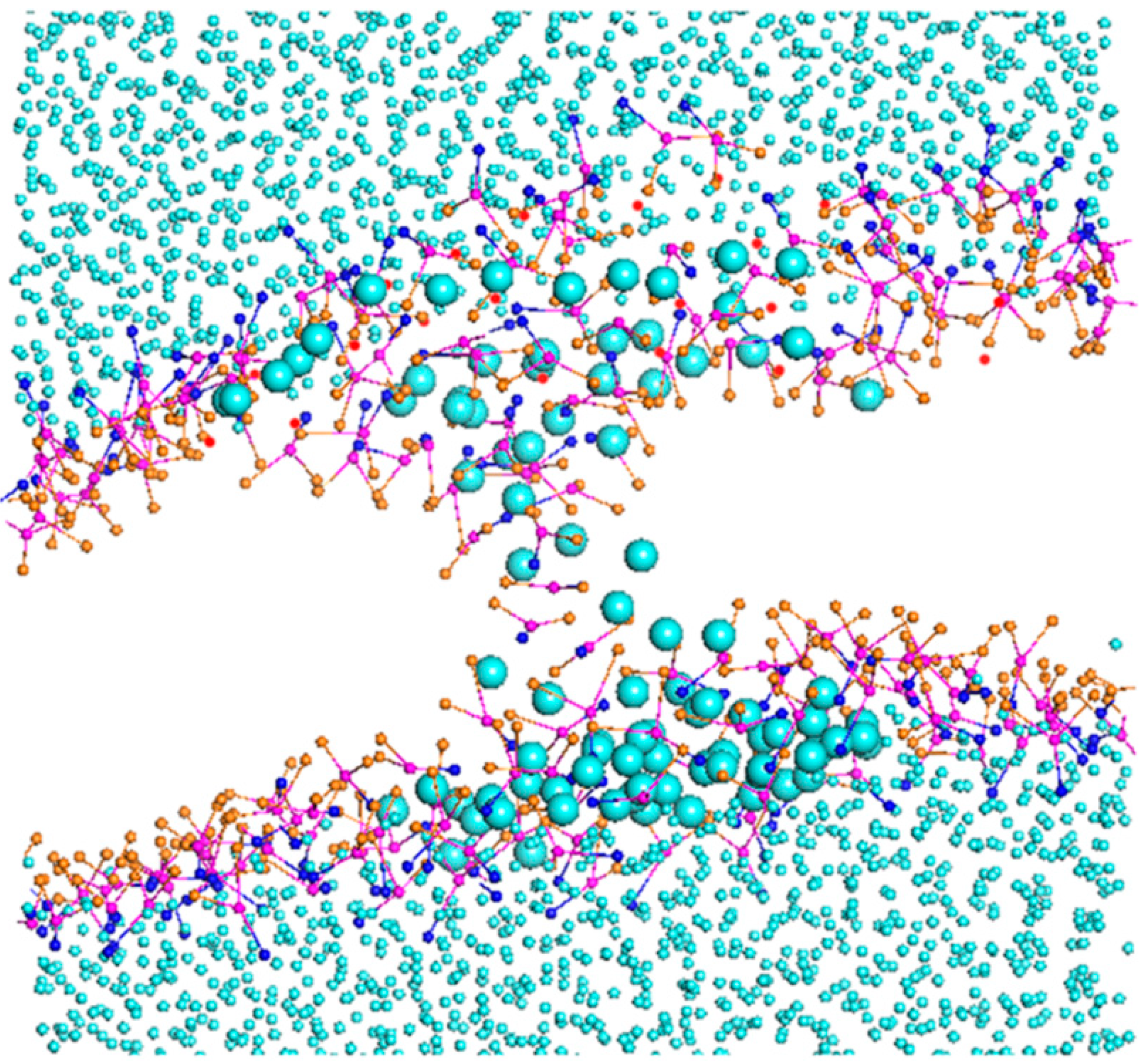
2.1.2. Morphology Perturbation of Bilayer Structure
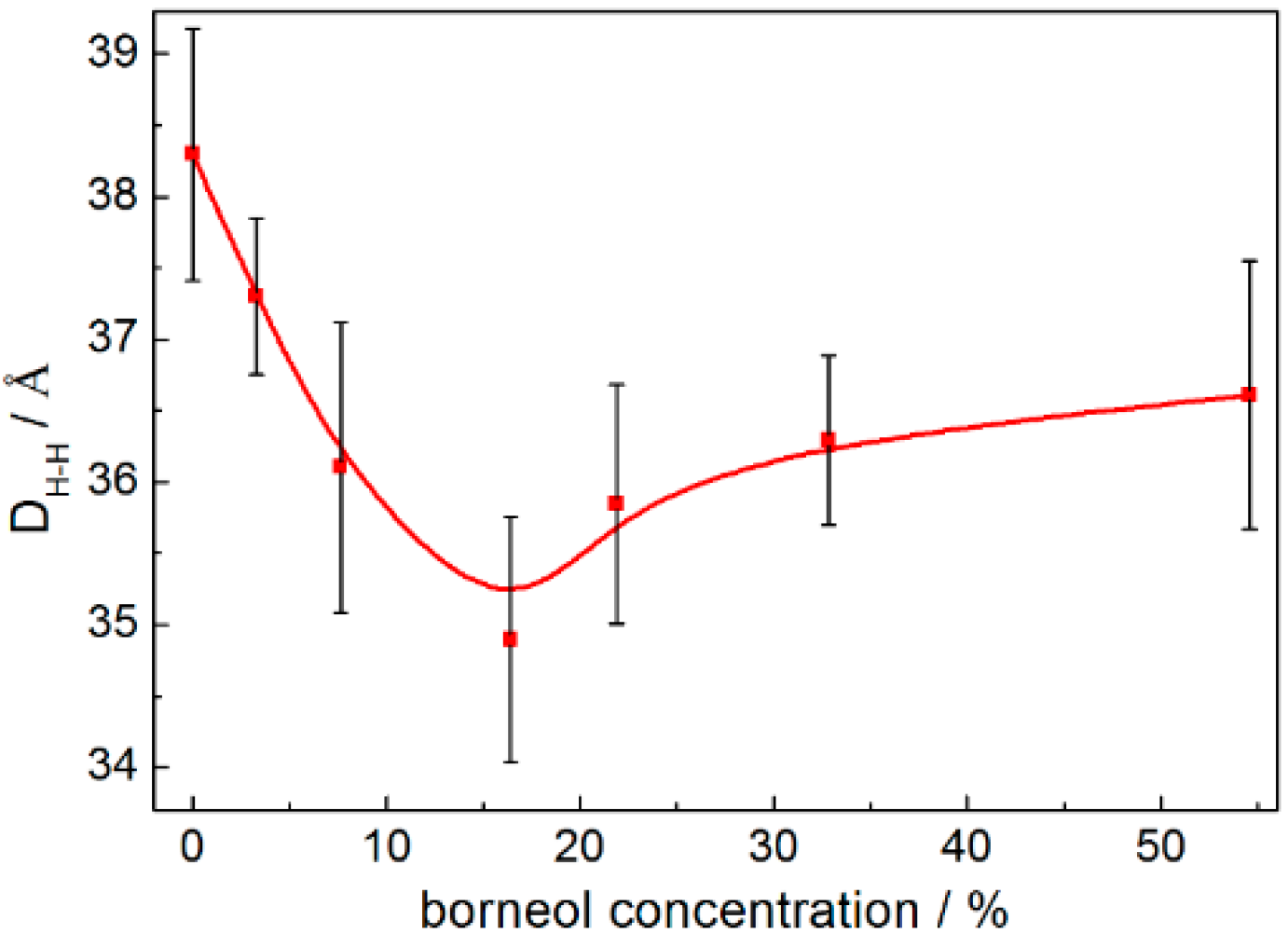


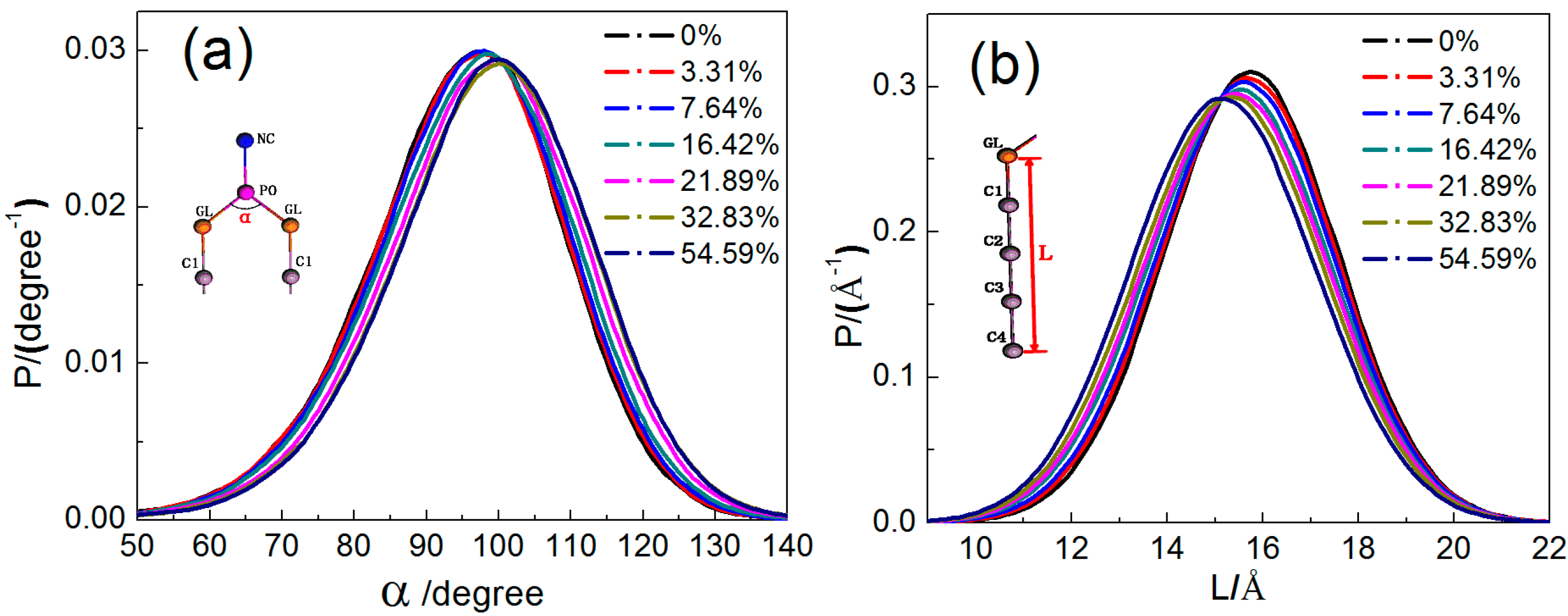
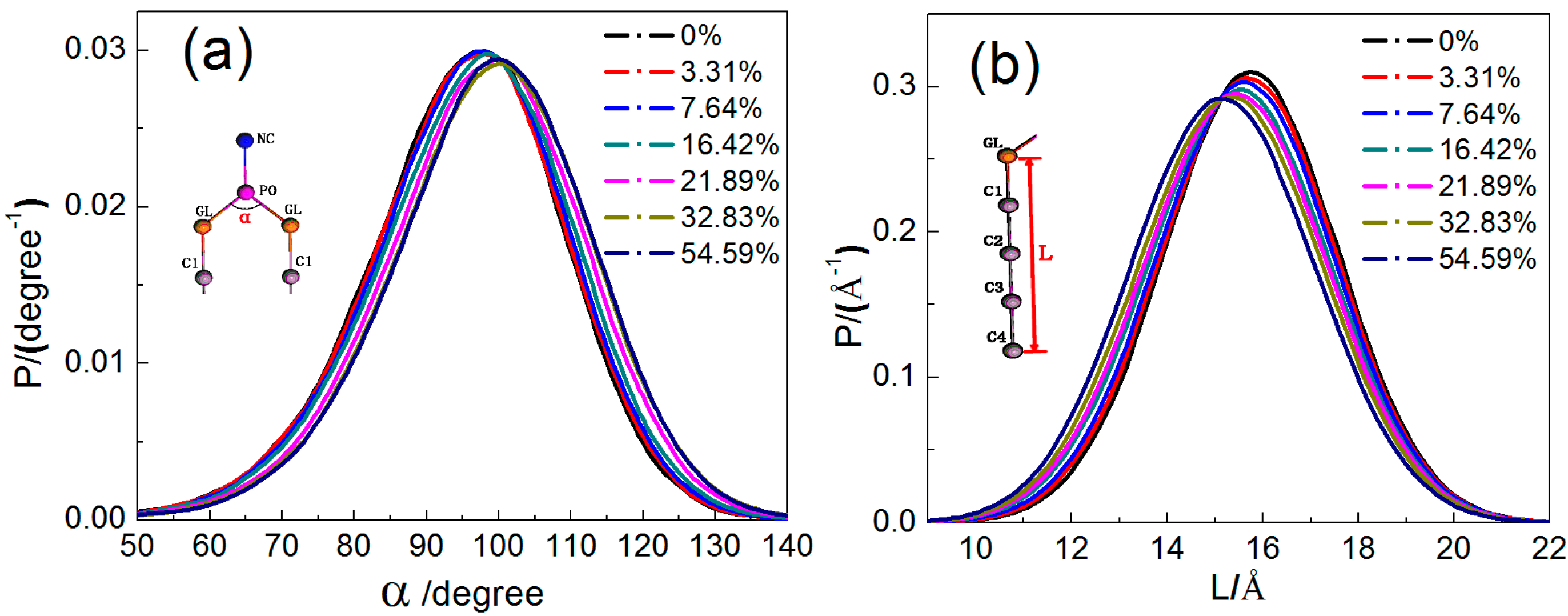
2.1.3. Configuration Changes of DPPC Molecules
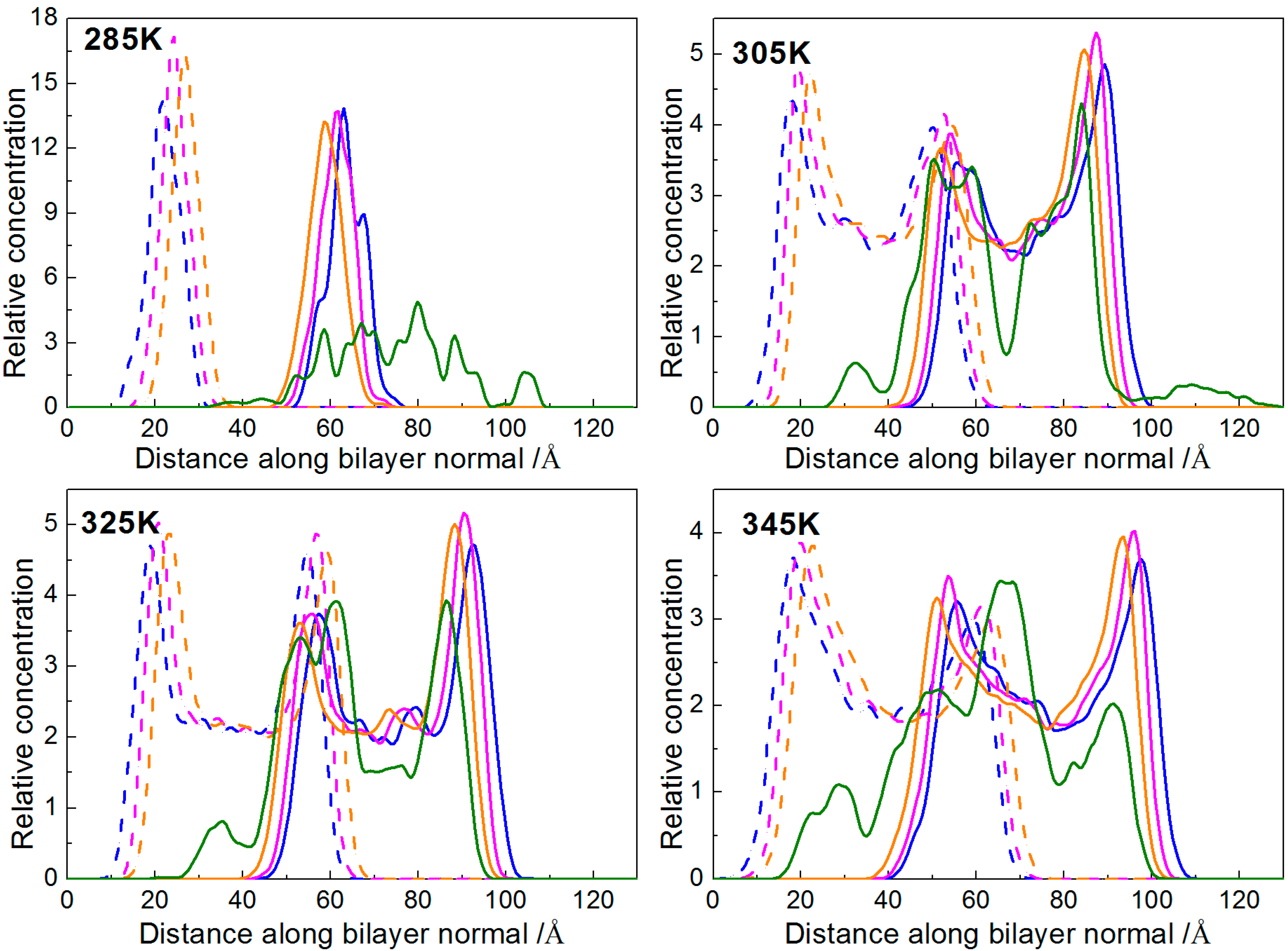
2.1.4. Distribution of Borneol within the DPPC Bilayer
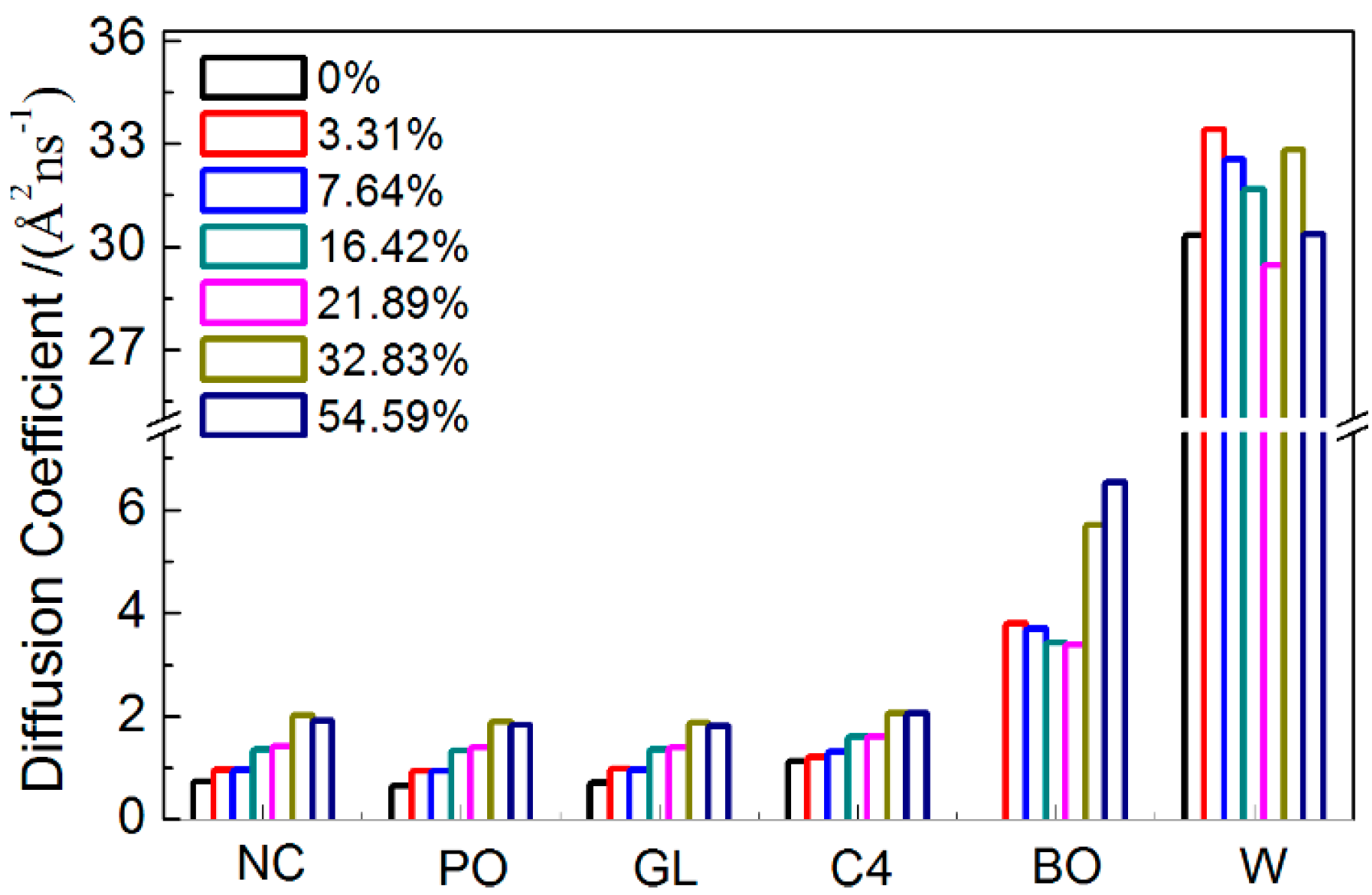
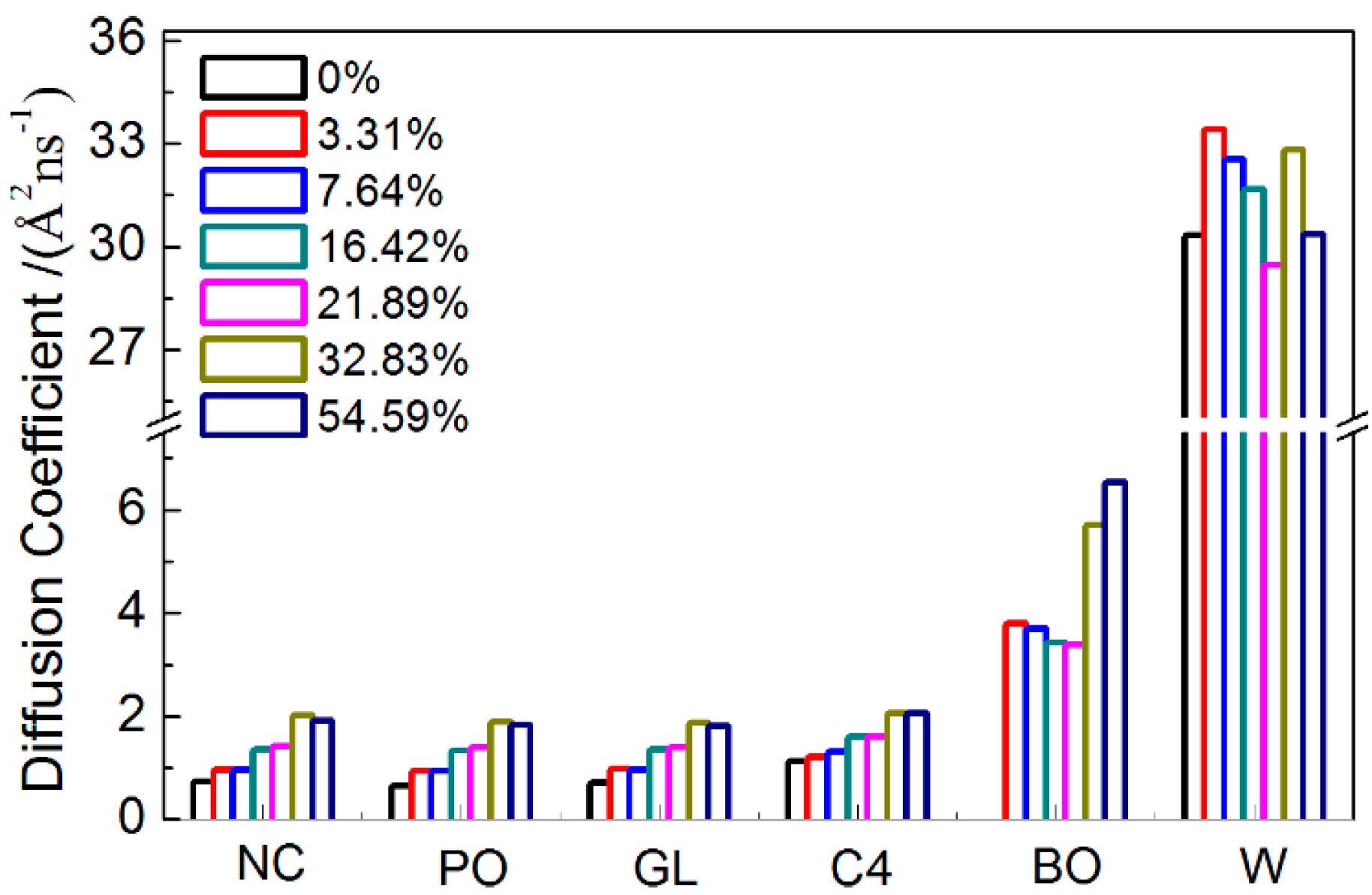
2.1.5. Dynamical Properties


2.1.6. A Temporary Water Pore

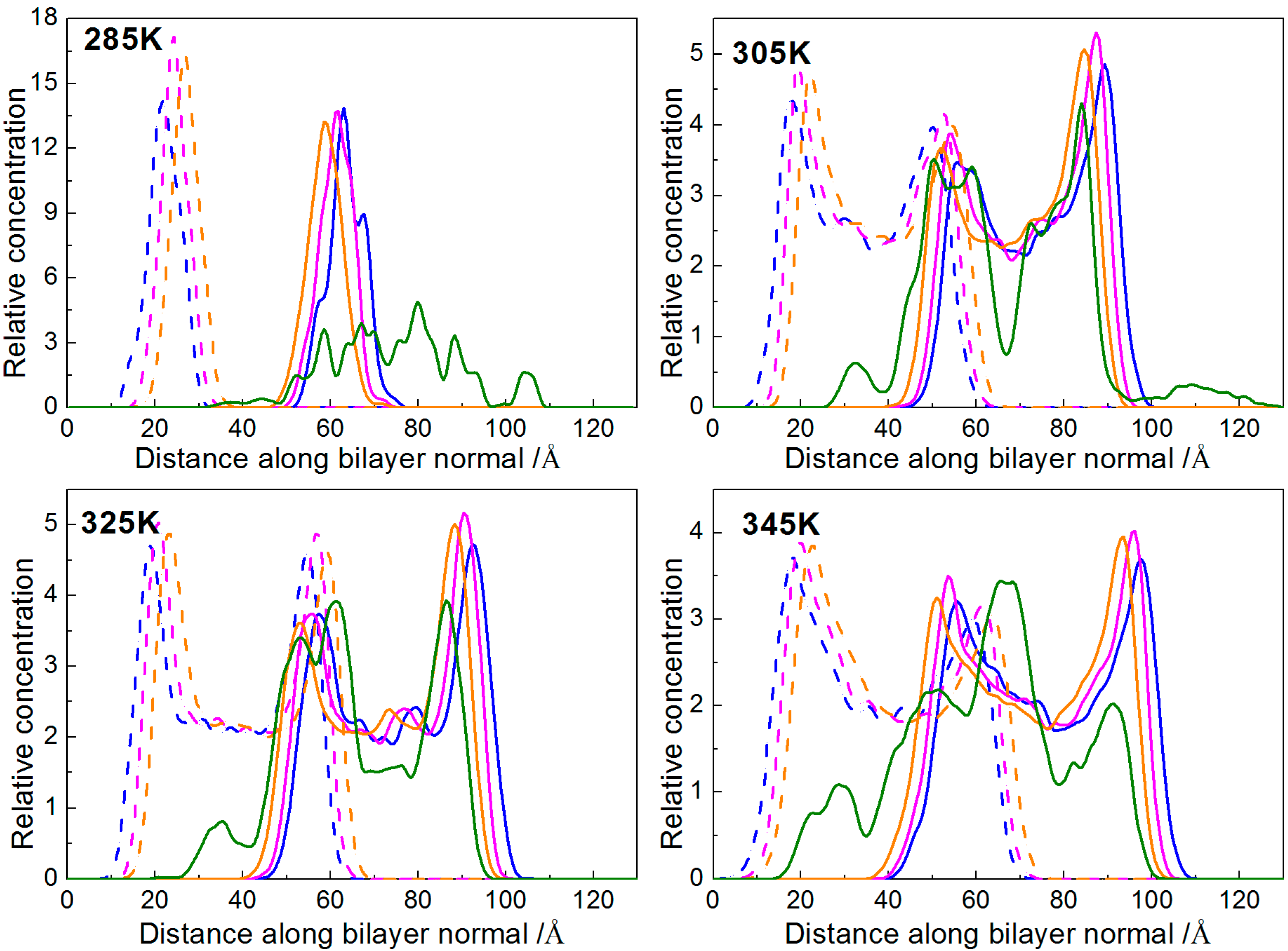
2.2. Temperature Effect
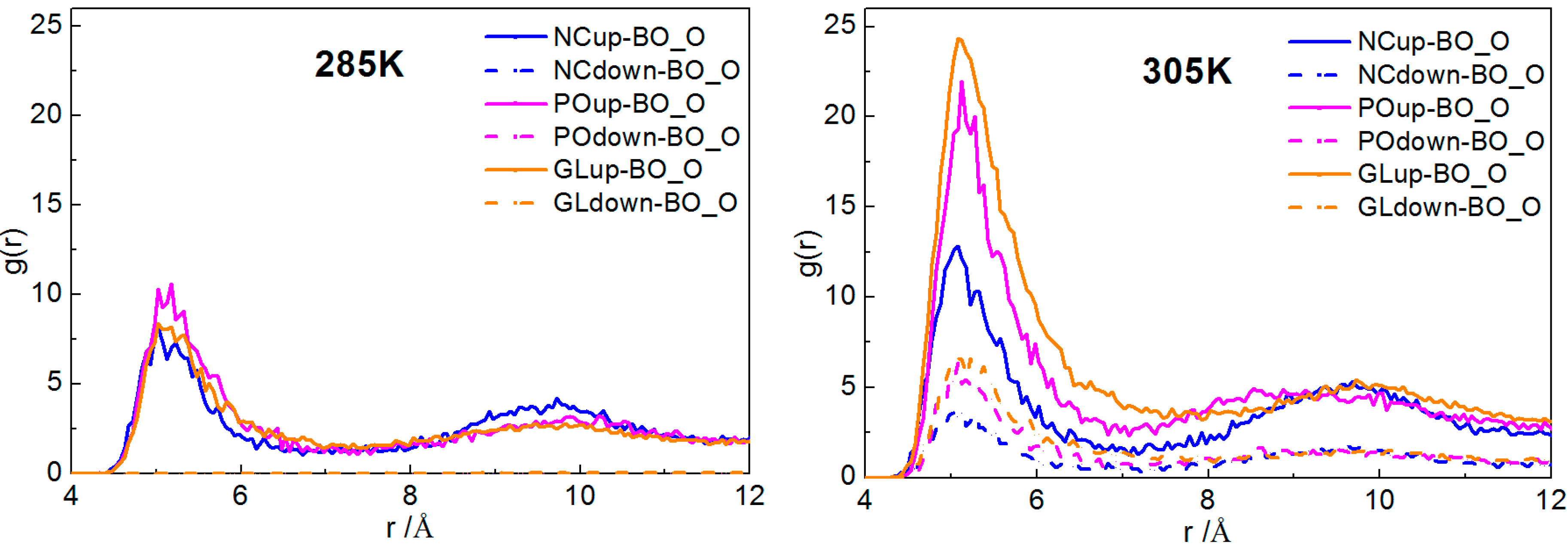
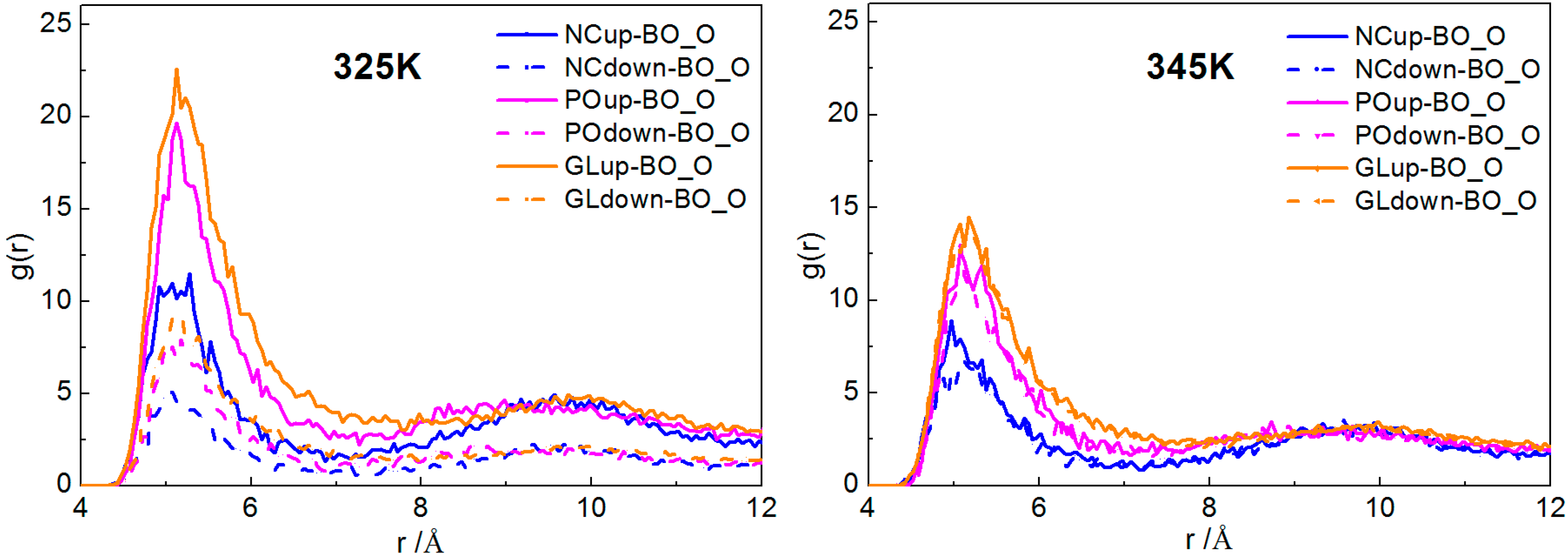
2.2.1. Interactions of Borneol with the DPPC Bilayer

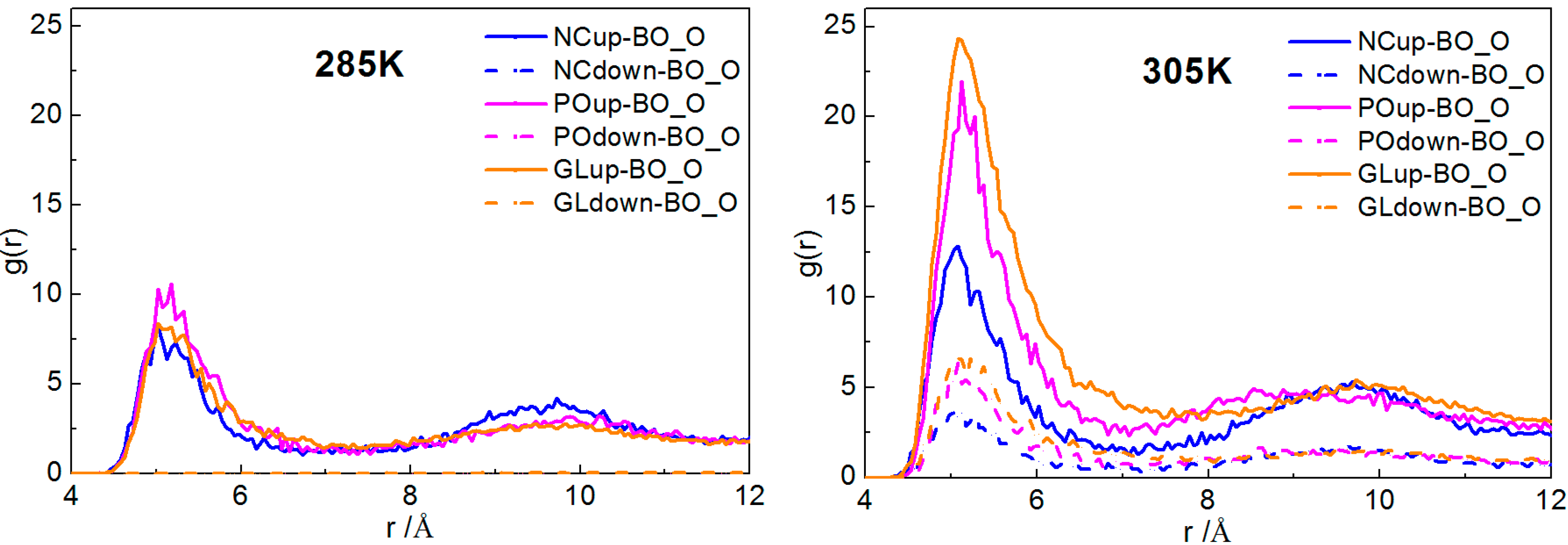
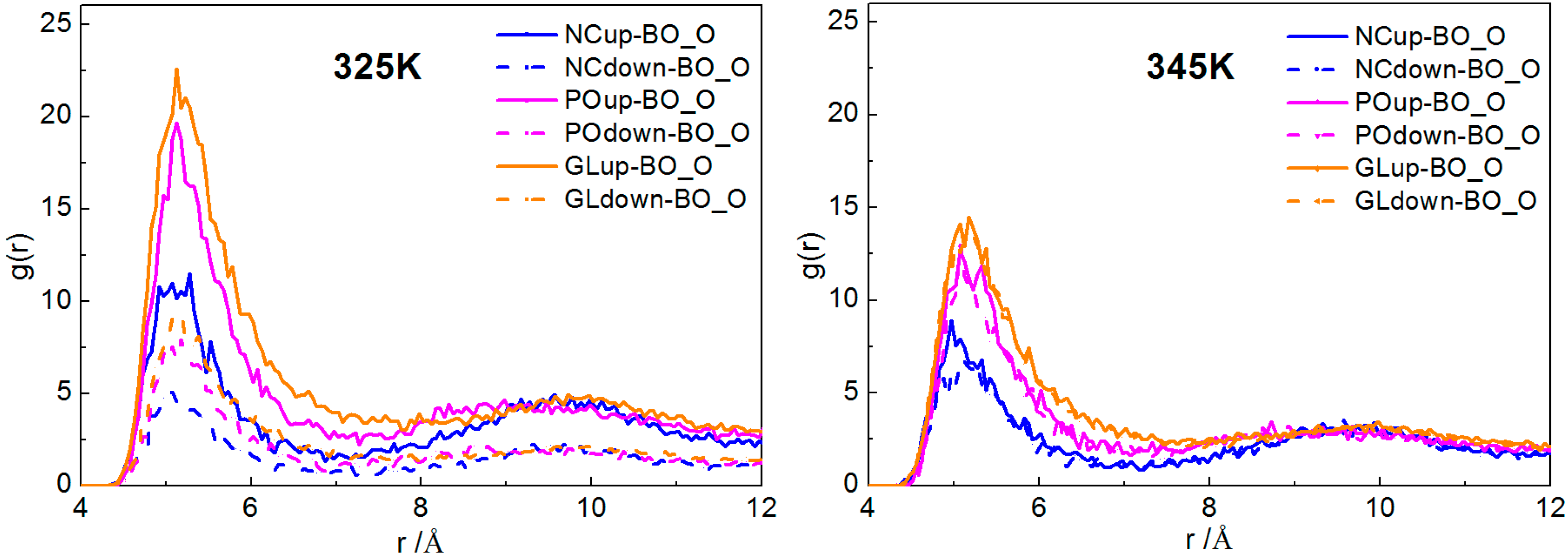

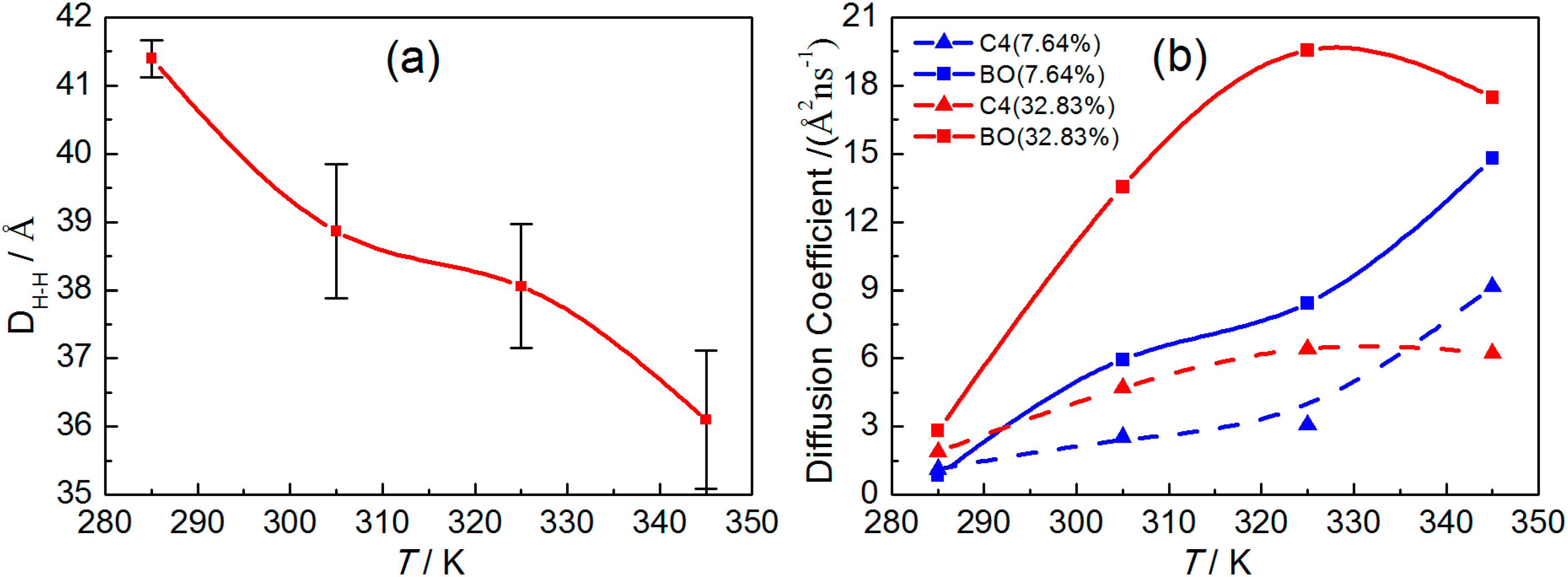
2.2.2. Dynamical Properties
3. Simulation Details

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Systems | DPPC | Bottom Water | Top Water | Extra Water | v% | mol % | |
|---|---|---|---|---|---|---|---|
| Water | Borneol | ||||||
| A | 362 | 3929 | 3929 | 0 | 1100 | 0 | 0 |
| B | 362 | 3929 | 3799 | 26 | 1100 | 3.31 | 2.09 |
| C | 362 | 3929 | 3629 | 60 | 1100 | 7.64 | 4.91 |
| D | 362 | 3929 | 3284 | 129 | 1100 | 16.42 | 10.93 |
| E | 362 | 3929 | 3069 | 172 | 1100 | 21.89 | 14.90 |
| F | 362 | 3929 | 2639 | 258 | 1100 | 32.83 | 23.40 |
| G | 362 | 3929 | 1784 | 429 | 1100 | 54.59 | 42.91 |
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- ChPC. In Chinese Pharmacopoeia; Chinese Medical Science and Technology Press: Beijing, China, 2010.
- Li, D.; Deng, W. Progress in studies on borneol’s penetration enhancing function. Pharmacol. Clin. Chin. Mater. Med. 2007, 23, 87–89. [Google Scholar]
- Qi, H.; Gao, X.; Zhang, L.; Wei, S.-Q.; Bid, S.; Yang, Z.-C.; Cui, H. In vitro evaluation of enhancing effect of borneol on transcorneal permeation of compounds with different hydrophilicities and molecular sizes. Eur. J. Pharmacol. 2013, 705, 20–25. [Google Scholar]
- Song, J.; Bi, H.; Xie, X.; Guo, J.; Wang, X.; Liu, D. Natural borneol enhances geniposide ophthalmic absorption in rabbits. Int. J. Pharm. 2013, 445, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fu, S.; Wei, N.; Hou, Y.; Zhang, X.; Cui, H. The effects of combined menthol and borneol on fluconazole permeation through the corneaex vivo. Eur. J. Pharmacol. 2012, 688, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Lu, Y.; Du, S.; Shang, K.; Cai, C. Influence of borneol and muscone on geniposide transport through MDCK and MDCK-MDR1 cells as blood—Brain barrier in vitro model. Int. J. Pharm. 2013, 456, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Bensikaddour, H.; Snoussi, K.; Lins, L. Interactions of ciprofloxacin with DPPC and DPPG: Fluorescence anisotropy, ATR-FTIR and 31P NMR spectroscopies and conformational analysis. BBA-Biomembranes 2008, 1778, 2535–2543. [Google Scholar] [CrossRef] [PubMed]
- Goksu, E.I.; Vanegas, J.M.; Blanchette, C.D.; Lin, W.; Longo, M.L. AFM for structure and dynamics of biomembranes. BBA-Biomembranes 2009, 1788, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Takajo, Y.; Matsuki, H.; Matsubara, H.; Tsuchiya, K.; Aratono, M.; Yamanaka, M. Structural and morphological transition of long-chain phospholipid vesicles induced by mixing with short-chain phospholipid. Coll. Surf. B 2010, 76, 571–576. [Google Scholar] [CrossRef]
- Smith, A.K.; Freed, J.H. Dynamics and ordering of lipid spin-labels along the coexistence curve of two membrane phases:An ESR study. Chem. Phys. Lipids 2012, 165, 348–361. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.F.; Chen, Y.Y.; Yang, N.N.; Zhu, X.J.; Sun, L.Z.; Li, G.X. Study of the interaction between curcumin and mimetic biomembrane. Sci. China Life Sci. 2012, 55, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Ing, O.; Lfsson, H.I.; Lopez, C.A.; Uusitalo, J.J. The power of coarse graining in biomolecular simulations. WIREs: Comput. Mol. Sci. 2013. [Google Scholar] [CrossRef]
- Marrink, S.J.; Risselada, H.J.; Yefimov, S.; Tieleman, D.P.; de Vries, A.H. The Martini force field: Coarse grained model for biomolecular simulations. J. Phys.Chem. B 2007, 111, 7812–7824. [Google Scholar] [CrossRef] [PubMed]
- Marrink, S.J.; Tieleman, D.P. Perspective on the Martini model. Chem. Soc. Rev. 2013, 42, 6801–6822. [Google Scholar] [CrossRef] [PubMed]
- López, C.A.; Sovova, Z.; van Eerden, F.J.; de Vries, A.H.; Marrink, S.J. Martini force field parameters for glycolipids. J. Chem. Theory Comput. 2013, 9, 1694–1708. [Google Scholar] [CrossRef]
- Yesylevskyy, S.O.; Schafer, L.V.; Sengupta, D.; Marrink, S.J. Polarizable water model for the coarse-grained Martini force field. PLoS Comput. Biol. 2010, 6, e10008106. [Google Scholar] [CrossRef]
- López, C.A.; Rzepiela, A.J.; de Vries, A.H.; Dijkhuizen, L.; Hünenberger, P.H.; Marrink, S.J. Martini coarse-grained force field: Extension to carbohydrates. J. Chem. Theory Comput. 2009, 5, 3195–3210. [Google Scholar] [CrossRef]
- Monticelli, L.; Kandasamy, S.K.; Periole, X.; Larson, R.G.; Tieleman, D.P.; Marrink, S. The Martini coarse-grained force field: Extension to proteins. J. Chem. Theory Comput. 2008, 4, 819–834. [Google Scholar] [CrossRef]
- Hui, G. The Study on the Interaction Between Ginsenoside Molecules and Biomembranes. Ph.D. Thesis, Jilin University, Changchun, China, June 2010. [Google Scholar]
- Nagle, J.F.; Tristram-Nagle, S. Structure of lipid bilayers. Biochim. Biophys. Acta 2000, 1469, 159–195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J-W.; Bian, F-Y.; Shi, G.-J.; Xu, S.-C. Molecular dynamics simulation of dopamine diffusion within and permeation through POPC phospholipid bilayer membrane. Acta Phys. Chim. Sin. 2013, 30, 183–193. [Google Scholar]
- Notman, R.; Anwar, J. Breaching the skin barrier—Insights from molecular simulation of model membranes. Adv. Drug Deliv. Rev. 2013, 65, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Nagle, J.F. Area/lipid of bilayers from NMR. Biophys. J. 1993, 64, 1476–1481. [Google Scholar] [CrossRef] [PubMed]
- Petrache, H.I.; Dodd, S.W.; Brown, M.F. Area per lipid and acyl length distribution in fluid phosphatidylcholines determined by 2H NMR Spectroscopy. Biophys. J. 2000, 79, 3172–3192. [Google Scholar] [CrossRef] [PubMed]
- Gurtovenko, A.A.; Anwar, J. Modulating the structure and properties of cell membranes: The molecular mechanism of action of dimethyl sulfoxide. J. Phys. Chem. B 2007, 111, 10453–10460. [Google Scholar] [CrossRef] [PubMed]
- Ly, H.V.; Longo, M.L. The influence of short-chain alcohols on interfacial tension, mechanical properties, area/molecule, and permeability of fluid lipid bilayers. Biophys. J. 2004, 87, 1013–1033. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Quinn, P.J. Solvation effects of dimethyl sulfoxide on the structure of phospholipid bilayers. Biophys. Chem. 1998, 70, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Gurtovenko, A.A.; Anwar, J. Interaction of ethanol with biological membranes: The formation of non-bilayer structures within the membrane interior and their significance. J. Phys. Chem. B 2009, 113, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Tieleman, D.P.; Marrink, S.J.; Berendsen, H.J.C. A computer perspective of membranes: Molecular dynamics studies of lipid bilayer systems. Biochim. Biophys. Acta 1997, 1331, 235–270. [Google Scholar] [CrossRef] [PubMed]
- Accelrys, I. Materials Studio; Accelrys Software Inc.: San Diego, CA, USA, 2010. [Google Scholar]
- Notman, R.; Noro, M.; O’Malley, B.; Anwar, J. Molecular basis for dimethylsulfoxide (DMSO) action on lipid membranes. J. Am. Chem. Soc. 2006, 128, 13982–13983. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.P.; McDonald, I.R. Theory of Simple Liquids; Academic Press: London, UK, 1990. [Google Scholar]
- Marrink, S.J.; de Vries, A.H.; Mark, A.E. Coarse grained model for semiquantitative lipid simulations. J. Phys. Chem. B 2004, 108, 750–760. [Google Scholar] [CrossRef]
- Baron, R.; de Vries, A.H.; Hünenberger, P.H.; van Gunsteren, W.F. Comparison of atomic-level and coarse-grained models for liquid hydrocarbons from molecular dynamics configurational entropy estimates. J. Phys. Chem. B 2006, 110, 8464–8473. [Google Scholar] [CrossRef] [PubMed]
- Baron, R.; de Vries, A.H.; Hünenberger, P.H.; van Gunsteren, W.F. Configurational entropies of lipids in pure and mixed bilayers from atomic-level and coarse-grained molecular dynamics simulations. J. Phys. Chem. B 2006, 110, 15602–15614. [Google Scholar] [CrossRef] [PubMed]
- Curry, H.B. The method of steepest descent for nonlinear minimization problems. Q. Appl. Math. 1944, 2, 250–261. [Google Scholar]
- Gilbert, J.C.; Nocedal, J. Global convergence properties of conjugate gradient methods for optimization. SIAM J. Optim. 1992, 2, 21–42. [Google Scholar] [CrossRef]
- Schlick, T.; Overton, M. A powerful truncated Newton method for potential energy minimization. J. Comput. Chem. 1987, 8, 1025–1039. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577. [Google Scholar] [CrossRef]
- Ewald, P.P. Die berechnung optischer and elektrostatischer gitterpotentiale. Ann. Phys. 1921, 64, 253–287. [Google Scholar] [CrossRef]
- Steinbach, P.J.; Brooks, B.R. New spherical-cutoff methods for long‐range forces in macromolecular simulation. J. Comput. Chem. 1994, 15, 667–683. [Google Scholar]
- Nosé, S. Constant temperature molecular dynamics methods. Prog. Theor. Phys. Suppl. 1991, 103, 1–46. [Google Scholar] [CrossRef]
- Nosé, S. A molecular dynamics method for simulations in the canonical ensemble. Mol. Phys. 1984, 52, 255–268. [Google Scholar] [CrossRef]
- Nosé, S. A unified formulation of the constant temperature molecular dynamics methods. J. Chem. Phys. 1984, 81, 511–519. [Google Scholar] [CrossRef]
- Mou, J.; Yang, J.; Huang, C.; Shao, Z. Alcohol induces tnterdigitated domains in unilamellar phosphatidylcholine bilayers. Biochemistry 1994, 33, 9981–9985. [Google Scholar] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, Q.; Shi, X.; Ding, H.; Dai, X.; Wan, G.; Qiao, Y. Interactions of Borneol with DPPC Phospholipid Membranes: A Molecular Dynamics Simulation Study. Int. J. Mol. Sci. 2014, 15, 20365-20381. https://doi.org/10.3390/ijms151120365
Yin Q, Shi X, Ding H, Dai X, Wan G, Qiao Y. Interactions of Borneol with DPPC Phospholipid Membranes: A Molecular Dynamics Simulation Study. International Journal of Molecular Sciences. 2014; 15(11):20365-20381. https://doi.org/10.3390/ijms151120365
Chicago/Turabian StyleYin, Qianqian, Xinyuan Shi, Haiou Ding, Xingxing Dai, Guang Wan, and Yanjiang Qiao. 2014. "Interactions of Borneol with DPPC Phospholipid Membranes: A Molecular Dynamics Simulation Study" International Journal of Molecular Sciences 15, no. 11: 20365-20381. https://doi.org/10.3390/ijms151120365
APA StyleYin, Q., Shi, X., Ding, H., Dai, X., Wan, G., & Qiao, Y. (2014). Interactions of Borneol with DPPC Phospholipid Membranes: A Molecular Dynamics Simulation Study. International Journal of Molecular Sciences, 15(11), 20365-20381. https://doi.org/10.3390/ijms151120365