Genomic and Phenotypic Alterations of the Neuronal-Like Cells Derived from Human Embryonal Carcinoma Stem Cells (NT2) Caused by Exposure to Organophosphorus Compounds Paraoxon and Mipafox


Abstract
:1. Introduction
2. Results
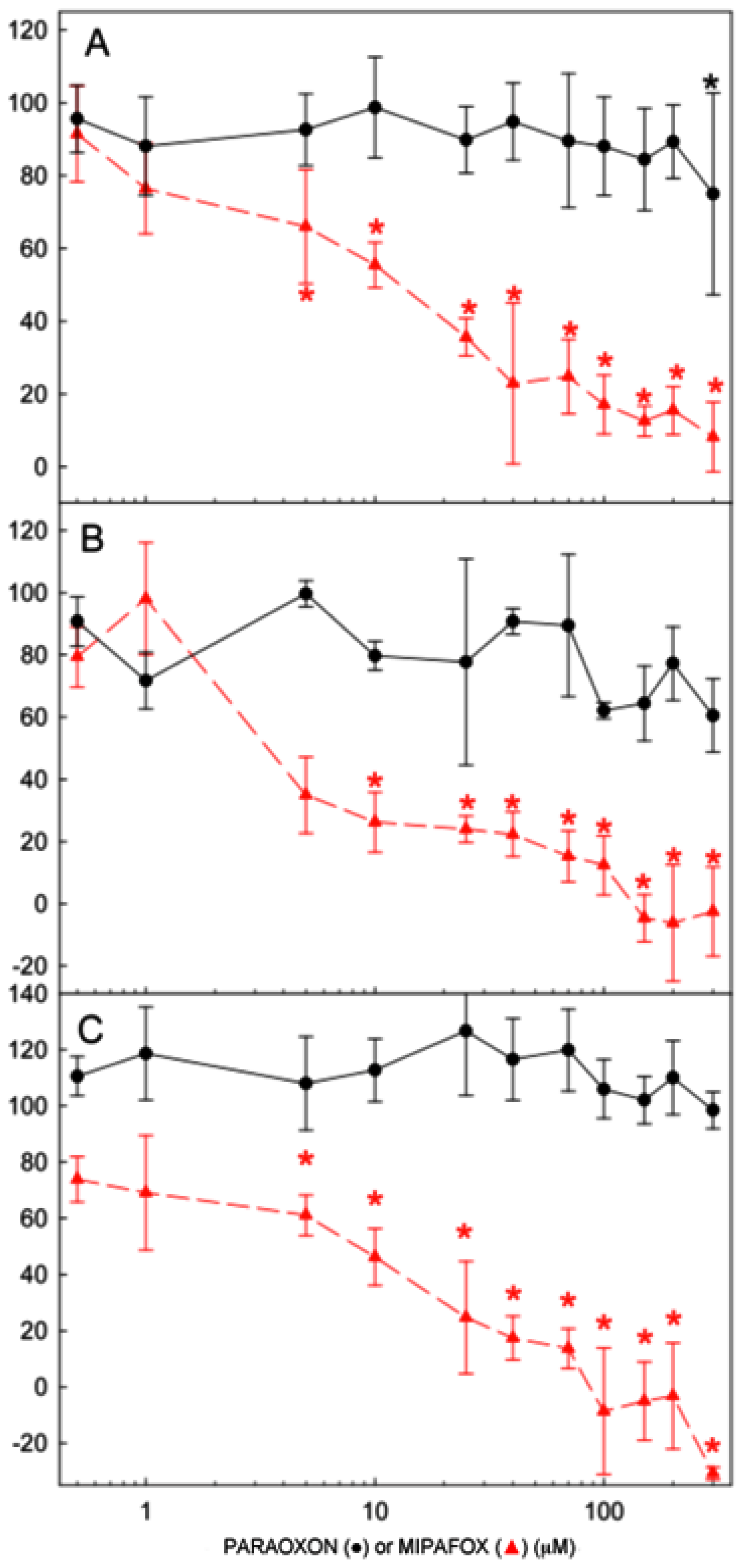
2.1. Cell Viability after Exposure to OPs
2.2. Effect of Paraoxon and Mipafox on NTE Activity
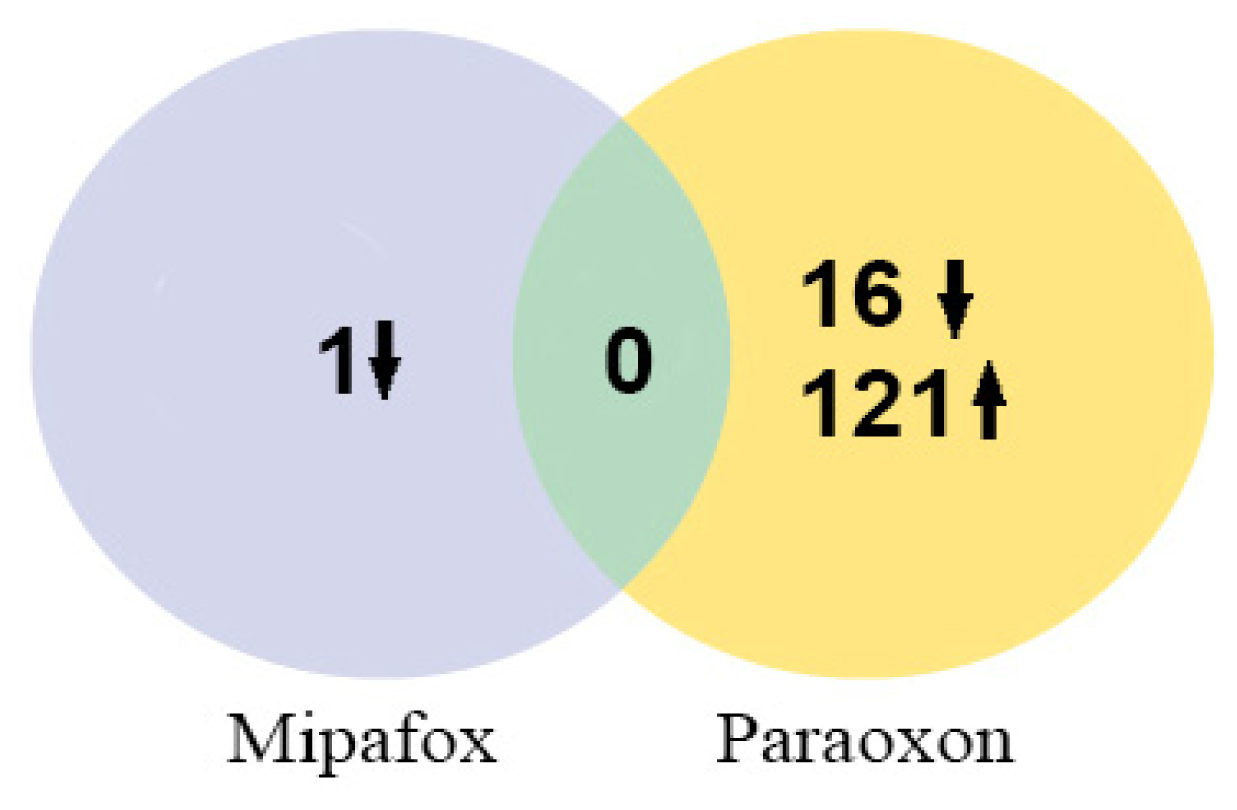
2.3. Microarray Analysis after 4-Day Exposure
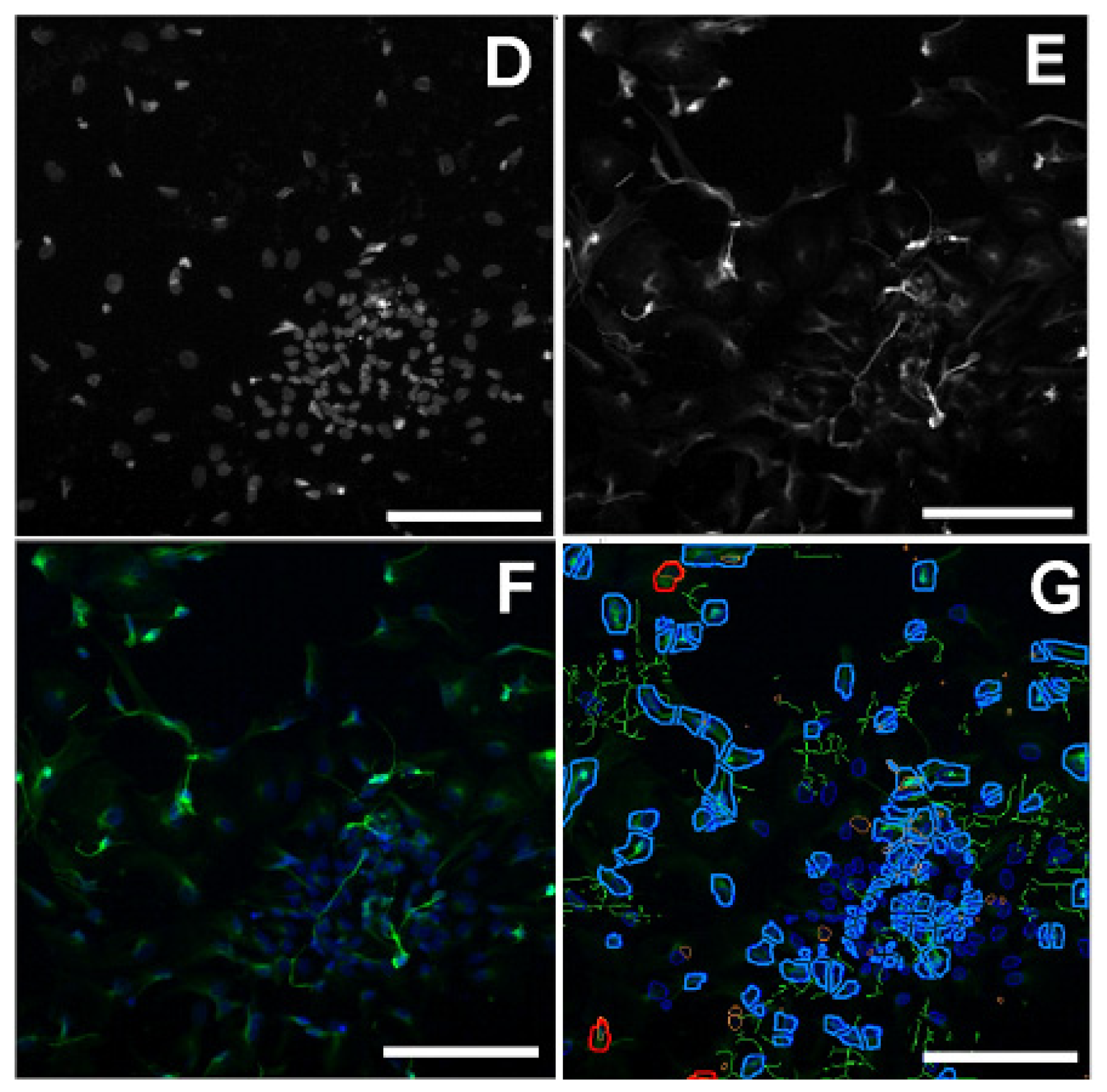
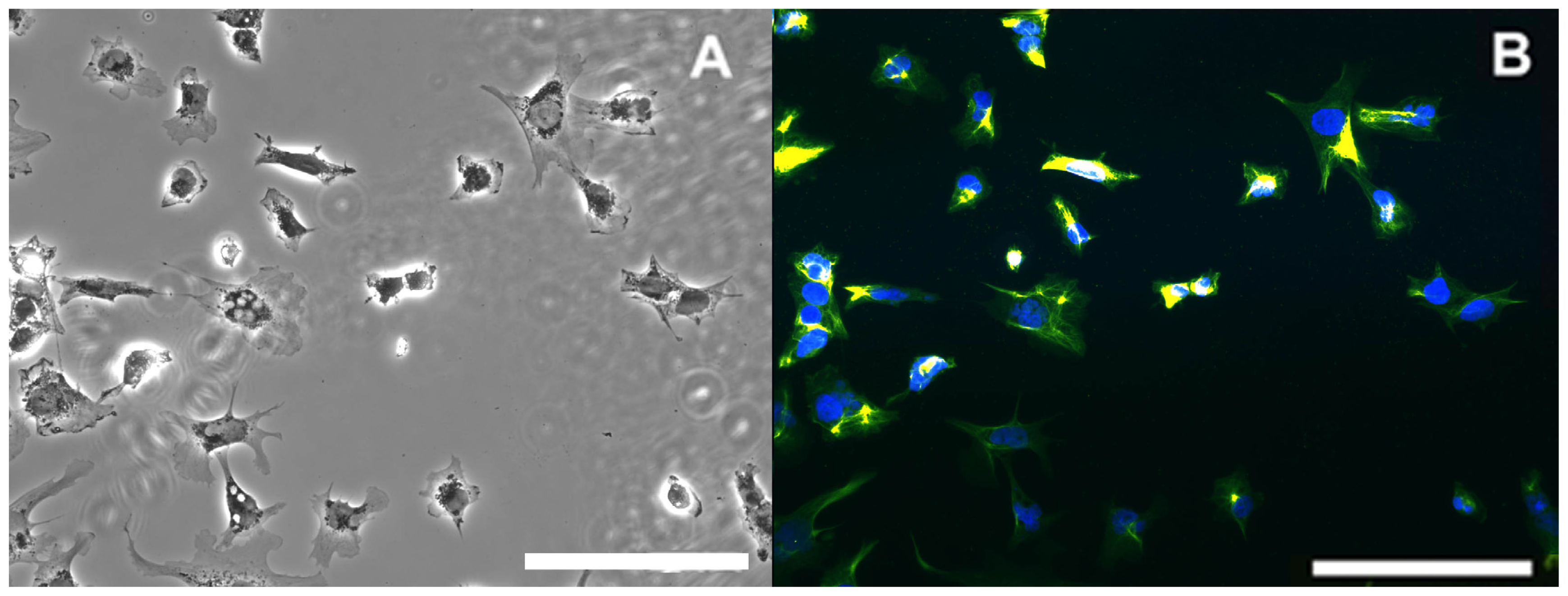
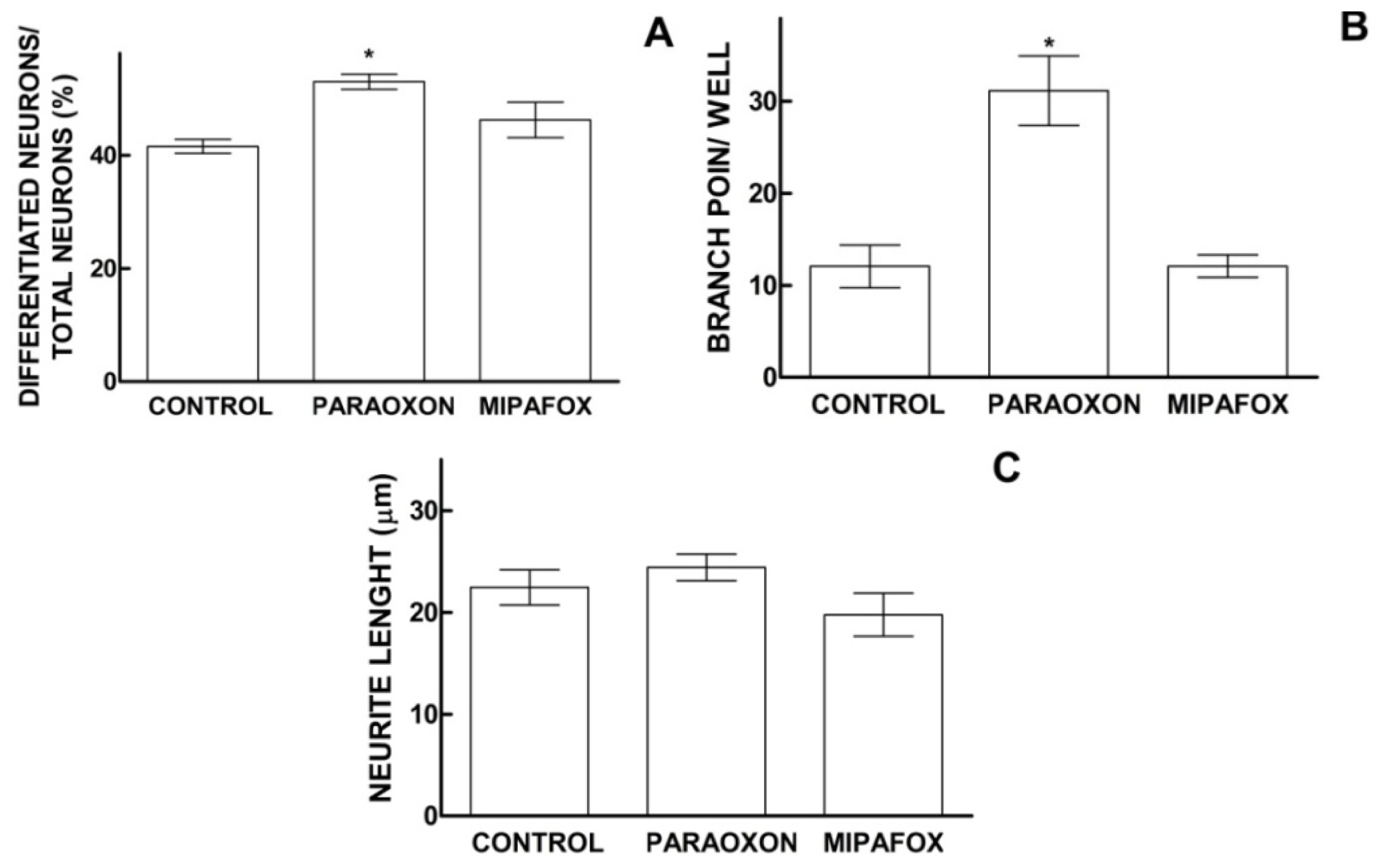
2.4. Effect of Paraoxon and Mipafox on the Morphology of NT2-Derived Neurons
3. Discussion
3.1. Effects of Mipafox during NT2 Neurodifferentiation
3.2. Effects of Paraoxon during Initial Stage of NT2 Neurodifferentiation
4. Material and Methods
4.1. Chemicals
4.2. NT2 Cell Differentiation into Neuronal-like Cells
4.3. Assessment of Cell Viability Using the MTT Assay
4.4. NTE Enzymatic Activity
4.5. Microarray Studies
4.6. Evaluation of Cell Morphology Using Fluorescence Imaging
5. Conclusions
Supplementary Information
ijms-15-00905-s001.pdfAcknowledgments
Conflicts of Interest
Abbreviations and organophosphorus compounds common names
| AChE | Acetylcholinesterase |
| DAPI | 4-6-diamidino-2-phenylindole |
| FBS | foetal bovine serum |
| FGFR1 | Fibroblasts growth factor receptor 1 |
| GFAP | Glial fibrillary acidic protein |
| NT2 | human-derived NTera2/D1 cells |
| MAP2 | Microtubule-associated protein 2 |
| Mipafox | N,N-diisopropyl diamidophosphorofluoridate |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NCBI | National Center for Biotechnology Information |
| NTE | neuropathy target esterase |
| OPs | organophosphorus compounds |
| Paraoxon | O,O′-diethyl p-nitrophenyl phosphate |
| PBS | phosphate buffered saline |
| PV | phenyl valerate |
| RA | retinoic acid |
| SEM | standard error of the mean |
| YAP1 | Yes-associated protein 1 |
References
- Business Analytic Center, Insecticides: European Union Market Outlook 2011 and Forecast till 2016; PRNewswire: New York, NY, USA, 2011.
- Bronstein, A.C.; Spyker, D.A.; Cantilena, L.R., Jr.; Green, J.L.; Rumack, B.H.; Dart, R.C. Annual Report of the American Association of Poison Control Centers’ National Poison Data System (NPDS): 28th Annual Report. Clin. Toxicol 49, 910–941.
- Bjørling-Poulsen, M.; Andersen, H.R.; Grandjean, P. Potential developmental neurotoxicity of pesticides used in Europe. Environ. Health 2008, 7, 50. [Google Scholar] [Green Version]
- Grandjean, P.; Landrigan, P.J. Developmental neurotoxicity of industrial chemicals. Lancet 2006, 368, 2167–2178. [Google Scholar]
- Hogberg, H.T.; Kinsner-Ovaskainen, A.; Hartung, T.; Coecke, S.; Bal-Price, A.K. Gene expression as a sensitive endpoint to evaluate cell differentiation and maturation of the developing central nervous system in primary cultures of rat cerebellar granule cells (CGCs) exposed to pesticides. Toxicol. Appl. Pharmacol 2009, 235, 268–286. [Google Scholar]
- Rohlman, D.S.; Arcury, T.A.; Quandt, S.A.; Lasarev, M.; Rothlein, J.; Travers, R.; Tamulinas, A.; Scherer, J.; Early, J.; Marín, A.; et al. Neurobehavioral performance in preschool children from agricultural and non-agricultural communities in Oregon and North Carolina. Neurotoxicology 2005, 26, 589–598. [Google Scholar]
- Grandjean, P.; Harari, R.; Barr, D.B.; Debes, F. Pesticide exposure and stunting as independent predictors of neurobehavioral deficits in Ecuadorian school children. Pediatrics 2006, 117, e546–e556. [Google Scholar]
- World Health Organization. Organophosphorus Insecticides: A general introduction. Environmental Health Criteria; World Health Organization, 1986; 63. Available online: http://www.inchem.org/documents/ehc/ehc/ehc63.htm (accessed on 26 December 2013).
- Glynn, P. NTE: One target protein for different toxic syndromes with distinct mechanisms? Bioessays 2003, 25, 742–745. [Google Scholar]
- Roldán-Tapia, L.; Nieto-Escámez, F.A.; del Águila, E.M.; Laynez, F.; Parrón, T.; Sánchez-Santed, F. Neuropsychological sequelae from acute poisoning and long-term exposure to carbamate and organophosphate pesticides. Neurotoxicol. Teratol 2006, 28, 694–703. [Google Scholar]
- Mackenzie Ross, S.J.; Brewin, C.R.; Curran, H.V.; Furlong, C.E.; Abraham-Smith, K.M.; Harrison, V. Neuropsychological and psychiatric functioning in sheep farmers exposed to low levels of organophosphate pesticides. Neurotoxicol. Teratol 2010, 32, 452–459. [Google Scholar]
- Parrón, T.; Requena, M.; Hernández, A.F.; Alarcón, R. Association between environmental exposure to pesticides and neurodegenerative diseases. Toxicol. Appl. Pharmacol 2011, 256, 379–385. [Google Scholar]
- Gupta, R.C.; Malik, J.K.; Milatovic, D. Organophosphate and Carbamate Pesticides. In Reproductive and Developmental Toxicology; Gupta, R.C., Ed.; Elsevier: London, UK, 2011; pp. 471–486. [Google Scholar]
- Flaskos, J. The developmental neurotoxicity of organophosphorus insecticides: A direct role for the oxon metabolites. Toxicol. Lett 2012, 209, 86–93. [Google Scholar]
- Lauder, J.M.; Schambra, U.B. Morphogenetic roles of acetylcholine. Environ. Health Perspect 1999, 107, 65–69. [Google Scholar]
- Johnson, F.O.; Chambers, J.E.; Nail, C.A.; Givaruangsawat, S.; Carr, R.L. Developmental chlorpyrifos and methyl parathion exposure alters radial-arm maze performance in juvenile and adult rats. Toxicol. Sci 2009, 109, 132–142. [Google Scholar]
- Moreira, E.G.; Yu, X.; Robinson, J.F.; Griffith, W.; Hong, S.W.; Beyer, R.P.; Bammler, T.K.; Faustman, E.M. Toxicogenomic profiling in maternal and fetal rodent brains following gestational exposure to chlorpyrifos. Toxicol. Appl. Pharmacol 2010, 245, 310–325. [Google Scholar]
- Estevan, C.; Vilanova, E.; Sogorb, M.A. Chlorpyrifos and its metabolites alter gene expression at non-cytotoxic concentrations in D3 mouse embryonic stem cells under in vitro differentiation: Considerations for embryotoxic risk assessment. Toxicol. Lett 2012, 217, 14–22. [Google Scholar]
- Guillette, E.A.; Meza, M.M.; Aquilar, M.G.; Soto, A.D.; Garcia, I.E. An anthropological approach to the evaluation of preschool children exposed to pesticides in Mexico. Environ. Health Perspect 1998, 106, 347–353. [Google Scholar]
- Hogberg, H.T.; Sobanski, T.; Novellino, A.; Whelan, M.; Weiss, D.G.; Bal-Price, A.K. Application of micro-electrode arrays (MEAs) as an emerging technology for developmental neurotoxicity: Evaluation of domoic acid-induced effects in primary cultures of rat cortical neurons. Neurotoxicology 2011, 32, 158–168. [Google Scholar]
- Bal-Price, A.K.; Hogberg, H.T.; Buzanska, L.; Lenas, P.; van Vliet, E.; Hartung, T. In vitro developmental neurotoxicity (DNT) testing: Relevant models and endpoints. Neurotoxicology 2009, 31, 545–554. [Google Scholar]
- Coyle, D.E.; Li, J.; Baccei, M. Regional differentiation of retinoic acid-induced human pluripotent embryonic carcinoma stem cell neurons. PLoS One 2011, 6, e16174. [Google Scholar]
- Andrews, P.W. From teratocarcinomas to embryonic stem cells. Philos. Trans. R. Soc. Lond. B 2002, 357, 405–417. [Google Scholar]
- Tegenge, M.A.; Roloff, F.M.; Bicker, G. Rapid differentiation of human embryonal carcinoma stem cells (NT2) into neurons for neurite outgrowth analysis. Cell. Mol. Neurobiol 2011, 31, 635–643. [Google Scholar]
- Stummann, T.C.; Hareng, L.; Bremer, S. Hazard assessment of methylmercury toxicity to neuronal induction in embryogenesis using human embryonic stem cells. Toxicology 2009, 257, 117–126. [Google Scholar]
- Buzanska, L.; Sypecka, J.; Nerini-Molteni, S.; Compagnoni, A.; Hogberg, H.T.; del Torchio, R.; Domanska-Janik, K.; Zimmer, J.; Coecke, S. A human stem cell-based model for identifying adverse effects of organic and inorganic chemicals on the developing nervous system. Stem Cells 2009, 27, 2591–2601. [Google Scholar]
- Slotkin, T.; Seidler, F. Transcriptional profiles reveal similarities and differences in the effects of developmental neurotoxicants on differentiation into neurotransmitter phenotypes in PC12 cells. Brain Res. Bull 2009, 78, 211–22530. [Google Scholar]
- Camon, E.; Barrell, D.; Lee, V.; Dimmer, E.; Apweiler, R. The Gene Ontology Annotation (GOA) Database—An integrated resource of GO annotations to the UniProt Knowledgebase. In Silico Biol 2004, 4, 5–6. [Google Scholar]
- Crumpton, T.L.; Seidler, F.J.; Slotkin, T.A. Developmental neurotoxicity of chlorpyrifos in vivo and in vitro: Effects on nuclear transcription factors involved in cell replication and differentiation. Brain. Res 2000, 857, 87–98. [Google Scholar]
- Ehrich, M.; Intropido, L.; Costa, L.G. Interaction of organophosphorus compounds with muscarinic receptors in SH-SY5Y human neuroblastoma cells. J. Toxicol. Environ. Health 1994, 43, 51–63. [Google Scholar]
- Garcia, S.J.; Seidler, F.J.; Crumpton, T.L.; Slotkin, T.A. Does the developmental neurotoxicity of chlorpyrifos involve glial targets? Macromolecule synthesis, adenylyl cyclase signaling, nuclear transcription factors, and formation of reactive oxygen in C6 glioma cells. Brain Res 2001, 891, 54–68. [Google Scholar]
- Pamies, D.; Vilanova, E.; Sogorb, M.A. Functional pathways altered after silencing Pnpla6 (the codifying gene of Neuropathy Target Esterase) in mouse embryonic stem cells under differentiation. In Vitro Cell. Dev. Biol. Anim 2013. [Google Scholar] [CrossRef]
- Moser, M.; Li, Y.; Vaupel, K.; Kretzschmar, D.; Kluge, R.; Glynn, P.; Buettner, R. Placental failure and impaired vasculogenesis result in embryonic lethality for neuropathy target esterase-deficient mice. Mol. Cell. Biol 2004, 24, 1667–1679. [Google Scholar]
- Lotti, M.; Moretto, A. Organophosphate-induced delayed polyneuropathy. Toxicol. Rev 2005, 24, 37–49. [Google Scholar]
- Estévez, J.; Mangas, I.; Sogorb, M.Á.; Vilanova, E. Interactions of neuropathy inducers and potentiators/promoters with soluble esterases. Chem. Biol. Interact 2013, 203, 245–250. [Google Scholar]
- Johnson, M.K. The delayed neurotoxic effect of some organophosphorus compounds. Identification of the phosphorylation site as an esterase. Biochem. J 1969, 114, 711–717. [Google Scholar]
- Pallocca, G.; Fabbri, M.; Sacco, M.G.; Gribaldo, L.; Pamies, D.; Laurenza, I.; Bal-Price, A. miRNA expression profiling in a human stem cell-based model as a tool for developmental neurotoxicity testing. Cell Biol. Toxicol 2013, 29, 239–257. [Google Scholar]
- Hong, M.S.; Hong, S.J.; Barhoumi, R.; Burghardt, R.C.; Donnelly, K.C.; Wild, J.R.; Venkatraj, V.; Tiffany-Castiglioni, E. Neurotoxicity induced in differentiated SK-N-SH-SY5Y human neuroblastoma cells by organophosphorus compounds. Toxicol. Appl. Pharmacol 2003, 186, 110–118. [Google Scholar]
- Sachana, M.; Flaskos, J.M.; Alexaki, E.M.; Hargreaves, A.J. Inhibition of neurite outgrowth in N2a cells by leptophos and carbaryl: effects on neurofilament heavy chain, GAP-43 and HSP-70. Toxicol. In Vitro 2003, 17, 115–120. [Google Scholar]
- Sachana, M.; Flaskos, J.; Sidiropoulou, E.; Yavari, C.A.; Hargreaves, A.J. Inhibition of extension outgrowth in differentiating rat C6 glioma cells by chlorpyrifos and chlorpyrifos oxon: Effects on microtubule proteins. Toxicol. In Vitro 2008, 22, 1387–1391. [Google Scholar]
- Howard, A.S.; Bucelli, R.; Jett, D.A.; Bruun, D.M.; Yang, D.; Lein, P.J. Chlorpyrifos exerts opposing effects on axonal and dendritic growth in primary neuronal cultures. Toxicol. Appl. Pharmacol 2005, 207, 112–124. [Google Scholar]
- Massicotte, C.; Jortner, B.S.; Ehrich, M. Morphological effects of neuropathy-inducing organophosphorus compounds in primary dorsal root ganglia cell cultures. Neurotoxicology 2003, 24, 787–796. [Google Scholar]
- Li, W.; Casida, J.E. Organophosphorus neuropathy target esterase inhibitors selectively block outgrowth of neurite-like and cell processes in cultured cells. Toxicol. Lett 1998, 98, 139–146. [Google Scholar]
- Henschler, D.; Schmuck, G.; van Aerssen, M.; Schiffmann, D. The inhibitory effect of neuropathic organophosphate esters on neurite outgrowth in cell cultures: A basisfor screening for delayed neurotoxicity. Toxicol. In Vitro 1992, 6, 327–235. [Google Scholar]
- Li, Z.; Szurek, P.F.; Jiang, C.; Pao, A.; Bundy, B.; Le, W.D.; Bradley, A.; Yu, Y.E. Neuronal differentiation of NTE-deficient embryonic stem cells. Biochem. Biophys. Res. Commun 2005, 330, 1103–1109. [Google Scholar]
- Slotkin, T.A.; Levin, E.D.; Seidler, F.J. Developmental neurotoxicity of parathion: Progressive effects on serotonergic systems in adolescence and adulthood. Neurotoxicol. Teratol 2009, 31, 11–17. [Google Scholar]
- Laurenza, I.; Pallocca, G.; Mennecozzi, M.; Scelfo, B.; Pamies, D.; Bal-Price, A. A human pluripotent carcinoma stem cell-based model for in vitro developmental neurotoxicity testing: Effects of methylmercury, lead and aluminum evaluated by gene expression studies. Int. J. Dev. Neurosci 2013. [Google Scholar] [CrossRef]
- Pleasure, S.J.; Page, C.; Lee, V.M. Pure, postmitotic, polarized human neurons derived from NTera 2 cells provide a system for expressing exogenous proteins in terminally differentiated neurons. J. Neurosci 1992, 12, 1802–1815. [Google Scholar]
- Li, B.; Zhou, J.; Liu, P.; Hu, J.; Jin, H.; Shimono, Y.; Takahashi, M.; Xu, G. Polycomb protein Cbx4 promotes SUMO modification of de novo DNA methyltransferase Dnmt3a. Biochem. J 2007, 405, 369–378. [Google Scholar]
- Marzluff, W.F.; Gongidi, P.; Woods, K.R.; Jin, J.; Maltais, L.J. The human and mouse replication-dependent histone genes. Genomics 2002, 80, 487–498. [Google Scholar]
- Camargo, F.D.; Gokhale, S.; Johnnidis, J.B.; Fu, D.; Bell, G.W.; Jaenisch, R.; Brummelkamp, T.R. YAP1 increases organ size and expands undifferentiated progenitor cells. Curr. Biol 2007, 17, 2054–2060, Erratum in: Curr. Biol. 2007, 17, 2094. [Google Scholar]
- Singh, S.; Kumar, V.; Thakur, S.; Banerjee, B.D.; Chandna, S.; Rautela, R.S.; Grover, S.S.; Rawat, D.S.; Pasha, S.T.; Jain, S.K.; et al. DNA damage and cholinesterase activity in occupational workers exposed to pesticides. Environ. Toxicol. Pharmacol 2011, 31, 278–285. [Google Scholar]
- Singh, S.; Kumar, V.; Vashisht, K.; Singh, P.; Banerjee, B.D.; Rautela, R.S.; Grover, S.S.; Rawat, D.S.; Pasha, S.T.; Jain, S.K.; et al. Role of genetic polymorphisms of CYP1A1, CYP3A5, CYP2C9, CYP2D6, and PON1 in the modulation of DNA damage in workers occupationally exposed to organophosphate pesticides. Toxicol. Appl. Pharmacol 2011, 257, 84–92. [Google Scholar]
- Singh, S.; Kumar, V.; Thakur, S.; Banerjee, B.D.; Rautela, R.S.; Grover, S.S.; Rawat, D.S.; Pasha, S.T.; Jain, S.K.; Ichhpujani, R.L.; et al. Paraoxonase-1 genetic polymorphisms and susceptibility to DNA damage in workers occupationally exposed to organophosphate pesticides. Toxicol. Appl. Pharmacol 2011, 252, 130–137. [Google Scholar]
- Hébert, J.M.; Lin, M.; Partanen, J.; Rossant, J.; McConnell, S.K. FGF signaling through FGFR1 is required for olfactory bulb morphogenesis. Development 2003, 130, 1101–1111. [Google Scholar]
- Zhang, H.; Deo, M.; Thompson, R.C.; Uhler, M.D.; Turner, D.L. Negative regulation of Yap during neuronal differentiation. Dev. Biol 2012, 361, 103–115. [Google Scholar]
- Teixidó, E.; Piqué, E.; Gómez-Catalán, J.; Llobet, J.M. Assessment of developmental delay in the zebrafish embryo teratogenicity assay. Toxicol. In Vitro 2013, 27, 469–478. [Google Scholar]
- Islas-González, K.; González-Horta, C.; Sánchez-Ramírez, B.; Reyes-Aragón, E.; Levario-Carrillo, M. In vitro assessment of the genotoxicity of ethyl paraoxon in newborns and adults. Hum. Exp. Toxicol 2005, 24, 319–324. [Google Scholar]
- Hreljac, I.; Zajc, I.; Lah, T.; Filipic, M. Effects of model organophosphorous pesticides on DNA damage and proliferation of HepG2 cells. Environ. Mol. Mutagen 2008, 49, 360–367. [Google Scholar]
- Sogorb, M.A.; González-González, I.; Pamies, D.; Vilanova, E. An alternative in vitro method for detecting neuropathic compounds based on acetylcholinesterase inhibition and on inhibition and aging of neuropathy target esterase (NTE). Toxicol. In Vitro 2010, 24, 942–952. [Google Scholar]
- Mangas, I.; Vilanova, E.; Estévez, J. NTE and non-NTE esterases in brain membrane: Kinetic characterization with organophosphates. Toxicology 2012, 297, 17–25. [Google Scholar]
- Mangas, I.; Vilanova, E.; Estévez, J. Kinetics of the inhibitory interaction of organophosphorus neuropathy inducers and non-inducers in soluble esterases in the avian nervous system. Toxicol. Appl. Pharmacol 2011, 256, 360–368. [Google Scholar]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol 2004, 3, 30. [Google Scholar]
- Sogorb, M.A.; García-Argüelles, S.; Carrera, V.; Vilanova, E. Serum albumin is as efficient as paraxonase in the detoxication of paraoxon at toxicologically relevant concentrations. Chem. Res. Toxicol 2008, 21, 1524–1529. [Google Scholar]
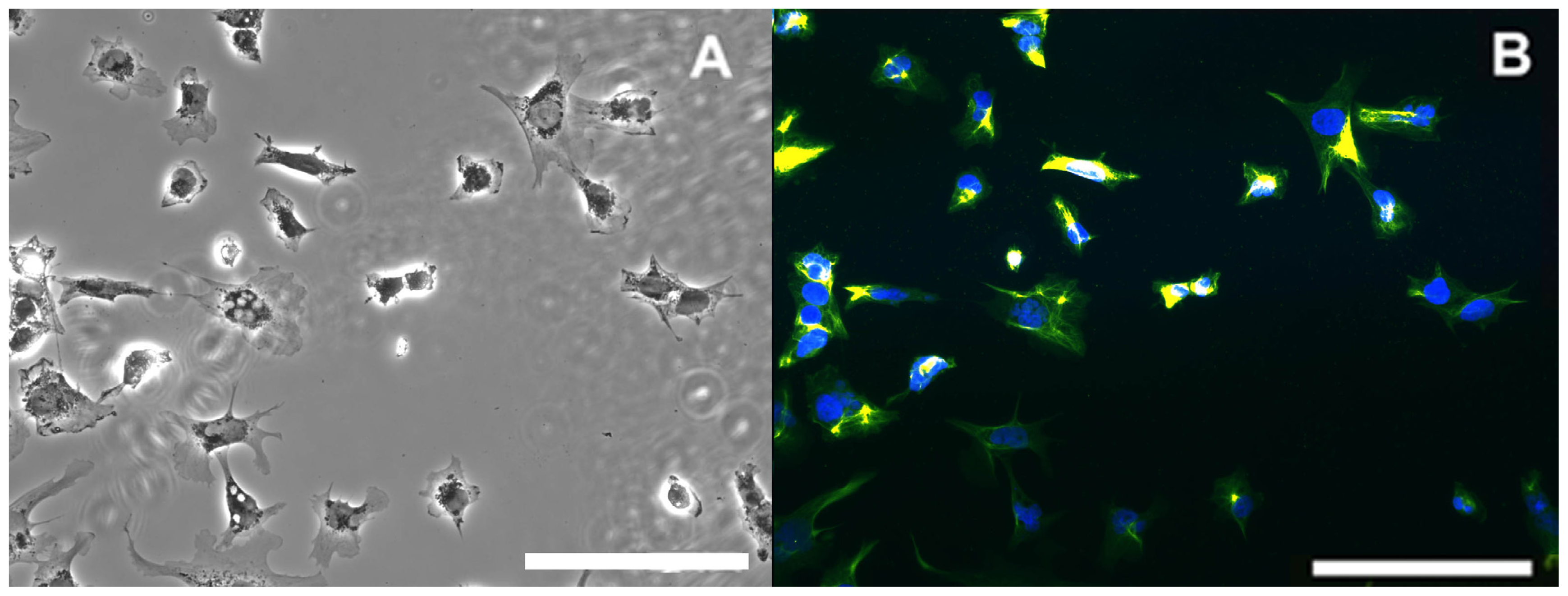
 ) for 4 days (A); 10 days (B); and 15 days (C). Data represent mean ± SEM of the four independent experiments with 16 independent technical replicates for each experimental condition run. (* statistically different from the controls for p < 0.05 in both paraoxon and mipafox by Dunnett’s test).
) for 4 days (A); 10 days (B); and 15 days (C). Data represent mean ± SEM of the four independent experiments with 16 independent technical replicates for each experimental condition run. (* statistically different from the controls for p < 0.05 in both paraoxon and mipafox by Dunnett’s test).
) for 4 days (A); 10 days (B); and 15 days (C). Data represent mean ± SEM of the four independent experiments with 16 independent technical replicates for each experimental condition run. (* statistically different from the controls for p < 0.05 in both paraoxon and mipafox by Dunnett’s test).
) for 4 days (A); 10 days (B); and 15 days (C). Data represent mean ± SEM of the four independent experiments with 16 independent technical replicates for each experimental condition run. (* statistically different from the controls for p < 0.05 in both paraoxon and mipafox by Dunnett’s test). ) for 4 days (A); 10 days (B); and 15 days (C). NTE activity is expressed as the percentage of activity determined in the time-matched control (non-exposed) cultures. Data represent mean ± SEM of three independent experiments with 8 technical replicates for each experimental condition. (* statistically different from the controls for p < 0.05 with a Dunnett’s test).
) for 4 days (A); 10 days (B); and 15 days (C). NTE activity is expressed as the percentage of activity determined in the time-matched control (non-exposed) cultures. Data represent mean ± SEM of three independent experiments with 8 technical replicates for each experimental condition. (* statistically different from the controls for p < 0.05 with a Dunnett’s test).
) for 4 days (A); 10 days (B); and 15 days (C). NTE activity is expressed as the percentage of activity determined in the time-matched control (non-exposed) cultures. Data represent mean ± SEM of three independent experiments with 8 technical replicates for each experimental condition. (* statistically different from the controls for p < 0.05 with a Dunnett’s test).
) for 4 days (A); 10 days (B); and 15 days (C). NTE activity is expressed as the percentage of activity determined in the time-matched control (non-exposed) cultures. Data represent mean ± SEM of three independent experiments with 8 technical replicates for each experimental condition. (* statistically different from the controls for p < 0.05 with a Dunnett’s test).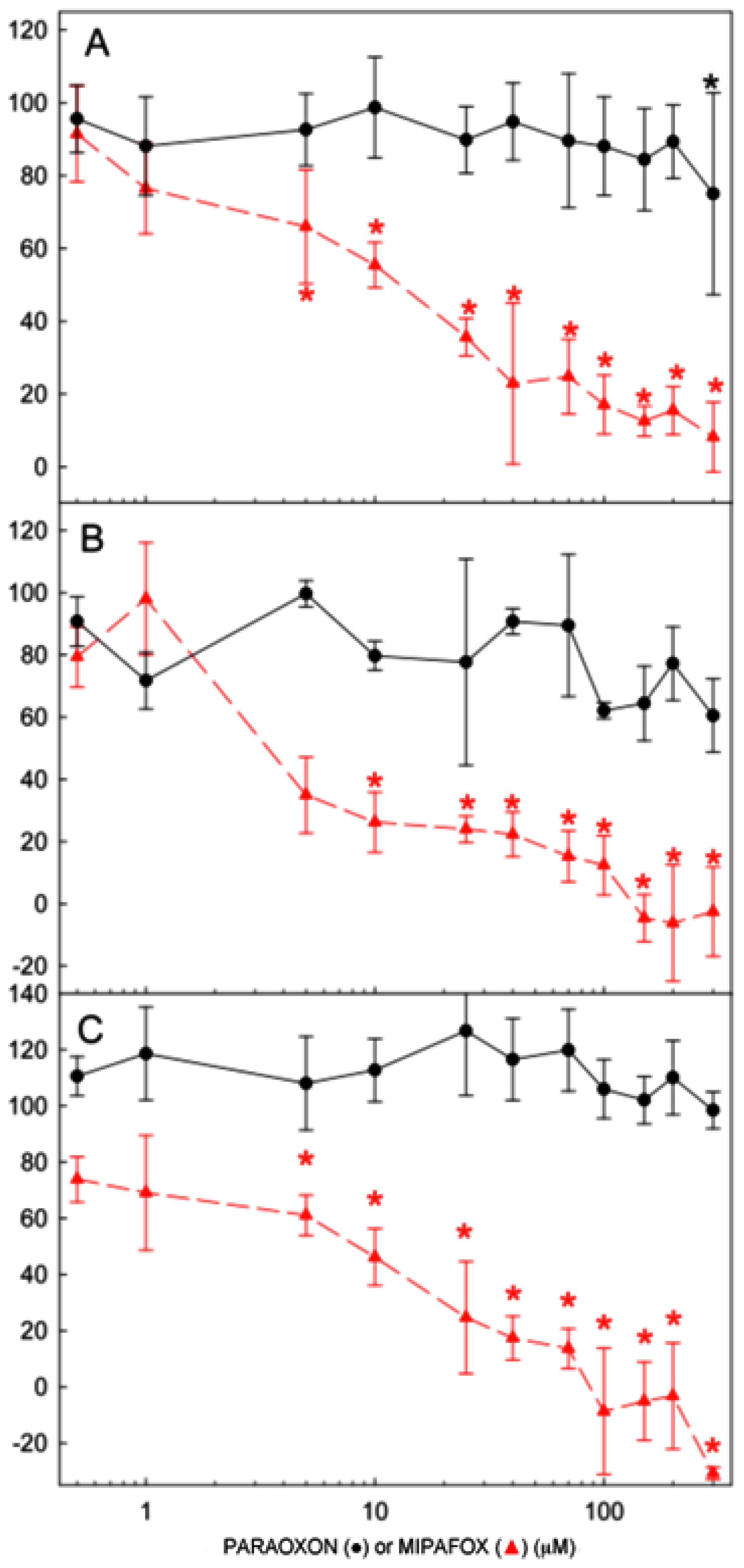
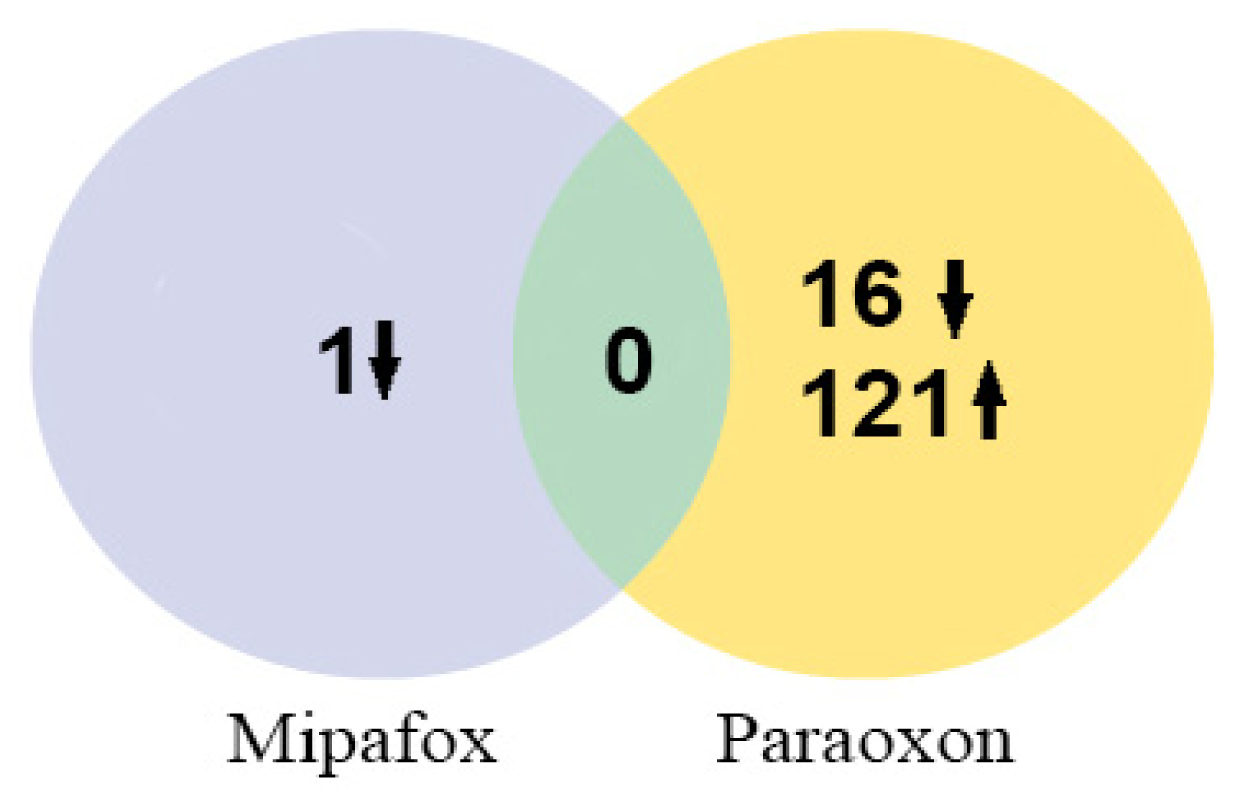


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Number of genes associated with processes altered | Corrected p-value |
|---|---|---|
| Cluster Enrichment score = 1.7 | ||
| Chromatin assembly | 4 | 0.001 * |
| Nucleosome assembly | 3 | 0.010 * |
| Protein-DNA complex assembly | 3 | 0.011 * |
| Nucleosome organization | 3 | 0.012 * |
| Cellular macromolecular complex assembly | 4 | 0.018 * |
| DNA package | 3 | 0.018 * |
| Cellular macromolecular complex subunit organization | 4 | 0.024 * |
| Chromatin organization | 4 | 0.028 * |
| Chromosome organization | 4 | 0.053 * |
| Cluster Enrichment score = 1.33 | ||
| Enzyme binding | 6 | 0.0019 * |
| Cell death | 5 | 0.036 * |
| Death | 5 | 0.037 * |
| Apoptosis | 3 | 0.29 |
| Programmed cell death | 3 | 0.3 |
| Cluster Enrichment score = 1.04 | ||
| Regulation of cell size | 3 | 0.051 * |
| Regulation of cellular component size | 3 | 0.083 |
| Neuro differentiation | 3 | 0.18 |
| Gene | Name | NCBI entry | FC real |
|---|---|---|---|
| HIST1H4E | histone cluster 1, H4e | 8367 | −1.69 |
| LUZP6 | leucine zipper protein 6 | 767558 | −1.57 |
| LOC400804 | hypothetical LOC400804 | 400804 | −1.24 |
| HIST1H2AB | histone cluster 1, H2ab | 8335 | −1.23 |
| C14orf162 | chromosome 14 open reading frame 162 | 56936 | −1.19 |
| YAP1 | Yes-associated protein 1 | 10413 | −1.15 |
| FGFR1 | fibroblast growth factor receptor 1 | 2260 | −1.05 |
| AKAP12 | A kinase (PRKA) anchor protein 12 | 9590 | −1.03 |
| UBE2Z | ubiquitin-conjugating enzyme E2Z | 65264 | −1.01 |
| SRGAP2P2 | SLIT-ROBO Rho GTPase activating protein 2 pseudogene 2 | 647135 | −1.01 |
| AXIN2 | axin 2 | 8313 | −1.00 |
| LOC646214 | p21-activated kinase 2 pseudogene | 646214 | 1.00 |
| LOC100130654 | hypothetical protein LOC100130654 | 100130654 | 1.00 |
| CCR6 | chemokine (C-C motif) receptor 6 | 1235 | 1.01 |
| C14orf135 | chromosome 14 open reading frame 135 | 64430 | 1.01 |
| KIRREL2 | kin of IRRE like 2 (Drosophila) | 84063 | 1.01 |
| C2orf27A | chromosome 2 open reading frame 27A | 29798 | 1.02 |
| BMP8B | bone morphogenetic protein 8b | 656 | 1.03 |
| RNF113B | ring finger protein 113B | 140432 | 1.05 |
| GCLM | glutamate-cysteine ligase, modifier subunit | 2730 | 1.06 |
| TTC16 | tetratricopeptide repeat domain 16 | 158248 | 1.07 |
| PARP4 | poly (ADP-ribose) polymerase family, member 4 | 143 | 1.07 |
| LOC100134868 | hypothetical LOC100134868 | 100134868 | 1.07 |
| SCD5 | stearoyl-CoA desaturase 5 | 79966 | 1.08 |
| PTK2B | PTK2B protein tyrosine kinase 2 beta | 2185 | 1.08 |
| LAT2 | linker for activation of T cells family, member 2 | 7462 | 1.10 |
| RPL13AP17 | ribosomal protein L13a pseudogene 17 | 399670 | 1.10 |
| ACBD5 | acyl-CoA binding domain containing 5 | 91452 | 1.16 |
| C1orf152 | profilin 1 pseudogene | 767846 | 1.17 |
| YY2 | YY2 transcription factor | 404281 | 1.18 |
| LOC100133791 | hypothetical protein LOC100133791 | 100133791 | 1.21 |
| LOC100233209 | hypothetical LOC100233209 | 100233209 | 1.24 |
| ILDR1 | immunoglobulin-like domain containing receptor 1 | 286676 | 1.26 |
| LOC648740 | actin, beta pseudogene | 648740 | 1.28 |
| SET | SET nuclear oncogene | 6418 | 1.29 |
| LOC100131581 | hypothetical LOC100131581 | 100131581 | 1.29 |
| CXCL5 | chemokine (C-X-C motif) ligand 5 nascent-polypeptide-associated complex alpha polypeptide | 6374 | 1.30 |
| NACAP1 | pseudogene 1 | 83955 | 1.30 |
| KRTAP10-9 | keratin associated protein 10-9 | 386676 | 1.33 |
| CBX3 | chromobox homolog 3 | 11335 | 1.37 |
| SNRPD2P2 | small nuclear ribonucleoprotein D2 pseudogene 2 | 645339 | 1.39 |
| SGSM2 | small G protein signaling modulator 2 | 9905 | 1.42 |
| ANXA2 | annexin A2 | 302 | 1.59 |
| REREP3 | arginine-glutamic acid dipeptide (RE) repeats pseudogene 3 | 646396 | 1.78 |
| FKSG2 | tumor protein, translationally-controlled 1 pseudogene | 59347 | 1.80 |
| TXNDC17 | thioredoxin domain containing 17 | 84817 | 1.94 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pamies, D.; Sogorb, M.A.; Fabbri, M.; Gribaldo, L.; Collotta, A.; Scelfo, B.; Vilanova, E.; Harris, G.; Bal-Price, A. Genomic and Phenotypic Alterations of the Neuronal-Like Cells Derived from Human Embryonal Carcinoma Stem Cells (NT2) Caused by Exposure to Organophosphorus Compounds Paraoxon and Mipafox. Int. J. Mol. Sci. 2014, 15, 905-926. https://doi.org/10.3390/ijms15010905
Pamies D, Sogorb MA, Fabbri M, Gribaldo L, Collotta A, Scelfo B, Vilanova E, Harris G, Bal-Price A. Genomic and Phenotypic Alterations of the Neuronal-Like Cells Derived from Human Embryonal Carcinoma Stem Cells (NT2) Caused by Exposure to Organophosphorus Compounds Paraoxon and Mipafox. International Journal of Molecular Sciences. 2014; 15(1):905-926. https://doi.org/10.3390/ijms15010905
Chicago/Turabian StylePamies, David, Miguel A. Sogorb, Marco Fabbri, Laura Gribaldo, Angelo Collotta, Bibiana Scelfo, Eugenio Vilanova, Georgina Harris, and Anna Bal-Price. 2014. "Genomic and Phenotypic Alterations of the Neuronal-Like Cells Derived from Human Embryonal Carcinoma Stem Cells (NT2) Caused by Exposure to Organophosphorus Compounds Paraoxon and Mipafox" International Journal of Molecular Sciences 15, no. 1: 905-926. https://doi.org/10.3390/ijms15010905