High-Level Expression of Pro-Form Lipase from Rhizopus oryzae in Pichia pastoris and Its Purification and Characterization

Abstract
:1. Introduction
2. Results and Discussion
2.1. Selection of Lipase Producing Clones
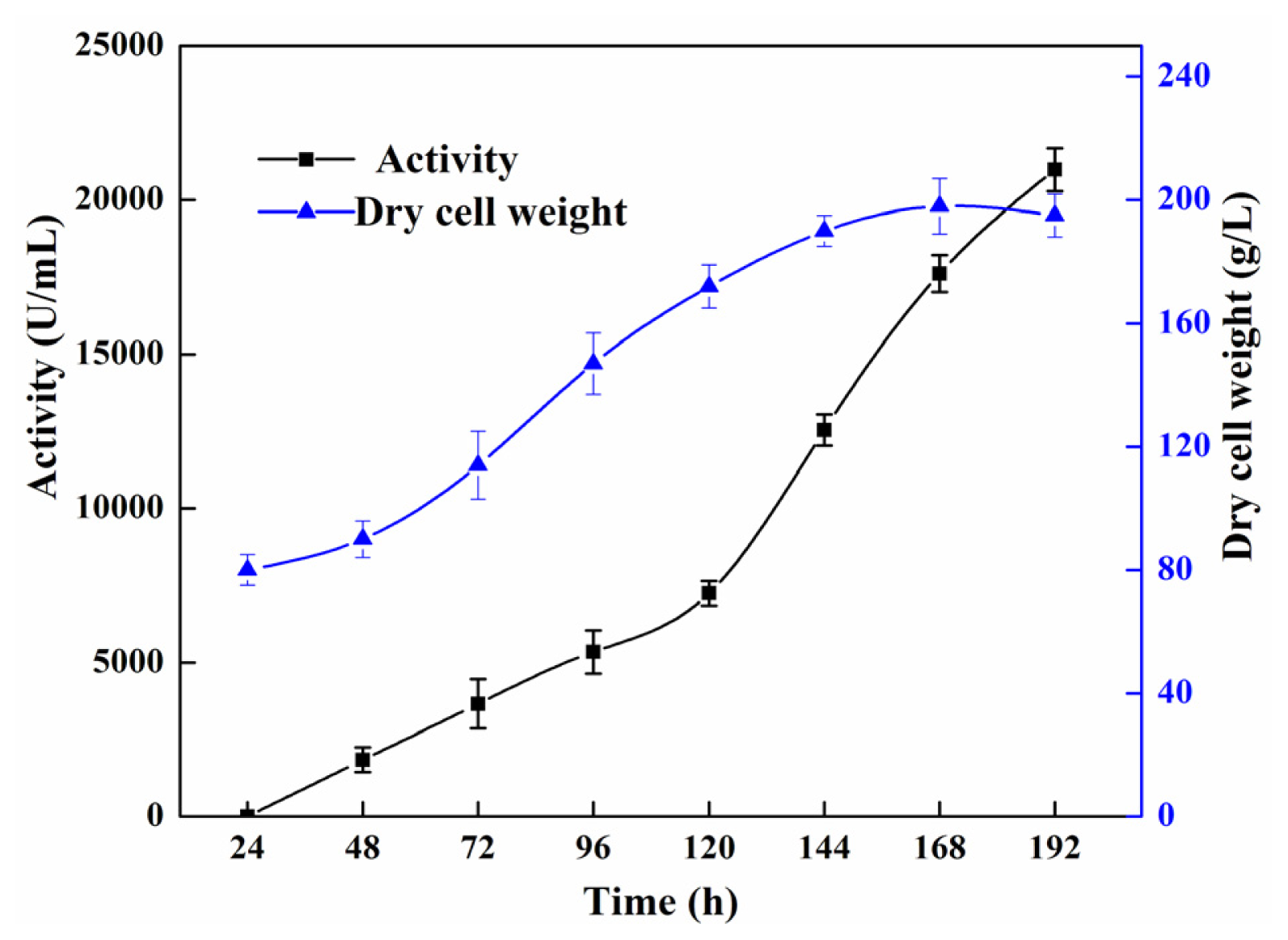
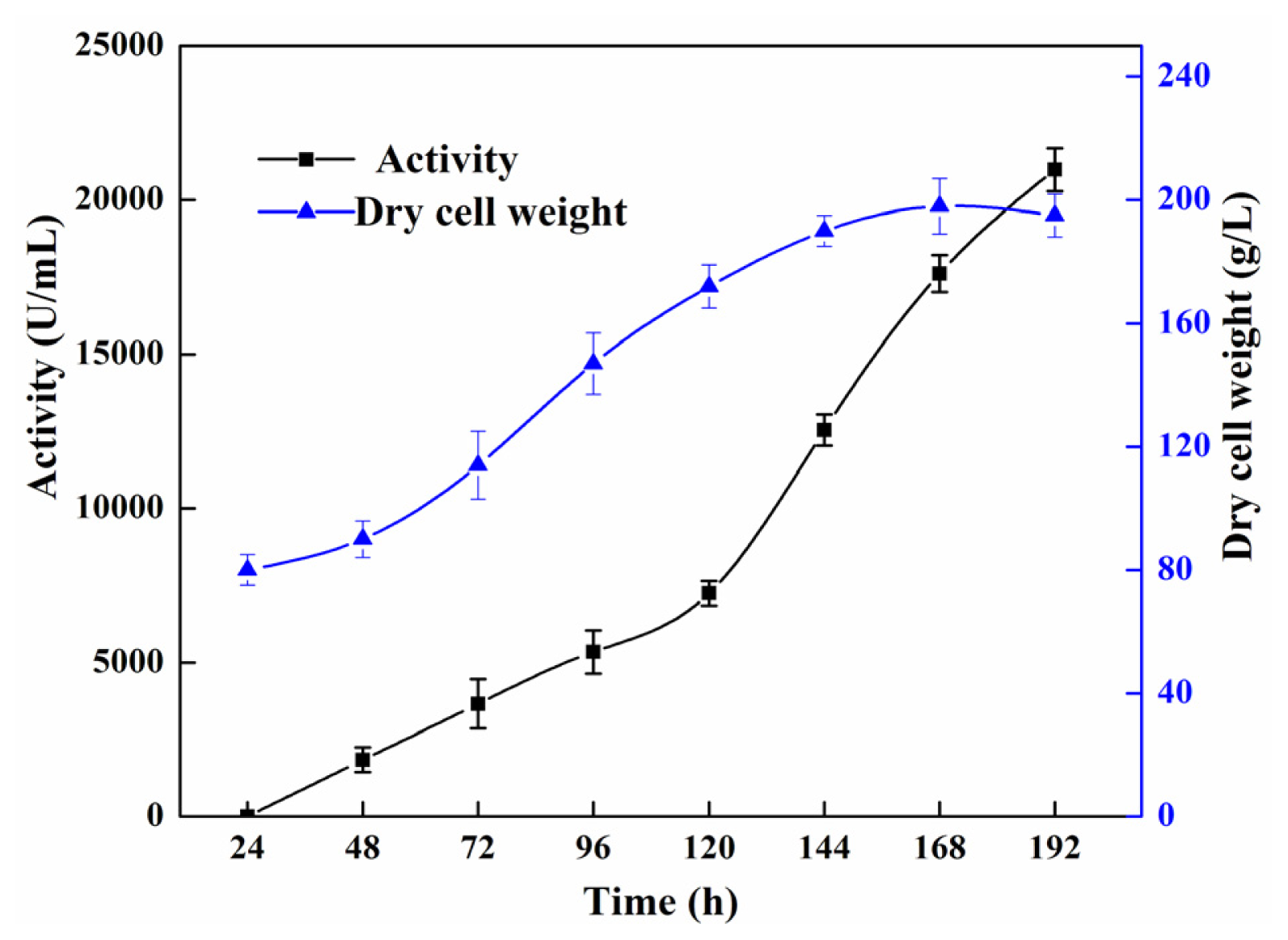
2.2. High Cell Density Fermentation for Production of Recombinant Lipase
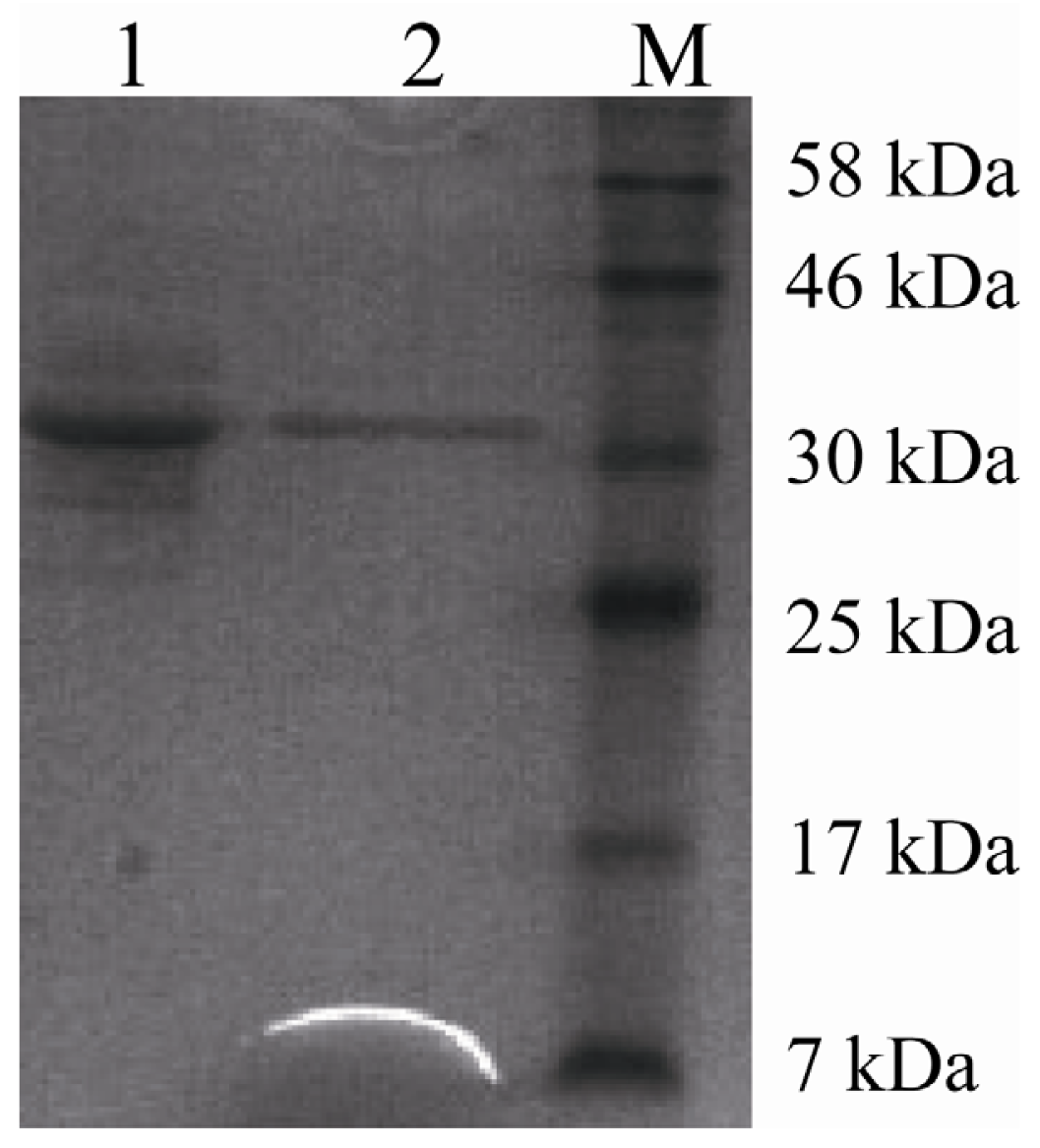
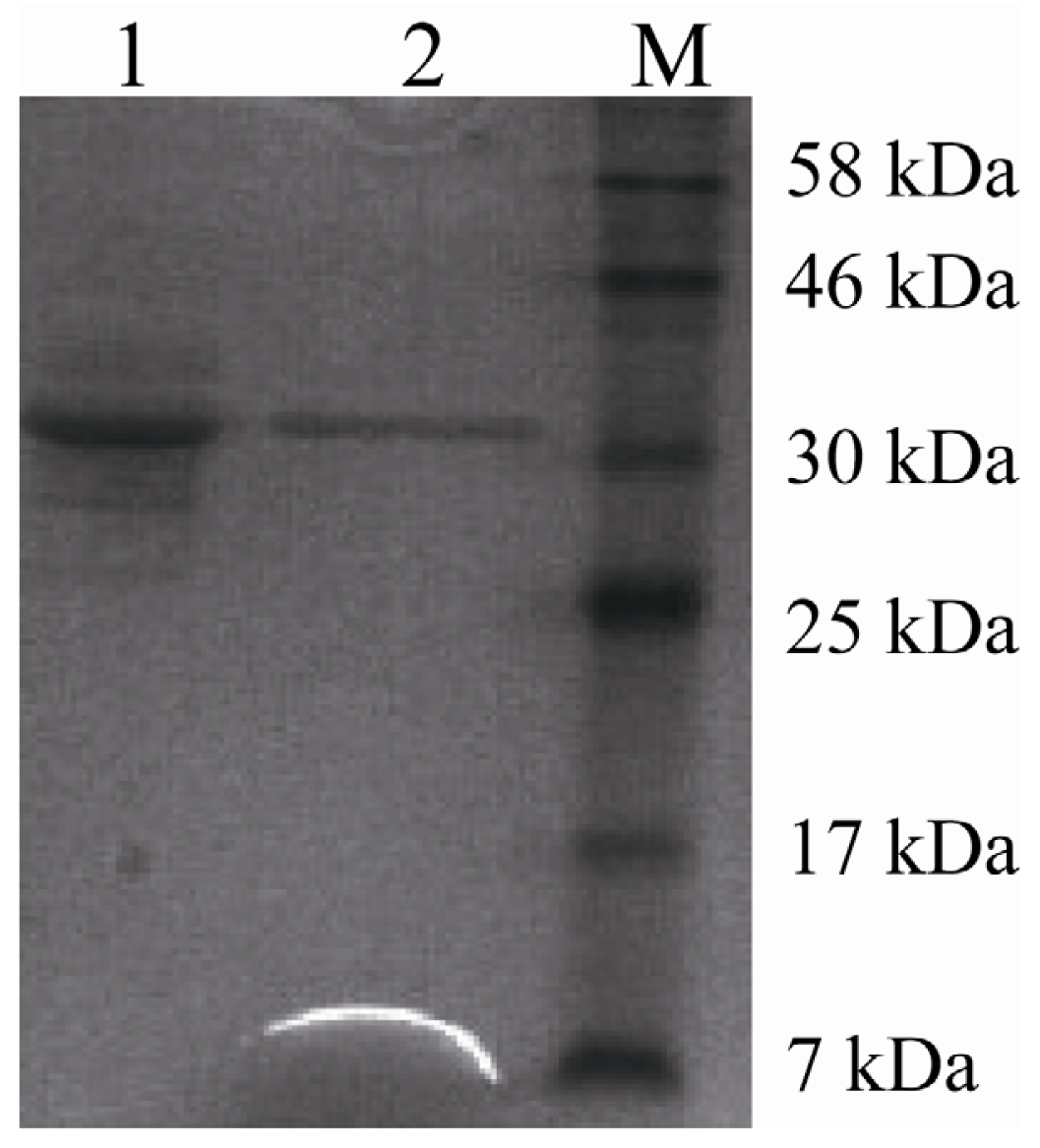
2.3. Purification of Recombinant Lipase
2.4. Biochemical Characteristics of Recombinant Lipase
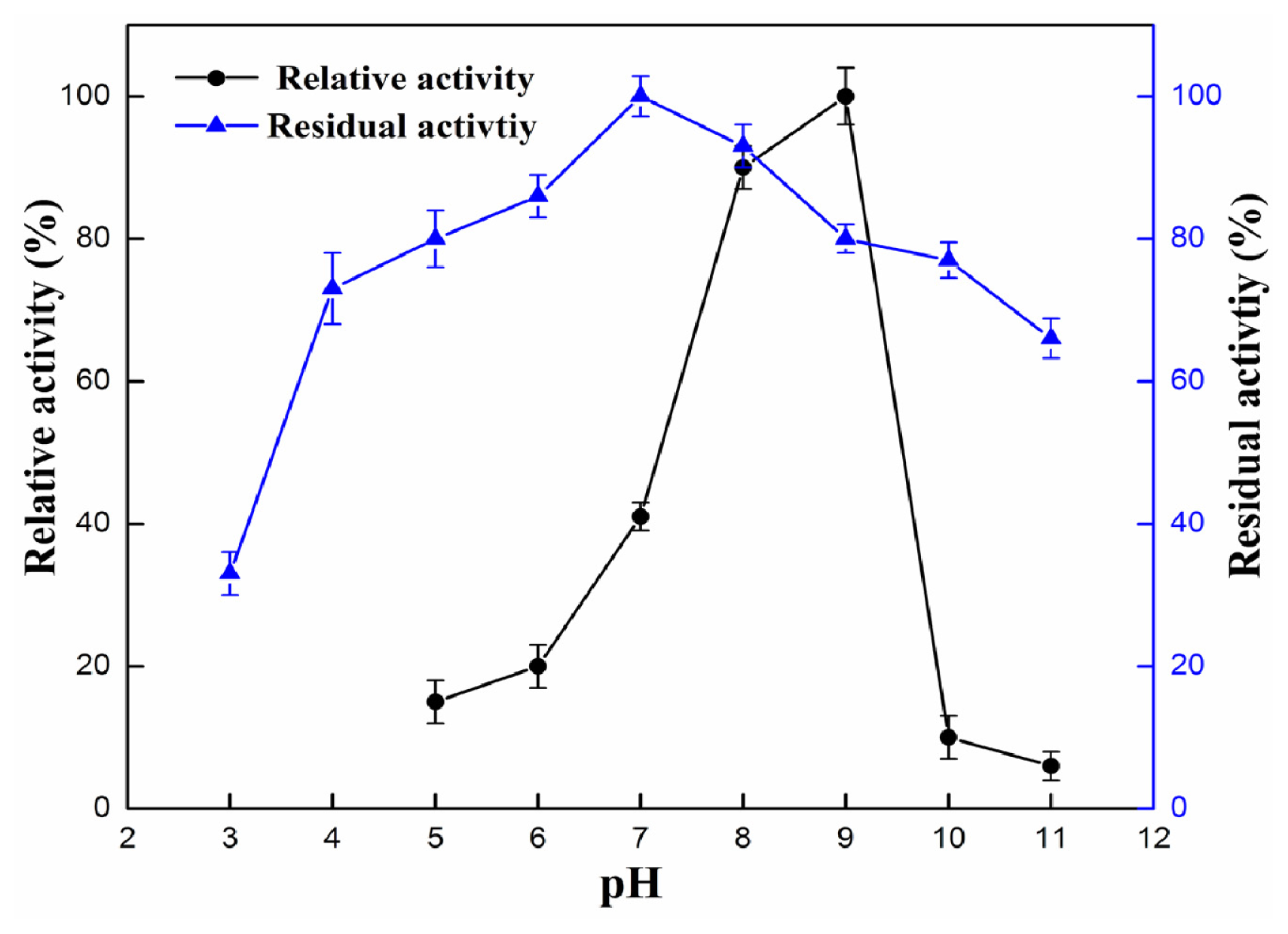
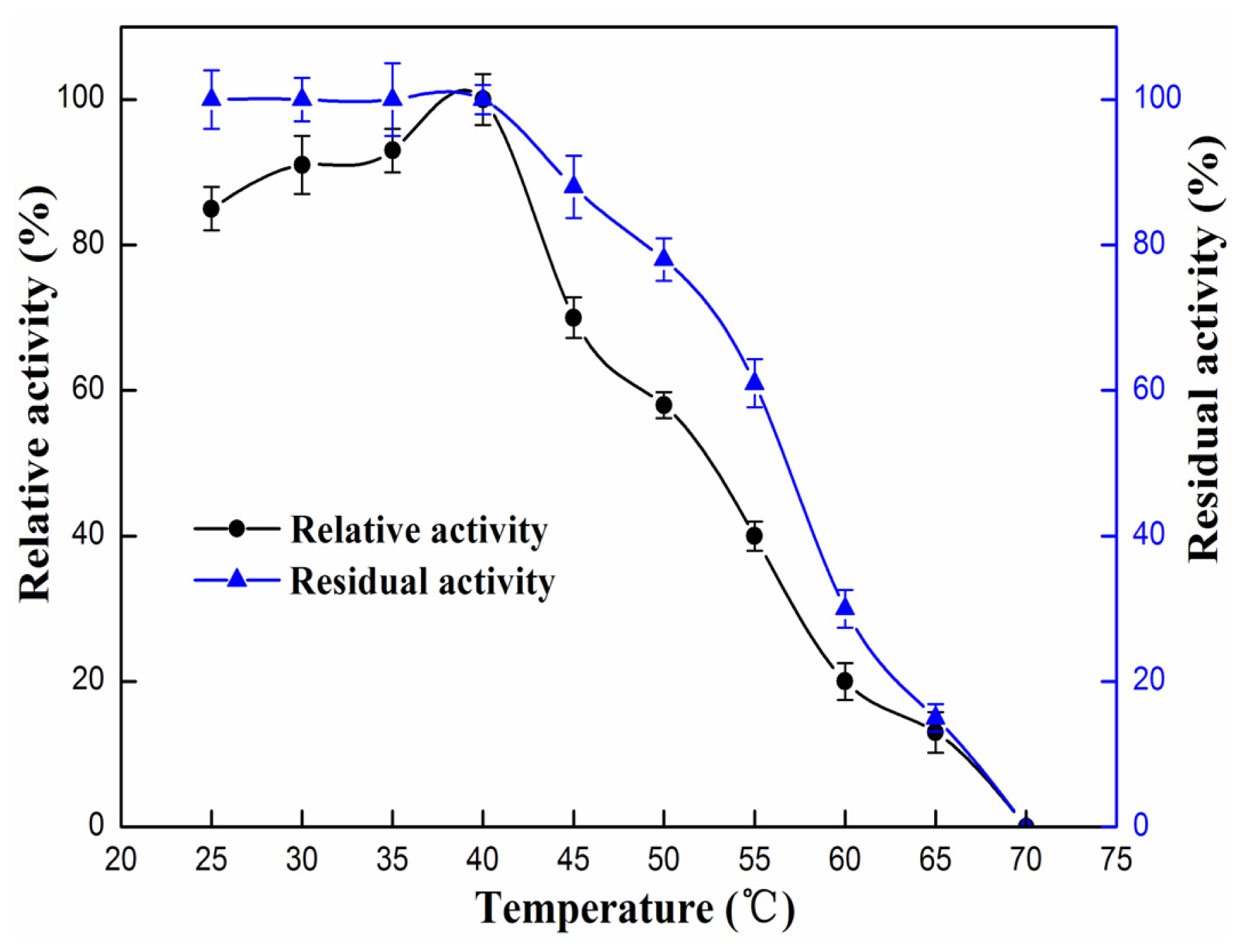
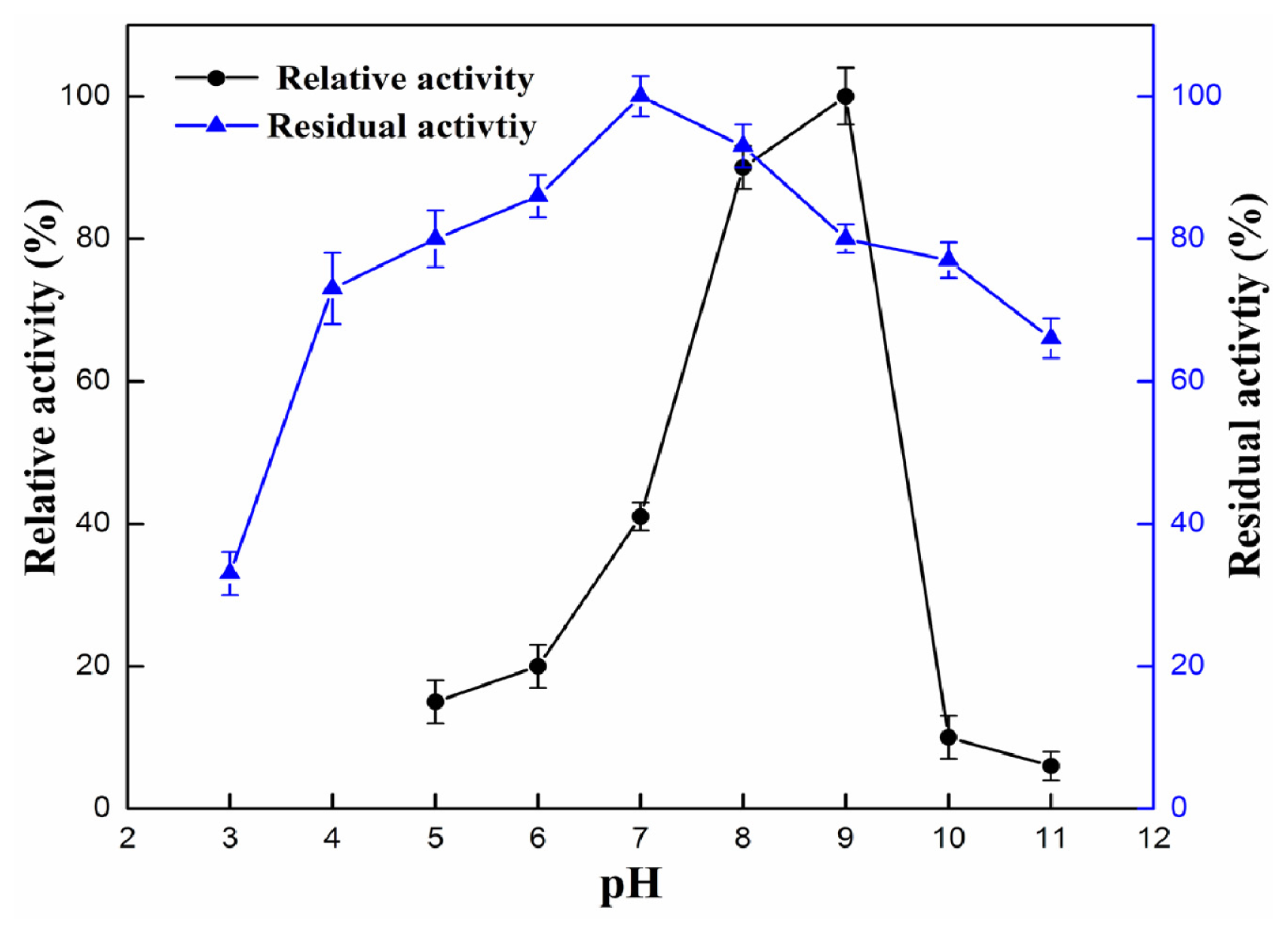
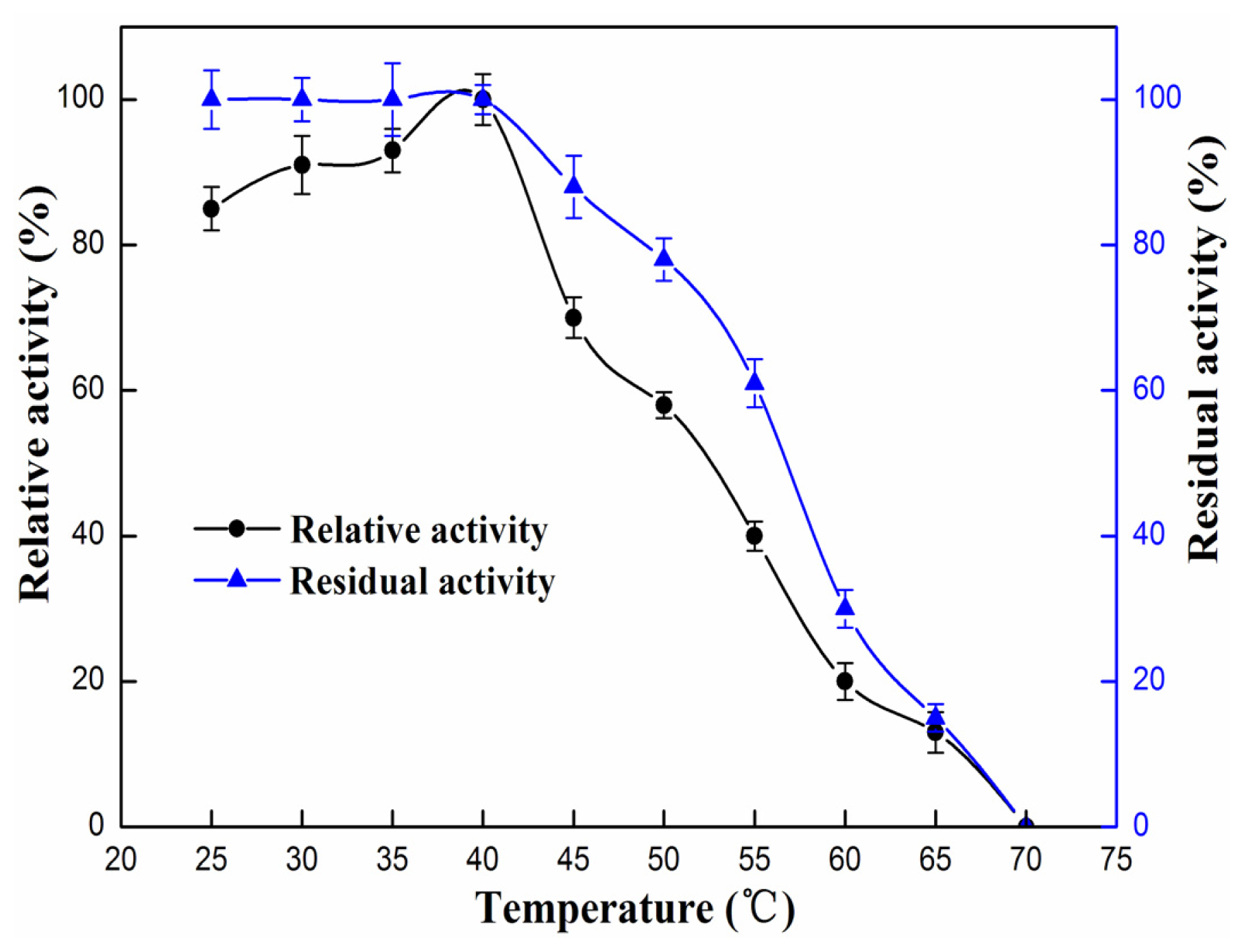
2.4.1. Effect of pH and Temperature
2.4.2. Effect of Metal Ions on Lipase
2.4.3. Effect of Detergents on Lipase
2.4.4. Effect of Organic Solvents
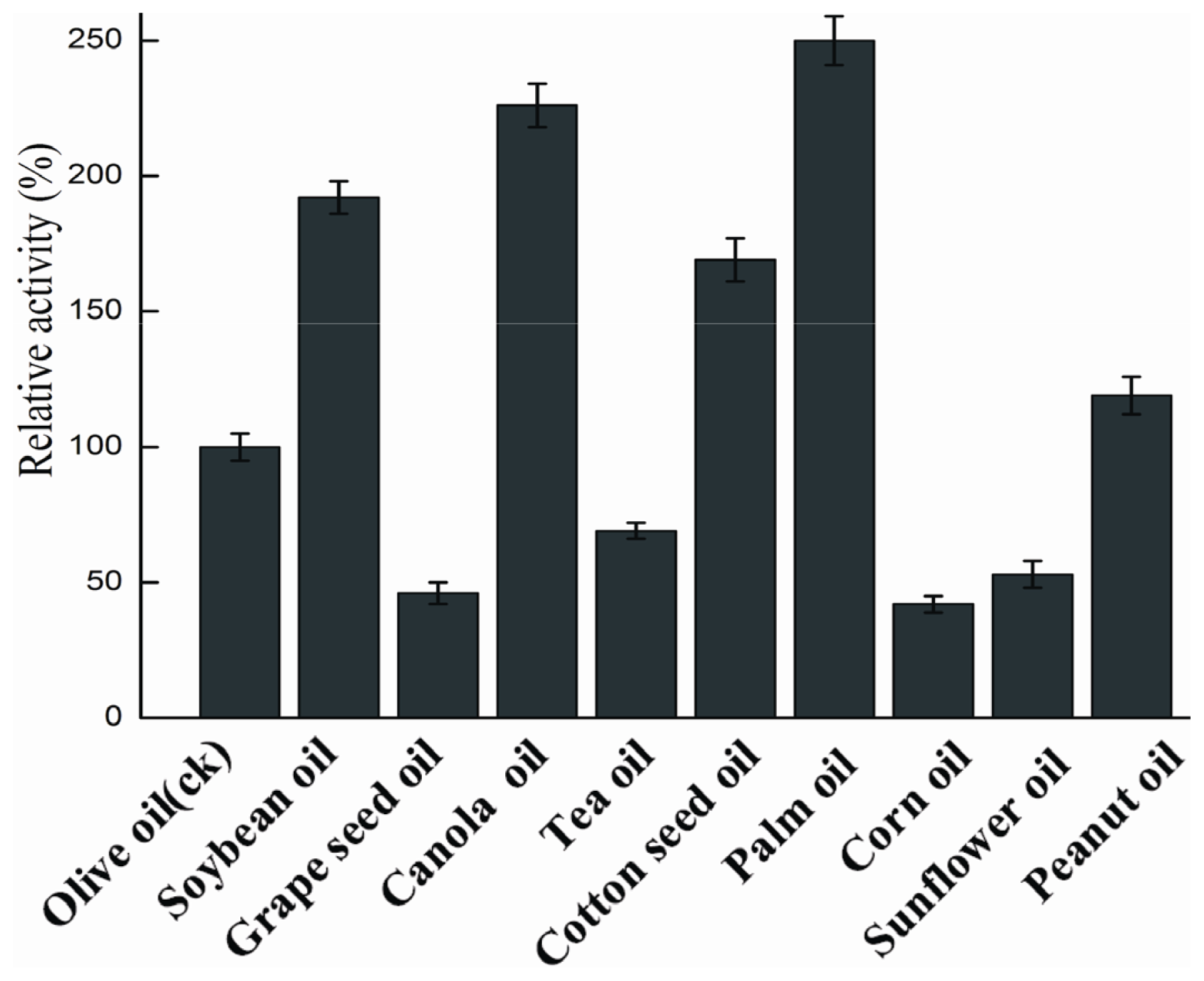
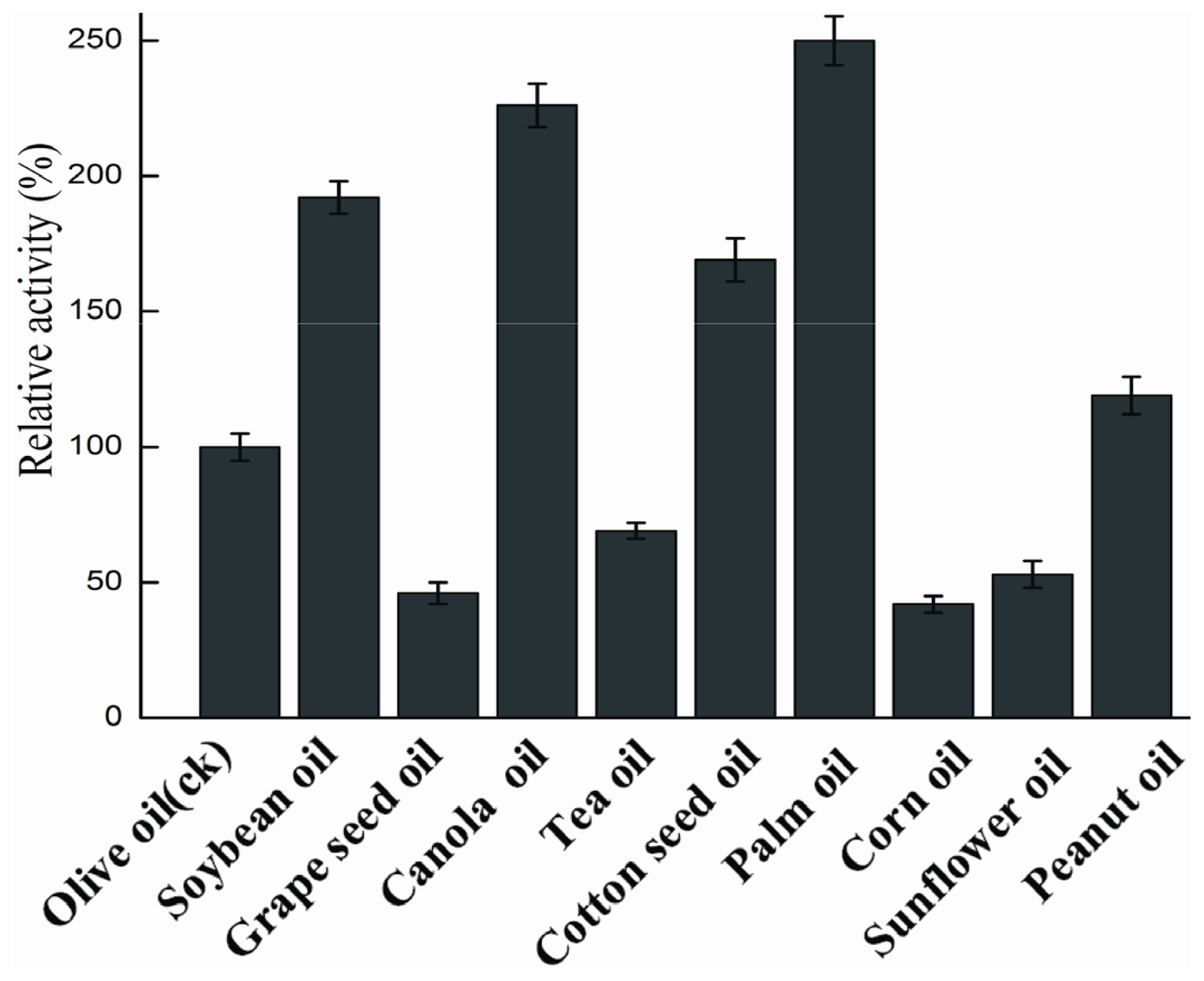
2.4.5. Substrate Specificity
3. Experimental Section
3.1. Strains, Plasmids and Materials
3.2. Vector Construction
3.3. Transformation and Selection of P. pastoris Clones
3.4. Shaking Flask Cultures
3.5. High Cell Density Fermentation
3.6. Biomass Analysis
3.7. Assay of Lipase Activity and Total Protein Concentration
3.8. Purification of Lipase
3.9. Biochemical Characteristics of Recombinant Lipase
3.9.1. Effect of pH and Temperature on Enzyme Activity and Stability
3.9.2. Effect of Metal Ions on Lipase Activity
3.9.3. Effect of Detergents on Lipase Activity
3.9.4. Effect of Organic Solvents on Lipase
3.9.5. Substrate Specificity
4. Conclusions
Supplementary Information
ijms-15-00203-s001.pdfAcknowledgments
Conflicts of Interest
References
- Jaeger, K.E.; Eggert, T. Lipases for biotechnology. Curr. Opin. Biotechnol 2002, 13, 390–397. [Google Scholar]
- Gupta, R.; Gupta, N.; Rathi, P. Bacterial lipases: An overview of production, purification and biochemical properties. Appl. Microbiol. Biotechnol 2004, 64, 763–781. [Google Scholar]
- Singh, A.K.; Mukhopadhyay, M. Overview of fungal lipase: A review. Appl. Biochem. Biotechnol 2012, 166, 486–520. [Google Scholar]
- Hama, S.; Yamaji, H.; Fukumizu, T.; Numata, T.; Tamalampudi, S.; Kondo, A.; Noda, H.; Fukuda, H. Biodiesel-fuel production in a packed-bed reactor using lipase-producing Rhizopus oryzae cells immobilized within biomass support particles. Biochem. Eng. J 2007, 34, 273–278. [Google Scholar]
- Kaieda, M.; Samukawa, T.; Matsumoto, T.; Ban, K.; Kondo, A.; Shimada, Y.; Noda, H.; Nomoto, F.; Ohtsuka, K.; Izumoto, E.; et al. Biodiesel fuel production from plant oil catalyzed by Rhizopus oryzae lipase in a water-containing system without an organic solvent. J. Biosci. Bioeng 1999, 88, 627–631. [Google Scholar]
- Niu, W.N.; Li, Z.P.; Tan, T. Secretion of pro-and mature Rhizopus arrhizus lipases by Pichia pastoris and properties of the proteins. Mol. Biotechnol 2006, 32, 73–81. [Google Scholar]
- Beer, H.D.; Wohlfahrt, G.; Schmid, R.D.; McCarthy, J.E.G. The folding and activity of the extracellular lipase of Rhizopus oryzae are modulated by a prosequence. Biochem. J 1996, 319, 351–359. [Google Scholar]
- Beer, H.D.; McCarthy, J.E.; Bornscheuer, U.T.; Schmid, R.D. Cloning, expression, characterization and role of the leader sequence of a lipase from Rhizopus oryzae. Biochim. Biophys. Acta 1998, 1399, 173–180. [Google Scholar]
- Yang, J.; Yan, X.; Huang, R.; Zhang, B. Codon optimization, expression and enzymatic comparison of Rhizopus oryzae lipases pro-ROL and m-ROL in Pichia pastoris. Chin. J. Biotech 2011, 27, 1780–1788. (in Chinese). [Google Scholar]
- Hidalgo, A.; Haas, M.; Bornscheuer, U.T. Heterologous production of functional forms of Rhizopus oryzae lipase in Escherichia coli. Appl. Environ. Microbiol 2005, 71, 8974–8977. [Google Scholar]
- Takahashi, S.; Ueda, M.; Atomi, H.; Beer, H.D.; Bornscheuer, U.T.; Schmid, R.D.; Tanaka, A. Extracellular production of active Rhizopus oryzae lipase by Saccharomyces cerevisiae. J. Ferment. Bioeng 1998, 86, 164–168. [Google Scholar]
- Macauley, S.; Fazenda, M.L.; McNeil, B.; Harvey, L.M. Heterologous protein production using the Pichia pastor is expression system. Yeast 2005, 22, 249–270. [Google Scholar]
- Lan, D.M.; Yang, N.; Wang, W.K.; Shen, Y.F.; Yang, B.; Wang, Y.H. A novel cold-active lipase from Candida albicans: Cloning, expression and characterization of the recombinant enzyme. Int. J. Mol. Sci 2011, 12, 3950–3965. [Google Scholar]
- Shi, B.H.; Zeng, L.Q.; Song, H.L.; Shi, Q.Q.; Wu, S.G. Cloning and expression of Aspergillus tamarii FS132 lipase gene in Pichia pastoris. Int. J. Mol. Sci 2010, 11, 2373–2382. [Google Scholar]
- Yu, M.R.; Lange, S.; Richter, S.; Tan, T.W.; Schmid, R.D. High-level expression of extracellular lipase Lip2 from Yarrowia lipolytica in Pichia pastoris and its purification and characterization. Protein Expr. Purif 2007, 53, 255–263. [Google Scholar]
- Yang, J.K.; Liu, L.Y. Codon optimization through a two-step gene synthesis leads to a high-level expression of Aspergillus niger lip 2 gene in Pichia pastoris. J. Mol. Catal. B Enzym 2010, 63, 164–169. [Google Scholar]
- Wang, S.L.; Lin, Y.T.; Liang, T.W.; Chio, S.H.; Ming, L.J.; Wu, P.C. Purification and characterization of extracellular lipases from Pseudomonas monteilii TKU009 by the use of soybeans as the substrate. J. Ind. Microbiol. Biotechnol 2009, 36, 65–73. [Google Scholar]
- Yoo, H.Y.; Simkhada, J.R.; Cho, S.S.; Park, D.H.; Kim, S.W.; Seong, C.N.; Yoo, J.C. A novel alkaline lipase from Ralstonia with potential application in biodiesel production. Bioresour. Technol 2011, 102, 6104–6111. [Google Scholar]
- Minning, S.; Schmidt-Dannert, C.; Schmid, R.D. Functional expression of Rhizopus oryzae lipase in Pichia pastoris: High-level production and some properties. J. Biotechnol 1998, 66, 147–156. [Google Scholar]
- Ji, Q.C.; Xiao, S.J.; He, B.F.; Liu, X.N. Purification and characterization of an organic solvent-tolerant lipase from Pseudomonas aeruginosa LX1 and its application for biodiesel production. J. Mol. Catal. B Enzym 2010, 66, 264–269. [Google Scholar]
- Yan, J.Y.; Yang, J.K.; Xu, L.; Yan, Y.J. Gene cloning, overexpression and characterization of a novel organic solvent tolerant and thermostable lipase from Galactomyces geotrichum Y05. J. Mol. Catal. B Enzym 2007, 49, 28–35. [Google Scholar]
- Yu, M.R.; Qin, S.W.; Tan, T.W. Purification and characterization of the extracellular lipase Lip2 from Yarrowia lipolytica. Process Biochem 2007, 42, 384–391. [Google Scholar]
- Sharon, C.; Furugoh, S.; Yamakido, T.; Ogawa, H.I.; Kato, Y. Purification and characterization of a lipase from Pseudomonas aeruginosa KKA-5 and its role in castor oil hydrolysis. J. Ind. Microbiol. Biotechnol 1998, 20, 304–307. [Google Scholar]
- Cheng, Y.Y.; Qian, Y.K.; Li, Z.F.; Wu, Z.H.; Liu, H.; Li, Y.Z. A novel cold-adapted lipase from Sorangium cellulosum strain So0157-2: Gene cloning, expression, and enzymatic characterization. Int. J. Mol. Sci 2011, 12, 6765–6780. [Google Scholar]
- Yang, K.S.; Sohn, J.H.; Kim, H.K. Catalytic properties of a lipase from Photobacterium lipolyticum for biodiesel production containing a high methanol concentration. J. Biosci. Bioeng 2009, 107, 599–604. [Google Scholar]
- Shimada, Y.; Sugihara, A.; Nakano, H.; Nagao, T.; Suenaga, M.; Nakai, S.; Tominaga, Y. Fatty acid specificity of Rhizopus delemar lipase in acidolysis. J. Ferment. Bioeng 1997, 83, 321–327. [Google Scholar]
- Du, W.; Xu, Y.Y.; Zeng, J.; Liu, D.H. Novozym 435-catalysed transesterification of crude soya bean oils for biodiesel production in a solvent-free medium. Biotechnol. Appl. Biochem 2004, 40, 187–190. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification steps | Total activity (U) | Total protein (mg) | Specific activity (U/mg) | Yield (%) | Purification factor |
|---|---|---|---|---|---|
| Supernatant | 2,100,000 | 950 | 2,210 | 100 | 1 |
| Ultrafiltration | 1,974,000 | 740 | 2,667 | 94 | 1.20 |
| SP-Sepharose FF | 1,512,000 | 226 | 6,690 | 72 | 3.02 |
| Metal ions (1 mM) | Relative activity (%) | Metal ions (1 mM) | Relative activity (%) |
|---|---|---|---|
| Control | 100 | Control | 100 |
| Ca2+ | 111 ± 2 | Co2+ | 84 ± 3 |
| Zn2+ | 42 ± 1 | K+ | 106 ± 3 |
| Li+ | 89 ± 3 | Na+ | 105 ± 4 |
| Fe3+ | 76 ± 3 | Hg+ | 0 |
| Mn2+ | 88 ± 4 | Ag+ | 5 ± 1 |
| Mg2+ | 107 ± 3 | EDTA | 90 ± 5 |
| Organic solvents | Relative activity (%) | Organic solvents | Relative activity (%) |
|---|---|---|---|
| Control | 100 | Control | 100 |
| Methanol | 22 ± 3 | Acetone | 27 ± 3 |
| Ethanol | 14 ± 1 | Butanol | 11 ± 2 |
| Isopropanol | 17 ± 1 | Chloroform | 16 ± 2 |
| Hexane | 61 ± 4 | Glycerol | 104 ± 6 |
| Solvent concentrations (%) | Relative activity (%) | |
|---|---|---|
| Methanol | Ethanol | |
| 0 | 100 | 100 |
| 5 | 98 ± 5 | 101 ± 3 |
| 10 | 97 ± 4 | 88 ± 4 |
| 20 | 91 ± 3 | 81 ± 4 |
| 30 | 61 ± 4 | 41 ± 3 |
| 40 | 22 ± 1 | 16 ± 2 |
| Substrates | Relative activity (%) | Substrates | Relative activity (%) |
|---|---|---|---|
| Olive oil | 100 | Olive oil | 100 |
| Tributyrin (C4) | 23 ± 2 | Methyl butyrate (C4) | 5 ± 2 |
| Tricaprylin (C8) | 134 ± 6 | Methyl caprylin (C8) | 8 ± 2 |
| Trilaurin (C12) | 156 ± 4 | Methyl laurate (C12) | 9 ± 1 |
| Tripalmitin (C16) | 178 ± 4 | Methyl palmitate (C16) | 11 ± 2 |
| Tristearin (C18) | 45 ± 3 | Methyl stearate (C18) | 10 ± 2 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, J.-R.; Li, Y.-Y.; Xu, S.-D.; Li, P.; Liu, J.-S.; Liu, D.-N. High-Level Expression of Pro-Form Lipase from Rhizopus oryzae in Pichia pastoris and Its Purification and Characterization. Int. J. Mol. Sci. 2014, 15, 203-217. https://doi.org/10.3390/ijms15010203
Wang J-R, Li Y-Y, Xu S-D, Li P, Liu J-S, Liu D-N. High-Level Expression of Pro-Form Lipase from Rhizopus oryzae in Pichia pastoris and Its Purification and Characterization. International Journal of Molecular Sciences. 2014; 15(1):203-217. https://doi.org/10.3390/ijms15010203
Chicago/Turabian StyleWang, Jian-Rong, Yang-Yuan Li, Shu-De Xu, Peng Li, Jing-Shan Liu, and Dan-Ni Liu. 2014. "High-Level Expression of Pro-Form Lipase from Rhizopus oryzae in Pichia pastoris and Its Purification and Characterization" International Journal of Molecular Sciences 15, no. 1: 203-217. https://doi.org/10.3390/ijms15010203