SnoRNA U50 Levels Are Regulated by Cell Proliferation and rRNA Transcription



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
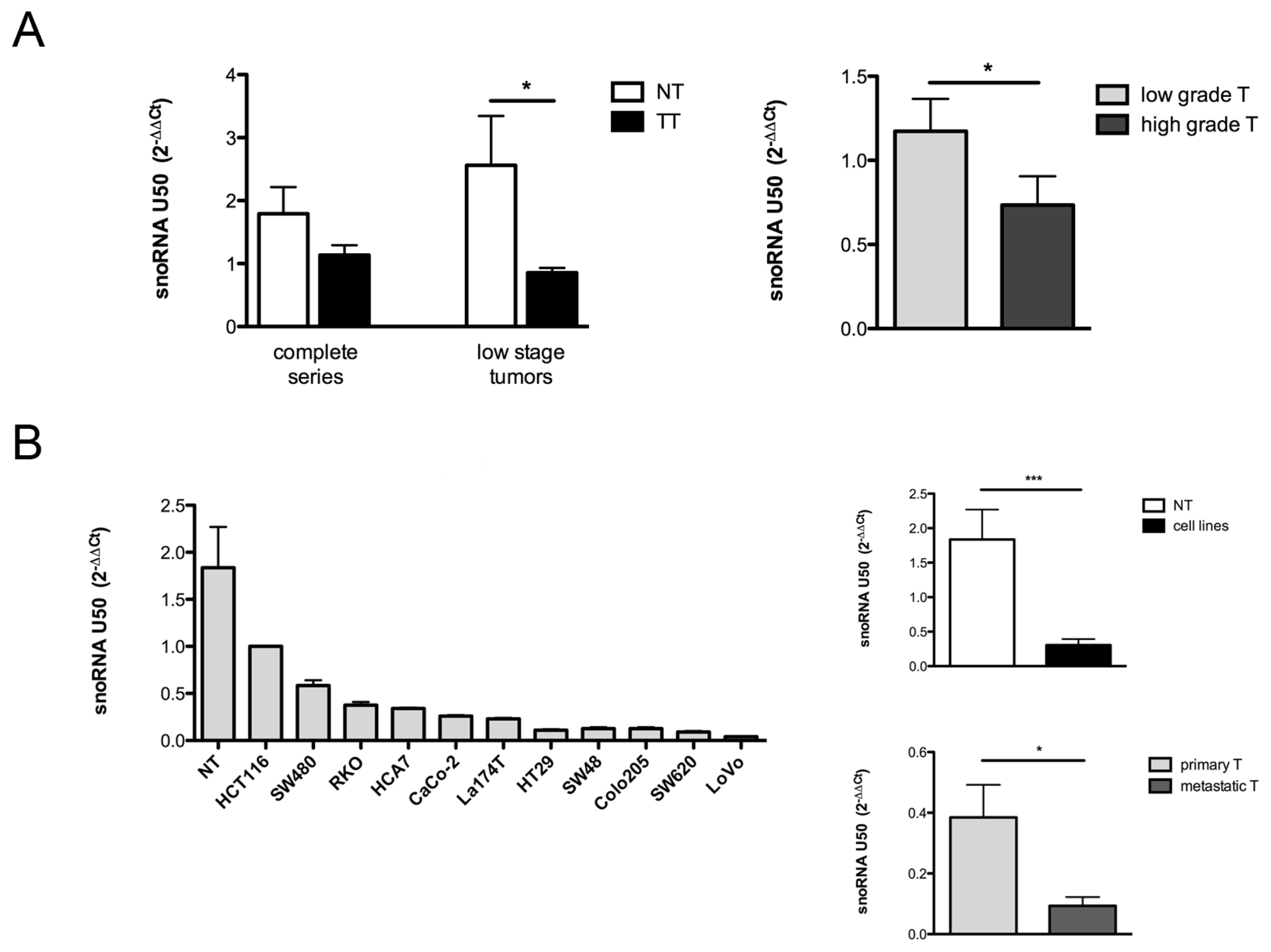
2.1. Evaluation of U50 Levels in Colorectal Cancer Tissues and Cell Lines
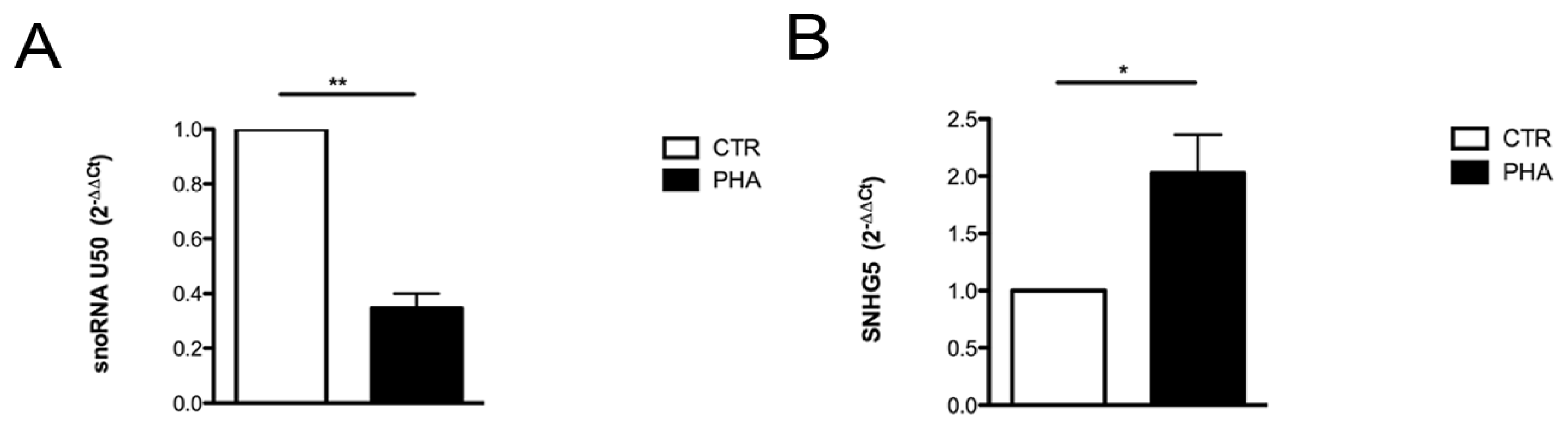
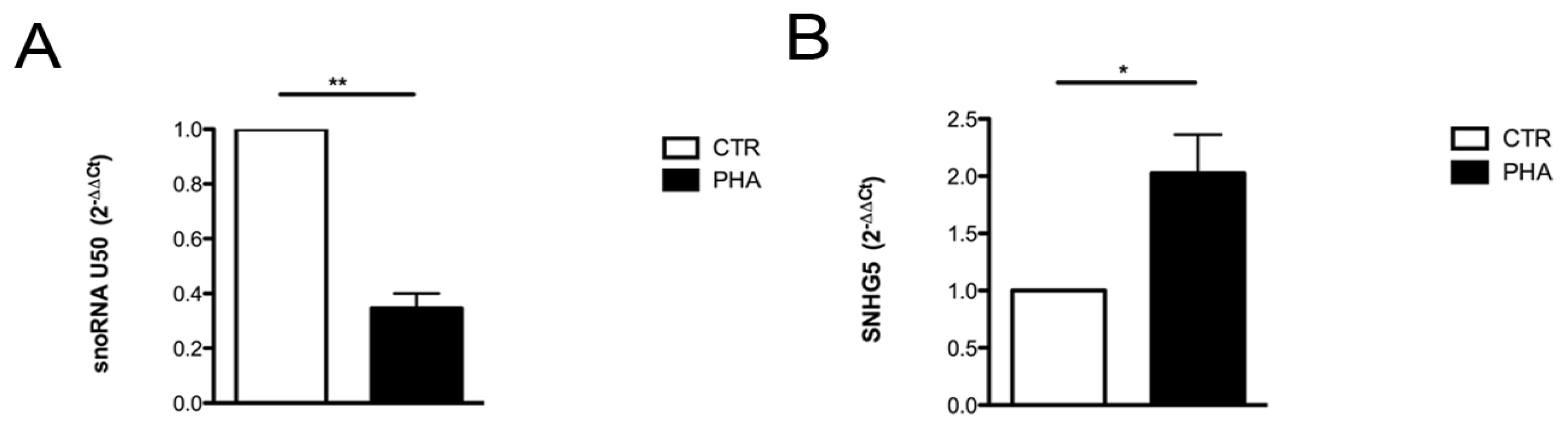
2.2. Relationship between U50 and Proliferation
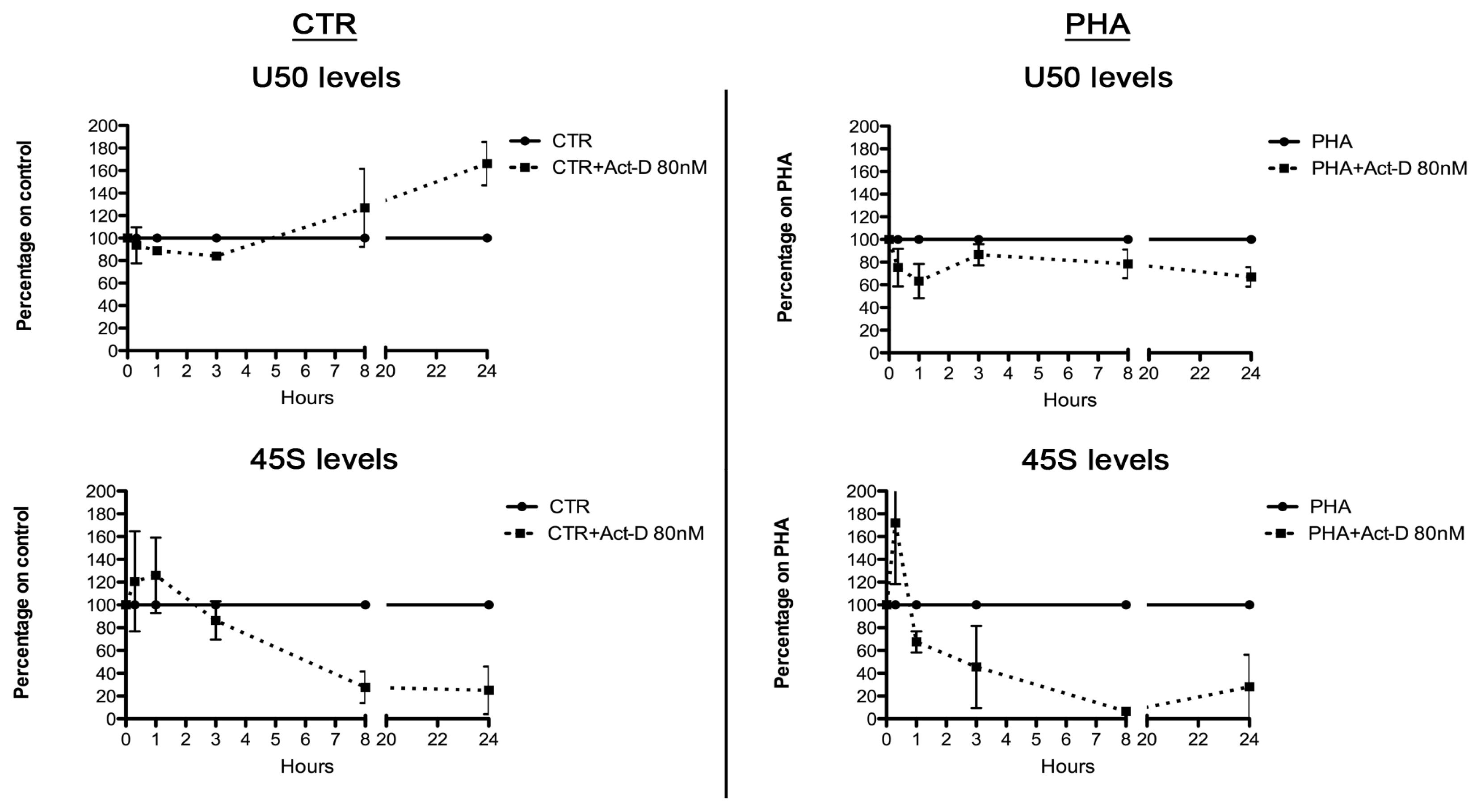
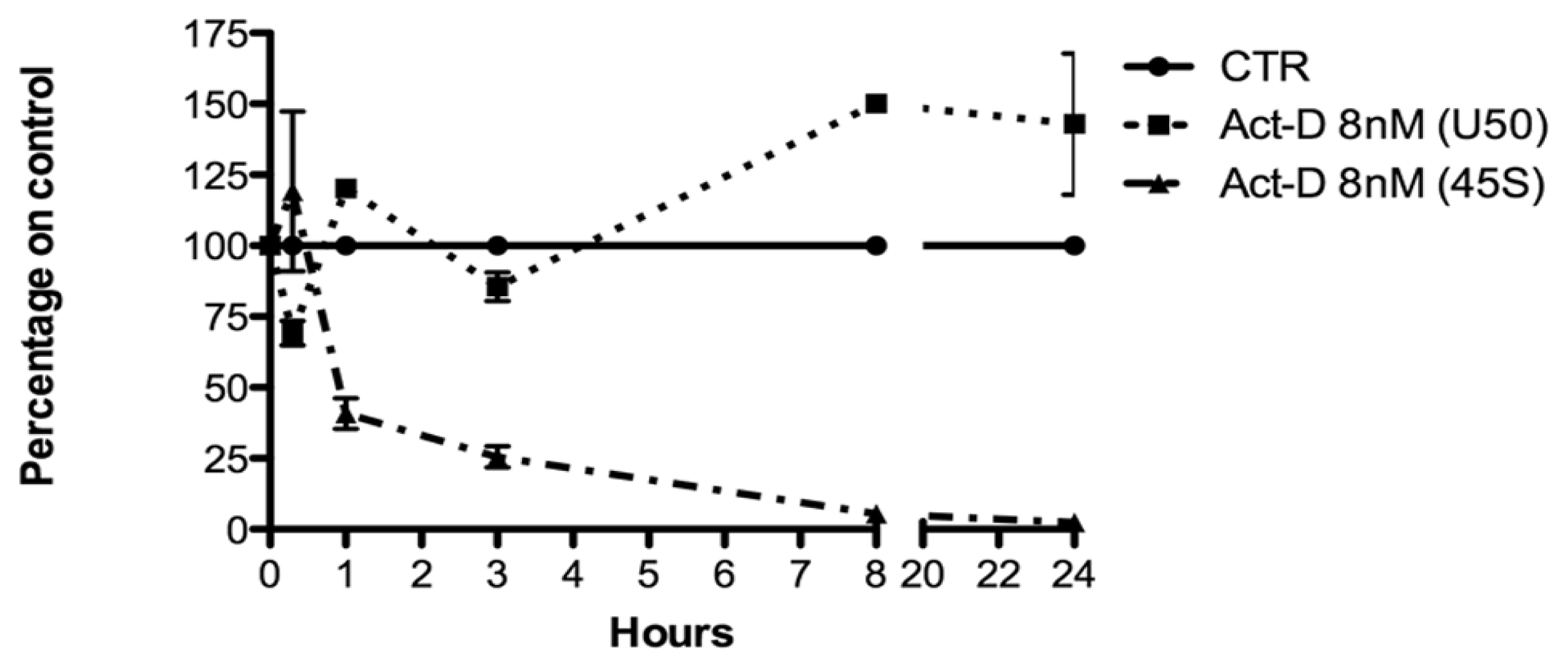
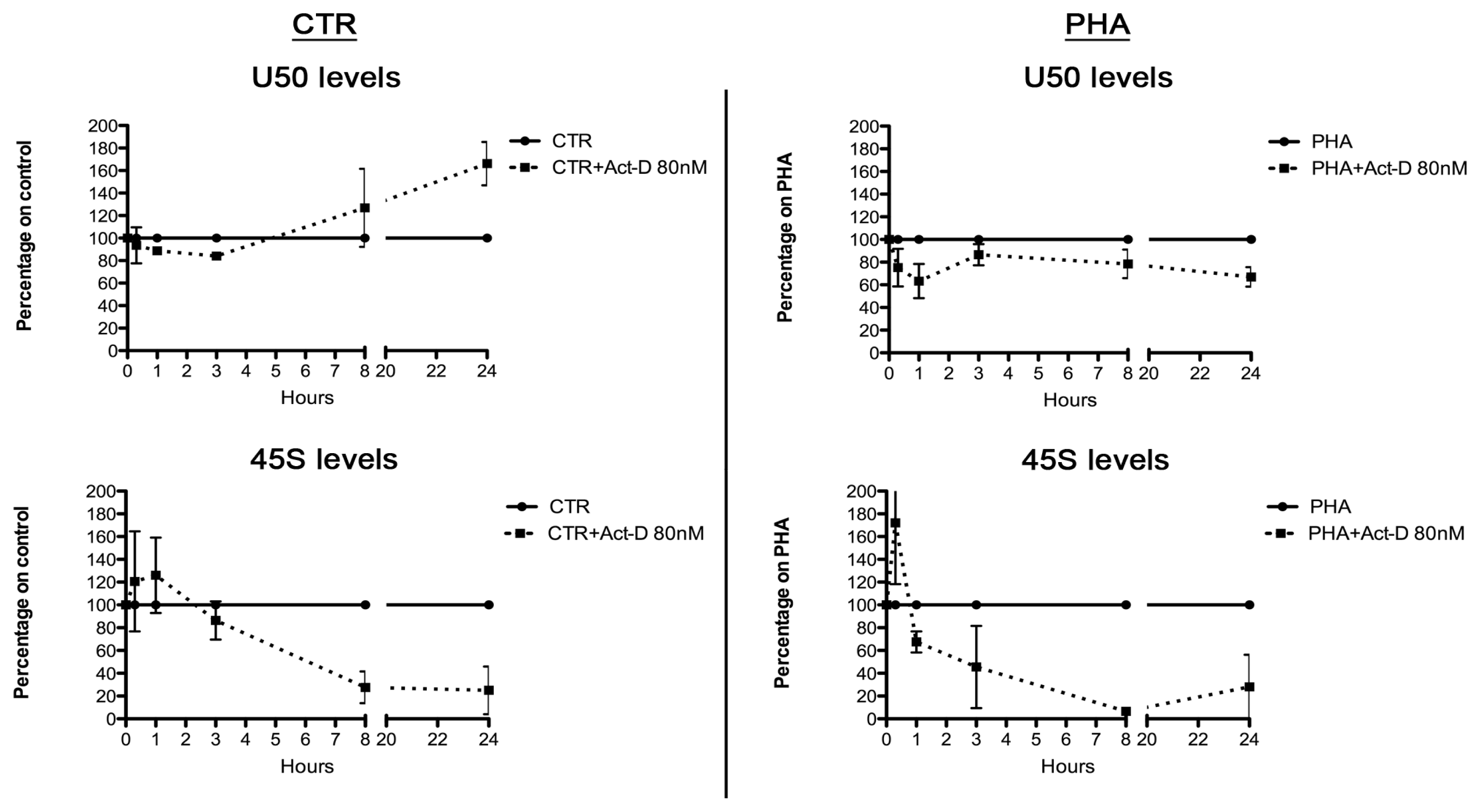
2.3. Relationship between U50 and rRNA Transcription
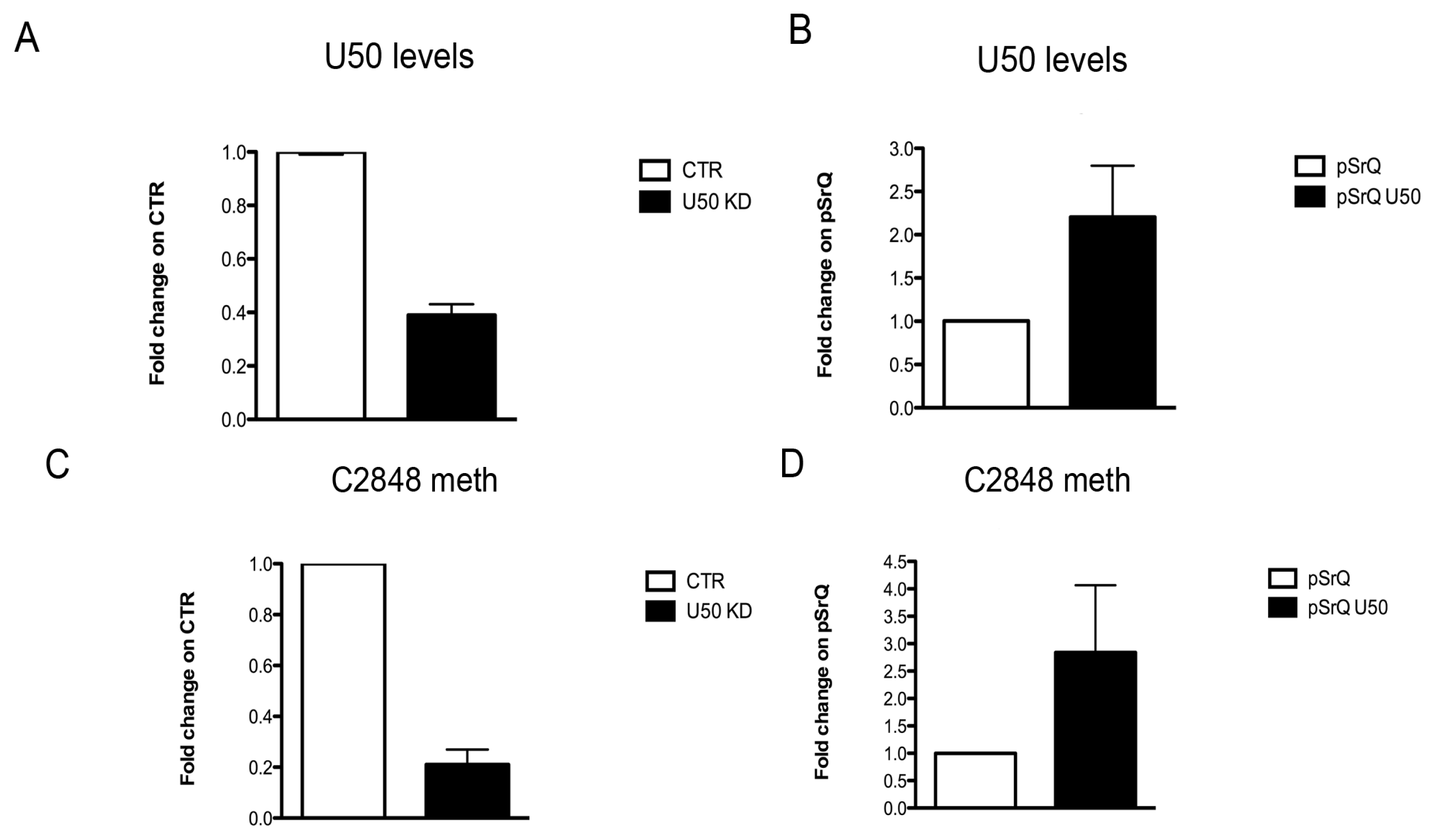
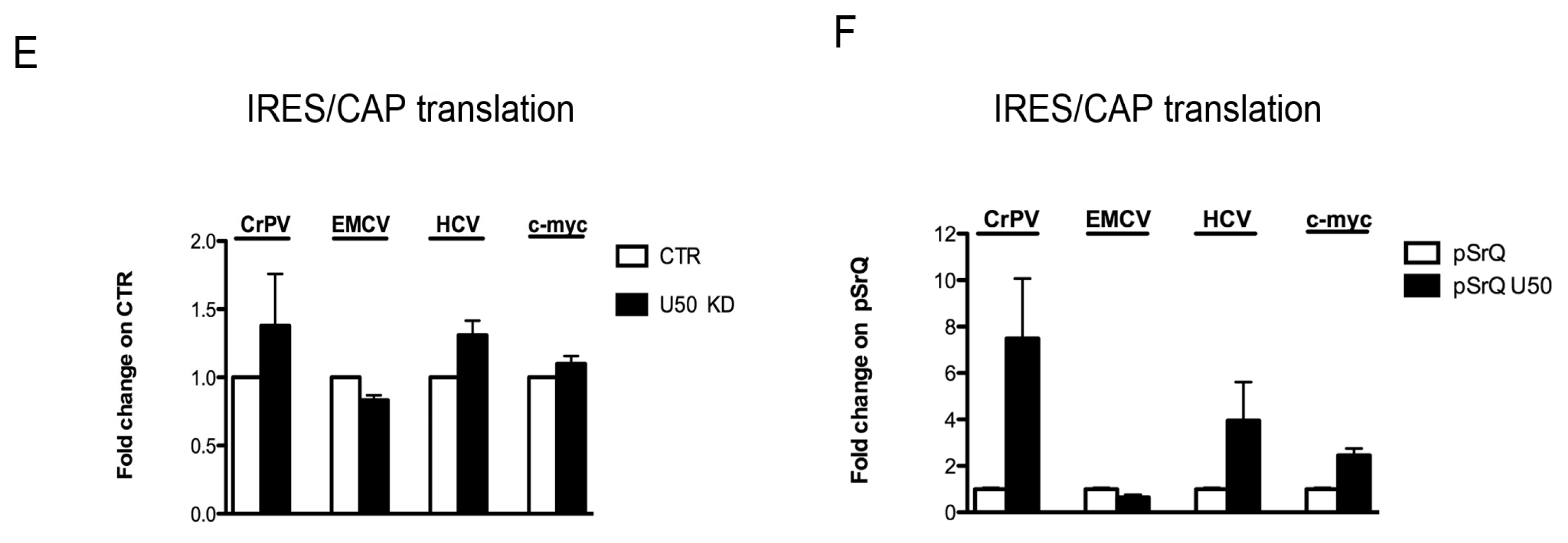
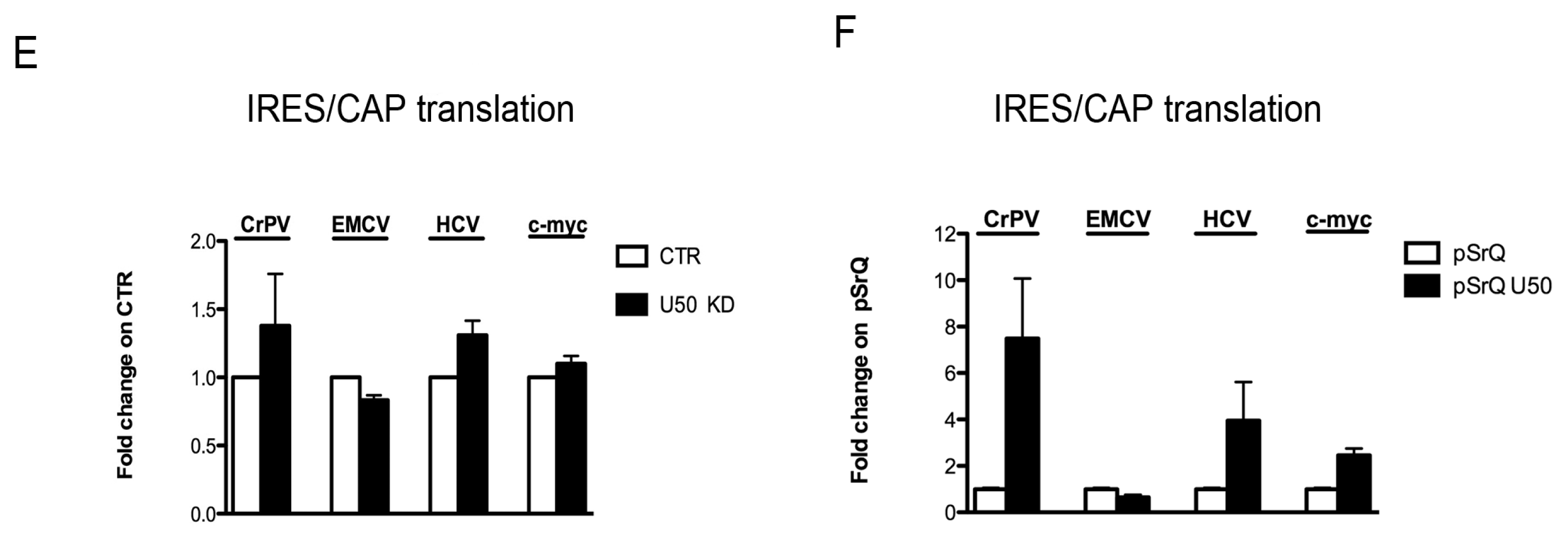
2.4. Effect of U50 Regulation on rRNA Site-Specific Methylation and Ribosome Activity
3. Experimental Section
3.1. Patient Materials
3.2. Cell Cultures and Treatments
3.3. Gene Expression Assays
3.4. SnoRNA U50 Knockdown and Upregulation
3.5. Semi-Quantitative Site-Specific Methylation Assay
3.6. mRNA Transfection and Internal Ribosome Entry Site (IRES) Translation
3.7. Statistical Analysis
4. Conclusions
Supplementary Information
ijms-14-14923-s001.pdfAcknowledgements
Conflict of Interest
References
- Filipowicz, W.; Pogacić, V. Biogenesis of small nucleolar ribonucleoproteins. Curr. Opin. Cell Biol 2002, 14, 319–327. [Google Scholar]
- Smith, C.M.; Steitz, J.A. Sno storm in the nucleolus: New roles for myriad small RNPs. Cell 1997, 89, 669–672. [Google Scholar]
- Ni, J.; Tien, A.L.; Fournier, M.J. Small nucleolar RNAs direct site-specific synthesis of pseudouridine in ribosomal RNA. Cell 1997, 89, 565–573. [Google Scholar]
- Kiss, T. Small nucleolar RNAs: An abundant group of noncoding RNAs with diverse cellular functions. Cell 2002, 109, 145–148. [Google Scholar]
- Kiss-Laszlo, Z.; Henry, Y.; Bachellerie, J.P.; Caizergues-Ferrer, M.; Kiss, T. Site-specific ribose methylation of preribosomal RNA: A novel function for small nucleolar RNAs. Cell 1996, 85, 1077–1088. [Google Scholar]
- Cavaille, J.; Bachellerie, J.P. SnoRNA-guided ribose methylation of rRNA: Structural features of the guide RNA duplex influencing the extent of the reaction. Nucleic Acids Res 1998, 26, 1576–1587. [Google Scholar]
- Bachellerie, J.P.; Michot, B.; Nicoloso, M.; Balakin, A.; Ni, J.; Fournier, M.J. Antisense snoRNAs: A family of nucleolar RNAs with long complementarities to rRNA. Trends Biochem. Sci 1995, 20, 261–264. [Google Scholar]
- Kiss, T. Small nucleolar RNA-guided posttranscriptional modification of cellular RNAs. EMBO J 2001, 20, 3617–3622. [Google Scholar]
- Weinstein, L.B.; Steitz, J.A. Guided tours: From precursor snoRNA to functional snoRNP. Curr. Opin. Cell Biol 1999, 11, 378–384. [Google Scholar]
- Meyuhas, O.; Avni, D.; Shama, S. Translational Control of Ribosomal Protein mRNAs in Eukaryotes. In Translational Control; Hershey, J.W.B., Mathews, M.B., Sonenberg, N., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1996; Volume 30, pp. 363–388. [Google Scholar]
- Smith, C.M.; Steitz, J.A. Classification of gas5 as a multi-small-nucleolar-RNA (snoRNA) host gene and a member of the 5′-terminal oligopyrimidine gene family reveals common features of snoRNA host genes. Mol. Cell Biol 1998, 18, 6897–6909. [Google Scholar]
- Decatur, W.A.; Fournier, M.J. rRNA modifications and ribosome function. Trends Biochem. Sci 2002, 27, 344–351. [Google Scholar]
- Yusupov, M.M.; Yusupova, G.Z.; Baucom, A.; Lieberman, K.; Earnest, T.N.; Cate, J.H.; Noller, H.F. Crystal structure of the ribosome at 5.5 Å resolution. Science 2001, 292, 883–896. [Google Scholar]
- Lane, B.G.; Ofengand, J.; Gray, M.W. Pseudouridine and O2′-methylated nucleosides. Significance of their selective occurrence in rRNA domains that function in ribosome-catalyzed synthesis of the peptide bonds in proteins. Biochimie 1995, 77, 7–15. [Google Scholar]
- Green, R.; Noller, H.F. Reconstitution of functional 50S ribosomes from in vitro transcripts of Bacillus stearothermophilus 23S rRNA. Biochemistry 1999, 38, 1772–1779. [Google Scholar]
- Khaitovich, P.; Tenson, T.; Kloss, P.; Mankin, A.S. Reconstitution of functionally active Thermus aquaticus large ribosomal subunits with in vitro-transcribed rRNA. Biochemistry 1999, 38, 1780–1788. [Google Scholar]
- Liu, B.; Liang, X.H.; Piekna-Przybylska, D.; Liu, Q.; Fournier, M.J. Mis-targeted methylation in rRNA can severely impair ribosome synthesis and activity. RNA Biol 2008, 5, 249–254. [Google Scholar]
- Montanaro, L.; Calienni, M.; Bertoni, S.; Rocchi, L.; Sansone, P.; Storci, G.; Santini, D.; Ceccarelli, C.; Taffurelli, M.; Carnicelli, D.; et al. Novel dyskerin-mediated mechanism of p53 inactivation through defective mRNA translation. Cancer Res 2010, 70, 4767–4777. [Google Scholar]
- Belin, S.; Beghin, A.; Solano-Gonzàlez, E.; Bezin, L.; Brunet-Manquat, S.; Textoris, J.; Prats, A.C.; Mertani, H.C.; Dumontet, C.; Diaz, J.J. Dysregulation of ribosome biogenesis and translational capacity is associated with tumor progression of human breast cancer cells. PLoS One 2009, 4. [Google Scholar] [CrossRef]
- Tanaka, R.; Satoh, H.; Moriyama, M.; Satoh, K.; Morishita, Y.; Yoshida, S.; Watanabe, T.; Nakamura, Y.; Mori, S. Intronic U50 small-nucleolar-RNA (snoRNA) host gene of no protein-coding potential is mapped at the chromosome breakpoint t(3;6)(q27; q15) of human B-cell lymphoma. Genes Cells 2000, 5, 277–287. [Google Scholar]
- Dong, X.Y.; Rodriguez, C.; Guo, P.; Sun, X.; Talbot, J.T.; Zhou, W.; Petros, J.; Li, Q.; Vessella, R.L.; Kibel, A.S.; et al. SnoRNA U50 is a candidate tumor-suppressor gene at 6q14.3 with a mutation associated with clinically significant prostate cancer. Hum. Mol. Genet 2008, 17, 1031–1042. [Google Scholar]
- Mourtada-Maarabouni, M.; Pickard, M.R.; Hedge, V.L.; Farzaneh, F.; Williams, G.T. GAS5, a non-protein-coding RNA, controls apoptosis and is downregulated in breast cancer. Oncogene 2009, 28, 195–208. [Google Scholar]
- Dong, X.Y.; Guo, P.; Boyd, J.; Sun, X.; Li, Q.; Zhou, W.; Dong, J.T. Implication of snoRNA U50 in human breast cancer. J. Genet. Genomics 2009, 36, 447–454. [Google Scholar]
- Nakamura, Y.; Takahashi, N.; Kakegawa, E.; Yoshida, K.; Ito, Y.; Kayano, H.; Niitsu, N.; Jinnai, I.; Bessho, M. The GAS5 (growth arrest-specific transcript 5) gene fuses to BCL6 as a result of t(1;3)(q25;q27) in a patient with B-cell lymphoma. Cancer Genet. Cytogenet 2008, 182, 144–149. [Google Scholar]
- Basu, A.; Das, P.; Chaudhuri, S.; Bevilacqua, E.; Andrews, J.; Barik, S.; Hatzoglou, M.; Komar, A.A.; Mazumder, B. Requirement of rRNA methylation for 80S ribosome assembly on a cohort of cellular internal ribosome Entry Sites. Mol. Cell Biol 2011, 31, 4482–4499. [Google Scholar]
- Nicoloso, M.; Qu, L.H.; Michot, B.; Bachellerie, J.P. Intron-encoded, antisense small nucleolar RNAs: The characterization of nine novel species points to their direct role as guides for the 2′-O-ribose methylation of rRNAs. J. Mol. Biol 1996, 260, 178–195. [Google Scholar]
- Maxwell, E.S.; Fournier, M.J. The small nucleolar RNAs. Annu. Rev. Biochem 1995, 64, 897–934. [Google Scholar]
- Maquat, L.E. When cells stop making sense: Effects of nonsense codons on RNA metabolism in vertebrate cells. RNA 1995, 1, 453–465. [Google Scholar]
- Kino, T.; Hurt, D.E.; Ichijo, T.; Nader, N.; Chrousos, G.P. Noncoding RNA Gas5 is a growth arrest and starvation-associated repressor of the glucocorticoid receptor. Sci. Signal 2010, 3. [Google Scholar] [CrossRef]
- Ploner, A.; Ploner, C.; Lukasser, M.; Niederegger, H.; Hüttenhofer, A. Methodological obstacles in knocking down small noncoding RNAs. RNA 2009, 15, 1797–1804. [Google Scholar]
- Yoon, A.; Peng, G.; Brandenburger, Y.; Zollo, O.; Xu, W.; Rego, E.; Ruggero, D. Impaired control of IRES-mediated translation in X-linked dyskeratosis congenita. Science 2006, 312, 902–906. [Google Scholar]
- Bochkov, Y.A.; Palmenberg, A.C. Translational efficiency of EMCV IRES in bicistronic vectors is dependent upon IRES sequence and gene location. Biotechniques 2006, 41, 283–284. [Google Scholar]
- Van Eden, M.E.; Byrd, M.P.; Sherrill, K.W.; Lloyd, R.E. Demonstrating internal ribosome entry sites in eukaryotic mRNAs using stringent RNA test procedures. RNA 2004, 10, 720–730. [Google Scholar]
- Thoma, C.; Fraterman, S.; Gentzel, M.; Wilm, M.; Hentze, M.W. Translation initiation by the c-Myc mRNA internal ribosome entry sequence and the poly(A) tail. RNA 2008, 14, 1579–1589. [Google Scholar]



© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pacilli, A.; Ceccarelli, C.; Treré, D.; Montanaro, L. SnoRNA U50 Levels Are Regulated by Cell Proliferation and rRNA Transcription. Int. J. Mol. Sci. 2013, 14, 14923-14935. https://doi.org/10.3390/ijms140714923
Pacilli A, Ceccarelli C, Treré D, Montanaro L. SnoRNA U50 Levels Are Regulated by Cell Proliferation and rRNA Transcription. International Journal of Molecular Sciences. 2013; 14(7):14923-14935. https://doi.org/10.3390/ijms140714923
Chicago/Turabian StylePacilli, Annalisa, Claudio Ceccarelli, Davide Treré, and Lorenzo Montanaro. 2013. "SnoRNA U50 Levels Are Regulated by Cell Proliferation and rRNA Transcription" International Journal of Molecular Sciences 14, no. 7: 14923-14935. https://doi.org/10.3390/ijms140714923