Quorum Quenching in Culturable Phyllosphere Bacteria from Tobacco


Abstract
:1. Introduction
2. Results and Discussion
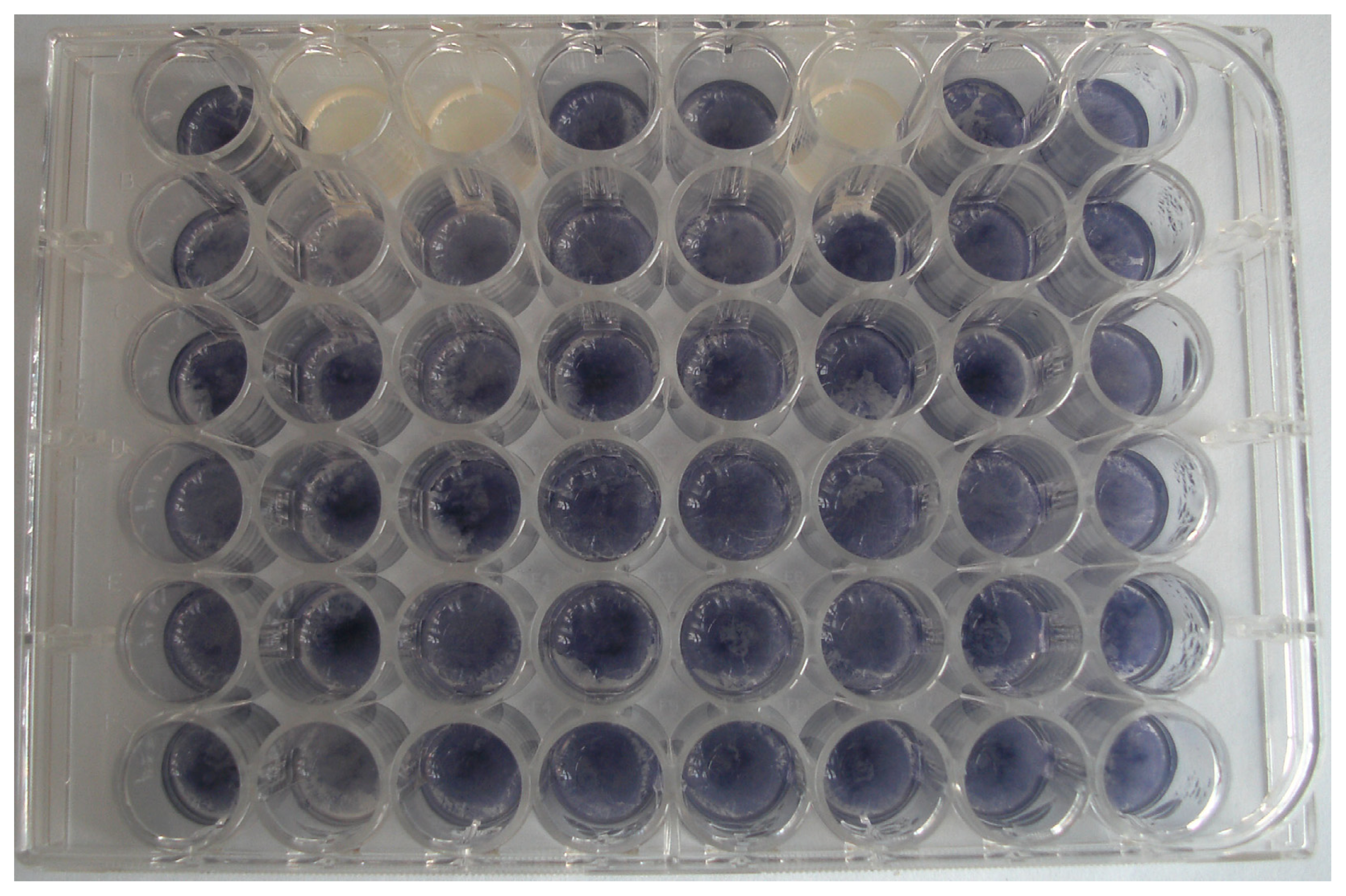
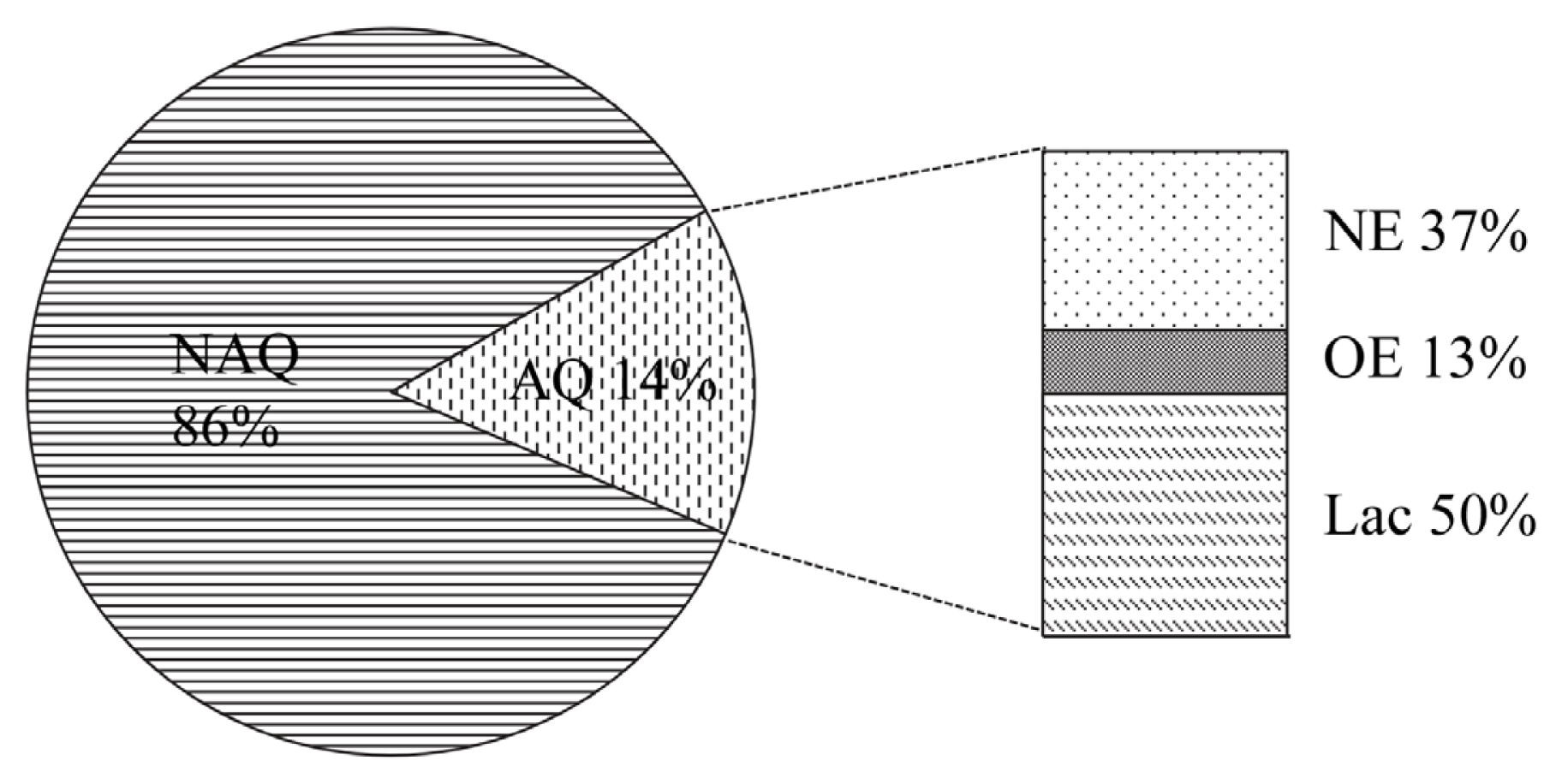
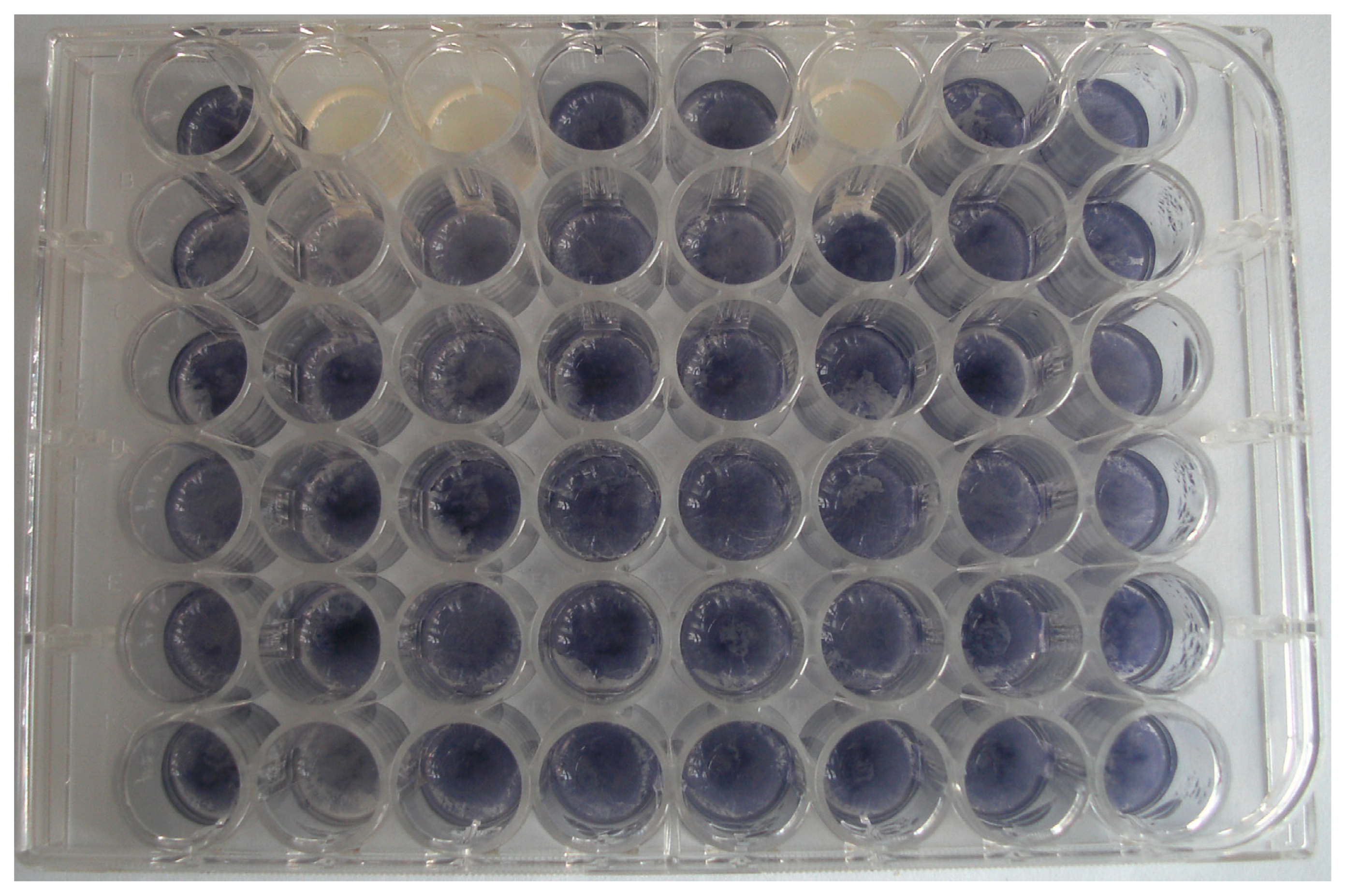
2.1. Isolation of Bacteria with QQ Activity
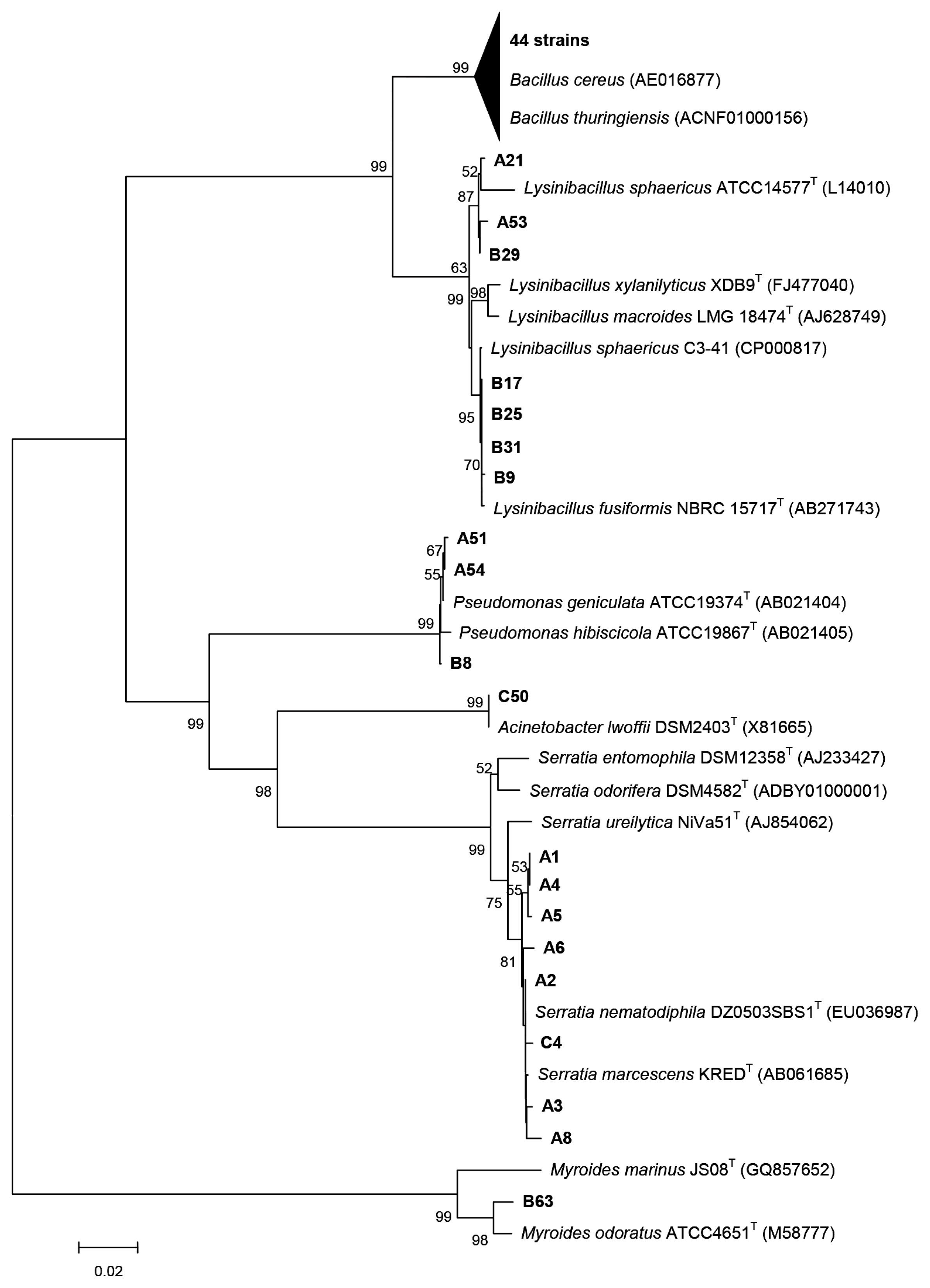
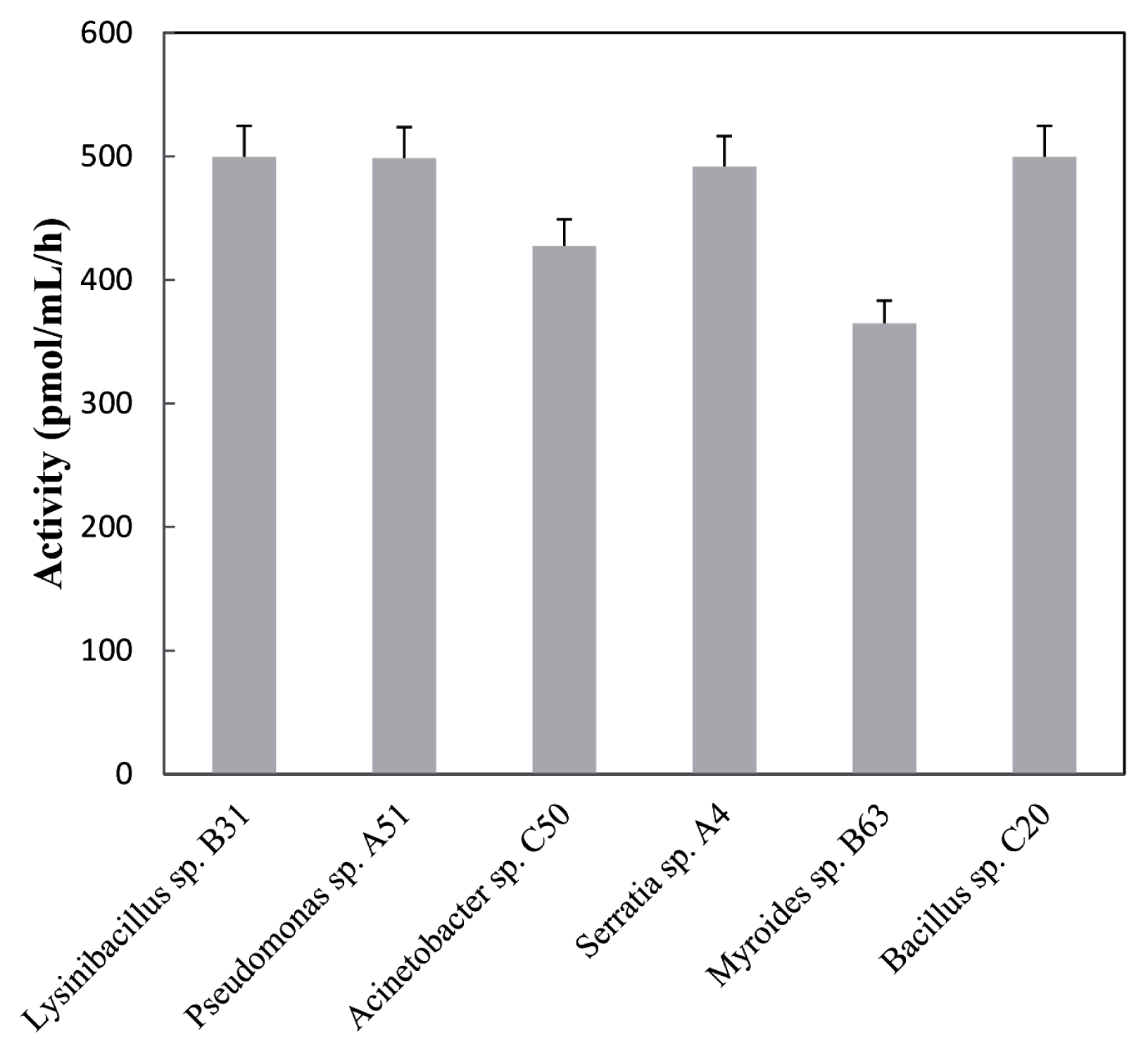
2.2. Identification and Characterization of Bacteria with AHL-Degrading Activity
3. Experimental Section
3.1. Isolation of Leaf Associated Culturable Bacteria
3.2. Screening for Bacteria with QQ Activity
3.3. PCR Amplification and Sequence Analyses
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol 2005, 21, 319–346. [Google Scholar]
- Pacheco, A.R.; Sperandio, V. Inter-kingdom signaling: Chemical language between bacteria and host. Curr. Opin. Microbiol 2009, 12, 192–198. [Google Scholar]
- Manefield, M.; Whiteley, A.S. Acylated homoserine lactones in the environment: Chameleons of bioactivity. Phil. Trans. R. Soc. B 2007, 362, 1235–1240. [Google Scholar]
- Loh, J.; Pierson, E.A.; Pierson, L.S., III; Stacey, G.; Chatterjee, A. Quorum sensing in plant-associated bacteria. Curr. Opin. Plant Biol. 2002, 5, 285–290. [Google Scholar]
- Cha, C.; Gao, P.; Chen, Y.C.; Shaw, P.D.; Farrand, S.K. Production of acyl-homoserine lactone quorum-sensing signals by gram-negative plant-associated bacteria. Mol. Plant Microbe Interact 1998, 11, 1119–1129. [Google Scholar]
- Quinones, B.; Dulla, G.; Lindow, S.E. Quorum sensing regulates exopolysaccharide production, motility, and virulence in Pseudomonas syringae. Mol. Plant Microbe Interact 2005, 18, 682–693. [Google Scholar]
- Morohoshi, T.; Nakamura, Y.; Yamazaki, G.; Ishida, A.; Kato, N.; Ikeda, T. The plant pathogen Pantoea ananatis produces N-acylhomoserine lactone and causes center rot disease of onion by quorum sensing. J. Bacteriol 2007, 189, 8333–8338. [Google Scholar]
- Vonbodman, S.B.; Farrand, S.K. Capsular polysaccharide biosynthesis and pathogenicity in Erwinia stewartii require induction by an N-acylhomoserine lactone autoinducer. J. Bacteriol 1995, 177, 5000–5008. [Google Scholar]
- Dong, Y.H.; Wang, L.H.; Zhang, L.H. Quorum-quenching microbial infections: Mechanisms and implications. Phil. Trans. R. Soc. B 2007, 362, 1201–1211. [Google Scholar]
- Uroz, S.; Dessaux, Y.; Oger, P. Quorum sensing and quorum quenching: The yin and yang of bacterial communication. ChemBioChem 2009, 10, 205–216. [Google Scholar]
- Dong, Y.H.; Wang, L.H.; Xu, J.L.; Zhang, H.B.; Zhang, X.F.; Zhang, L.H. Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature 2001, 411, 813–817. [Google Scholar]
- Varzakas, T.H.; Arvanitoyannis, I.S.; Baltas, H. The politics and science behind GMO acceptance. Crit. Rev. Food Sci. Nutr 2007, 47, 335–361. [Google Scholar]
- Molina, L.; Constantinescu, F.; Michel, L.; Reimmann, C.; Duffy, B.; Defago, G. Degradation of pathogen quorum-sensing molecules by soil bacteria: A preventive and curative biological control mechanism. FEMS Microbiol. Ecol 2003, 45, 71–81. [Google Scholar]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol 2003, 69, 1875–1883. [Google Scholar]
- Diallo, S.; Crepin, A.; Barbey, C.; Orange, N.; Burini, J.F.; Latour, X. Mechanisms and recent advances in biological control mediated through the potato rhizosphere. FEMS Microbiol. Ecol 2011, 75, 351–364. [Google Scholar]
- Dong, Y.H.; Xu, J.L.; Li, X.Z.; Zhang, L.H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar]
- D’Angelo-Picard, C.; Faure, D.; Penot, I.; Dessaux, Y. Diversity of N-acyl homoserine lactone-producing and -degrading bacteria in soil and tobacco rhizosphere. Environ. Microbiol 2005, 7, 1796–1808. [Google Scholar]
- Schipper, C.; Hornung, C.; Bijtenhoorn, P.; Quitschau, M.; Grond, S.; Streit, W.R. Metagenome-derived clones encoding two novel lactonase family proteins involved in biofilm inhibition in Pseudomonas aeruginosa. Appl. Environ. Microbiol 2009, 75, 224–233. [Google Scholar]
- Hunter, P.J.; Hand, P.; Pink, D.; Whipps, J.M.; Bending, G.D. Both leaf properties and microbe-microbe interactions influence within-species variation in bacterial population diversity and structure in the lettuce (Lactuca Species) phyllosphere. Appl. Environ. Microbiol 2010, 76, 8117–8125. [Google Scholar]
- Huang, J.J.; Han, J.I.; Zhang, L.H.; Leadbetter, J.R. Utilization of acyl-homoserine lactone quorum signals for growth by a soil pseudomonad and Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol 2003, 69, 5941–5949. [Google Scholar]
- Decho, A.W.; Visscher, P.T.; Ferry, J.; Kawaguchi, T.; He, L.J.; Przekop, K.M.; Norman, R.S.; Reid, R.P. Autoinducers extracted from microbial mats reveal a surprising diversity of N-acylhomoserine lactones (AHLs) and abundance changes that may relate to diel pH. Environ. Microbiol 2009, 11, 409–420. [Google Scholar]
- Lyon, G.J.; Mayville, P.; Muir, T.W.; Novick, R.P. Rational design of a global inhibitor of the virulence response in Staphylococcus aureus, based in part on localization of the site of inhibition to the receptor-histidine kinase, AgrC. Proc. Natl. Acad. Sci. USA 2000, 97, 13330–13335. [Google Scholar]
- Chernin, L.; Toklikishvili, N.; Ovadis, M.; Kim, S.; Ben-Ari, J.; Khmel, I.; Vainstein, A. Quorum-sensing quenching by rhizobacterial volatiles. Environ. Microbiol. Rep 2011, 3, 698–704. [Google Scholar]
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv 2013, 31, 224–245. [Google Scholar]
- Uroz, S.; Heinonsalo, J. Degradation of N-acyl homoserine lactone quorum sensing signal molecules by forest root-associated fungi. FEMS Microbiol. Ecol 2008, 65, 271–278. [Google Scholar]
- Lee, S.J.; Park, S.Y.; Lee, J.J.; Yum, D.Y.; Koo, B.T.; Lee, J.K. Genes encoding the N-acyl homoserine lactone-degrading enzyme are widespread in many subspecies of Bacillus thuringiensis. Appl. Environ. Microbiol 2002, 68, 3919–3924. [Google Scholar]
- Redford, A.J.; Bowers, R.M.; Knight, R.; Linhart, Y.; Fierer, N. The ecology of the phyllosphere: Geographic and phylogenetic variability in the distribution of bacteria on tree leaves. Environ. Microbiol 2010, 12, 2885–2893. [Google Scholar]
- Dong, Y.H.; Gusti, A.R.; Zhang, Q.; Xu, J.L.; Zhang, L.H. Identification of quorum-quenching N-acyl homoserine lactonases from Bacillus species. Appl. Environ. Microbiol 2002, 68, 1754–1759. [Google Scholar]
- Dong, Y.H.; Zhang, X.F.; Xu, J.L.; Zhang, L.H. Insecticidal Bacillus thuringiensis silences Erwinia carotovora virulence by a new form of microbial antagonism, signal interference. Appl. Environ. Microbiol 2004, 70, 954–960. [Google Scholar]
- Melnick, R.L.; Suarez, C.; Bailey, B.A.; Backman, P.A. Isolation of endophytic endospore-forming bacteria from Theobroma cacao as potential biological control agents of cacao diseases. Biol. Control 2011, 57, 236–245. [Google Scholar]
- Van Houdt, R.; Givskov, M.; Michiels, C.W. Quorum sensing in Serratia. FEMS Microbiol. Rev 2007, 31, 407–424. [Google Scholar]
- Whitehead, N.A.; Barnard, A.M.L.; Slater, H.; Simpson, N.J.L.; Salmond, G.P.C. Quorum-sensing in gram-negative bacteria. FEMS Microbiol. Rev 2001, 25, 365–404. [Google Scholar]
- Akutsu, K.; Hirat, M.; Yamamota, M.; Hirayae, K.; Okuyama, S.; Hibi, T. Growth inhibition of Botrytis spp. by Serratia marcescens B2 isolated from tomato phylloplane. Ann. Rev. Phytopathol. Soc. Jpn 1993, 59, 18–25. [Google Scholar]
- Van Houdt, R.; Moons, P.; Aertsen, A.; Jansen, A.; Vanoirbeek, K.; Daykin, M.; Williams, P.; Michiels, C.W. Characterization of a luxI/luxR-type quorum sensing system and N-acyl-homoserine lactone-dependent regulation of exo-enzyme and antibacterial component production in Serratia plymuthica RVH1. Res. Microbiol 2007, 158, 150–158. [Google Scholar]
- Pang, Y.D.; Liu, X.G.; Ma, Y.X.; Chernin, L.; Berg, G.; Gao, K.X. Induction of systemic resistance, root colonisation and biocontrol activities of the rhizospheric strain of Serratia plymuthica are dependent on N-acyl homoserine lactones. Eur. J. Plant Pathol 2009, 124, 261–268. [Google Scholar]
- Al-Thani, R.F.; Abd-El-Haleem, D.A.M.; Al-Shammri, M. Isolation and characterization of polyaromatic hydrocarbons-degrading bacteria from different Qatari soils. Afr. J. Microbiol. Res 2009, 3, 761–766. [Google Scholar]
- Chan, K.G.; Atkinson, S.; Mathee, K.; Sam, C.K.; Chhabra, S.R.; Camara, M.; Koh, C.L.; Williams, P. Characterization of N-acylhomoserine lactone-degrading bacteria associated with the Zingiber officinale(ginger) rhizosphere: Co-existence of quorum quenching and quorum sensing in Acinetobacter and. Burkholderia. BMC Microbiol. 2011, 11. [Google Scholar] [CrossRef]
- Fravel, D.R. Commercialization and implementation of biocontrol. Annu. Rev. Phytopathol 2005, 43, 337–359. [Google Scholar]
- Francis, I.; Holsters, M.; Vereecke, D. The Gram-positive side of plant-microbe interactions. Environ. Microbiol 2010, 12, 1–12. [Google Scholar]
- Chankhamhaengdecha, S.; Hongvijit, S.; Srichaisupakit, A.; Charnchai, P.; Panbangred, W. Endophytic actinomycetes: A novel source of potential acyl homoserine lactone degrading enzymes. BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef]
- Riaz, K.; Elmerich, C.; Moreira, D.; Raffoux, A.; Dessaux, Y.; Faure, D. A metagenomic analysis of soil bacteria extends the diversity of quorum-quenching lactonases. Environ. Microbiol 2008, 10, 560–570. [Google Scholar]
- Huang, J.J.; Petersen, A.; Whiteley, M.; Leadbetter, J.R. Identification of QuiP, the product of gene PA1032, as the second acyl-homoserine lactone acylase of Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol 2006, 72, 1190–1197. [Google Scholar]
- Lv, D.; Ma, A.; Tang, X.; Bai, Z.; Qi, H.; Zhuang, G. Profile of the culturable microbiome capable of producing acyl-homoserine lactone in the tobacco phyllosphere. J. Environ. Sci 2013, 25, 357–366. [Google Scholar]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar]
- Brons, J.K.; van Ems, J.D. Analysis of bacterial communities in soil by use of denaturing gradient gel electrophoresis and clone libraries, as influenced by different reverse primers. Appl. Environ. Microbiol 2008, 74, 2717–2727. [Google Scholar]
- Ashelford, K.E.; Chuzhanova, N.A.; Fry, J.C.; Jones, A.J.; Weightman, A.J. New screening software shows that most recent large 16S rRNA gene clone libraries contain chimeras. Appl. Environ. Microbiol 2006, 72, 5734–5741. [Google Scholar]
- Chun, J.; Lee, J.H.; Jung, Y.; Kim, M.; Kim, S.; Kim, B.K.; Lim, Y.W. EzTaxon: A web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences. Int. J. Syst. Evol. Microbiol 2007, 57, 2259–2261. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol 2011, 28, 2731–2739. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Related bacteria | Similarity (%) | C6-HSL | 3OC6-HSL | aiiA |
|---|---|---|---|---|---|
| B31 | Lysinibacillus fusiformis NBRC15717 | 99.9 | +++ | +++ | NA |
| A51 | Pseudomonas geniculata ATCC19374 | 99.9 | +++ | +++ | NA |
| C50 | Acinetobacter lwoffii DSM2403 | 99.5 | ++ | ++ | NA |
| A4 | Serratia marcescens | 99.8 | +++ | +++ | NA |
| B63 | Myroides odoratus ATCC4651 | 98.4 | + | + | NA |
| C20 | Bacillus cereus ATCC14579 | 99.9 | +++ | +++ | + |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ma, A.; Lv, D.; Zhuang, X.; Zhuang, G. Quorum Quenching in Culturable Phyllosphere Bacteria from Tobacco. Int. J. Mol. Sci. 2013, 14, 14607-14619. https://doi.org/10.3390/ijms140714607
Ma A, Lv D, Zhuang X, Zhuang G. Quorum Quenching in Culturable Phyllosphere Bacteria from Tobacco. International Journal of Molecular Sciences. 2013; 14(7):14607-14619. https://doi.org/10.3390/ijms140714607
Chicago/Turabian StyleMa, Anzhou, Di Lv, Xuliang Zhuang, and Guoqiang Zhuang. 2013. "Quorum Quenching in Culturable Phyllosphere Bacteria from Tobacco" International Journal of Molecular Sciences 14, no. 7: 14607-14619. https://doi.org/10.3390/ijms140714607