Uterine Micro-Environment and Estrogen-Dependent Regulation of Osteopontin Expression in Mouse Blastocyst

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
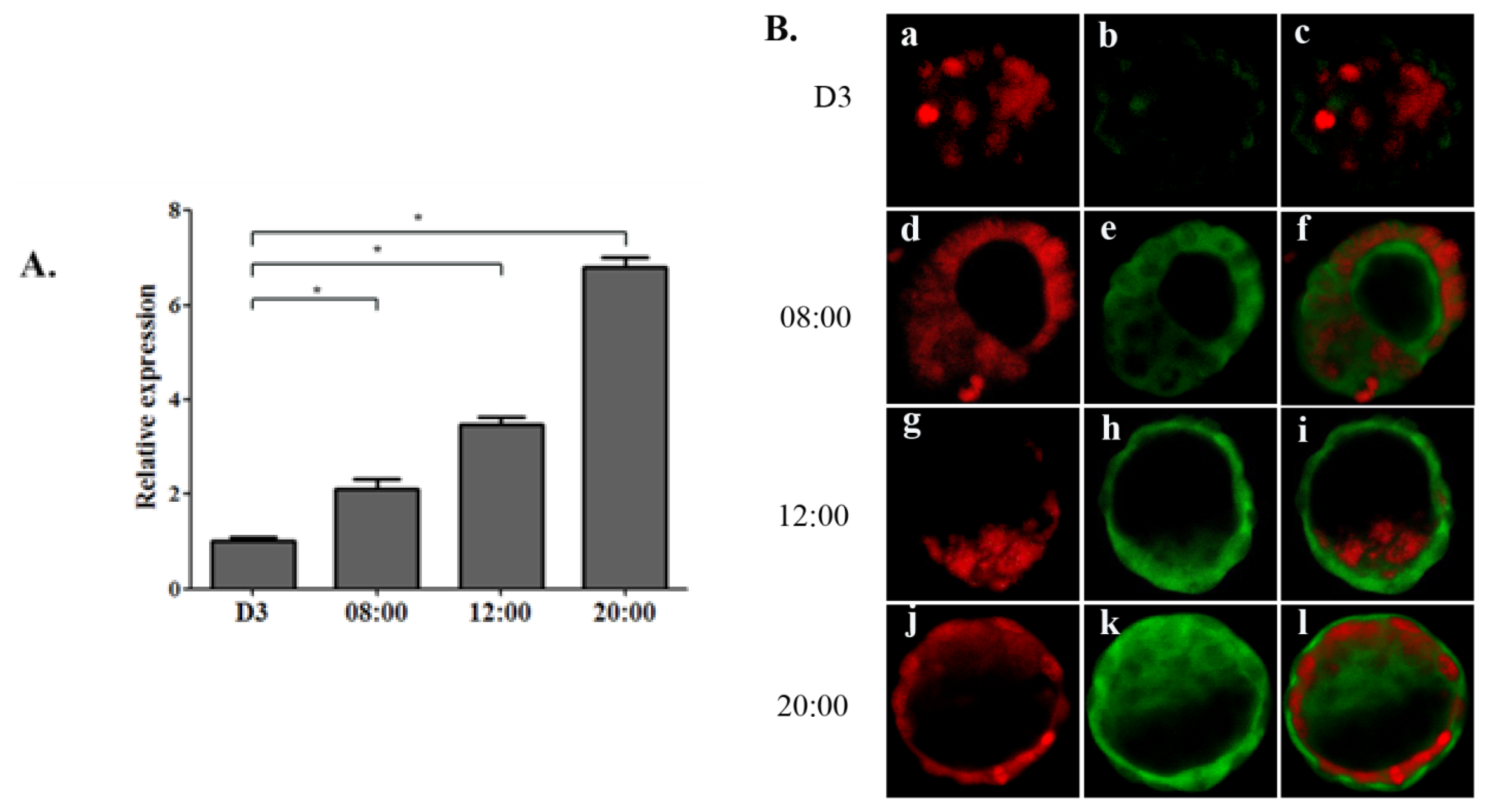
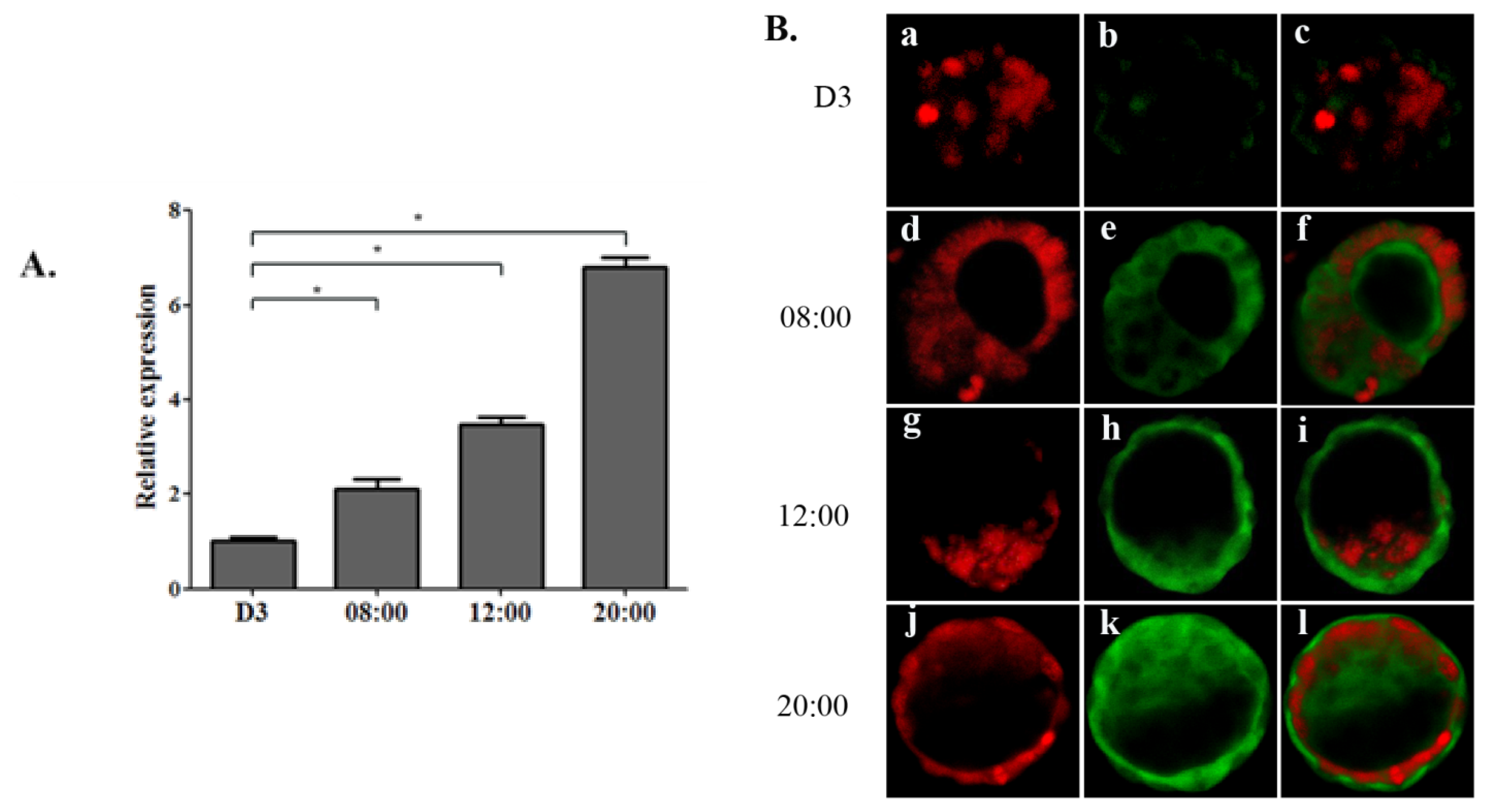
2.1.1. Increase of OPN Expression Corresponds to Ovarian Estrogen Surge
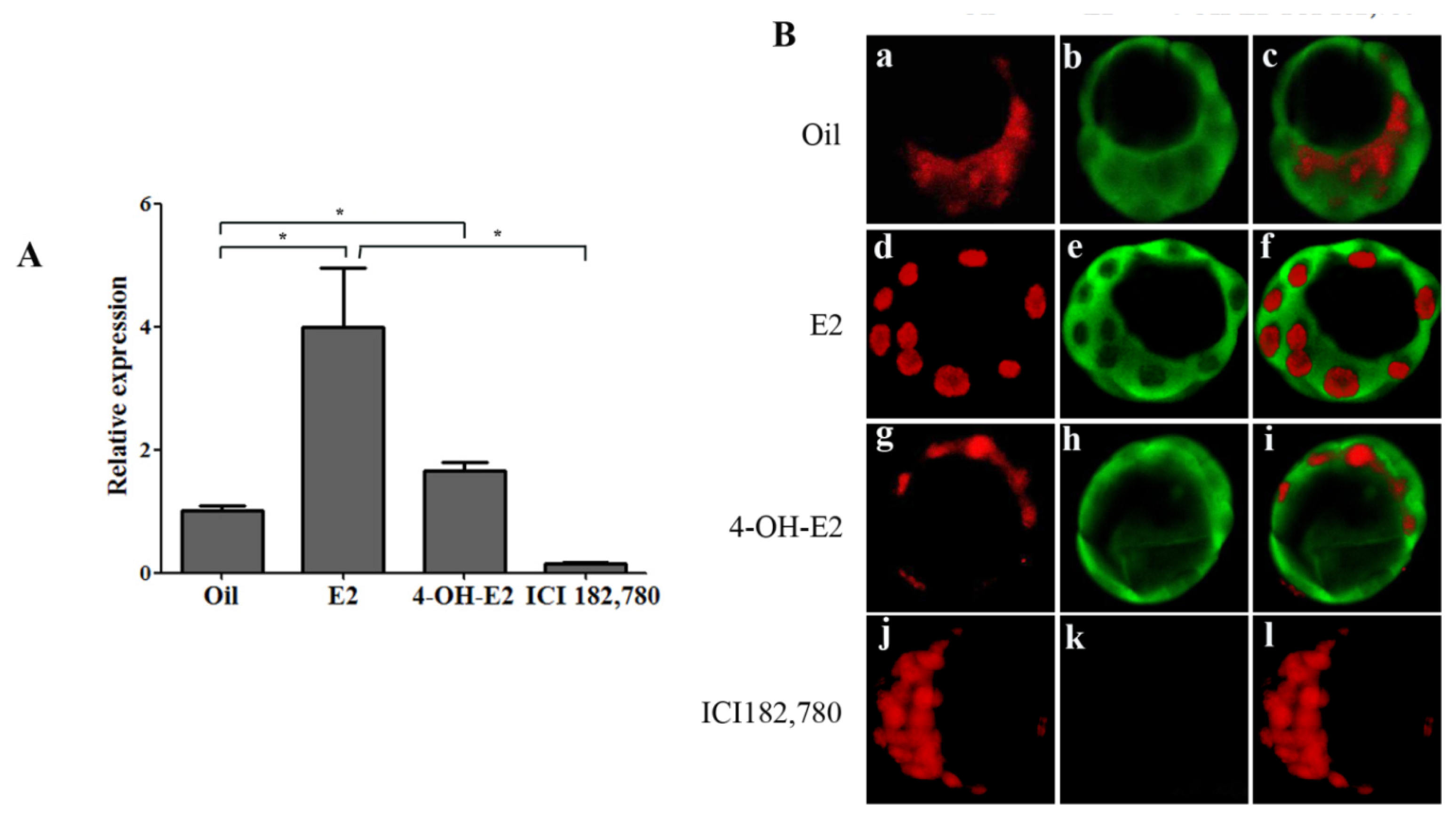
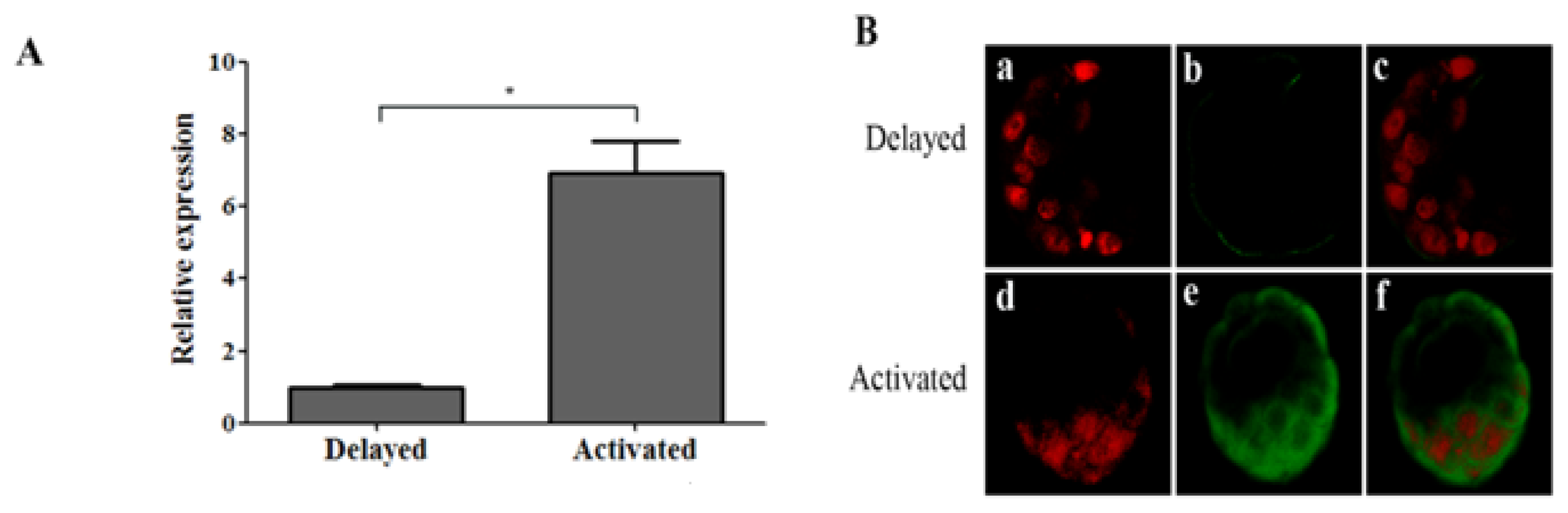
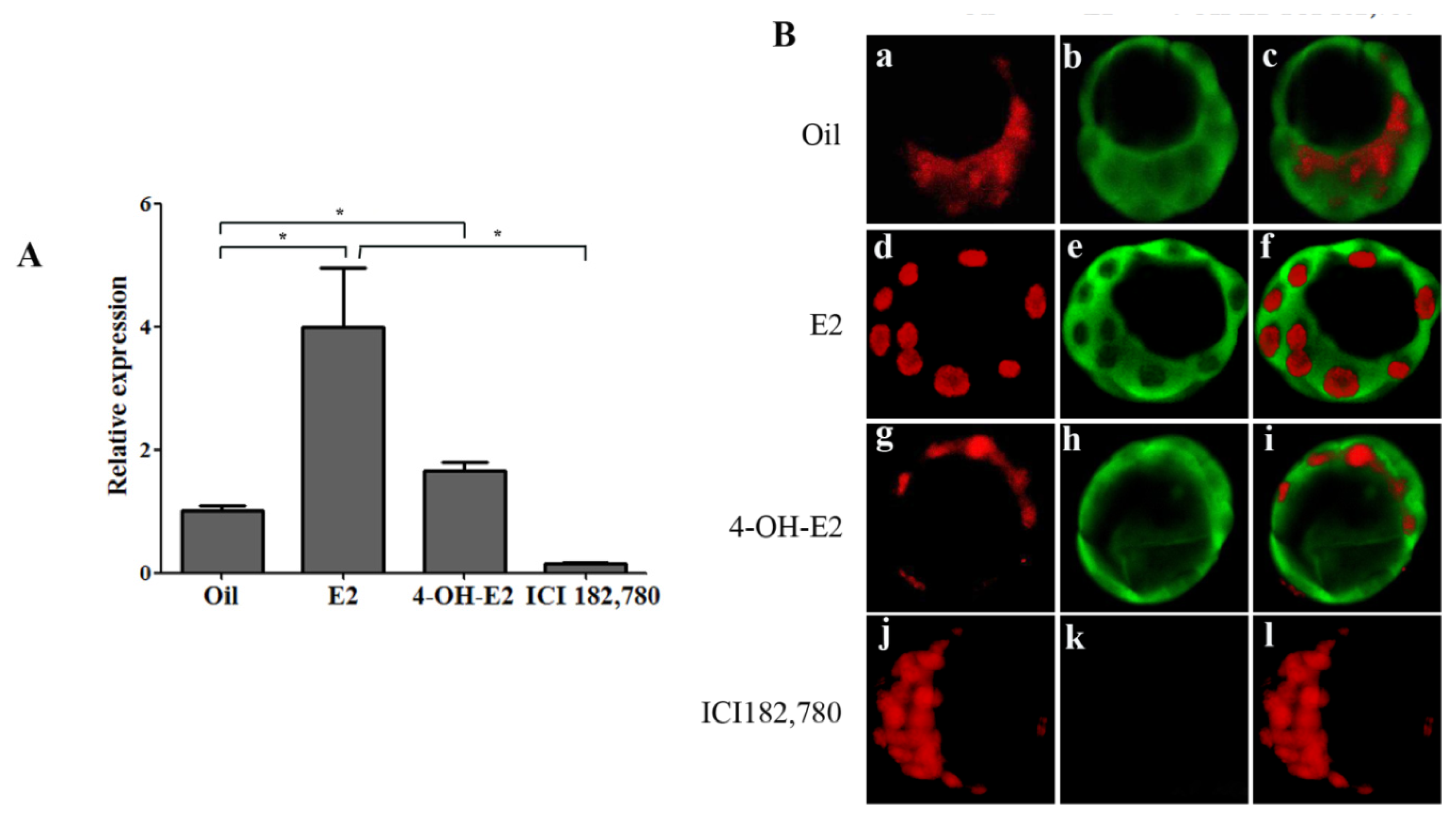
2.1.2. Ovarian Estrogen Is Necessary for Up-regulating OPN Expression in Mouse Blastocyst
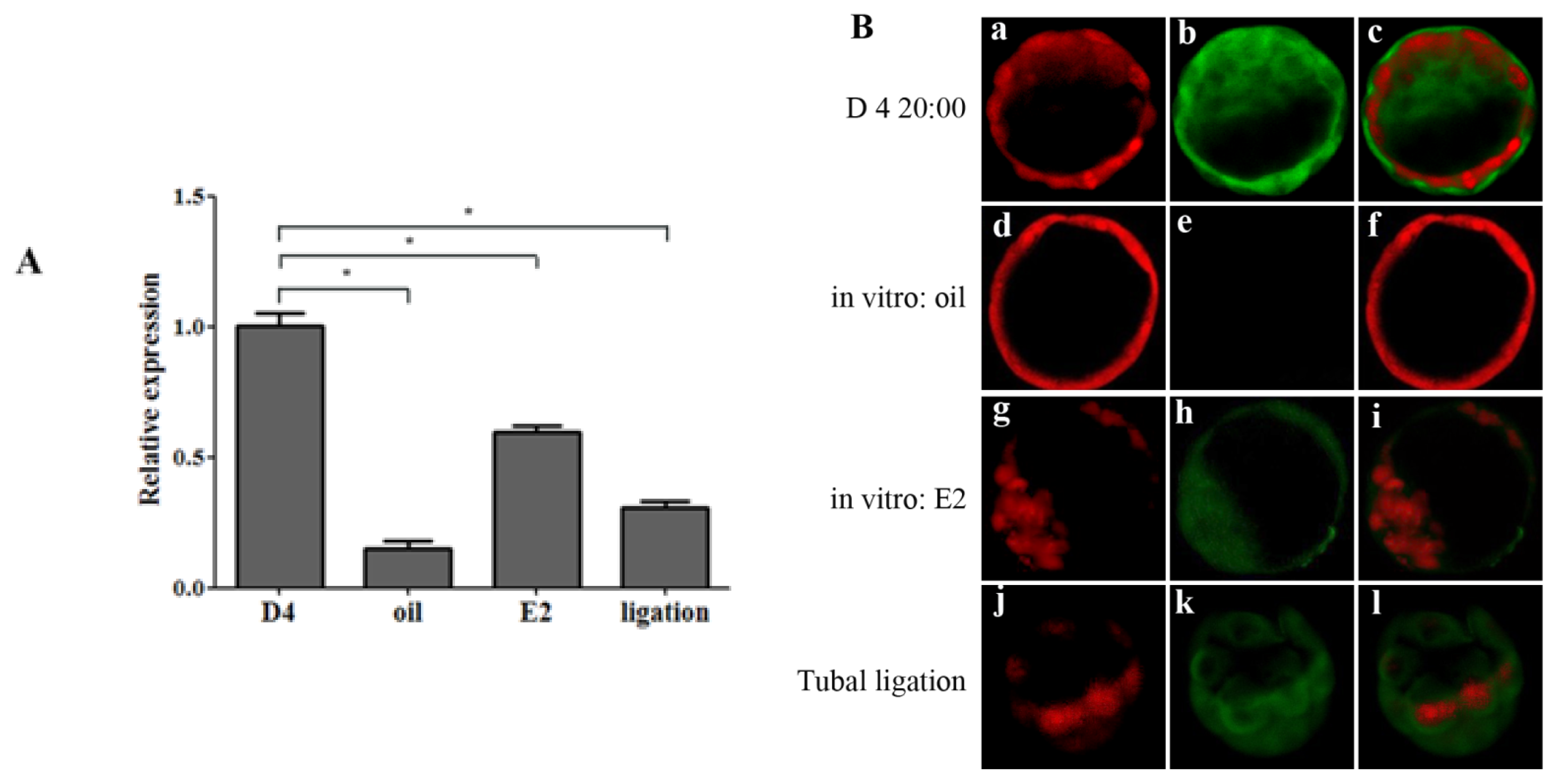
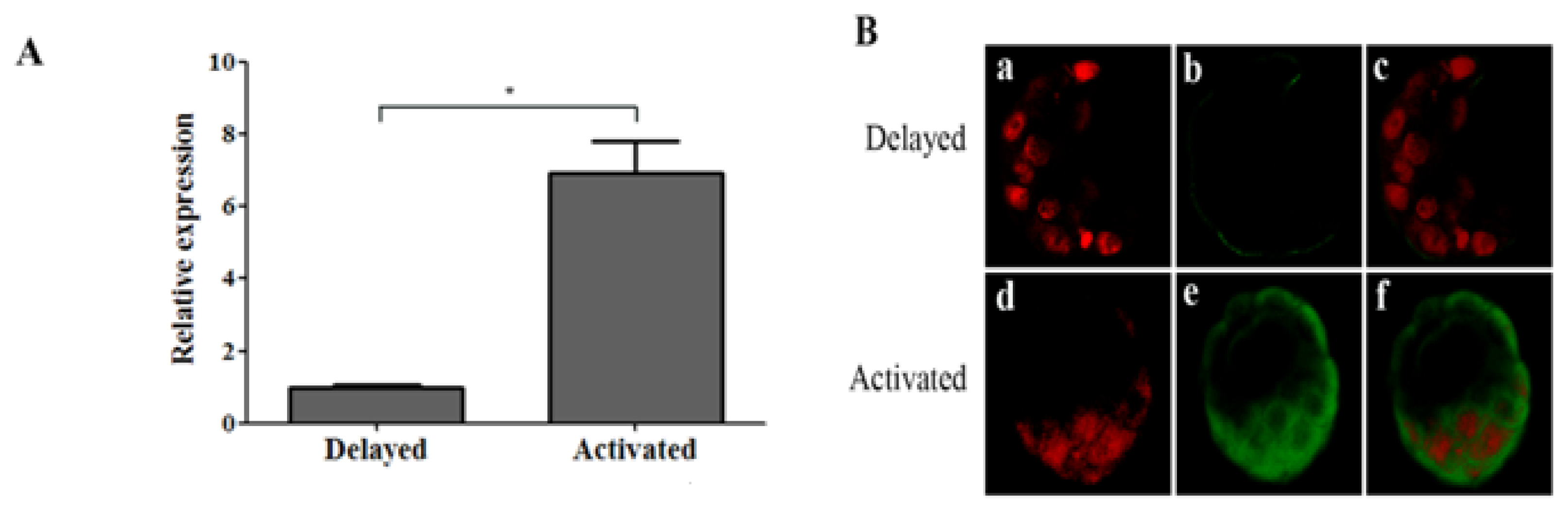
2.1.3. Ovarian Estrogen Induced OPN Expression Is Uterine Micro-Environment Dependent
2.2. Discussion
3. Experimental Section
3.1. Animal and Treatments
3.2. RNA Extraction and Real-Time PCR
3.3. Indirect Immunofluorescence
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Revel, A. Defective endometrial receptivity. Fertil. Steril 2012, 97, 1028–1132. [Google Scholar]
- Hewitt, S.C.; Korach, K.S. Cell biology. A hand to support the implantation window. Science 2011, 331, 863–864. [Google Scholar]
- Daya, S. Luteal support: Progestogens for pregnancy protection. Maturitas 2009, 65, 29–34. [Google Scholar]
- Wang, H.; Dey, S.K. Roadmap to embryo implantation: Clues from mouse models. Nat. Rev. Genet 2006, 7, 185–199. [Google Scholar]
- Zhang, S.; Lin, H.; Kong, S.; Wang, S.; Wang, H.; Wang, H.; Armant, D.R. Physiological and molecular determinants of embryo implantation. Mol. Asp. Med. 2013. [Google Scholar] [CrossRef]
- Bazer, F.W.; Spencer, T.E.; Johnson, G.A.; Burghardt, R.C. Uterine receptivity to implantation of blastocysts in mammals. Front. Biosci 2011, 3, 745–767. [Google Scholar]
- Basak, S.; Dubanchet, S.; Zourbas, S.; Chaouat, G.; Das, C. Expression of pro-inflammatory cytokines in mouse blastocysts during implantation: Modulation by steroid hormones. Am. J. Reprod. Immunol 2002, 47, 2–11. [Google Scholar]
- Das, A.; Mantena, S.R.; Kannan, A.; Evans, D.B.; Bagchi, M.K.; Bagchi, I.C. De novo synthesis of estrogen in pregnant uterus is critical for stromal decidualization and angiogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 12542–12547. [Google Scholar]
- O’Brien, J.E.; Peterson, T.J.; Tong, M.H.; Lee, E.J.; Pfaff, L.E.; Hewitt, S.C.; Korach, K.S.; Weiss, J.; Jameson, J.L. Estrogen-induced proliferation of uterine epithelial cells is independent of estrogen receptor alpha binding to classical estrogen response elements. J. Biol. Chem 2006, 281, 26683–26692. [Google Scholar]
- Hou, Q.; Paria, B.C.; Mui, C.; Dey, S.K.; Gorski, J. Immunolocalization of estrogen receptor protein in the mouse blastocyst during normal and delayed implantation. Proc. Natl. Acad. Sci. USA 1996, 93, 2376–2381. [Google Scholar]
- Ramathal, C.; Wang, W.; Hunt, E.; Bagchi, I.C.; Bagchi, M.K. Transcription factor CCAAT enhancer- binding protein beta (C/EBPbeta) regulates the formation of a unique extracellular matrix that controls uterine stromal differentiation and embryo implantation. J. Biol. Chem 2011, 286, 19860–19871. [Google Scholar]
- Klaffky, E.J.; Gonzales, I.M.; Sutherland, A.E. Trophoblast cells exhibit differential responses to laminin isoforms. Dev. Biol 2006, 292, 277–289. [Google Scholar]
- Zhu, F.; Shen, F.; Fan, Y.; Xie, Y.; Xia, Y.; Kong, Y. Osteopontin increases the expression of beta1, 4-galactosyltransferase-I and promotes adhesion in human RL95-2 cells. Glycoconj. J 2012, 29, 347–356. [Google Scholar]
- Franchi, A.; Zaret, J.; Zhang, X.; Bocca, S.; Oehninger, S. Expression of immunomodulatory genes, their protein products and specific ligands/receptors during the window of implantation in the human endometrium. Mol. Hum. Reprod 2008, 14, 413–421. [Google Scholar]
- Omigbodun, A.; Ziolkiewicz, P.; Tessler, C.; Hoyer, J.R.; Coutifaris, C. Progesterone regulates osteopontin expression in human trophoblasts: A model of paracrine control in the placenta? Endocrinology 1997, 138, 4308–4315. [Google Scholar]
- Casals, G.; Ordi, J.; Creus, M.; Fábregues, F.; Carmona, F.; Casamitjana, R.; Balasch, J. Osteopontin and alphavbeta3 integrin as markers of endometrial receptivity: The effect of different hormone therapies. Reprod. Biomed. Online 2010, 21, 349–359. [Google Scholar]
- Johnson, G.A.; Bazer, F.W.; Jaeger, L.A.; Ka, H.; Garlow, J.E.; Pfarrer, C.; Spencer, T.E.; Burghardt, R.C. Muc-1, integrin, and osteopontin expression during the implantation cascade in sheep. Biol. Reprod 2001, 65, 820–828. [Google Scholar]
- Garlow, J.E.; Ka, H.; Johnson, G.A.; Burghardt, R.C.; Jaeger, L.A.; Bazer, F.W. Analysis of osteopontin at the maternal-placental interface in pigs. Biol. Reprod 2002, 66, 718–725. [Google Scholar]
- Chaen, T.; Konno, T.; Egashira, M.; Bai, R.; Nomura, N.; Nomura, S.; Hirota, Y.; Sakurai, T.; Imakawa, K. Estrogen-dependent uterine secretion of osteopontin activates blastocyst adhesion competence. PLoS One 2012, e48933. [Google Scholar]
- White, F.J.; Burghardt, R.C.; Hu, J.; Joyce, M.M.; Spencer, T.E.; Johnson, G.A. Secreted phosphoprotein 1 (osteopontin) is expressed by stromal macrophages in cyclic and pregnant endometrium of mice, but is induced by estrogen in luminal epithelium during conceptus attachment for implantation. Reproduction 2006, 132, 919–929. [Google Scholar]
- Weintraub, A.S.; Lin, X.; Itskovich, V.V.; Aguinaldo, J.G.; Chaplin, W.F.; Denhardt, D.T.; Fayad, Z.A. Prenatal detection of embryo resorption in osteopontin-deficient mice using serial noninvasive magnetic resonance microscopy. Pediatr. Res 2004, 55, 419–424. [Google Scholar]
- Botquin, V.; Hess, H.; Fuhrmann, G.; Anastassiadis, C.; Gross, M.K.; Vriend, G.; Schöler, H.R. New POU dimer configuration mediates antagonistic control of an osteopontin preimplantation enhancer by Oct-4 and Sox-2. Genes Dev 1998, 12, 2073–2090. [Google Scholar]
- Armant, D.R. Life and death responses to trophinin-mediated adhesion during blastocyst implantation. Cell Cycle 2011, 10, 574–575. [Google Scholar]
- Fukuda, M.N.; Sugihara, K. An integrated view of L-selectin and trophinin function in human embryo implantation. J. Obstet. Gynaecol. Res 2008, 34, 129–136. [Google Scholar]
- Daikoku, T.; Cha, J.; Sun, X.; Tranguch, S.; Xie, H.; Fujita, T.; Hirota, Y.; Lydon, J.; DeMayo, F.; Maxson, R.; et al. Conditional deletion of Msx homeobox genes in the uterus inhibits blastocyst implantation by altering uterine receptivity. Dev. Cell 2011, 21, 1014–1025. [Google Scholar]
- Platzer, G.; Schedlbauer, A.; Chemelli, A.; Ozdowy, P.; Coudevylle, N.; Auer, R.; Kontaxis, G.; Hartl, M.; Miles, A.J.; Wallace, B.A.; et al. The metastasis-associated extracellular matrix protein osteopontin forms transient structure in ligand interaction sites. Biochemistry 2011, 50, 6113–6124. [Google Scholar]
- Singh, H.; Aplin, J.D. Adhesion molecules in endometrial epithelium: Tissue integrity and embryo implantation. J. Anat 2009, 215, 3–13. [Google Scholar]
- Zou, C.; Luo, Q.; Qin, J.; Shi, Y.; Yang, L.; Ju, B.; Song, G. Osteopontin Promotes Mesenchymal Stem Cell Migration and Lessens Cell Stiffness via Integrin beta1, FAK, and ERK Pathways. Cell Biochem. Biophys 2013, 65, 455–462. [Google Scholar]
- Casals, G.; Ordi, J.; Creus, M.; Fábregues, F.; Carmona, F.; Casamitjana, R.; Balasch, J. Expression pattern of osteopontin and alphavbeta3 integrin during the implantation window in infertile patients with early stages of endometriosis. Hum. Reprod 2012, 27, 805–813. [Google Scholar]
- Johnson, G.A.; Burghardt, R.C.; Bazer, F.W.; Spencer, T.E. Osteopontin: Roles in implantation and placentation. Biol. Reprod 2003, 69, 1458–1471. [Google Scholar]
- Carson, D.D.; Lagow, E.; Thathiah, A.; Al-Shami, R.; Farach-Carson, M.C.; Vernon, M.; Yuan, L.; Fritz, M.A.; Lessey, B. Changes in gene expression during the early to mid-luteal (receptive phase) transition in human endometrium detected by high-density microarray screening. Mol. Hum. Reprod 2002, 8, 871–879. [Google Scholar]
- Das, A.; Li, Q.; Laws, M.J.; Kaya, H.; Bagchi, M.K.; Bagchi, I.C. Estrogen-induced expression of Fos-related antigen 1 (FRA-1) regulates uterine stromal differentiation and remodeling. J. Biol. Chem 2012, 287, 19622–19630. [Google Scholar]
- Wetendorf, M.; DeMayo, F.J. The progesterone receptor regulates implantation, decidualization, and glandular development via a complex paracrine signaling network. Mol. Cell Endocrinol 2012, 357, 108–118. [Google Scholar]
- Ziecik, A.J.; Waclawik, A.; Kaczmarek, M.M.; Blitek, A.; Jalali, B.M.; Andronowska, A. Mechanisms for the establishment of pregnancy in the pig. Reprod. Domest. Anim 2011, 46, 31–41. [Google Scholar]
- Spencer, T.E.; Johnson, G.A.; Bazer, F.W.; Burghardt, R.C.; Palmarini, M. Pregnancy recognition and conceptus implantation in domestic ruminants: Roles of progesterone, interferons and endogenous retroviruses. Reprod. Fertil. Dev 2007, 19, 65–78. [Google Scholar]
- Zhang, S.; Kong, S.; Lu, J.; Wang, Q.; Chen, Y.; Wang, W.; Wang, B.; Wang, H. Deciphering the molecular basis of uterine receptivity. Mol. Reprod. Dev 2013, 80, 8–21. [Google Scholar]
- Banerjee, A.; Padh, H.; Nivsarkar, M. Hormonal crosstalk with calcium channel blocker during implantation. Syst. Biol. Reprod. Med 2011, 57, 186–189. [Google Scholar]
- Cha, J.; Sun, X.; Dey, S.K. Mechanisms of implantation: Strategies for successful pregnancy. Nat. Med 2012, 18, 1754–1767. [Google Scholar]
- Das, S.K.; Das, N.; Wang, J.; Lim, H.; Schryver, B.; Plowman, G.D.; Dey, S.K. Expression of betacellulin and epiregulin genes in the mouse uterus temporally by the blastocyst solely at the site of its apposition is coincident with the “window” of implantation. Dev. Biol 1997, 190, 178–190. [Google Scholar]
- Petracco, R.G.; Kong, A.; Grechukhina, O.; Krikun, G.; Taylor, H.S. Global gene expression profiling of proliferative phase endometrium reveals distinct functional subdivisions. Reprod. Sci 2012, 19, 1138–1145. [Google Scholar]
- Shuya, L.L.; Menkhorst, E.M.; Yap, J.; Li, P.; Lane, N.; Dimitriadis, E. Leukemia inhibitory factor enhances endometrial stromal cell decidualization in humans and mice. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Teng, C.B.; Diao, H.L.; Ma, X.H.; Xu, L.B.; Yang, Z.M. Differential expression and activation of Stat3 during mouse embryo implantation and decidualization. Mol. Reprod. Dev 2004, 69, 1–10. [Google Scholar]
- Basak, S.; Dhar, R.; Das, C. Steroids modulate the expression of alpha4 integrin in mouse blastocysts and uterus during implantation. Biol. Reprod 2002, 66, 1784–1789. [Google Scholar]
- Bazer, F.W.; Wu, G.; Johnson, G.A.; Kim, J.; Song, G. Uterine histotroph and conceptus development: Select nutrients and secreted phosphoprotein 1 affect mechanistic target of rapamycin cell signaling in ewes. Biol. Reprod 2011, 85, 1094–1107. [Google Scholar]
- Hannan, N.J.; Paiva, P.; Meehan, K.L.; Rombauts, L.J.; Gardner, D.K.; Salamonsen, L.A. Analysis of fertility-related soluble mediators in human uterine fluid identifies VEGF as a key regulator of embryo implantation. Endocrinology 2011, 152, 4948–4956. [Google Scholar]
- Xiao, Y.; Sun, X.; Yang, X.; Zhang, J.; Xue, Q.; Cai, B.; Zhou, Y. Leukemia inhibitory factor is dysregulated in the endometrium and uterine flushing fluid of patients with adenomyosis during implantation window. Fertil. Steril 2010, 94, 85–89. [Google Scholar]
- Bang, C.; Thum, T. Exosomes: New players in cell-cell communication. Int. J. Biochem. Cell Biol 2012, 44, 2060–2064. [Google Scholar]
- Ng, Y.H.; Rome, S.; Jalabert, A.; Forterre, A.; Singh, H.; Hincks, C.L.; Salamonsen, L.A. Endometrial exosomes/microvesicles in the uterine microenvironment: A new paradigm for embryo-endometrial cross talk at implantation. PLoS One 2013, 8, e58502. [Google Scholar]
- Johnson, G.A.; Burghardt, R.C.; Joyce, M.M.; Spencer, T.E.; Bazer, F.W.; Gray, C.A.; Pfarrer, C. Osteopontin is synthesized by uterine glands and a 45-kDa cleavage fragment is localized at the uterine-placental interface throughout ovine pregnancy. Biol. Reprod 2003, 69, 92–98. [Google Scholar]

© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xie, Q.-Z.; Qi, Q.-R.; Chen, Y.-X.; Xu, W.-M.; Liu, Q.; Yang, J. Uterine Micro-Environment and Estrogen-Dependent Regulation of Osteopontin Expression in Mouse Blastocyst. Int. J. Mol. Sci. 2013, 14, 14504-14517. https://doi.org/10.3390/ijms140714504
Xie Q-Z, Qi Q-R, Chen Y-X, Xu W-M, Liu Q, Yang J. Uterine Micro-Environment and Estrogen-Dependent Regulation of Osteopontin Expression in Mouse Blastocyst. International Journal of Molecular Sciences. 2013; 14(7):14504-14517. https://doi.org/10.3390/ijms140714504
Chicago/Turabian StyleXie, Qing-Zhen, Qian-Rong Qi, Ying-Xian Chen, Wang-Ming Xu, Qian Liu, and Jing Yang. 2013. "Uterine Micro-Environment and Estrogen-Dependent Regulation of Osteopontin Expression in Mouse Blastocyst" International Journal of Molecular Sciences 14, no. 7: 14504-14517. https://doi.org/10.3390/ijms140714504